
A Tricky Trait: Applying the Fruits of the “Function Debate” in the Philosophy of Biology to the “Venom Debate” in the Science of Toxinology


Abstract
:1. Introduction
1.1. Function, “Why” Questions and Teleonomic Explanations
1.2. Theories of Function
1.3. Venom Is a Complex Functional Trait
2. Theories of Function Applied to Venom
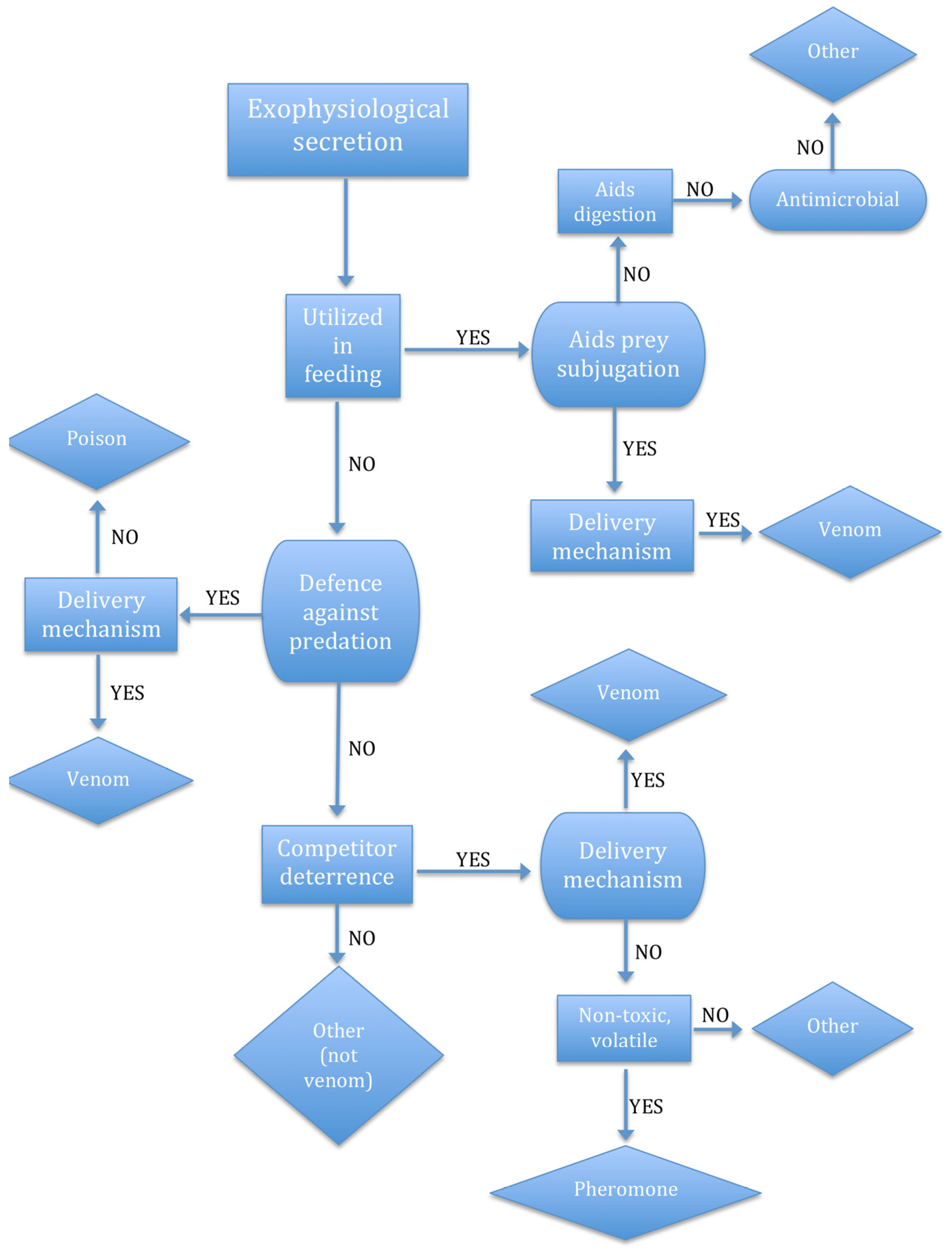
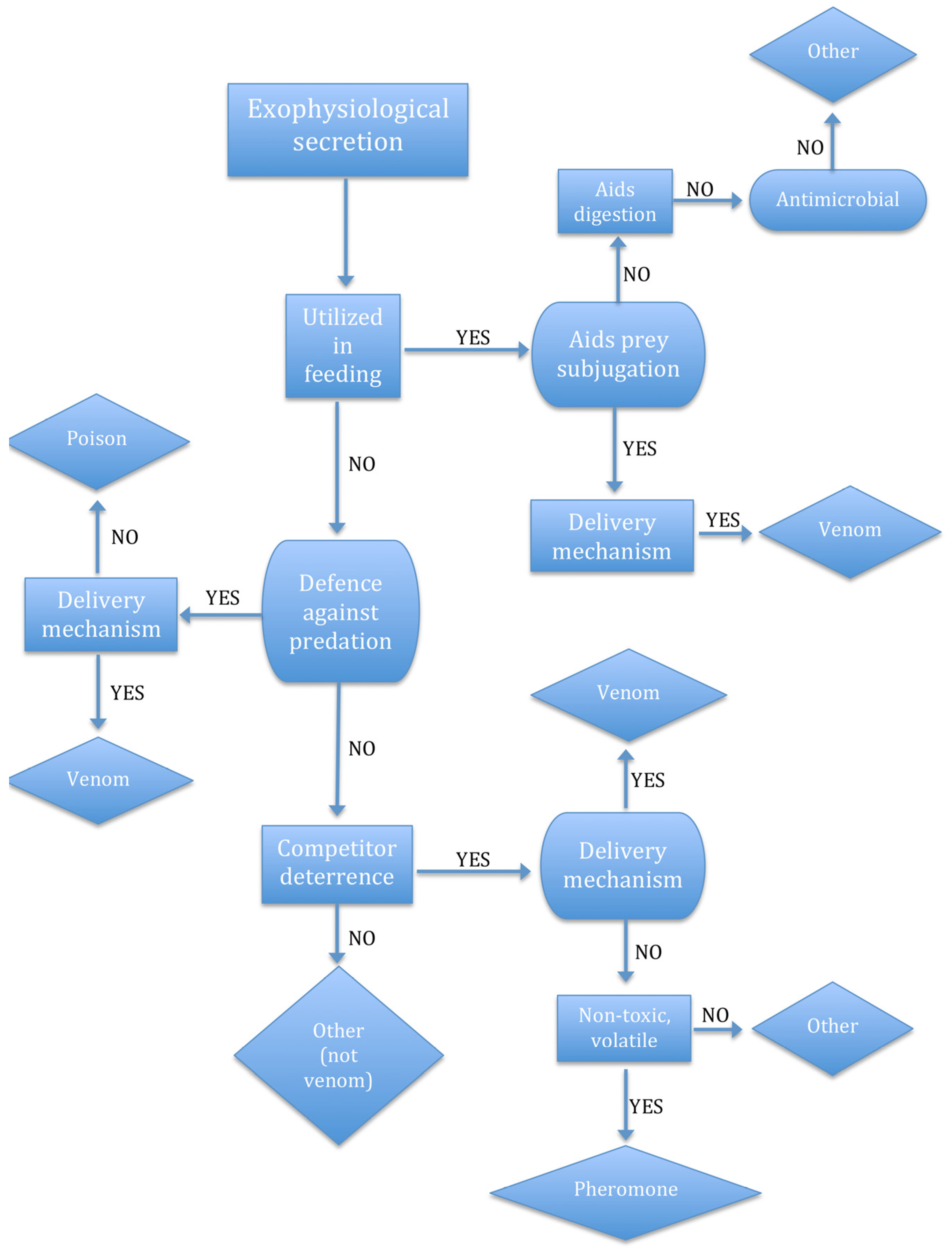
2.1. The Venom System
2.2. Death as an Epiphenomenal Consequence of Envenomation
2.3. Use It (for Predation) or Lose It
2.4. The Context-Specificity of Functions
2.5. Defective Functions—Vestigial and Incipient Traits
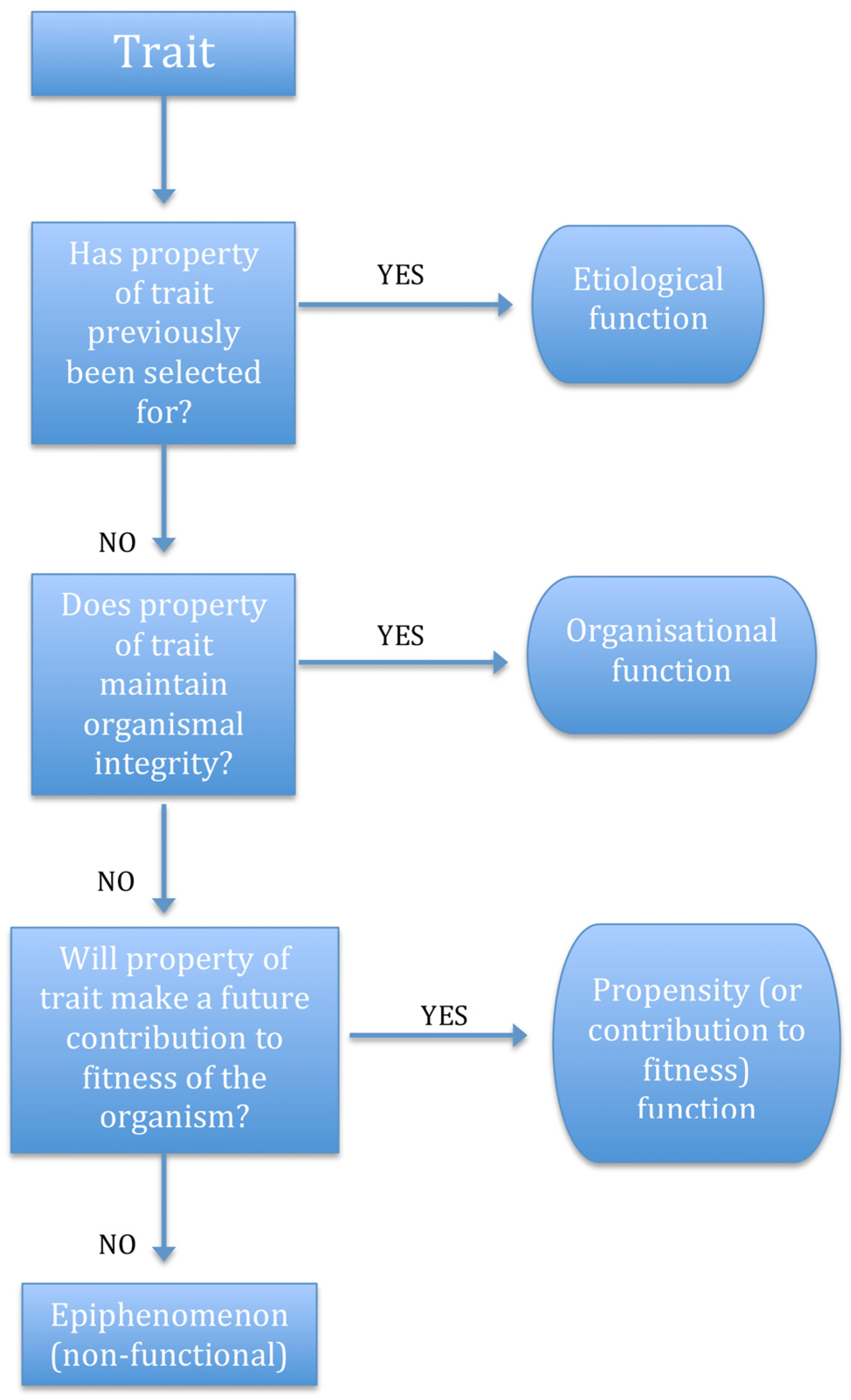
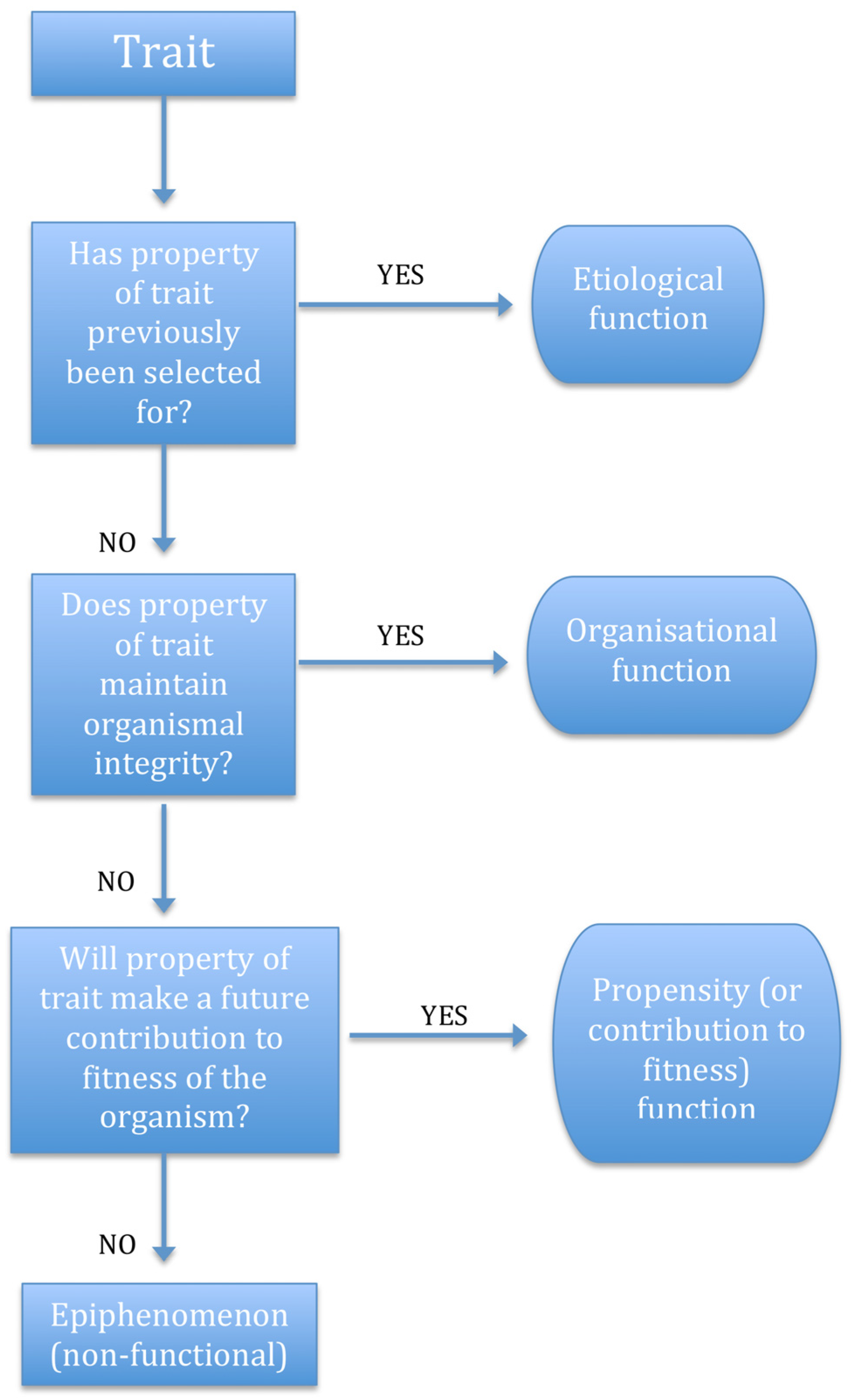
2.6. Function Categories
2.7. Vestigial and Incipient Traits Revisited
3. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Hohwy, J.; Kallestrup, J. Being Reduced: New Essays on Reduction, Explanation, and Causation; Oxford University Press: Oxford, UK, 2008. [Google Scholar]
- Ayala, F.J.; Arp, R. Contemporary Debates in Philosophy of Biology; Wiley-Blackwell Pub: Chichester, UK, 2010. [Google Scholar]
- Dennett, D.C. Darwin’s Dangerous Idea: Evolution and the Meanings of Life; Simon & Schuster: New York, NY, USA, 1995. [Google Scholar]
- Wouters, A.G. The Function Debate in Philosophy. Acta Biotheor. 2005, 53, 123–151. [Google Scholar] [CrossRef] [PubMed]
- Gilbert, W.H. Teleological vs teleonomic. Nature 1975, 254, 176–176. [Google Scholar] [CrossRef]
- Jelliffe, D.B. Teleonomic Explanation. BioScience 1978, 28, 681–681. [Google Scholar] [CrossRef]
