
Biotechnological Trends in Spider and Scorpion Antivenom Development

 ,
,
Abstract
:
1. Introduction
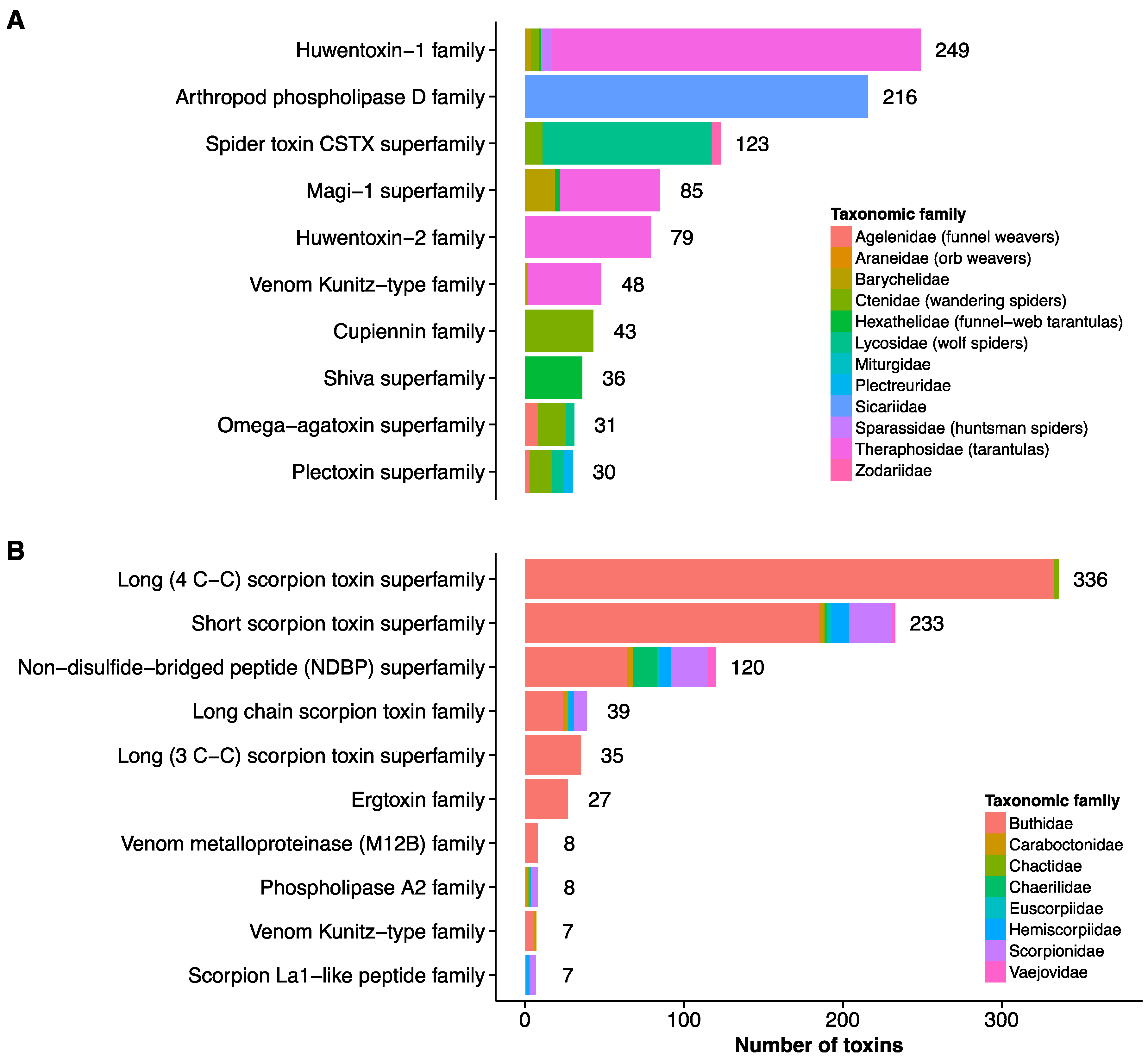
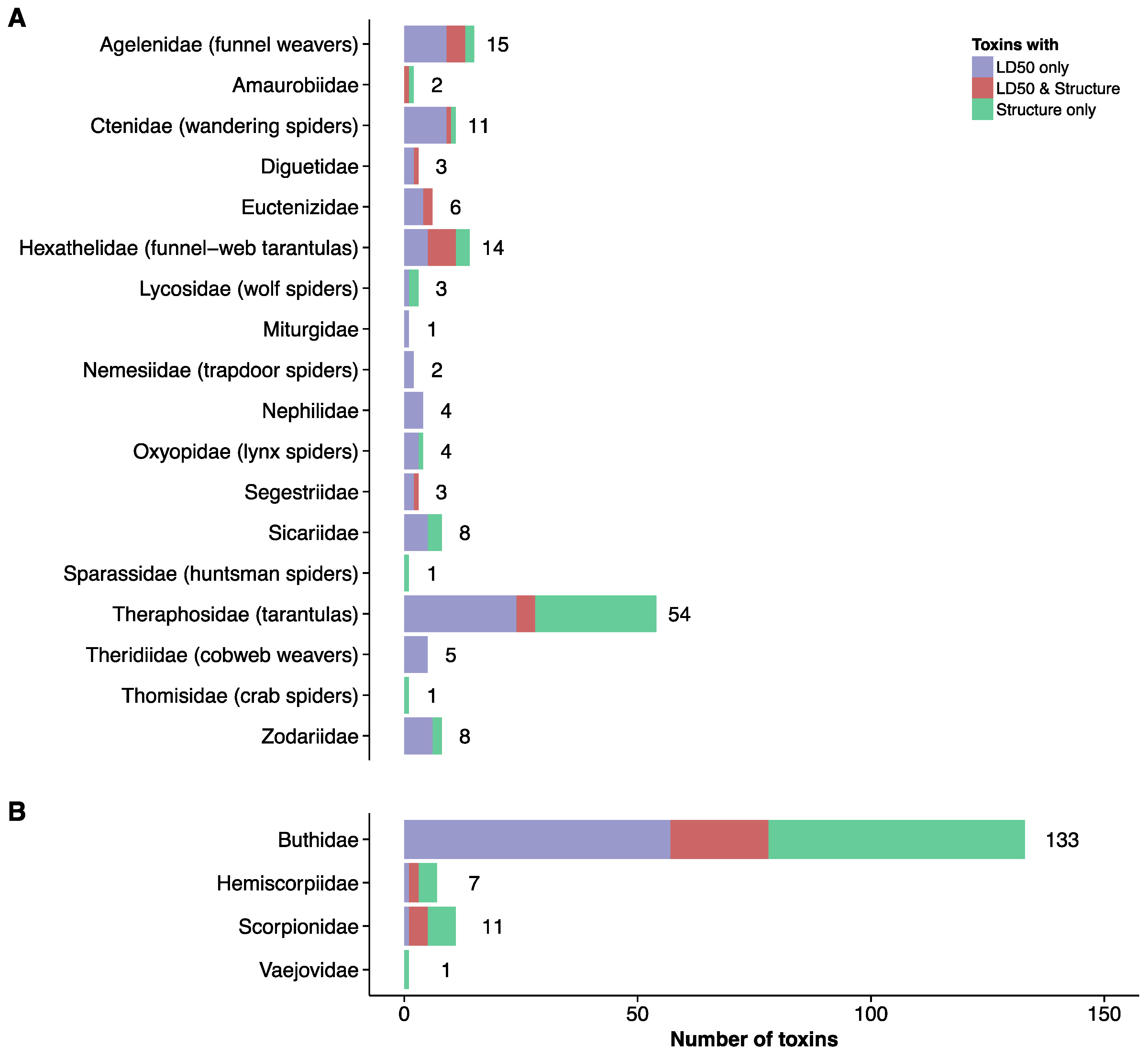
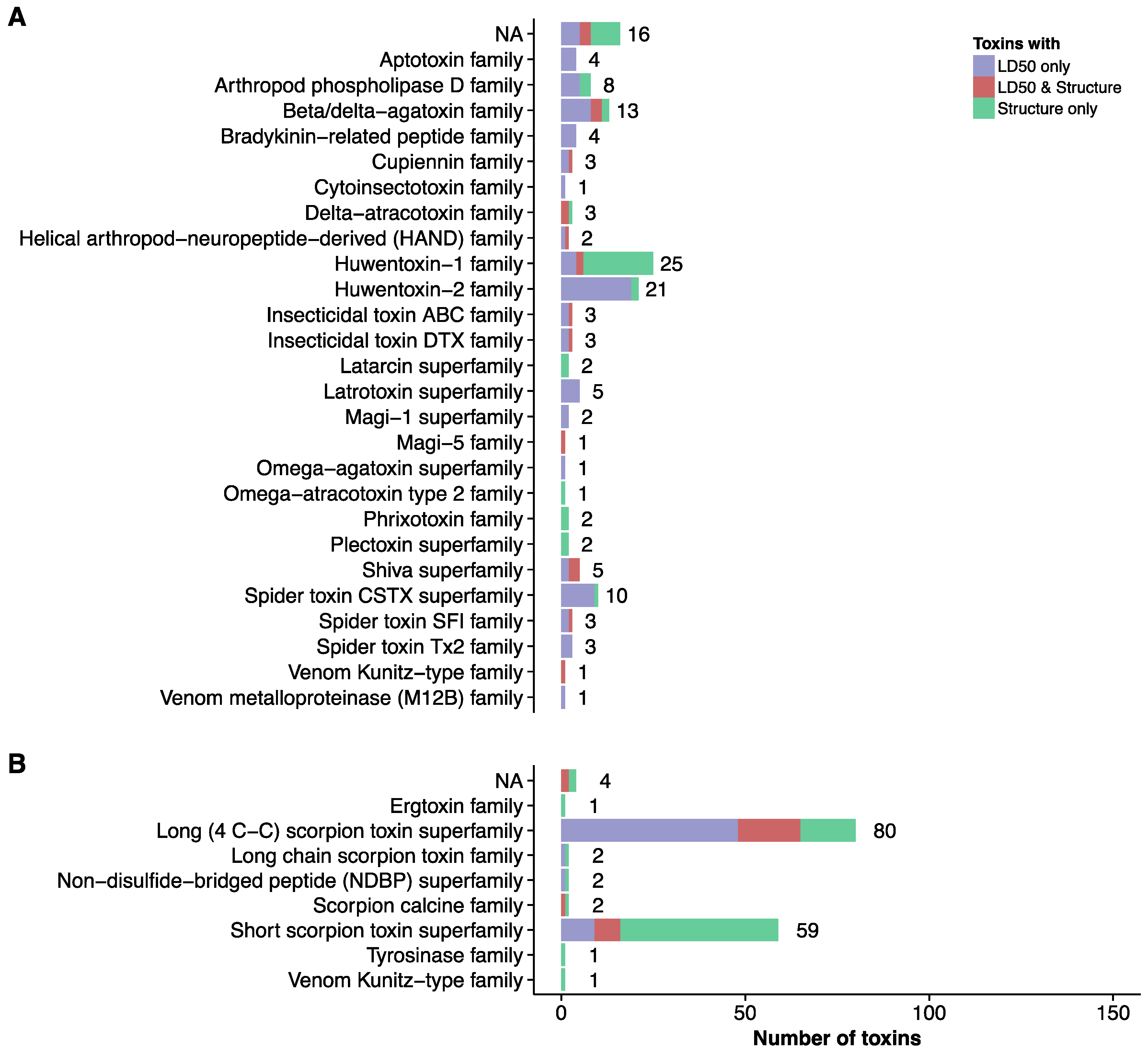
2. Current Knowledge of Toxins and Venom Proteomes
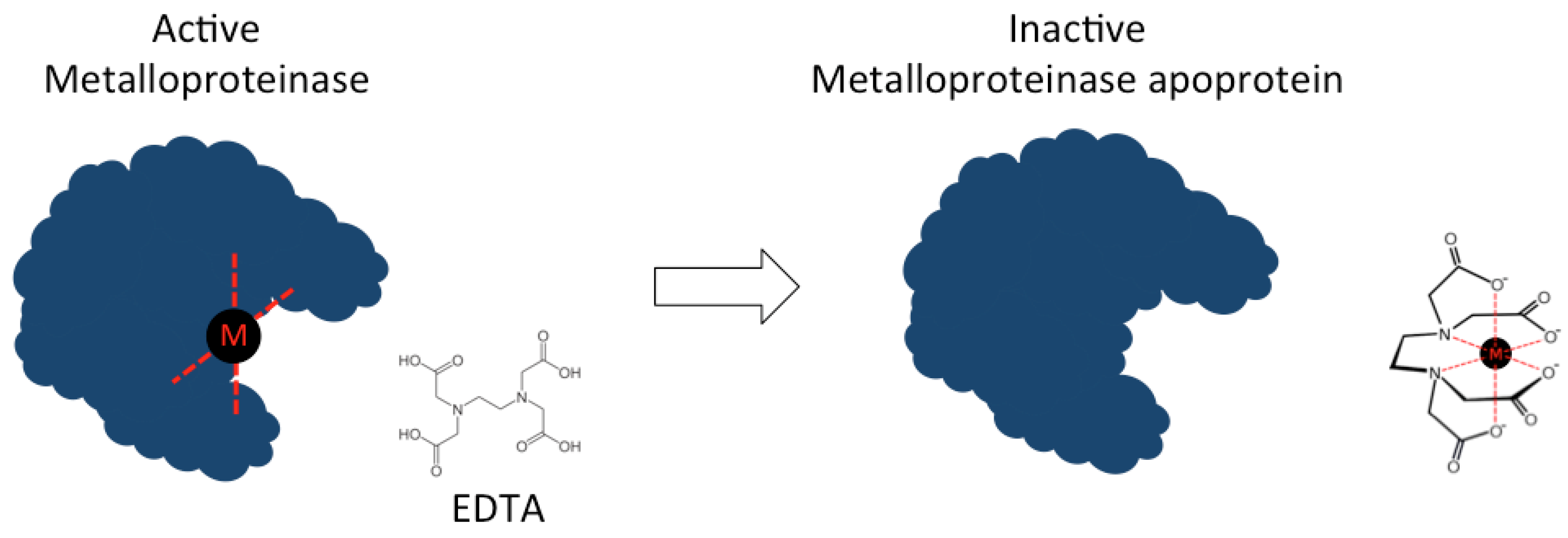
3. Toxin Inhibition by Small Molecules
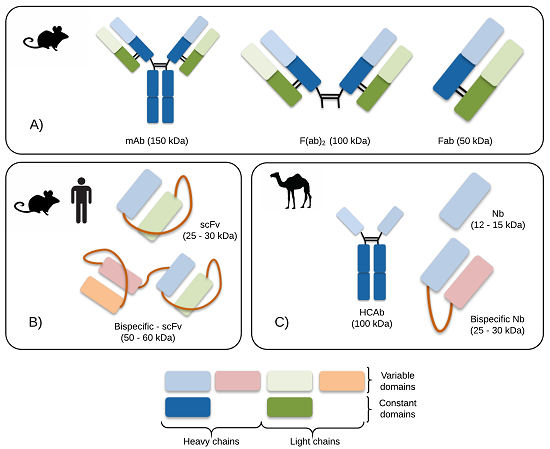
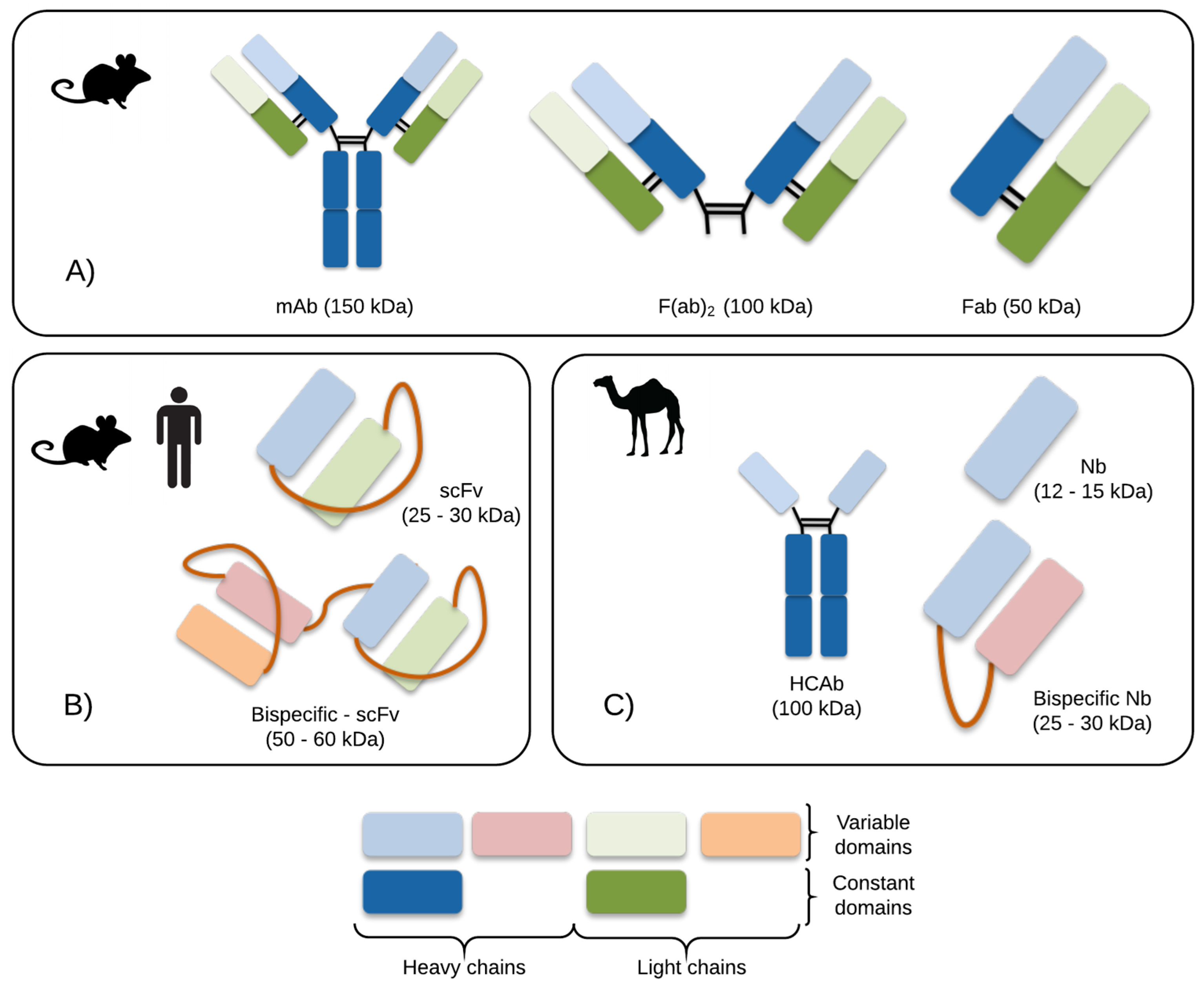
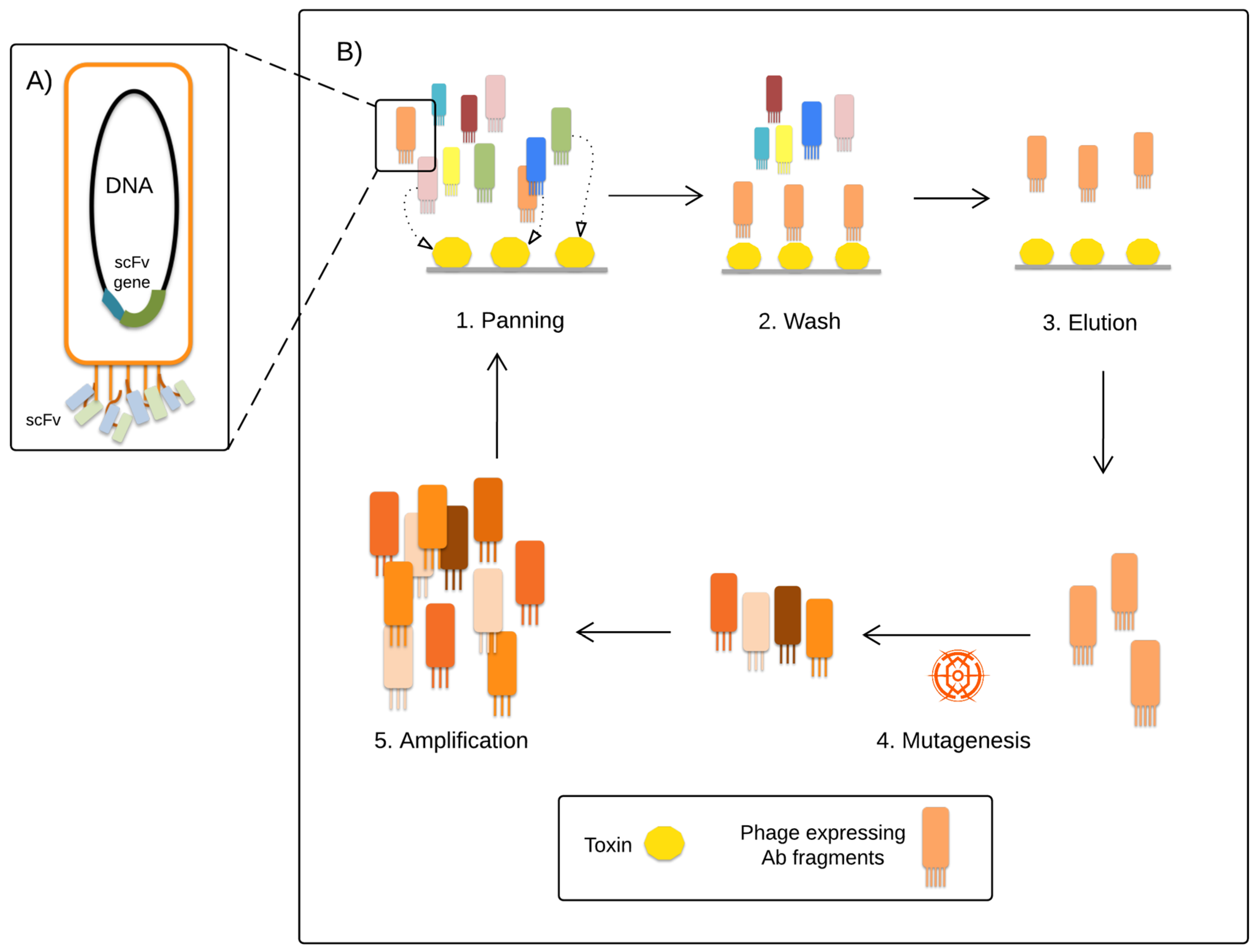
4. Research Efforts within Antibodies and Antibody Fragments
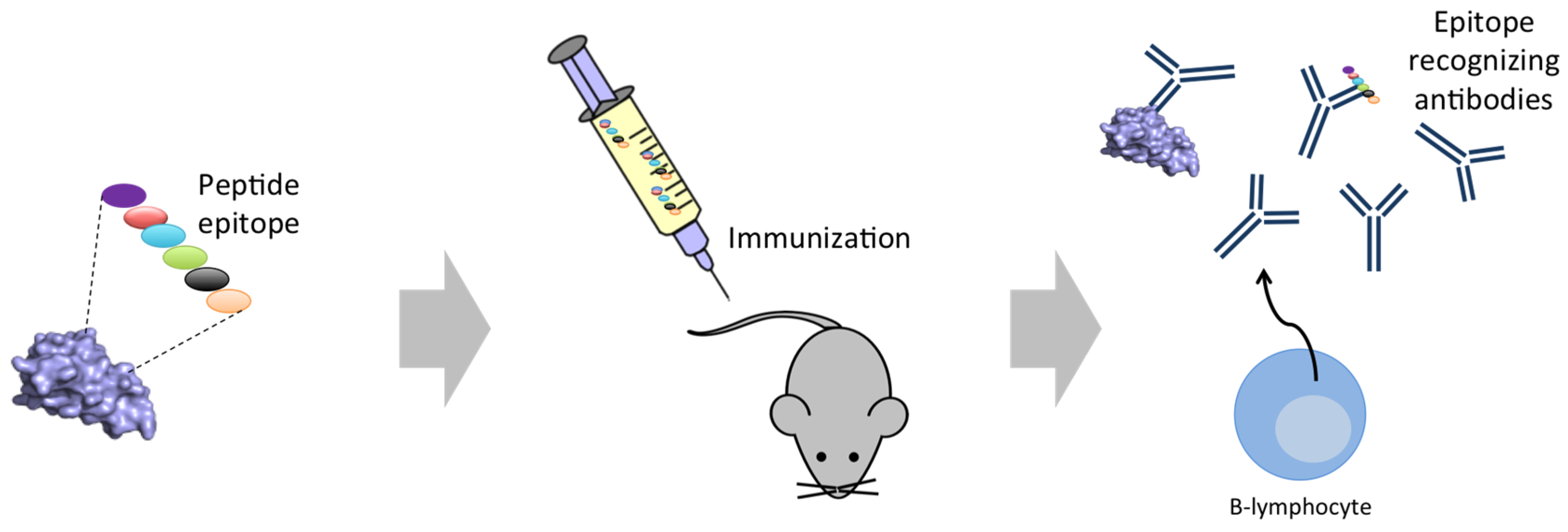
5. Next Generation Immunization Strategies
6. Future Perspectives
Acknowledgments
Conflicts of Interest
References
- Warrell, D.; Gutiérrez, J.M.; Padilla, A. Rabies and Envenomings: A Neglected Public Health Issue: Report of a Consultative Meeting; World Health Organization: Geneva, Spain, 2007. [Google Scholar]
- Carmo, A.O.; Chatzaki, M.; Horta, C.C.R.; Magalhães, B.F.; Oliveira-Mendes, B.B.R.; Chávez-Olórtegui, C.; Kalapothakis, E. Evolution of alternative methodologies of scorpion antivenoms production. Toxicon 2015, 97, 64–74. [Google Scholar] [CrossRef] [PubMed]
- Laraba-Djebari, F.; Adi-Bessalem, S.; Hammoudi-Triki, D. Scorpion Venoms: Pathogenesis and Biotherapies. In Toxinology - Scorpion Venoms; Springer Netherlands: Berlin, Germany, 2015; Volume 4, pp. 63–85. [Google Scholar]
- Diaz, J.H. The Global Epidemiology, Syndromic Classification, Management, and Prevention of Spider Bites. Am. J. Trop. Med. Hyg. 2004, 71, 239–250. [Google Scholar] [PubMed]
- Bettini, S. Epidemiology of latrodectism. Toxicon 1964, 2, 93–102. [Google Scholar] [CrossRef]
- Goyffon, M.; Tournier, J.-N. Scorpions: A Presentation. Toxins 2014, 6, 2137–2148. [Google Scholar] [CrossRef] [PubMed]
- Medical Entomology; Eldridge, B.F.; Edman, J.D. (Eds.) Springer Netherlands: Berlin, Germany, 2004.