- Saborido, C. New Directions in the Philosophy of Biology: A New Taxonomy of Functions. In New Directions in the Philosophy of Science; Galavotti, C.M., Dieks, D., Gonzalez, J.W., Hartmann, S., Uebel, T., Weber, M., Eds.; Springer International Publishing: Cham, Switzerland, 2014; pp. 235–251. [Google Scholar]
- Ariew, A.; Cummins, R.; Perlman, M. Functions: New Essays in the Philosophy of Psychology and Biology; Oxford University Press: Oxford, UK, 2002. [Google Scholar]
- Walsh, D.M. Fitness and Function. Br. J. Philos. Sci. 1996, 47, 553–574. [Google Scholar] [CrossRef]
- Gould, S.J. The Panda’s Thumb: More Reflections in Natural History; Norton: New York, NY, USA, 1980. [Google Scholar]
- Godfrey-Smith, P. A Modern History Theory of Functions. Noûs 1994, 28, 344–362. [Google Scholar] [CrossRef]
- Gould, S.J.; Vrba, E.S. Exaptation—A missing term in the science of form. Paleobiology 1982, 8, 4–15. [Google Scholar] [CrossRef]
- Mossio, M.; Saborido, C.; Moreno, A. An Organizational Account of Biological Functions. Br. J. Philos. Sci. 2009, 60, 813–841. [Google Scholar] [CrossRef] [Green Version]
- Griffiths, P.E. Functional Analysis and Proper Functions. Br. J. Philos. Sci. 1993, 44, 409–422. [Google Scholar] [CrossRef]
- Godfrey-Smith, P. Functions: Consensus without unity. Pac. Philos. Q. 1993, 74, 196–208. [Google Scholar]
- Kardong, K.V. Replies to Fry et al. (Toxicon 2012, 60/4, 434–448). Part B. Properties and biological roles of squamate oral products: The “venomous lifestyle” and preadaptation. Toxicon 2012, 60. [Google Scholar] [CrossRef] [PubMed]