- Chippaux, J.-P. Emerging options for the management of scorpion stings. Drug Des. Devel. Ther. 2012, 6, 165–173. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez, E.R.R.; Umbarila, L.R.; Possani, L.D.; Becerril, B. Recombinant Neutralizing Antibodies, A New Generation of Antivenoms. In Toxinology - Scorpion Venoms; Springer: Berlin, Germany, 2015; Volume 4, pp. 139–159. [Google Scholar]
- Pucca, M.B.; Carlos, J.; Roncolato, E.C.; Bertolini, T.B.; Fossa, C.M.; Varanda, W.A.; Arantes, E.C.; Barbosa, J.E. Monoclonal antibody (Scfv) against the venom of the scorpion Tityus serrulatus, produced by phage display technic, is capable to recognize and inhibit the action of the ts1 toxin. Epeq/Fafibe 2011, 1, 18–23. [Google Scholar]
- Rodríguez-Rodríguez, E.R.; Olamendi-Portugal, T.; Serrano-Posada, H.; Arredondo-López, J.N.; Gómez-Ramírez, I.; Fernández-Taboada, G.; Possani, L.D.; Anguiano-Vega, G.A.; Riaño-Umbarila, L.; Becerril, B. Broadening the neutralizing capacity of a family of antibody fragments against different toxins from Mexican scorpions. Toxicon 2016, 119, 52–63. [Google Scholar] [CrossRef] [PubMed]
- Meadows, P.E.; Russell, F.E. Milking of arthropods. Toxicon 1970, 8, 311–312. [Google Scholar] [CrossRef]
- Product Information. Funnel Web Spider Antivenom. Available online: http://www.csl.com.au/docs/822/652/Funnel%20Web%20Spider%20AV%20PI%20V6%20clean%20TGA%20approved%2018%20Sept%202013.pdf (accessed on 15 July 2016).
- Product Information. Red Back Spider Antivenom. Available online: http://www.csl.com.au/docs/1002/757/Red-Back-Spider-AV_PI_V5_Clean_TGA-Approved_8-January-2014.pdf (accessed on 15 July 2016).
- Our Products—Aracmyn PLUS. Available online: http://www.bioclon.com.mx/bioclon/html/aracmynP_en.html (accessed on 15 July 2016).
- Our Products—Reclusmyn. Available online: http://www.bioclon.com.mx/bioclon/html/reclusmyn_en.html (accessed on 15 July 2016).
- Package Leaflet: Soro Antiarachnidico—Butantan. Available online: http://www.toxinfo.org/antivenoms/resources/antivenom_brasil-butantan-antiarachnidico_2011-07-28.pdf (accessed on 15 July 2016).
- WCH Clinical Toxinology Resources—Latrodectus mactans. Available online: http://www.toxinology.com/fusebox.cfm?fuseaction=main.spiders.display&id=SP00061 (accessed on 15 July 2016).
- Detalle del Producto. Suero Antiloxoscélico Monovalente. Available online: http://www.ins.gob.pe/insvirtual/hdetprod.aspx?1596 (accessed on 15 July 2016).
- Instituto Vital Brazil—Soro Antilatrodéctico. Available online: http://www.ivb.rj.gov.br/soros_produzidos.html (accessed on 15 July 2016).
- Antivenin (Latrodectus mactans). Available online: http://www.merck.com/product/usa/pi_circulars/a/antivenin/antivenin_pi.pdf (accessed on 15 July 2016).
- NHLS SAVP | Products. Available online: http://www.savp.co.za/?page=products&id=11 (accessed on 15 July 2016).
- BIRMEX. Nuestros Productos. Available online: http://www.birmex.gob.mx/comercializacion.html (accessed on 15 July 2016).
- Suero Antiescorpionico—Centro de Biotecnologia de la Universidad Central de Venezuela. Available online: http://www.biotecfar.com.ve/biotecfar/pdf/suero_antiescorpionico.pdf (accessed on 15 July 2016).
- Institut Pasteur D’Algerie. Available online: http://www.pasteur.dz/presentation.php?page=32 (accessed on 15 July 2016).
- WCH Clinical Toxinology Resources—Buthus Occitanus. Available online: http://www.toxinology.com/fusebox.cfm?fuseaction=main.scorpions.display&mode=PrintFriendly&id=SC0038 (accessed on 15 July 2016).
- Scorpion Antivenom Production. Turk Hij Den Biyol Derg. Available online: http://www.turkhijyen.org/eng/jvi.aspx?pdir=turkhijyen&plng=eng&un=THDBD-74046 (accessed on 15 July 2016).
- Instructions for Use. Alacramyn®. Available online: http://www.toxinfo.org/antivenoms/resources/antivenom_mexico-bioclon-alacramyn_2011-07-28.pdf (accessed on 15 July 2016).
- Package Leaflet: Soro Antiescorpionico—Butantan. Available online: http://www.toxinfo.org/antivenoms/resources/antivenom_brasil-butantan-scorpionico_2011-07-05.pdf (accessed on 15 July 2016).
- Polyvalent Scorpion Antivenom | National Antivenom and Vaccine Production Center. Available online: http://www.antivenom-center.com/navpc-products/polyvalent-scorpion-antivenom/ (accessed on 15 July 2016).
- Institut Pasteur de Tunis—Production. Available online: http://www.pasteur.tn/index.php?option=com_content&view=article&id=161&Itemid=199 (accessed on 15 July 2016).
- Razi Vaccine & Serum Research Institute - Medical Antisera. Available online: http://www.rvsri.ac.ir/Portal/Home/Default.aspx?CategoryID=50c3cada-f4f3-424f-955f-e2b04ca488f2 (accessed on 15 July 2016).
- MAVIN Poison Centre Munich—Scorpifav. Available online: http://www.toxinfo.org/antivenoms/productinfo/SCORPIFAV.html (accessed on 15 July 2016).
- MAVIN Poison Centre Munich - Scorpion antivenom Twyford (North Africa). Available online: http://www.toxinfo.org/antivenoms/productinfo/SCORPION_ANTIVENOM_TWYFORD__NORTH_AFRICA_.html (accessed on 15 July 2016).
- Vacsera—Products: Anti-Scorpion Serum. Available online: http://www.tradeegypt.com/vacsera/?ID=19730&CatalogID=3186&link=products&catname=Medical+%26+Pharmaceutical&subid=877&catid=32 (accessed on 15 July 2016).
- Scorpion Anti Serum | VINS BioProducts Limited. Available online: http://vinsbio.in/antiscorpion/ (accessed on 15 July 2016).
- Identificação do Medicamento—Soro Antiescorpiônico. Available online: http://www.anvisa.gov.br/datavisa/fila_bula/frmVisualizarBula.asp?pNuTransacao=1233402015&pIdAnexo=2447866 (accessed on 15 July 2016).
- Haffkine Antitoxins & Sera - antiscorpion venom serum. Available online: http://www.vaccinehaffkine.com/products/antitoxins-sera/antiscorpion-venom-serum-detail.html (accessed on 15 July 2016).
- Sannaningaiah, D.; Subbaiah, G.K.; Kempaiah, K. Pharmacology of spider venom toxins. Toxin Rev. 2014, 33, 206–220. [Google Scholar] [CrossRef]
- Palagi, A.; Koh, J.M.S.; Leblanc, M.; Wilson, D.; Dutertre, S.; King, G.F.; Nicholson, G.M.; Escoubas, P. Unravelling the complex venom landscapes of lethal Australian funnel-web spiders (Hexathelidae: Atracinae) using LC-MALDI-TOF mass spectrometry. J. Proteomics 2013, 80, 292–310. [Google Scholar] [CrossRef] [PubMed]
- Cao, Z.; Yu, Y.; Wu, Y.; Hao, P.; Di, Z.; He, Y.; Chen, Z.; Yang, W.; Shen, Z.; He, X.; et al. The genome of Mesobuthus martensii reveals a unique adaptation model of arthropods. Nat. Commun. 2013, 4. [Google Scholar] [CrossRef] [PubMed]
- Consortium, T.U. UniProt: A hub for protein information. Nucleic Acids Res. 2015, 43, D204–D212. [Google Scholar] [CrossRef] [PubMed]
- Lewis, R.J.; Garcia, M.L. Therapeutic potential of venom peptides. Nat. Rev. Drug Discov. 2003, 2, 790–802. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Yarov-Yarovoy, V.; Kahn, R.; Gordon, D.; Gurevitz, M.; Scheuer, T.; Catterall, W.A. Mapping the receptor site for α-scorpion toxins on a Na+ channel voltage sensor. Proc. Natl. Acad. Sci. USA 2011, 108, 15426–15431. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez-Rodríguez, E.R.; Olamendi-Portugal, T.; Serrano-Posada, H.; Arredondo-López, J.N.; Gómez-Ramírez, I.; Fernández-Taboada, G.; Possani, L.D.; Anguiano-Vega, G.A.; Riaño-Umbarila, L.; Becerril, B. Broadening the neutralizing capacity of a family of antibody fragments against different toxins from Mexican scorpions. Toxicon 2016, 119, 52–63. [Google Scholar] [CrossRef] [PubMed]
- Laustsen, A.H.; Lohse, B.; Lomonte, B.; Engmark, M.; Gutiérrez, J.M. Selecting key toxins for focused development of elapid snake antivenoms and inhibitors guided by a Toxicity Score. Toxicon 2015, 104, 43–45. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abdel-Rahman, M.A.; Harrison, P.L.; Strong, P.N. Snapshots of scorpion venomics. J. Arid Environ. 2015, 112 Pt B, 170–176. [Google Scholar] [CrossRef]
- Shlyapnikov, Y.M.; Kozlov, S.A.; Fedorov, A.A.; Grishin, E.V. A comparison of polypeptide compositions of individual Agelena orientalis spider venoms. Russ. J. Bioorganic Chem. 2010, 36, 73–80. [Google Scholar] [CrossRef]
- Duan, Z.; Cao, R.; Jiang, L.; Liang, S. A combined de novo protein sequencing and cDNA library approach to the venomic analysis of Chinese spider Araneus ventricosus. J. Proteomics 2013, 78, 416–427. [Google Scholar] [CrossRef] [PubMed]
- Undheim, E.A.B.; Sunagar, K.; Herzig, V.; Kely, L.; Low, D.H.W.; Jackson, T.N.W.; Jones, A.; Kurniawan, N.; King, G.F.; Ali, S.A.; et al. A Proteomics and Transcriptomics Investigation of the Venom from the Barychelid Spider Trittame loki (Brush-Foot Trapdoor). Toxins 2013, 5, 2488–2503. [Google Scholar] [CrossRef] [PubMed]
- Trachsel, C.; Siegemund, D.; Kämpfer, U.; Kopp, L.S.; Bühr, C.; Grossmann, J.; Lüthi, C.; Cunningham, M.; Nentwig, W.; Kuhn-Nentwig, L.; et al. Multicomponent venom of the spider Cupiennius salei: A bioanalytical investigation applying different strategies. FEBS J. 2012, 279, 2683–2694. [Google Scholar] [CrossRef] [PubMed]
- Estrada-Gomez, S.; Muñoz, L.J.V.; Lanchero, P.; Latorre, C.S. Partial Characterization of Venom from the Colombian Spider Phoneutria Boliviensis (Aranae: Ctenidae). Toxins 2015, 7, 2872–2887. [Google Scholar] [CrossRef] [PubMed]
- Diniz, M.R.V.; Machado, C.R.L.; Mudado, M.A.; Paiva, A.L.B. 257. Comparative Analysis of Transcriptomes of Phoneutria pertyi and P. nigriventer Venom Glands. Toxicon 2012, 60, 127–128. [Google Scholar] [CrossRef]
- Liberato, T.; Troncone, L.R.P.; Yamashiro, E.T.; Serrano, S.M.T.; Zelanis, A. High-resolution proteomic profiling of spider venom: Expanding the toxin diversity of Phoneutria nigriventer venom. Amino Acids 2016, 48, 901–906. [Google Scholar] [CrossRef] [PubMed]
- Richardson, M.; Pimenta, A.M.C.; Bemquerer, M.P.; Santoro, M.M.; Beirao, P.S.L.; Lima, M.E.; Figueiredo, S.G.; Bloch, C., Jr.; Vasconcelos, E.A.R.; Campos, F.A.P.; et al. Comparison of the partial proteomes of the venoms of Brazilian spiders of the genus Phoneutria. Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2006, 142, 173–187. [Google Scholar] [CrossRef] [PubMed]
- Pineda, S.S.; Jones, A.; Nicholson, G.M.; Escoubas, P.; Mattick, J.S.; King, G.F. Understanding the Chemical Diversity of Spider Venoms Using a Combined Genomic, Transcriptomic and Proteomic Approach. Toxicon 2012, 60. [Google Scholar] [CrossRef]
- Zhang, Y.; Chen, J.; Tang, X.; Wang, F.; Jiang, L.; Xiong, X.; Wang, M.; Rong, M.; Liu, Z.; Liang, S. Transcriptome analysis of the venom glands of the Chinese wolf spider Lycosa singoriensis. Zoology 2010, 113, 10–18. [Google Scholar] [CrossRef] [PubMed]
- Zhang, F.; Liu, C.; Tan, H.; Wang, H.; Jiang, Y.; Liang, S.; Zhang, F.; Liu, Z. A survey of the venom of the spider Lycosa vittata by biochemical, pharmacological and transcriptomic analyses. Toxicon 2015, 107 Pt B, 335–343. [Google Scholar] [CrossRef] [PubMed]
- Kozlov, S.A.; Lazarev, V.N.; Kostryukova, E.S.; Selezneva, O.V.; Ospanova, E.A.; Alexeev, D.G.; Govorun, V.M.; Grishin, E.V. Comprehensive analysis of the venom gland transcriptome of the spider Dolomedes fimbriatus. Sci. Data 2014, 1, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Jiang, L.; Liu, C.; Duan, Z.; Deng, M.; Tang, X.; Liang, S. Transcriptome analysis of venom glands from a single fishing spider Dolomedes mizhoanus. Toxicon 2013, 73, 23–32. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.; Wang, H.; Zhang, F.; Hu, Z.; Liang, S.; Liu, Z. A Comparative Analysis of the Venom Gland Transcriptomes of the Fishing Spiders Dolomedes mizhoanus and Dolomedes sulfurous. PLoS ONE 2015, 10. [Google Scholar] [CrossRef] [PubMed]
- Zobel-Thropp, P.A.; Thomas, E.Z.; David, C.L.; Breci, L.A.; Binford, G.J. Plectreurys tristis venome: A proteomic and transcriptomic analysis. J. Venom Res. 2014, 5, 33–47. [Google Scholar] [PubMed]
- Machado, L.F.; Laugesen, S.; Botelho, E.D.; Ricart, C.A.O.; Fontes, W.; Barbaro, K.C.; Roepstorff, P.; Sousa, M.V. Proteome analysis of brown spider venom: identification of loxnecrogin isoforms in Loxosceles gaucho venom. Proteomics 2005, 5, 2167–2176. [Google Scholar] [CrossRef] [PubMed]
- dos Santos, L.D.; Dias, N.B.; Roberto, J.; Pinto, A.S.; Palma, M.S. Brown recluse spider venom: proteomic analysis and proposal of a putative mechanism of action. Protein Pept. Lett. 2009, 16, 933–943. [Google Scholar] [CrossRef] [PubMed]
- Gremski, L.H.; da Silveira, R.B.; Chaim, O.M.; Probst, C.M.; Ferrer, V.P.; Nowatzki, J.; Weinschutz, H.C.; Madeira, H.M.; Gremski, W.; Nader, H.B.; et al. A novel expression profile of the Loxosceles intermedia spider venomous gland revealed by transcriptome analysis. Mol. Biosyst. 2010, 6, 2403–2416. [Google Scholar] [CrossRef] [PubMed]
- Fernandes-Pedrosa, M. de F.; Junqueira-de-Azevedo, I. de L.; Gonçalves-de-Andrade, R.M.; Kobashi, L.S.; Almeida, D.D.; Ho, P.L.; Tambourgi, D.V. Transcriptome analysis of Loxosceles laeta (Araneae, Sicariidae) spider venomous gland using expressed sequence tags. BMC Genomics 2008, 9. [Google Scholar] [CrossRef] [PubMed]
- Zobel-Thropp, P.A.; Correa, S.M.; Garb, J.E.; Binford, G.J. Spit and Venom from Scytodes Spiders: A Diverse and Distinct Cocktail. J. Proteome Res. 2014, 13, 817–835. [Google Scholar] [CrossRef] [PubMed]