- Weinstein, S.A. “Venomous” Bites from Non-Venomous Snakes: A Critical Analysis of Risk and Management of “Colubrid” Snake Bites; Elsevier: London, UK, 2011. [Google Scholar]
- Weinstein, S.A.; Keyler, D.E.; White, J. Replies to Fry et al. (Toxicon 2012, 60/4, 434–448). Part A. Analyses of squamate reptile oral glands and their products: A call for caution in formal assignment of terminology designating biological function. Toxicon 2012, 60. [Google Scholar] [CrossRef] [PubMed]
- Fry, B.G.; Casewell, N.R.; Wüster, W.; Vidal, N.; Young, B.; Jackson, T.N.W. The structural and functional diversification of the Toxicofera reptile venom system. Toxicon 2012, 60, 434–448. [Google Scholar] [CrossRef] [PubMed]
- Weinstein, S.A. Snake venoms: A brief treatise on etymology, origins of terminology, and definitions. Toxicon 2015, 103, 188–195. [Google Scholar] [CrossRef] [PubMed]
- Olivera, B.M.; Gray, W.R.; Zeikus, R.; McIntosh, J.M.; Varga, J.; Rivier, J.; de Santos, V.; Cruz, L.J. Peptide neurotoxins from fish-hunting cone snails. Science 1985, 230, 1338–1343. [Google Scholar] [CrossRef] [PubMed]
- Quicke, D.L.J. Parasitic Wasps; Chapman & Hall: London, UK, 1997. [Google Scholar]
- Dawkins, R. The Selfish Gene; Oxford University Press: Oxford, UK, 2006. [Google Scholar]
- Doolittle, W.F.; Sapienza, C. Selfish genes, the phenotype paradigm and genome evolution. Nature 1980, 284, 601–603. [Google Scholar] [CrossRef] [PubMed]