- Mourão, C.B.F.; Oliveira, F.N.; e Carvalho, A.C.; Arenas, C.J.; Duque, H.M.; Gonçalves, J.C.; Macêdo, J.K.A.; Galante, P.; Schwartz, C.A.; Mortari, M.R.; et al. Venomic and pharmacological activity of Acanthoscurria paulensis (Theraphosidae) spider venom. Toxicon 2013, 61, 129–138. [Google Scholar]
- Liao, Z.; Cao, J.; Li, S.; Yan, X.; Hu, W.; He, Q.; Chen, J.; Tang, J.; Xie, J.; Liang, S. Proteomic and peptidomic analysis of the venom from Chinese tarantula Chilobrachys jingzhao. Proteomics 2007, 7, 1892–1907. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Zhao, L.; Jiang, L.; Meng, E.; Zhang, Y.; Xiong, X.; Liang, S. Transcriptome analysis revealed novel possible venom components and cellular processes of the tarantula Chilobrachys jingzhao venom gland. Toxicon 2008, 52, 794–806. [Google Scholar] [CrossRef] [PubMed]
- Diego-García, E.; Peigneur, S.; Waelkens, E.; Debaveye, S.; Tytgat, J. Venom components from Citharischius crawshayi spider (Family Theraphosidae): Exploring transcriptome, venomics, and function. Cell. Mol. Life Sci. 2010, 67, 2799–2813. [Google Scholar] [CrossRef] [PubMed]
- Borges, M.H.; Figueiredo, S.G.; Leprevost, F.V.; De Lima, M.E.; Cordeiro, M. do N.; Diniz, M.R.V.; Moresco, J.; Carvalho, P.C.; Yates, J.R. Venomous extract protein profile of Brazilian tarantula Grammostola iheringi: Searching for potential biotechnological applications. J. Proteomics 2016, 136, 35–47. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.-Y.; Huang, Y.; He, Q.-Z.; Luo, J.; Zhu, L.; Lu, S.-S.; Liu, J.-Y.; Huang, P.-F.; Zeng, X.-Z.; Liang, S.-P. Structural and Functional Diversity of Peptide Toxins from Tarantula Haplopelma hainanum (Ornithoctonus hainana) Venom Revealed by Transcriptomic, Peptidomic, and Patch Clamp Approaches. J. Biol. Chem. 2015, 290, 14192–14207. [Google Scholar] [CrossRef] [PubMed]
- Tang, X.; Zhang, Y.; Hu, W.; Xu, D.; Tao, H.; Yang, X.; Li, Y.; Jiang, L.; Liang, S. Molecular Diversification of Peptide Toxins from the Tarantula Haplopelma hainanum (Ornithoctonus hainana) Venom Based on Transcriptomic, Peptidomic, and Genomic Analyses. J. Proteome Res. 2010, 9, 2550–2564. [Google Scholar] [CrossRef] [PubMed]
- Cheng, T.-C.; Long, R.-W.; Wu, Y.-Q.; Guo, Y.-B.; Liu, D.-L.; Peng, L.; Li, D.-Q.; Yang, D.-W.; Xu, X.; Liu, F.-X.; Xia, Q.-Y. Identification and characterization of toxins in the venom gland of the Chinese bird spider, Haplopelma hainanum, by transcriptomic analysis. Insect Sci. 2016, 23, 487–499. [Google Scholar] [CrossRef] [PubMed]
- Jiang, L.; Zhang, D.; Zhang, Y.; Peng, L.; Chen, J.; Liang, S. Venomics of the spider Ornithoctonus huwena based on transcriptomic versus proteomic analysis. Comp. Biochem. Physiol. Part D Genomics Proteomics 2010, 5, 81–88. [Google Scholar] [CrossRef] [PubMed]
- Yuan, C.; Jin, Q.; Tang, X.; Hu, W.; Cao, R.; Yang, S.; Xiong, J.; Xie, C.; Xie, J.; Liang, S. Proteomic and Peptidomic Characterization of the Venom from the Chinese Bird Spider, Ornithoctonus huwena Wang. J. Proteome Res. 2007, 6, 2792–2801. [Google Scholar] [CrossRef] [PubMed]
- Liang, S.; Li, X.; Cao, M.; Xie, J.; Chen, P.; Huang, R. Indentification of venom proteins of spider S. huwena on two-dimensional electrophoresis gel by N-terminal microsequencing and mass spectrometric peptide mapping. J. Protein Chem. 2000, 19, 225–229. [Google Scholar] [CrossRef] [PubMed]
- Zhou, T.; Zhou, J.; Xiong, J.; Wang, X. A preliminary analysis of the toxinome of spider O. huwena based on isoelectric focusing-RP HPLC. Chin. J. Biochem. Mol. Biol. 2006, 22, 823–831. [Google Scholar]
- Zhang, Y.; Huang, Y.; He, Q.; Liu, J.; Luo, J.; Zhu, L.; Lu, S.; Huang, P.; Chen, X.; Zeng, X.; et al. Toxin Diversity Revealed by a Transcriptomic Study of Ornithoctonus huwena. PLoS ONE 2014, 9. [Google Scholar] [CrossRef] [PubMed]
- Haney, R.A.; Clarke, T.H.; Gadgil, R.; Fitzpatrick, R.; Hayashi, C.Y.; Ayoub, N.A.; Garb, J.E. Effects of Gene Duplication, Positive Selection, and Shifts in Gene Expression on the Evolution of the Venom Gland Transcriptome in Widow Spiders. Genome Biol. Evol. 2016, 8, 228–242. [Google Scholar] [CrossRef] [PubMed]
- Haney, R.A.; Ayoub, N.A.; Clarke, T.H.; Hayashi, C.Y.; Garb, J.E. Dramatic expansion of the black widow toxin arsenal uncovered by multi-tissue transcriptomics and venom proteomics. BMC Genomics 2014, 15. [Google Scholar] [CrossRef] [PubMed]
- Duan, Z.; Yan, X.; Cao, R.; Liu, Z.; Wang, X.; Liang, S. Proteomic analysis of Latrodectus tredecimguttatus venom for uncovering potential latrodectism-related proteins. J. Biochem. Mol. Toxicol. 2008, 22, 328–336. [Google Scholar] [CrossRef] [PubMed]
- Duan, Z.G.; Yan, X.J.; He, X.Z.; Zhou, H.; Chen, P.; Cao, R.; Xiong, J.X.; Hu, W.J.; Wang, X.C.; Liang, S.P. Extraction and protein component analysis of venom from the dissected venom glands of Latrodectus tredecimguttatus. Comp. Biochem. Physiol. B Biochem. Mol. Biol. 2006, 145, 350–357. [Google Scholar] [CrossRef] [PubMed]
- He, Q.; Duan, Z.; Yu, Y.; Liu, Z.; Liu, Z.; Liang, S. The Venom Gland Transcriptome of Latrodectus tredecimguttatus Revealed by Deep Sequencing and cDNA Library Analysis. PLoS ONE 2013, 8. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Shi, W.; Zeng, X.-C.; Ge, F.; Yang, M.; Nie, Y.; Bao, A.; Wu, S.; E, G. Unique diversity of the venom peptides from the scorpion Androctonus bicolor revealed by transcriptomic and proteomic analysis. J. Proteomics 2015, 128, 231–250. [Google Scholar] [CrossRef] [PubMed]
- Oukkache, N.; Rosso, J.P.; Alami, M.; Ghalim, N.; Saïle, R.; Hassar, M.; Bougis, P.E.; Martin-Eauclaire, M.F. New analysis of the toxic compounds from the Androctonus mauretanicus mauretanicus scorpion venom. Toxicon 2008, 51, 835–852. [Google Scholar] [CrossRef] [PubMed]
- Caliskan, F.; Quintero-Hernández, V.; Restano-Cassulini, R.; Batista, C.V.F.; Zamudio, F.Z.; Coronas, F.I.; Possani, L.D. Turkish scorpion Buthacus macrocentrus: General characterization of the venom and description of Bu1, a potent mammalian Na+-channelα-toxin. Toxicon 2012, 59, 408–415. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Zhang, X.; Guo, Z.; Yan, J.; Yu, L.; Li, X.; Xue, X.; Liang, X. Short-chain peptides identification of scorpion Buthus martensi Karsch venom by employing high orthogonal 2D-HPLC system and tandem mass spectrometry. Proteomics 2012, 12, 3076–3084. [Google Scholar] [CrossRef] [PubMed]
- Valdez-Velázquez, L.L.; Quintero-Hernández, V.; Romero-Gutiérrez, M.T.; Coronas, F.I.V.; Possani, L.D. Mass Fingerprinting of the Venom and Transcriptome of Venom Gland of Scorpion Centruroides tecomanus. PLoS ONE 2013, 8. [Google Scholar] [CrossRef]
- Mille, B.G.; Peigneur, S.; Predel, R.; Tytgat, J. Trancriptomic approach reveals the molecular diversity of Hottentotta conspersus (Buthidae) venom. Toxicon 2015, 99, 73–79. [Google Scholar] [CrossRef] [PubMed]
- Morgenstern, D.; Rohde, B.H.; King, G.F.; Tal, T.; Sher, D.; Zlotkin, E. The tale of a resting gland: Transcriptome of a replete venom gland from the scorpion Hottentotta judaicus. Toxicon 2011, 57, 695–703. [Google Scholar] [CrossRef] [PubMed]
- Ma, Y.; He, Y.; Zhao, R.; Wu, Y.; Li, W.; Cao, Z. Extreme diversity of scorpion venom peptides and proteins revealed by transcriptomic analysis: Implication for proteome evolution of scorpion venom arsenal. J. Proteomics 2012, 75, 1563–1576. [Google Scholar] [CrossRef] [PubMed]
- Nascimento, D.G.; Rates, B.; Santos, D.M.; Verano-Braga, T.; Barbosa-Silva, A.; Dutra, A.A.A.; Biondi, I.; Martin-Eauclaire, M.F.; De Lima, M.E.; Pimenta, A.M.C. Moving pieces in a taxonomic puzzle: Venom 2D-LC/MS and data clustering analyses to infer phylogenetic relationships in some scorpions from the Buthidae family (Scorpiones). Toxicon 2006, 47, 628–639. [Google Scholar] [CrossRef] [PubMed]
- Kuzmenkov, A.I.; Vassilevski, A.A.; Kudryashova, K.S.; Nekrasova, O.V.; Peigneur, S.; Tytgat, J.; Feofanov, A.V.; Kirpichnikov, M.P.; Grishin, E.V. Variability of potassium channel blockers in Mesobuthus eupeus scorpion venom with focus on Kv1.1: An integrated transcriptomic and proteomic study. J. Biol. Chem. 2015, 290, 12195–12209. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez-Ravelo, R.; Batista, C.V.F.; Coronas, F.I.V.; Zamudio, F.Z.; Hernández-Orihuela, L.; Espinosa-López, G.; Ruiz-Urquiola, A.; Possani, L.D. Comparative proteomic analysis of male and female venoms from the Cuban scorpion Rhopalurus junceus. Toxicon 2015, 107, 327–334. [Google Scholar]
- Rodríguez-Ravelo, R.; Coronas, F.I.V.; Zamudio, F.Z.; González-Morales, L.; López, G.E.; Urquiola, A.R.; Possani, L.D. The Cuban scorpion Rhopalurus junceus (Scorpiones, Buthidae): Component variations in venom samples collected in different geographical areas. J. Venom. Anim. Toxins Trop. Dis. 2013, 19. [Google Scholar] [CrossRef] [PubMed]
- García-Gómez, B.I.; Coronas, F.I.V.; Restano-Cassulini, R.; Rodríguez, R.R.; Possani, L.D. Biochemical and molecular characterization of the venom from the Cuban scorpion Rhopalurus junceus. Toxicon 2011, 58, 18–27. [Google Scholar] [CrossRef] [PubMed]
- de Oliveira, U.C.; Candido, D.M.; Coronado Dorce, V.A.; Junqueira-de-Azevedo, I. de L.M. The transcriptome recipe for the venom cocktail of Tityus bahiensis scorpion. Toxicon 2015, 95, 52–61. [Google Scholar] [CrossRef] [PubMed]
- Batista, C.V.F.; Del Pozo, L.; Zamudio, F.Z.; Contreras, S.; Becerril, B.; Wanke, E.; Possani, L.D. Proteomics of the venom from the Amazonian scorpion Tityus cambridgei and the role of prolines on mass spectrometry analysis of toxins. J. Chromatogr. B Analyt. Technol. Biomed. Life. Sci. 2004, 803, 55–66. [Google Scholar] [CrossRef] [PubMed]
- Diego-García, E.; Batista, C.V.F.; García-Gómez, B.I.; Lucas, S.; Candido, D.M.; Gómez-Lagunas, F.; Possani, L.D. The Brazilian scorpion Tityus costatus Karsch: Genes, peptides and function. Toxicon 2005, 45, 273–283. [Google Scholar] [CrossRef] [PubMed]
- Batista, C.V.F.; D’Suze, G.; Gómez-Lagunas, F.; Zamudio, F.Z.; Encarnación, S.; Sevcik, C.; Possani, L.D. Proteomic analysis of Tityus discrepans scorpion venom and amino acid sequence of novel toxins. Proteomics 2006, 6, 3718–3727. [Google Scholar] [CrossRef] [PubMed]
- Barona, J.; Batista, C.V.F.; Zamudio, F.Z.; Gomez-Lagunas, F.; Wanke, E.; Otero, R.; Possani, L.D. Proteomic analysis of the venom and characterization of toxins specific for Na+- and K+-channels from the Colombian scorpion Tityus pachyurus. Biochim. Biophys. Acta Proteins Proteomics 2006, 1764, 76–84. [Google Scholar] [CrossRef] [PubMed]
- Arantes, E.C.; Cologna, C.T.; Cardoso, I.A.; De Pauw, E. Identification and characterization of peptides and proteins from Tityus serrulatus venom by mass spectrometry. Amino Acids 2015, 47, 1608–1609. [Google Scholar]
- Alvarenga, E.R.; Mendes, T.M.; Magalhaes, B.F.; Siqueira, F.F.; Dantas, A.E.; Barroca, T.M.; Horta, C.C.; Kalapothakis, E. Transcriptome analysis of the Tityus serrulatus scorpion venom gland. Open J. Genet. 2012, 2, 210–220. [Google Scholar] [CrossRef]
- Fuzita, F.J.; Pinkse, M.W.H.; Patane, J.S.L.; Juliano, M.A.; Verhaert, P.D.E.M.; Lopes, A.R. Biochemical, Transcriptomic and Proteomic Analyses of Digestion in the Scorpion Tityus serrulatus : Insights into Function and Evolution of Digestion in an Ancient Arthropod. PLoS ONE 2015, 10. [Google Scholar] [CrossRef] [PubMed]
- Nascimento, D.G.; Rates, B.; Santos, D.M.; Verano-Braga, T.; Barbosa-Silva, A.; Dutra, A.A.A.; Biondi, I.; Martin-Eauclaire, M.F.; Lima, M.E.D.; Pimenta, A.M.C. Moving pieces in a taxonomic puzzle: Venom 2D-LC/MS and data clustering analyses to infer phylogenetic relationships in some scorpions from the Buthidae family (Scorpiones). Toxicon 2006, 47, 628–639. [Google Scholar] [CrossRef] [PubMed]
- Pimenta, A.M.C.; Almeida, F.D.M.; de Lima, M.E.; Martin-Eauclaire, M.F.; Bougis, P.E. Individual variability in Tityus serrulatus (Scorpiones, Buthidae) venom elicited by matrix-assisted laser desorption/ionization time-of-flight mass spectrometry. Rapid Commun. Mass Spectrom. 2003, 17, 413–418. [Google Scholar] [CrossRef] [PubMed]
- Pimenta, A.M.C.; Stcklin, R.; Favreau, P.; Bougis, P.E.; Martin-Eauclaire, M.F. Moving pieces in a proteomic puzzle: Mass fingerprinting of toxic fractions from the venom of Tityus serrulatus (Scorpiones, buthidae). Rapid Commun. Mass Spectrom. 2001, 15, 1562–1572. [Google Scholar] [CrossRef] [PubMed]
- Rates, B.; Ferraz, K.K.F.; Borges, M.H.; Richardson, M.; De Lima, M.E.; Pimenta, A.M.C. Tityus serrulatus venom peptidomics: Assessing venom peptide diversity. Toxicon 2008, 52, 611–618. [Google Scholar] [CrossRef] [PubMed]
- Verano-Braga, T.; Dutra, A.A.A.; León, I.R.; Melo-Braga, M.N.; Roepstorff, P.; Pimenta, A.M.C.; Kjeldsen, F. Moving Pieces in a Venomic Puzzle: Unveiling Post-translationally Modified Toxins from Tityus serrulatus. J. Proteome Res. 2013, 12, 3460–3470. [Google Scholar] [CrossRef] [PubMed]
- Verano-Braga, T.; Rocha-Resende, C.; Silva, D.M.; Ianzer, D.; Martin-Eauclaire, M.F.; Bougis, P.E.; de Lima, M.E.; Santos, R.A.S.; Pimenta, A.M.C. Tityus serrulatus Hypotensins: A new family of peptides from scorpion venom. Biochem. Biophys. Res. Commun. 2008, 371, 515–520. [Google Scholar] [CrossRef] [PubMed]
- Almeida, D.D.; Scortecci, K.C.; Kobashi, L.S.; Agnez-Lima, L.F.; Medeiros, S.R.B.; Silva-Junior, A.A.; Junqueira-de-Azevedo, I. de L.M.; Fernandes-Pedrosa, M. de F. Profiling the resting venom gland of the scorpion Tityus stigmurus through a transcriptomic survey. BMC Genomics 2012, 13. [Google Scholar] [CrossRef] [PubMed]
- Batista, C.V.F.; Román-González, S.A.; Salas-Castillo, S.P.; Zamudio, F.Z.; Gómez-Lagunas, F.; Possani, L.D. Proteomic analysis of the venom from the scorpion Tityus stigmurus: Biochemical and physiological comparison with other Tityus species. Comp. Biochem. Physiol. C Toxicol. Pharmacol. 2007, 146, 147–157. [Google Scholar] [CrossRef] [PubMed]
- Schwartz, E.F.; Diego-García, E.; Rodríguez de la Vega, R.C.; Possani, L.D. Transcriptome analysis of the venom gland of the Mexican scorpion Hadrurus gertschi (Arachnida: Scorpiones). BMC Genomics 2007, 8, 865–879. [Google Scholar] [CrossRef] [PubMed]
- He, Y.; Zhao, R.; Di, Z.; Li, Z.; Xu, X.; Hong, W.; Wu, Y.; Zhao, H.; Li, W.; Cao, Z. Molecular diversity of Chaerilidae venom peptides reveals the dynamic evolution of scorpion venom components from Buthidae to non-Buthidae. J. Proteomics 2013, 89, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Ma, Y.; Zhao, R.; He, Y.; Li, S.; Liu, J.; Wu, Y.; Cao, Z.; Li, W. Transcriptome analysis of the venom gland of the scorpion Scorpiops jendeki: Implication for the evolution of the scorpion venom arsenal. BMC Genomics 2009, 10. [Google Scholar] [CrossRef] [PubMed]
- Taghavi Moghadam, A. The comparison of the proteinic pattern of the venoms of scorpion Hemiscorpius lepturus and Hemiscorpius persicus. Clin. Biochem. 2011, 44. [Google Scholar] [CrossRef]
- Schwartz, E.F.; Camargos, T.S.; Zamudio, F.Z.; Silva, L.P.; Bloch, C.; Caixeta, F.; Schwartz, C.A.; Possani, L.D. Mass spectrometry analysis, amino acid sequence and biological activity of venom components from the Brazilian scorpion Opisthacanthus cayaporum. Toxicon 2008, 51, 1499–1508. [Google Scholar] [CrossRef] [PubMed]
- Estrada-Gómez, S.; Vargas Muñoz, L.J.; Saldarriaga-Córdoba, M.; Quintana Castillo, J.C. Venom from Opisthacanthus elatus scorpion of Colombia, could be more hemolytic and less neurotoxic than thought. Acta Trop. 2016, 153, 70–78. [Google Scholar] [CrossRef] [PubMed]
- Bringans, S.; Eriksen, S.; Kendrick, T.; Gopalakrishnakone, P.; Livk, A.; Lock, R.; Lipscombe, R. Proteomic analysis of the venom of Heterometrus longimanus (Asian black scorpion). Proteomics 2008, 8, 1081–1096. [Google Scholar] [CrossRef] [PubMed]
- Ma, Y.; Zhao, Y.; Zhao, R.; Zhang, W.; He, Y.; Wu, Y.; Cao, Z.; Guo, L.; Li, W. Molecular diversity of toxic components from the scorpion Heterometrus petersii venom revealed by proteomic and transcriptome analysis. Proteomics 2010, 10, 2471–2485. [Google Scholar] [CrossRef] [PubMed]
- Diego-García, E.; Peigneur, S.; Clynen, E.; Marien, T.; Czech, L.; Schoofs, L.; Tytgat, J. Molecular diversity of the telson and venom components from Pandinus cavimanus (Scorpionidae Latreille 1802): Transcriptome, venomics and function. Proteomics 2012, 12, 313–328. [Google Scholar] [CrossRef] [PubMed]
- Abdel-Rahman, M.A.; Quintero-Hernandez, V.; Possani, L.D. Venom proteomic and venomous glands transcriptomic analysis of the Egyptian scorpion Scorpio maurus palmatus (Arachnida: Scorpionidae). Toxicon 2013, 74, 193–207. [Google Scholar] [CrossRef] [PubMed]
- Ramirez, K.L.; Hernandez, V.Q.; Romero, E.M.; Winkel, K.; Batista, C.F.; Possani, L.D. 173. Characterisation of the Venom of an Australian Scorpion, Urodacus yaschenkoi: Proteome and Transcriptome Analysis. Toxicon 2012, 60, 184–185. [Google Scholar] [CrossRef]
- Luna-Ramírez, K.; Quintero-Hernández, V.; Juárez-González, V.R.; Possani, L.D. Whole Transcriptome of the Venom Gland from Urodacus yaschenkoi Scorpion. PLoS ONE 2015, 10. [Google Scholar] [CrossRef]
- Luna-Ramírez, K.; Quintero-Hernández, V.; Vargas-Jaimes, L.; Batista, C.V.F.; Winkel, K.D.; Possani, L.D. Characterization of the venom from the Australian scorpion Urodacus yaschenkoi: Molecular mass analysis of components, cDNA sequences and peptides with antimicrobial activity. Toxicon 2013, 63, 44–54. [Google Scholar] [CrossRef] [PubMed]
- Laustsen, A.H.; Engmark, M.; Milbo, C.; Johannesen, J.; Lomonte, B.; Gutiérrez, J.M.; Lohse, B. From Fangs to Pharmacology: The Future of Snakebite Envenoming Therapy. Curr. Pharm. Des. 2016, 22. [Google Scholar]
- Feng, L.; Gao, R.; Gopalakrishnakone, P. Isolation and characterization of a hyaluronidase from the venom of Chinese red scorpion Buthus martensi. Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2008, 148, 250–257. [Google Scholar] [CrossRef] [PubMed]
- Morey, S.S.; Kiran, K.M.; Gadag, J.R. Purification and properties of hyaluronidase from Palamneus gravimanus (Indian black scorpion) venom. Toxicon 2006, 47, 188–195. [Google Scholar] [CrossRef] [PubMed]
- Parthasarathy, P.; Venkaiah, B.; Ramanaiah, M. Isolation and characterization of hyaluronidase from scorpion (heterometrus-fulvipes) venom. Biochem. Int. 1990, 20, 301–310. [Google Scholar]
- Horta, C.C.R.; Magalhães, B. de F.; Oliveira-Mendes, B.B.R.; do Carmo, A.O.; Duarte, C.G.; Felicori, L.F.; Machado-de-Ávila, R.A.; Chávez-Olórtegui, C.; Kalapothakis, E. Molecular, Immunological, and Biological Characterization of Tityus serrulatus Venom Hyaluronidase: New Insights into Its Role in Envenomation. PLoS Negl. Trop. Dis. 2014, 8. [Google Scholar] [CrossRef] [PubMed]
- Borchani, L.; Sassi, A.; Ben Gharsa, H.; Safra, I.; Shahbazzadeh, D.; Ben Lasfar, Z.; El Ayeb, M. The pathological effects of Heminecrolysin, a dermonecrotic toxin from Hemiscorpius lepturus scorpion venom are mediated through its lysophospholipase D activity. Toxicon 2013, 68, 30–39. [Google Scholar] [CrossRef] [PubMed]
- Feitosa, L.; Gremski, W.; Veiga, S.S.; Elias, M.C.Q.B.; Graner, E.; Mangili, O.C.; Brentani, R.R. Detection and characterization of metalloproteinases with gelatinolytic, fibronectinolytic and fibrinogenolytic activities in Brown spider (Loxosceles intermedia) venom. Toxicon 1998, 36, 1039–1051. [Google Scholar] [CrossRef]
- Amir, R.; Alam, J.M.; Khan, M.A. Isolation and biological properties of scorpionvenom proteins: IV-comparative study on four proteinases isolated from the venom of isometrus vittatus. Pak. J. Pharm. Sci. 1994, 7, 21–32. [Google Scholar] [PubMed]
- Carmo, A.O.; Oliveira-Mendes, B.B.R.; Horta, C.C.R.; Magalhães, B.F.; Dantas, A.E.; Chaves, L.M.; Chávez-Olórtegui, C.; Kalapothakis, E. Molecular and functional characterization of metalloserrulases, new metalloproteases from the Tityus serrulatus venom gland. Toxicon 2014, 90, 45–55. [Google Scholar] [CrossRef] [PubMed]
- Gimenez, G.S.; Coutinho-Neto, A.; Kayano, A.M.; Simões-Silva, R.; Trindade, F.; de Almeida e Silva, A.; Marcussi, S.; da Silva, S.L.; Fernandes, C.F.C.; Zuliani, J.P.; et al. Biochemical and Functional Characterization of Parawixia bistriata Spider Venom with Potential Proteolytic and Larvicidal Activities, Biochemical and Functional Characterization of Parawixia bistriata Spider Venom with Potential Proteolytic and Larvicidal Activities. Biomed. Res. Int. 2014, 2014. [Google Scholar] [CrossRef]
- Nagaraju, S.; Girish, K.S.; Fox, J.W.; Kemparaju, K. “Partitagin” a hemorrhagic metalloprotease from Hippasa partita spider venom: Role in tissue necrosis. Biochimie 2007, 89, 1322–1331. [Google Scholar] [CrossRef] [PubMed]
- Fletcher, P.L.; Fletcher, M.D.; Weninger, K.; Anderson, T.E.; Martin, B.M. Vesicle-associated Membrane Protein (VAMP) Cleavage by a New Metalloprotease from the Brazilian Scorpion Tityus serrulatus. J. Biol. Chem. 2010, 285, 7405–7416. [Google Scholar] [CrossRef] [PubMed]
- Ramanaiah, M.; Parthasarathy, Pr.; Venkaiah, B. Isolation and characterization of hyaluronidase from scorpion (Heterometrus fulvipes) venom. Biochem. Int. 1989, 20, 301–310. [Google Scholar]
- Atkinson, R.K.; Wright, L.G. A study of the necrotic actions of the venom of the wolf spider, Lycosa godeffroyi, on mouse skin. Comp. Biochem. Physiol. Part C Comp. Pharmacol. 1990, 95, 319–325. [Google Scholar] [CrossRef]
- Rees, R.; Shack, R.B.; Withers, E.; Madden, J.; Franklin, J.; Lynch, J.B. Management of the brown recluse spider bite. Plast. Reconstr. Surg. 1981, 68, 768–773. [Google Scholar] [CrossRef] [PubMed]
- Farace, F.; Lissia, M.; Mele, A.; Masia, D.R.; Rubino, C. Local cutaneous arachnidism: A report of three cases and their management. J. Plast. Reconstr. Aesthet. Surg. 2006, 59, 197–201. [Google Scholar] [CrossRef] [PubMed]
- El-Kik, C.Z.; Fernandes, F.F.A.; Tomaz, M.A.; Gaban, G.A.; Fonseca, T.F.; Calil-Elias, S.; Oliveira, S.D.S.; Silva, C.L.M.; Martinez, A.M.B.; Melo, P.A. Neutralization of Apis mellifera bee venom activities by suramin. Toxicon 2010, 67, 55–62. [Google Scholar]
- Petricevich, V.L. Scorpion venom and the inflammatory response. Mediators Inflamm. 2010, 2010, 903295. [Google Scholar] [CrossRef] [PubMed]
- Bahraoui, E.; Pichon, J.; Muller, J.M.; Darbon, H.; Elayeb, M.; Granier, C.; Marvaldi, J.; Rochat, H. Monoclonal antibodies to scorpion toxins. Characterization and molecular mechanisms of neutralization. J. Immunol. 1988, 141, 214–220. [Google Scholar] [PubMed]
- Köhler, G.; Milstein, C. Continuous cultures of fused cells secreting antibody of predefined specificity. Nature 1975, 256, 495–497. [Google Scholar] [CrossRef] [PubMed]
- Clot-Faybesse, O.; Juin, M.; Rochat, H.; Devaux, C. Monoclonal antibodies against the Androctonus australis hector scorpion neurotoxin I: Characterisation and use for venom neutralisation. FEBS Lett. 1999, 458, 313–318. [Google Scholar] [CrossRef]
- Alvarenga, L.M.; Martins, M.S.; Moura, J.F.; Kalapothakis, E.; Oliveira, J.C.; Mangili, O.C.; Granier, C.; Chávez-Olórtegui, C. Production of monoclonal antibodies capable of neutralizing dermonecrotic activity of Loxosceles intermedia spider venom and their use in a specific immunometric assay. Toxicon 2003, 42, 725–731. [Google Scholar] [CrossRef] [PubMed]