- Ruxton, G.D.; Sherratt, T.N.; Speed, M.P. Avoiding Attack: The Evolutionary Ecology of Crypsis, Warning Signals, and Mimicry; Oxford University Press: Oxford, UK, 2004. [Google Scholar]
- Casewell, N.R.; Wüster, W.; Vonk, F.J.; Harrison, R.A.; Fry, B.G. Complex cocktails: The evolutionary novelty of venoms. Trends Ecol. Evol. 2013, 28, 219–229. [Google Scholar] [CrossRef] [PubMed]
- Exnerová, A.; Stys, P.; Fucíková, E.; Veselá, S.; Svádová, K.; Prokopová, M.; Jarosík, V.; Fuchs, R.; Landová, E. Avoidance of aposematic prey in European tits (Paridae): Learned or innate? Behav. Ecol. 2006, 18, 148–156. [Google Scholar] [CrossRef]
- Shine, R.; Bonnet, X.; Elphick, M.J.; Barrott, E.G. A Novel Foraging Mode in Snakes: Browsing by the Sea Snake Emydocephalus annulatus (Serpentes, Hydrophiidae). Funct. Ecol. 2004, 18, 16–24. [Google Scholar] [CrossRef]
- Li, M.; Fry, B.G.; Kini, R.M. Eggs-Only Diet: Its Implications for the Toxin Profile Changes and Ecology of the Marbled Sea Snake (Aipysurus eydouxii). J. Mol. Evol. 2005, 60, 81–89. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Fry, B.G.; Kini, R.M. Putting the brakes on snake venom evolution: The unique molecular evolutionary patterns of Aipysurus eydouxii (Marbled sea snake) phospholipase A2 toxins. Mol. Biol. Evol. 2005, 22, 934–941. [Google Scholar] [CrossRef] [PubMed]
- Tu, A.T.; Fulde, G. 17 Sea snake bites. Clin. Dermatol. 1987, 5, 118–126. [Google Scholar] [CrossRef]