- Bugli, F.; Graffeo, R.; Sterbini, F.P.; Torelli, R.; Masucci, L.; Sali, M.; Grasso, A.; Rufini, S.; Ricci, E.; Fadda, G.; Pescatori, M. Monoclonal antibody fragment from combinatorial phage display library neutralizes alpha-latrotoxin activity and abolishes black widow spider venom lethality, in mice. Toxicon 2008, 51, 547–554. [Google Scholar] [CrossRef] [PubMed]
- Dias-Lopes, C.; Felicori, L.; Rubrecht, L.; Cobo, S.; Molina, L.; Nguyen, C.; Galéa, P.; Granier, C.; Molina, F.; Chávez-Olortegui, C. Generation and molecular characterization of a monoclonal antibody reactive with conserved epitope in sphingomyelinases D from Loxosceles spider venoms. Vaccine 2014, 32, 2086–2092. [Google Scholar] [CrossRef] [PubMed]
- Zamudio, F.; Saavedra, R.; Martin, B.M.; Gurrola-Briones, G.; Hérion, P.; Possani, L.D. Amino acid sequence and immunological characterization with monoclonal antibodies of two toxins from the venom of the scorpion Centruroides noxius Hoffmann. Eur. J. Biochem. 1992, 204, 281–292. [Google Scholar] [CrossRef] [PubMed]
- Licea, A.F.; Becerril, B.; Possani, L.D. Fab fragments of the monoclonal antibody BCF2 are capable of neutralizing the whole soluble venom from the scorpion Centruroides noxius Hoffmann. Toxicon 1996, 34, 843–847. [Google Scholar] [CrossRef]
- Mousli, M.; Devaux, C.; Rochat, H.; Goyffon, M.; Billiald, P. A recombinant single-chain antibody fragment that neutralizes toxin II from the venom of the scorpion Androctonus australis hector. FEBS Lett. 1999, 442, 183–188. [Google Scholar] [CrossRef]
- Jia, L.-Y.; Xie, H.-F.; Ji, Y.-H. Characterization of four distinct monoclonal antibodies specific to BmK AS-1, a novel scorpion bioactive polypeptide. Toxicon 2000, 38, 605–617. [Google Scholar] [CrossRef]
- Devaux, C.; Moreau, E.; Goyffon, M.; Rochat, H.; Billiald, P. Construction and functional evaluation of a single-chain antibody fragment that neutralizes toxin AahI from the venom of the scorpion Androctonus australis hector. Eur. J. Biochem. 2001, 268, 694–702. [Google Scholar] [CrossRef] [PubMed]
- Selisko, B.; Cosı́o, G.; Garcı́a, C.; Becerril, B.; Possani, L.D.; Horjales, E. Bacterial expression, purification and functional characterization of a recombinant chimeric Fab derived from murine mAb BCF2 that neutralizes the venom of the scorpion Centruroides noxius hoffmann. Toxicon 2004, 43, 43–51. [Google Scholar] [CrossRef] [PubMed]
- Aubrey, N.; Muzard, J.; Christophe Peter, J.; Rochat, H.; Goyffon, M.; Devaux, C.; Billiald, P. Engineering of a recombinant Fab from a neutralizing IgG directed against scorpion neurotoxin AahI, and functional evaluation versus other antibody fragments. Toxicon 2004, 43, 233–241. [Google Scholar] [CrossRef] [PubMed]
- Juárez-González, V.R.; Riaño-Umbarila, L.; Quintero-Hernández, V.; Olamendi-Portugal, T.; Ortiz-León, M.; Ortíz, E.; Possani, L.D.; Becerril, B. Directed Evolution, Phage Display and Combination of Evolved Mutants: A Strategy to Recover the Neutralization Properties of the scFv Version of BCF2 a Neutralizing Monoclonal Antibody Specific to Scorpion Toxin Cn2. J. Mol. Biol. 2005, 346, 1287–1297. [Google Scholar] [CrossRef] [PubMed]
- Alvarenga, L.M.; Machado de Avila, R.A.; Amim, P.R.; Martins, M.S.; Kalapothakis, E.; de Lima, M.E.; Santos, R.G.; Granier, C.; Chávez-Olórtegui, C. Molecular characterization of a neutralizing murine monoclonal antibody against Tityus serrulatus scorpion venom. Toxicon 2005, 46, 664–671. [Google Scholar] [CrossRef] [PubMed]
- Juste, M.; Martin-Eauclaire, M.F.; Devaux, C.; Billiald, P.; Aubrey, N. Using a recombinant bispecific antibody to block Na+ -channel toxins protects against experimental scorpion envenoming. Cell. Mol. Life Sci. CMLS 2007, 64, 206–218. [Google Scholar] [CrossRef] [PubMed]
- Riaño-Umbarila, L.; Juárez-González, V.R.; Olamendi-Portugal, T.; Ortíz-León, M.; Possani, L.D.; Becerril, B. A strategy for the generation of specific human antibodies by directed evolution and phage display. FEBS J. 2005, 272, 2591–2601. [Google Scholar] [CrossRef] [PubMed]
- Hmila, I.; Abdallah R, B.A.-B.; Saerens, D.; Benlasfar, Z.; Conrath, K.; Ayeb, M.E.; Muyldermans, S.; Bouhaouala-Zahar, B. VHH, bivalent domains and chimeric Heavy chain-only antibodies with high neutralizing efficacy for scorpion toxin AahI′. Mol. Immunol. 2008, 45, 3847–3856. [Google Scholar] [CrossRef] [PubMed]
- Abderrazek, R.B.; Hmila, I.; Vincke, C.; Benlasfar, Z.; Pellis, M.; Dabbek, H.; Saerens, D.; El Ayeb, M.; Muyldermans, S.; Bouhaouala-Zahar, B. Identification of potent nanobodies to neutralize the most poisonous polypeptide from scorpion venom. Biochem. J. 2009, 424, 263–272. [Google Scholar] [CrossRef] [PubMed]
- Hmila, I.; Saerens, D.; Abderrazek, R.B.; Vincke, C.; Abidi, N.; Benlasfar, Z.; Govaert, J.; Ayeb, M.E.; Bouhaouala-Zahar, B.; Muyldermans, S. A bispecific nanobody to provide full protection against lethal scorpion envenoming. FASEB J. 2010, 24, 3479–3489. [Google Scholar] [CrossRef] [PubMed]
- Riaño-Umbarila, L.; Contreras-Ferrat, G.; Olamendi-Portugal, T.; Morelos-Juárez, C.; Corzo, G.; Possani, L.D.; Becerril, B. Exploiting Cross-reactivity to Neutralize Two Different Scorpion Venoms with One Single Chain Antibody Fragment. J. Biol. Chem. 2011, 286, 6143–6151. [Google Scholar] [CrossRef] [PubMed]
- Amaro, I.; Riaño-Umbarila, L.; Becerril, B.; Possani, L.D. Isolation and characterization of a human antibody fragment specific for Ts1 toxin from Tityus serrulatus scorpion. Immunol. Lett. 2011, 139, 73–79. [Google Scholar] [CrossRef] [PubMed]
- Hmila, I.; Cosyns, B.; Tounsi, H.; Roosens, B.; Caveliers, V.; Abderrazek, R.B.; Boubaker, S.; Muyldermans, S.; El Ayeb, M.; Bouhaouala-Zahar, B.; et al. Pre-clinical studies of toxin-specific Nanobodies: Evidence of in vivo efficacy to prevent fatal disturbances provoked by scorpion envenoming. Toxicol. Appl. Pharmacol. 2012, 264, 222–231. [Google Scholar] [CrossRef] [PubMed]
- Di Tommaso, A.; Juste, M.O.; Martin-Eauclaire, M.-F.; Dimier-Poisson, I.; Billiald, P.; Aubrey, N. Diabody Mixture Providing Full Protection against Experimental Scorpion Envenoming with Crude Androctonus australis Venom. J. Biol. Chem. 2012, 287, 14149–14156. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez-Rodríguez, E.R.; Ledezma-Candanoza, L.M.; Contreras-Ferrat, L.G.; Olamendi-Portugal, T.; Possani, L.D.; Becerril, B.; Riaño-Umbarila, L. A Single Mutation in Framework 2 of the Heavy Variable Domain Improves the Properties of a Diabody and a Related Single-Chain Antibody. J. Mol. Biol. 2012, 423, 337–350. [Google Scholar] [CrossRef] [PubMed]
- Pucca, M.B.; Zoccal, K.F.; Roncolato, E.C.; Bertolini, T.B.; Campos, L.B.; Cologna, C.T.; Faccioli, L.H.; Arantes, E.C.; Barbosa, J.E. Serrumab: A human monoclonal antibody that counters the biochemical and immunological effects of Tityus serrulatus venom. J. Immunotoxicol. 2012, 9, 173–183. [Google Scholar] [CrossRef] [PubMed]
- Pucca, M.B.; Cerni, F.A.; Peigneur, S.; Arantes, E.C.; Tytgat, J.; Barbosa, J.E. Serrumab: A novel human single chain-fragment antibody with multiple scorpion toxin-neutralizing capacities. J. Immunotoxicol. 2014, 11, 133–140. [Google Scholar] [CrossRef] [PubMed]
- Riaño-Umbarila, L.; Olamendi-Portugal, T.; Morelos-Juárez, C.; Gurrola, G.B.; Possani, L.D.; Becerril, B. A novel human recombinant antibody fragment capable of neutralizing Mexican scorpion toxins. Toxicon 2013, 76, 370–376. [Google Scholar] [CrossRef] [PubMed]
- Riaño-Umbarila, L.; Ledezma-Candanoza, L.M.; Serrano-Posada, H.; Fernández-Taboada, G.; Olamendi-Portugal, T.; Rojas-Trejo, S.; Gómez-Ramírez, I.V.; Rudiño-Piñera, E.; Possani, L.D.; Becerril, B. Optimal Neutralization of Centruroides noxius Venom Is Understood through a Structural Complex between Two Antibody Fragments and the Cn2 Toxin. J. Biol. Chem. 2016, 291, 1619–1630. [Google Scholar] [CrossRef] [PubMed]
- Wörn, A.; Plückthun, A. Different Equilibrium Stability Behavior of ScFv Fragments: Identification, Classification, and Improvement by Protein Engineering. Biochemistry (Mosc.) 1999, 38, 8739–8750. [Google Scholar] [CrossRef] [PubMed]
- McCafferty, J.; Griffiths, A.D.; Winter, G.; Chiswell, D.J. Phage antibodies: Filamentous phage displaying antibody variable domains. Nature 1990, 348, 552–554. [Google Scholar] [CrossRef] [PubMed]
- Muyldermans, S. Nanobodies: Natural Single-Domain Antibodies. Annu. Rev. Biochem. 2013, 82, 775–797. [Google Scholar] [CrossRef] [PubMed]
- Cook, D.A.N.; Samarasekara, C.L.; Wagstaff, S.C.; Kinne, J.; Wernery, U.; Harrison, R.A. Analysis of camelid IgG for antivenom development: Immunoreactivity and preclinical neutralisation of venom-induced pathology by IgG subclasses, and the effect of heat treatment. Toxicon 2010, 56, 596–603. [Google Scholar] [CrossRef] [PubMed]
- Eyer, L.; Hruska, K. Single-domain antibody fragments derived from heavy-chain antibodies: a review. Vet. Med. (Praha) 2012, 57, 439–513. [Google Scholar]
- Richard, G.; Meyers, A.J.; McLean, M.D.; Arbabi-Ghahroudi, M.; MacKenzie, R.; Hall, J.C. In Vivo Neutralization of α-Cobratoxin with High-Affinity Llama Single-Domain Antibodies (VHHs) and a VHH-Fc Antibody. PLoS ONE 2013, 8. [Google Scholar] [CrossRef] [PubMed]
- Morrison, S.L.; Johnson, M.J.; Herzenberg, L.A.; Oi, V.T. Chimeric human antibody molecules: mouse antigen-binding domains with human constant region domains. Proc. Natl. Acad. Sci. USA. 1984, 81, 6851–6855. [Google Scholar] [CrossRef] [PubMed]
- Vaswani, S.K.; Hamilton, R.G. Humanized Antibodies as Potential Therapeutic Drugs. Ann. Allergy Asthma Immunol. 1998, 81, 105–119. [Google Scholar] [CrossRef]
- Vincke, C.; Loris, R.; Saerens, D.; Martinez-Rodriguez, S.; Muyldermans, S.; Conrath, K. General Strategy to Humanize a Camelid Single-domain Antibody and Identification of a Universal Humanized Nanobody Scaffold. J. Biol. Chem. 2009, 284, 3273–3284. [Google Scholar] [CrossRef] [PubMed]
- Birch, J.R.; Racher, A.J. Antibody production. Adv. Drug Deliv. Rev. 2006, 58, 671–685. [Google Scholar] [CrossRef] [PubMed]
- Kelley, B. Industrialization of mAb production technology: The bioprocessing industry at a crossroads. mAbs 2009, 1, 443–452. [Google Scholar] [CrossRef] [PubMed]
- Araujo, S.C.; Castanheira, P.; Alvarenga, L.M.; Mangili, O.C.; Kalapothakis, E.; Chávez-Olórtegui, C. Protection against dermonecrotic and lethal activities of Loxosceles intermedia spider venom by immunization with a fused recombinant protein. Toxicon 2003, 41, 261–267. [Google Scholar] [CrossRef]
- Felicori, L.; Fernandes, P.B.; Giusta, M.S.; Duarte, C.G.; Kalapothakis, E.; Nguyen, C.; Molina, F.; Granier, C.; Chávez-Olórtegui, C. An in vivo protective response against toxic effects of the dermonecrotic protein from Loxoscelesintermedia spider venom elicited by synthetic epitopes. Vaccine 2009, 27, 4201–4208. [Google Scholar] [CrossRef] [PubMed]
- Comis, A.; Tyler, M.; Mylecharane, E.; Spence, I.; Howden, M. Immunization with a synthetic robustoxin derivative lacking disulphide bridges protects against a potentially lethal challenge with funnel-web spider (Atrax robustus) venom. J. Biosci. 2009, 34, 35–44. [Google Scholar] [CrossRef] [PubMed]
- Mendes, T.M.; Oliveira, D.; Figueiredo, L.F.M.; Machado-de-Avila, R.A.; Duarte, C.G.; Dias-Lopes, C.; Guimarães, G.; Felicori, L.; Minozzo, J.C.; Chávez-Olortegui, C. Generation and characterization of a recombinant chimeric protein (rCpLi) consisting of B-cell epitopes of a dermonecrotic protein from Loxosceles intermedia spider venom. Vaccine 2013, 31, 2749–2755. [Google Scholar] [CrossRef] [PubMed]