- Pawlak, J.; Mackessy, S.P.; Sixberry, N.M.; Stura, E.A.; Le Du, M.H.; Ménez, R.; Foo, C.S.; Ménez, A.; Nirthanan, S.; Kini, R.M. Irditoxin, a novel covalently linked heterodimeric three-finger toxin with high taxon-specific neurotoxicity. FASEB J. 2009, 23, 534–545. [Google Scholar] [CrossRef] [PubMed]
- Rochelle, M.J.; Kardong, K.V. Constriction versus Envenomation in Prey Capture by the Brown Tree Snake, Boiga irregularis (Squamata: Colubridae). Herpetologica 1993, 49, 301–304. [Google Scholar]
- Millikan, R.G. In Defense of Proper Functions. Philos. Sci. 1989, 56, 288–302. [Google Scholar] [CrossRef]
- Fry, B.G.; Undheim, E.A.B.; Ali, S.A.; Jackson, T.N.W.; Debono, J.; Scheib, H.; Ruder, T.; Morgenstern, D.; Cadwallader, L.; Whitehead, D.; et al. Squeezers and leaf-cutters: Differential diversification and degeneration of the venom system in toxicoferan reptiles. Mol. Cell. Proteom. MCP 2013, 12, 1881–1899. [Google Scholar] [CrossRef] [PubMed]
- Kochva, E. The origin of snakes and evolution of the venom apparatus. Toxicon 1987, 25, 65–106. [Google Scholar] [CrossRef]
- Kelly, C.M.R.; Barker, N.P.; Villet, M.H.; Broadley, D.G. Phylogeny, biogeography and classification of the snake superfamily Elapoidea: A rapid radiation in the late Eocene. Cladistics 2009, 25, 38–63. [Google Scholar] [CrossRef]
- McCarthy, C.J. Monophyly of elapid snakes (Serpentes: Elapidae). An assessment of the evidence. Zool. J. Linn. Soc. 1985, 83, 79–93. [Google Scholar] [CrossRef]
- McDowell, S.B. The Architecture of the Corner of the Mouth of Colubroid Snakes. J. Herpetol. 1986, 20, 353–407. [Google Scholar] [CrossRef]
- Randal Bollinger, R.; Barbas, A.S.; Bush, E.L.; Lin, S.S.; Parker, W. Biofilms in the large bowel suggest an apparent function of the human vermiform appendix. J. Theor. Biol. 2007, 249, 826–831. [Google Scholar] [CrossRef] [PubMed]
- Shivik, J.A. Are Vultures Birds, and Do Snakes Have Venom, because of Macro- and Microscavenger Conflict? BioScience 2006, 56, 819–823. [Google Scholar] [CrossRef]
- Fry, B.G.; Vidal, N.; Norman, J.A.; Vonk, F.J.; Scheib, H.; Ramjan, S.F.; Kuruppu, S.; Fung, K.; Hedges, S.B.; Richardson, M.K.; et al. Early evolution of the venom system in lizards and snakes. Nature 2006, 439, 584–588. [Google Scholar] [CrossRef] [PubMed]
- Plato; Rowe, C.J. Phaedrus; Penguin: London, UK, 2005. [Google Scholar]
- Slater, M.H.; O’Rourke, M.; Campbell, J.K. Carving Nature at Its Joints: Natural Kinds in Metaphysics and Science; MIT Press: Cambridge, UK, 2011. [Google Scholar]
- Machery, E. Précis of Doing without Concepts. Behav. Brain Sci. 2010, 33, 195–206. [Google Scholar] [CrossRef] [PubMed]
- Richards, R.A. The Species Problem: A Philosophical Analysis; Cambridge University Press: Cambridge, UK, 2010. [Google Scholar]
- Dennett, D.C. Kinds of Minds: Toward an Understanding of Consciousness; Basic Books: New York, NY, USA, 1996. [Google Scholar]

{kind=link}
{kind=link}
| Theory | Defines “Function” as | Potential Limitations |
|---|---|---|
| Causal role | Effect of trait that contributes/facilitates/is integral to achievement of organismal ‘goals’. | Potential failure to distinguish between adaptations and epiphenomena. |