- Bahraoui, E.M.; Granier, C.; Rietschoten, J.V.; Rochat, H.; El Ayeb, M. Specificity and neutralizing capacity of antibodies elicited by a synthetic peptide of scorpion toxin. J. Immunol. 1986, 136, 3371–3377. [Google Scholar] [PubMed]
- Calderon-Aranda, E.S.; Olamendi-Portugal, T.; Possani, L.D. The use of synthetic peptides can be a misleading approach to generate vaccines against scorpion toxins. Vaccine 1995, 13, 1198–1206. [Google Scholar] [CrossRef]
- Bouhaouala-Zahar, B.; Ducancel, F.; Zenouaki, I.; Ben Khalifa, R.; Borchani, L.; Pelhate, M.; Boulain, J.-C.; El Ayeb, M.; Ménez, A.; Karoui, H. A Recombinant Insect-Specific α-Toxin of Buthus occitanus tunetanus Scorpion Confers Protection Against Homologous Mammal Toxins. Eur. J. Biochem. 1996, 238, 653–660. [Google Scholar] [CrossRef] [PubMed]
- Devaux, C.; Clot-Faybesse, O.; Juin, M.; Mabrouk, K.; Sabatier, J.-M.; Rochat, H. Monoclonal antibodies neutralizing the toxin II from Androctonus australis hector scorpion venom: Usefulness of a synthetic, non-toxic analog. FEBS Lett. 1997, 412, 456–460. [Google Scholar] [CrossRef]
- Calderon-Aranda, E.S.; Selisko, B.; York, E.J.; Gurrola, G.B.; Stewart, J.M.; Possani, L.D. Mapping of an epitope recognized by a neutralizing monoclonal antibody specific to toxin Cn2 from the scorpion Centruroides noxius, using discontinuous synthetic peptides. Eur. J. Biochem. 1999, 264, 746–755. [Google Scholar] [CrossRef] [PubMed]
- Guatimosim, S.C.F.; Kalapothakis, E.; Diniz, C.R.; Chávez-Olórtegui, C. Induction of neutralizing antibodies against Tityus serrulatus toxins by immunization with a recombinant nontoxic protein. Toxicon 2000, 38, 113–121. [Google Scholar] [CrossRef]
- Legros, C.; Kaabi, H.; El Ayeb, M.; Céard, B.; Vacher, H.; Bougis, P.E.; Martin-Eauclaire, M.-F. Use of fusion protein constructs to generate potent immunotherapy and protection against scorpion toxins. Vaccine 2001, 20, 934–942. [Google Scholar] [CrossRef]
- Benkhadir, K.; Mejri, T.; Bel Haj Rhouma, R.; EI Ayeb, M.; Karoui, H. In vivo and in vitro protection against lethal activity of Buthus occitanus tunetanus venom with a recombinant protein. Arch. Inst. Pasteur Tunis 2002, 79, 19–26. [Google Scholar] [PubMed]
- Alvarenga, L.M.; Diniz, C.R.; Granier, C.; Chávez-Olórtegui, C. Induction of neutralizing antibodies against Tityus serrulatus scorpion toxins by immunization with a mixture of defined synthetic epitopes. Toxicon 2002, 40, 89–95. [Google Scholar] [CrossRef]
- Garcia, C.; Calderón-Aranda, E.S.; Anguiano, G.A.V.; Becerril, B.; Possani, L.D. Analysis of the immune response induced by a scorpion venom sub-fraction, a pure peptide and a recombinant peptide, against toxin Cn2 of Centruroides noxius Hoffmann. Toxicon 2003, 41, 417–427. [Google Scholar] [CrossRef]
- Inceoglu, B.; Lango, J.; Rabinovich, A.; Whetstone, P.; Hammock, B.D. The neutralizing effect of a polyclonal antibody raised against the N-terminal eighteen-aminoacid residues of birtoxin towards the whole venom of Parabuthus transvaalicus. Toxicon 2006, 47, 144–149. [Google Scholar] [CrossRef] [PubMed]
- Mendes, T.M.; Dias, F.; Horta, C.C.R.; Pena, I.F.; Arantes, E.C.; Kalapothakis, E. Effective Tityus serrulatus anti-venom produced using the Ts1 component. Toxicon 2008, 52, 787–793. [Google Scholar] [CrossRef] [PubMed]
- García-Gómez, B.I.; Olamendi-Portugal, T.C.; Paniagua, J.; van der Walt, J.; Dyason, K.; Possani, L.D. Heterologous expression of a gene that codes for Pg8, a scorpion toxin of Parabuthus granulatus, capable of generating protecting antibodies in mice. Toxicon 2009, 53, 770–778. [Google Scholar] [CrossRef] [PubMed]
- Hernández-Salgado, K.; Estrada, G.; Olvera, A.; Coronas, F.I.; Possani, L.D.; Corzo, G. Heterologous expressed toxic and non-toxic peptide variants of toxin CssII are capable to produce neutralizing antibodies against the venom of the scorpion Centruroides suffusus suffusus. Immunol. Lett. 2009, 125, 93–99. [Google Scholar] [CrossRef] [PubMed]
- Duarte, C.G.; Alvarenga, L.M.; Dias-Lopes, C.; Machado-de-Ávila, R.A.; Nguyen, C.; Molina, F.; Granier, C.; Chávez-Olórtegui, C. In vivo protection against Tityus serrulatus scorpion venom by antibodies raised against a discontinuous synthetic epitope. Vaccine 2010, 28, 1168–1176. [Google Scholar] [CrossRef] [PubMed]
- Machado de Avila, R.A.; Alvarenga, L.M.; Tavares, C.A.P.; Molina, F.; Granier, C.; Chávez-Olórtegui, C. Molecular characterization of protective antibodies raised in mice by Tityus serrulatus scorpion venom toxins conjugated to bovine serum albumin. Toxicon 2004, 44, 233–241. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Product Name | Producer | Country | Type | Spiders | Link Reference |
|---|---|---|---|---|---|
| Funnel web spider antivenom | CSL Ltd. | Australia | Equine F(ab’)2 | Hexathelidae family (funnel-web spiders) | [13] |
| Red Back Spider antivenom | CSL Ltd. | Australia | Equine F(ab’)2 | Latrodectus hasselti (redback spider) | [14] |
| Aracmyn | Instituto Bioclon | Mexico | Equine F(ab’)2 | Latrodectus mactans (black widow spider), Loxosceles spp. (recluse spiders) | [15] |
| Reclusmyn | Instituto Bioclon | Mexico | Equine F(ab’)2 | Loxosceles spp. (recluse spiders) | [16] |
| Soro antiarachnidico | Instituto Butantan | Brazil | Equine F(ab’)2 | Loxosceles spp. (recluse spiders), Phoneutria spp. (Brazilian wandering spiders) | [17] |
| Anti Latrodectus antivenom | Instituto Nacional de Biologics A.N.L.I.S. | Argentina | N/A | Latrodectus mactans (black widow spider) | [18] |
| Suero antiloxoscélico monovalente | Instituto Nacional de Salud, Perú | Perú | Equine IgG | Loxosceles spp. (recluse spiders) | [19] |
| Soro Antilatrodéctico | Instituto Vital Brazil | Brazil | Equine F(ab’)2 | Latrodectus mactans (black widow spider) | [20] |
| Antivenin (Latrodectus mactans) | Merck Sharp and Dohme International | USA | Equine IgG | Latrodectus mactans (black widow spider) | [21] |
| SAIMR Spider Antivenom | South African Vaccine Producers | South Africa | N/A | Latrodectus indistinctus (black button spider) | [22] |
| Product Name | Producer | Country | Type | Scorpions | Link Reference |
|---|---|---|---|---|---|
| Suero antialacran | BIRMEX | Mexico | N/A | Centruroides spp. (bark scorpions) | [23] |
| Suero antiescorpiónico | Centro de Biotecnologia de la Universidad central de Venezuela | Venezuela | Equine F(ab’)2 | Tityus spp. (thin-tailed scorpions) | [24] |
| Le sérum antiscorpionique (monovalent) | Institut Pasteur d’Algerie | Algeria | N/A | Androctonus australis (fat-tailed scorpion), Buthus occitanus (common yellow scorpion), Androctonus crasicauda (Arabian fat-tailed scorpion) | [25] |
| Scorpion antivenom | Institut Pasteur du Maroc | Morocco | Equine F(ab’)2 | Buthus occitanus (common yellow scorpion), Androctonus mauritanicus (Moroccan fat-tailed scorpion) | [26] |
| Le sérum antiscorpionique | Refik Saydam Hygiene Center | Turkey | Equine | Androctonus crassicauda (Arabian fat-tailed scorpion), Leiurus quinquestriatus (Israeli yellow scorpion) | [27] |
| Alacramyn | Instituto Bioclon | Mexico | Equine Fab | Centruroides spp. (bark scorpions) | [28] |
| Soro antiarachnidico | Instituto Butantan | Brazil | Equine F(ab’)2 | Tityus spp. (thin-tailed scorpions) | [17] |
| Soro antiescorpionico | Instituto Butantan | Brazil | Equine F(ab’)2 | Tityus bahiensis (black scorpion), Tityus serrulatus (Brazilian yellow scorpion) | [29] |
| Soro antiescorpiônico | Instituto Vital Brazil | Brazil | Equine F(ab’)2 | Buthus occitanus (common yellow scorpion) | [20] |
| Polyvalent Scorpion Antivenom | National Antivenom and Vaccine Production Center | Saudi Arabia | Equine F(ab’)2 | Leiurus quinquestriatus (Israeli yellow scorpion), Androctonus crassicauda (Arabian fat-tailed scorpion), Buthus arenicola, Buthus mimax, Buthus occitanus (common yellow scorpion), Androctonus amoreuxi (fat-tailed scorpion) | [30] |
| Le sérum antiscorpionique | Pasteur Tunis | North Africa | Equine F(ab’)2 | Androctonus australis (fat-tailed scorpion), Buthus occitanus (common yellow scorpion) | [31] |
| Monovalent Scorpion Antivenom | Razi Vaccine and Serum Research Institute | Iran | Equine | N/A | [32] |
| Polyvalent Scorpion Antivenom | Razi Vaccine and Serum Research Institute | Iran | Equine | Androctonus crasicauda (Arabian fat-tailed scorpion), Hemiscorpius lepturus, Hottentotta saulcyi, Hottentotta schach, Mesobuthus eupeus, Odontobuthus doriae | [32] |
| Scorpifav | Sanofi Pasteur | North Africa and Middle East | Equine F(ab’)2 | Androctonus australis (fat-tailed scorpion), Leiurus quinquestriatus (Israeli yellow scorpion), Buthus occitanus (common yellow scorpion) | [33] |
| SAIMR Scorpion Antivenom | South African Vaccine Producer | South Africa | Equine | Parabuthus transvaalicus (dark scorpion) | [22] |
| Scorpion antivenom Twyford | Twyford Pharmaceuticals | North Africa | N/A | Androctonus australis (fat-tailed scorpion), Buthus occitanus (common yellow scorpion), Leiurus quinquestriatus (Israeli yellow scorpion) | [34] |
| Purified Polyvalent anti-scorpion serum | VACSERA | Egypt | Equine F(ab’)2 | Leiurus quinquestriatus (Israeli yellow scorpion), Scorpio maurus (large-clawed scorpion), Androctonus crasicauda (Arabian fat-tailed scorpion), Buthus occitanus (common yellow scorpion) | [35] |
| Scorpion Venom Antiserum | Vins Bioproducts Ltd. | India | Equine IgG | Leiurus quinquestraitus (Israeli yellow scorpion), Androctonus amoreuxi (fat-tailed scorpion) | [36] |
| Soro Antiescorpiônico (FUNED) | Fundação Ezequiel Dias | Brazil | Equine F(ab’)2 | Tityus serrulatus, (Brazilian yellow scorpion) | [37] |
| Anti-scorpion Venom Serum | Haffkine Bio-Pharmaceutical Corporation Ltd. | India | Equine IgG | Buthus tamulus (red scorpion) | [38] |
| Family | Genus | Species | Prot. | Tran. | Reference |
|---|---|---|---|---|---|
| Agelenidae | Agelena | A. orientalis (funnel weaver spider) | √ | - | [48] |
| Araneidae | Araneus | A. ventricosus (Chinese orb-weaving spider) | √ | √ | [49] |
| Barychelidae | Trittame | T. loki (brush-foot trapdoor) | √ | √ | [50] |
| Ctenidae | Cupiennius | C. salei (tiger wandering spider) | √ | - | [51] |
| Phoneutria | P. boliviensis (male) (Brazilian wandering spider) | √ | - | [52] | |
| P. boliviensis (female) | √ | - | [52] | ||
| P. nigriventer (Brazilian wandering spider) | √ | √ | [53,54] | ||
| P. nigriventer (Minas Gerais, Brazil) | √ | - | [55] | ||
| P. pertyi | √ | [53] | |||
| P. keyserlingi (Minas Gerais, Brazil) | √ | - | [55] | ||
| P. reidyi (Amazonas, Brazil) (Brazilian wandering spider) | √ | - | [55] | ||
| P. reidyi (Para, Brazil) | √ | - | [55] | ||
| P. reidyi (Roraima, Brazil) | √ | - | [55] | ||
| Hexathelidae | Hadronyche | H. cerberea (male and female) (Australian funnel-web spider) | √ | - | [40] |
| H. infensa (Australian funnel-web spider) | √ | √ | [40,56] | ||
| Illawarra | I. wisharti (male) (Australian funnel-web spider) | √ | - | [40] | |
| Lycosidae | Lycosa | L. singoriensis (Chinese wolf spider) | - | √ | [57] |
| L. vittata (wolf spider) | - | √ | [58] | ||
| Pisauridae | Dolomedes | D. fimbriatus (raft spider) | - | √ | [59] |
| D. mizhoanus (fishing spider) | - | √ | [60,61] | ||
| D. sulfurous (fishing spider) | - | √ | [61] | ||
| Plectreuridae | Plectreurys | P. tristis (primitive hunting spiders) | √ | √ | [62] |
| Sicariidae | Loxosceles | L. gaucho (Brown spider) | √ | - | [63] |
| L. intermedia (brown recluse spider) | √ | √ | [64,65] | ||
| L. laeta (Chilean recluse spider) | - | √ | [66] | ||
| Scytodidae | Scytodes | S. thoracica (spitting spider) | √ | √ | [67] |
| Theraphosidae | Acanthoscurria | A. paulensis (Brazilian giant black tarantula) | √ | - | [68] |
| Chilobrachys | C. jingzhao (Chinese earth tiger tarantula) | √ | √ | [69,70] | |
| Citharischius | C. crawshayi (king baboon spider) | √ | √ | [71] | |
| Grammostola | G. iheringi (Argentinean black tarantula) | √ | - | [72] | |
| Haplopelma | H. hainanum (Chinese bird spider) | √ | √ | [73,74,75] | |
| H. schmidti (Chinese bird spider) | √ | √ | [76,77,78,79,80] | ||
| Theridiidae | Latrodectus | L. geometricus (brown widow spider) | - | √ | [81] |
| L. hesperus (Western black widow spider) | √ | √ | [81,82] | ||
| L. tredecimguttatus (Mediterranean black widow) | √ | √ | [83,84,85] | ||
| Steatoda | S. grossa (cupboard spider) | - | √ | [81] |
| Family | Genus | Species | Prot. | Tran. | Reference |
|---|---|---|---|---|---|
| Buthidae | Androctonus | A. bicolor (black fat-tailed scorpion) | √ | √ | [86] |
| A. mauretanicus mauretanicus (fat-tailed scorpion) | √ | - | [87] | ||
| Buthacus | B. macrocentrus (Turkish scorpion) | √ | - | [88] | |
| Buthus | B martensi (Chinese Scorpion) | √ | - | [89] | |
| Centruroides | C. tecomanus | √ | √ | [90] | |
| Hottentotta | H. conspersus (Sesriem scorpion) | - | √ | [91] | |
| H. judaicus (black scorpion) | - | √ | [92] | ||
| Isometrus | I. maculatus (lesser brown scorpion) | - | √ | [93] | |
| Leiurus | L. quinquestriatus hebraeus (yellow scorpion) | √ | - | [94] | |
| Leiurus | L. quinquestriatus quinquestriatus (deathstalker scorpion) | √ | - | [94] | |
| Lychas | L. mucronatus (Chinese Swimming Scorpion) | - | √ | [93] | |
| Mesobuthus | M. eupeus (lesser Asian scorpion) | √ | √ | [95] | |
| Rhoplaurus | R. junceus (Caribbean blue scorpion) | √ | - | [96,97,98] | |
| Tityus | T. bahiensis (Brazilian scorpion) | √ | √ | [94,99] | |
| T. cambridgei (Amazonian scorpion) | √ | - | [100] | ||
| T. costatus (Brazilian scorpion) | √ | - | [101] | ||
| T. discrepans (Venezuelan scorpion) | √ | - | [102] | ||
| T. pachyurus (Colombian scorpion) | √ | - | [103] | ||
| T. serrulatus (Brazilian scorpion) | √ | √ | [104,105,106,107,108,109,110,111,112] | ||
| T. stigmurus (Brazilian scorpion) | √ | - | [94,113,114] | ||
| Caraboctonidae | Hadrurus | H. gertschi | - | √ | [115] |
| Chaerilidae | Chaerilus | C. tricostatus | - | √ | [116] |
| C. tryznai | - | √ | [116] | ||
| Euscorpiidae | Scorpiops | S. jendeki | - | √ | [117] |
| S. margerisonae | - | √ | [93] | ||
| Hemiscorpiidae | Hemiscorpius | H. lepturus | √ | - | [118] |
| H. persicus | √ | - | [118] | ||
| Opisthacanthus | O. cayaporum (Brazilian scorpion) | √ | - | [119] | |
| O. elatus | √ | - | [120] | ||
| Scorpionidae | Heterometrus | H. longimanus (black emperor scorpion) | √ | - | [121] |
| H. petersii (Asian forest scorpion) | √ | √ | [122] | ||
| Pandinus | P. cavimanus (Tanzanian red clawed scorpion) | - | √ | [123] | |
| Scorpio | S. maurus palmatus (chactoid scorpion) | √ | √ | [124] | |
| Urodacus | U. yaschenkoi (inland robust scorpion) | √ | √ | [125,126,127] |
| Antitoxin | Chemical class | Molecular Formula | Structure | MW (Da) | Target | Target | Ref. |
|---|---|---|---|---|---|---|---|
| Species | Toxin Family | ||||||
| Heparin | Sulfated glyco-aminglycan | C12H19NO20S3 |  | 12–15 kDa | Buthus martensi (Chinese red scorpion), Palamneus gravimanus (Indian black scorpion), Heterometrus fulvipes (giant forrest scorpion) | Hyaluronidase | [129,130,131] |
| Aristolochic acid | Alkaloid | C17H11NO7 |  | 341.27 | Tityus serrulatus (Brazilian yellow scorpion) | Hyaluronidase PLA2 | [132] |
| EDTA | Acyclic | C10H16N2O8 |  | 292.24 | Buthus martensi (Chinese red scorpion), Heterometrus fulvipes (giant forrest scorpion), Isonietrus vittatus (scorpion), Hemiscorpius lepturus (scorpion), Tityus serrulatus (Brazilian yellow scorpion) Loxosceles intermedia (brown spider), Hippasa partita (spider), Parawixia bistriata (spider) | Metalloprotease Hyaluronidase PLA2 | [129,131,133,134,135,136,137,138,139] |
| 1,10-phenanthroline | Heterocyclic, 3-ring | C12H8N2 |  | 180.21 | Hemiscorpius lepturus (scorpion) Loxosceles intermedia (brown spider), Hippasa partita (spider) | Metalloprotease Hyaluronidase PLA2 | [133,134,138] |
| Name | Target | Type | Author | Year | Ref. |
|---|---|---|---|---|---|
| LiMAb(7) | Loxoscelesintermedia (recluse spider). | mAb | Alvarenga et al. | 2003 | [149] |
| FM1 | Alpha-latrotoxin from Latrodectus sp. (black widow spiders). | Fab | Bugli et al. | 2008 | [150] |
| LiD1mAb16 | Sphingomyelinases D from Loxosceles intermedia, L. laeta and L. gaucho (brown or recluse spiders). | mAb | Dias-Lopes et al. | 2014 | [151] |
| Name | Target | Type | Author | Year | Ref. |
|---|---|---|---|---|---|
| mAb 4C1 | Aah I from Androctonus australis (fat-tailed scorpion) | mAb | Bahraoui et al. | 1988 | [146] |
| mAb BCF2 | Cn2 from Centruroides noxius (Mexican bark scorpion) | mAb | Zamudio et al. | 1992 | [152,153] |
| Licea et al. | 1996 | ||||
| mAb 9C2 | Aah II from Androctonus australis (fat-tailed scorpion) | mAb | Clot-Faybesse et al. | 1999 | [148] |
| scFv 4C1 | Aah I from Androctonus australis (fat-tailed scorpion) | scFv | Mousli et al. | 1999 | [154] |
| mAbs BmK AS-1 | BmK AS-1 from Buthus martensii karsch (Chinese scorpion) | mAb | Jia et al. | 2000 | [155] |
| scFv 9C2 | Aah II from Androctonus australis (fat-tailed scorpion) | scFv | Devaux et al. | 2001 | [156] |
| chFab-BCF2 | Cn2 from Centruroides noxius (Mexican bark scorpion) | chFab | Selisko et al. | 2004 | [157] |
| rFab 9C2 | Aah I from Androctonus australis (fat-tailed scorpion) | rFab | Aubrey et al. | 2004 | [158] |
| Triple mutant (G5 + B7) | Cn2 from Centruroides noxius hoffmann (Mexican bark scorpion) | scFvs | Juárez-González et al. | 2005 | [159] |
| mAbTs1 | TsVII, TsIV and TsNTxP from Tityus serrulatus (brazilian yellow scorpion) | mAb | Alvarenga et al. | 2005 | [160] |
| T94H6 | Aah I and Aah II from Androctonus australis (fat-tailed scorpion) | Tandem-scFv | Juste et al. | 2007 | [161] |
| Name | Target | Type | Author | Year | Ref. |
|---|---|---|---|---|---|
| scFv 3F, scFv 6009F | Cn2 from Centruroides noxiushoffmann (Mexican bark scorpion) | Human scFv | Riaño-Umbarila et al. | 2005 | [162] |
| NbAahI’22 | AahI’ from Androctonus australis (Sahara scorpion) | Camelid Nb | Hmila et al. | 2008 | [163] |
| NbAahII10 | AahII from Androctonus australis hector (Sahara scorpion) | Camelid Nb | Abderrazek et al. | 2009 | [164] |
| NbAah’F12 | AahI from Androctonus australis (Sahara scorpion) | Camelid Nb | Hmila et al | 2010 | [165] |
| scFv 9004G | Cn2 from Centruroides noxius hoffmann and Css2 from Centruroides suffusus suffusus (Mexican bark scorpions) | Human scFv | Riaño-Umbarila et al. | 2011 | [166] |
| scFv LR | - | - | - | ||
| scFv 15e | Ts1 or gamma-toxin from Tityus serrulatus (Brazilian yellow scorpion) | Human scFv | Amaro et al. | 2011 | [167] |
| NbF12-10 | AahI and AahII from Androctonus australis (Sahara scorpion) | Bispecific Nb | Hmila et al. | 2012 | [168] |
| Db 9C2 + Db4 C1 | AahI and AahII from Androctonus australis (Sahara scorpion) | Diabody mixture | Di Tommaso et al. | 2012 | [169] |
| Diabody D4, scFv LER | Cn2 from Centruroides noxiushoffmann (Mexican bark scorpion) | Diabody | Rodríguez-Rodríguez et al. | 2012 | [170] |
| Serrumab | Ts1 and Ts2 from Tityus serrulatus (Brazilian yellow scorpion) | Human scFv | Pucca et al. | 2012, 2014 | [171,172] |
| scFv C1 | Cn2 from Centruroides noxius hoffmann (Mexican bark scorpion) | Human scFv | Riaño-Umbarila et al. | 2013 | [173] |
| scFv 202F | CII1 from Centruroides limpidus limpidus (Mexican bark scorpion) | - | - | - | |
| scFv RU1 | Cn2 from Centruroides noxius hoffmann (Mexican bark scorpion), and CII1 from Centruroides limpidus limpidus (Mexican bark scorpion) | Human scFv | Riaño-Umbarila et al. | 2016 | [174] |
| scFv ER-5 | Cn2 from Centruroides noxius Hoffmann (Mexican bark scorpion), CII1 from Centruroides limpidus limpidus (Mexican bark scorpion), and Css2 from Centruroides suffuses suffuses (Mexican bark scorpion) | Human scFv | Rodríguez-Rodrígues et al. | 2016 | [45] |
| Immunization Strategy | Target | Author | Year | Ref. |
|---|---|---|---|---|
| Recombinant toxin | A dermonecrotic toxin from Loxoscelesintermedia (recluse spider) | Araujo et al. | 2003 | [186] |
| Synthetic epitope | A dermonecrotic toxin from Loxoscelesintermedia (recluse spider) | Felicori et al. | 2009 | [187] |
| Synthetic toxin | Robustoxin from Atrax robustus (Sydney funnel-web spider) | Comis et al. | 2009 | [188] |
| Recombinant toxin | A dermonecrotic toxin from Loxoscelesintermedia (recluse spider) | Mendes et al. | 2013 | [189] |
| Immunization Strategy | Target | Author | Year | Ref. |
|---|---|---|---|---|
| Synthetic epitope | AaH2 from Androctonus australis (Sahara scorpion) | Bahroui et al. | 1986 | [163] |
| Synthetic epitope | Cn2 from Centruroides noxius (Mexican bark scorpion) | Calderon-Aranda et al. | 1995 | [191] |
| Recombinant toxin | BotXIV from Buthus occitanus tunetanus (common European scorpion) | Bouhaouala-Zahar et al. | 1996 | [192] |
| Synthetic epitope | AaH2 from Androctonus australis hector (Sahara scorpion) | Devaux et al. | 1997 | [193] |
| Synthetic epitope | Cn2 from Centruroides noxius Hoffmann (Mexican bark scorpion) | Calderon-Aranda et al. | 1999 | [194] |
| Recombinant toxin | TsNTxP from Tityus serrulatus (Brazilian yellow scorpion) | Guatimosim et al. | 2000 | [195] |
| Recombinant toxin | AaH1, AaH2 and AaH3 from Androctonus australis (Sahara scorpion) | Legros et al. | 2001 | [196] |
| Recombinant toxin | Bot III from Buthus occitanus tunetanus (common European scorpion) | Benkhadir et al. | 2001 | [197] |
| Synthetic epitope | TsNTxP and TsIV from Tityus serrulatus (Brazilian yellow scorpion) | Alvarenga et al. | 2002 | [198] |
| Recombinant toxin | Cn5 from Centruroides noxius Hoffmann (Mexican bark scorpion) | Garcia et al. | 2003 | [199] |
| Synthetic epitope | Birtoxin from Parabuthus transvaalicus (South African fat-tail Scorpion) | Inceoglu et al. | 2006 | [200] |
| Recombinant toxin | Ts1 from Tityus serrulatus (Brazilian yellow scorpion) | Mendes et al. | 2008 | [201] |
| Recombinant toxin | PG8 from Parabuthus granulatus (granulated thick-tailed scorpion) | García-Gómez et al. | 2009 | [202] |
| Recombinant toxin | Css2 from Centruroides suffusus suffusus (Mexican bark scorpion) | Hernández-Salgado et al. | 2009 | [203] |
| Synthetic epitope | TsNTxP from Tityus serrulatus (Brazilian yellow scorpion) | Duarte et al. | 2010 | [204] |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Laustsen, A.H.; Solà, M.; Jappe, E.C.; Oscoz, S.; Lauridsen, L.P.; Engmark, M. Biotechnological Trends in Spider and Scorpion Antivenom Development. Toxins 2016, 8, 226. https://doi.org/10.3390/toxins8080226
Laustsen AH, Solà M, Jappe EC, Oscoz S, Lauridsen LP, Engmark M. Biotechnological Trends in Spider and Scorpion Antivenom Development. Toxins. 2016; 8(8):226. https://doi.org/10.3390/toxins8080226
Chicago/Turabian StyleLaustsen, Andreas Hougaard, Mireia Solà, Emma Christine Jappe, Saioa Oscoz, Line Præst Lauridsen, and Mikael Engmark. 2016. "Biotechnological Trends in Spider and Scorpion Antivenom Development" Toxins 8, no. 8: 226. https://doi.org/10.3390/toxins8080226