| Etiological | Effect of trait selected for during the evolutionary past that explains its current presence. | Inability to account for de novo appearance of functions. |
| Propensity | Effect of trait that makes contribution to the future fitness of the organism that possesses it. | Failure to consider current disposition of trait. |
| Organisational | Effect of trait that contributes to maintaining the integrity of the system (organism) that possesses it. | Conflict between maintenance of organism and maximisation of organism’s fitness. |
| Contribution to fitness | Effect of trait that makes a contribution to the fitness of the organism that possesses it. | Must include considerations of the trait’s evolutionary history as well as its current disposition |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jackson, T.N.W.; Fry, B.G. A Tricky Trait: Applying the Fruits of the “Function Debate” in the Philosophy of Biology to the “Venom Debate” in the Science of Toxinology. Toxins 2016, 8, 263. https://doi.org/10.3390/toxins8090263
Jackson TNW, Fry BG. A Tricky Trait: Applying the Fruits of the “Function Debate” in the Philosophy of Biology to the “Venom Debate” in the Science of Toxinology. Toxins. 2016; 8(9):263. https://doi.org/10.3390/toxins8090263
Chicago/Turabian StyleJackson, Timothy N. W., and Bryan G. Fry. 2016. "A Tricky Trait: Applying the Fruits of the “Function Debate” in the Philosophy of Biology to the “Venom Debate” in the Science of Toxinology" Toxins 8, no. 9: 263. https://doi.org/10.3390/toxins8090263