Microbial Inhibition of Fusarium Pathogens and Biological Modification of Trichothecenes in Cereal Grains


1
Department of Entomology, Phytopathology and Molecular Diagnostics, University of Warmia and Mazury in Olsztyn, ul. Prawocheńskiego 17, 10-720 Olsztyn, Poland
2
Department of Plant Breeding and Seed Production, University of Warmia and Mazury in Olsztyn, pl. Łódzki 3, 10-724 Olsztyn, Poland
*
Author to whom correspondence should be addressed.
Toxins 2017, 9(12), 408; https://doi.org/10.3390/toxins9120408
Submission received: 1 November 2017
/
Revised: 15 December 2017
/
Accepted: 18 December 2017
/
Published: 20 December 2017
(This article belongs to the Collection Understanding Mycotoxin Occurrence in Food and Feed Chains)
Abstract
:Fungi of the genus Fusarium infect cereal crops during the growing season and cause head blight and other diseases. Their toxic secondary metabolites (mycotoxins) contaminate grains. Several dozen toxic compounds produced by fungal pathogens have been identified to date. Type B trichothecenes—deoxynivalenol, its acetyl derivatives and nivalenol (produced mainly by F. graminearum and F. culmorum)—are most commonly detected in cereal grains. “T-2 toxin” (produced by, among others, F. sporotrichioides) belongs to type-A trichothecenes which are more toxic than other trichothecenes. Antagonistic bacteria and fungi can affect pathogens of the genus Fusarium via different modes of action: direct (mycoparasitism or hyperparasitism), mixed-path (antibiotic secretion, production of lytic enzymes) and indirect (induction of host defense responses). Microbial modification of trichothecenes involves acetylation, deacetylation, oxidation, de-epoxidation, and epimerization, and it lowers the pathogenic potential of fungi of the genus Fusarium. Other modifing mechanisms described in the paper involve the physical adsorption of mycotoxins in bacterial cells and the conjugation of mycotoxins to glucose and other compounds in plant and fungal cells. The development of several patents supports the commercialization and wider application of microorganisms biodegrading mycotoxins in grains and, consequently, in feed additives.
1. Introduction
In temperate climates, pathogens of the genus Fusarium are the most dangerous producers of toxic metabolites in cereal grains. They develop during the growing season on spikes or panicles to cause Fusarium head blight (FHB). A number of secondary metabolites produced by those fungi have an adverse influence on mammals: they cause damage to internal organs and lead to poisoning in humans and animals [1]. Fungi of the genus Fusarium are not always effectively controlled with fungicides because FHB epidemics develop rapidly and involve at least several fungal species [2]. New strategies in crop protection aim to limit the number of fungicide treatments, and they have led to the introduction of integrated pest management requirements in the relevant legal acts and regulations [3] which favor biological and biotechnological approaches. Statutory regulations significantly limit the use of chemical substances in organic farms [4]. The mycotoxin content of grains intended for consumption should not exceed the levels determined to be safe by toxicological considerations [5,6]. In unprocessed soft wheat grains, the maximum concentrations of deoxynivalenol (DON) are set at 1250 µg kg−1, in durum and oat grains at 1750 µg kg−1 [5], and the total content of T-2 and HT-2 toxins (indicative value) may not exceed 100 µg kg−1 [7]. Due to food safety concerns, mycotoxin levels in food and feed have to be monitored regularly to evaluate the risk of exceeding the tolerable daily intake (TDI) in humans and animals, expressed in mg kg−1 body weight (bw) [8]. Food and feed producers are, thus, forced to search for new methods of inhibiting the growth and development of toxin-producing fungi of the genus Fusarium.
Genetic analyses investigating the interactions between the genes of toxin-producing fungi, bacteria, antagonistic fungi and host plants supply valuable data for designing new biotechnological methods for eliminating trichothecenes from plant tissue. Okubara et al. [9] described a wheat line transformed by the TRI 101 gene from F. sporotrichioides. TRI101 encodes an enzyme that transfers an acetyl moiety to the C3 hydroxyl group of trichothecenes. Acetyltransferase in this plant confers partial protection against the spread of F. graminearum in inoculated wheat heads. Ohsato et al. [10] obtained resistant rice genotypes showing DON acetylase activity. Genetic transformation can significantly increase cereal resistance to Fusarium fungi [10]. However, this method is rarely used, and it is subject to numerous legal limitations in Europe. Genetic transformation complements the natural defense mechanism of cereals, which involves the conjugation of DON to glucose to produce (deoxynivalenol-3-β-d-glucopyranoside) D3G [11,12]. This mechanism probably differentiates the susceptibility of different wheat species to F. culmorum infections [13]. The presence of DON-glutathione conjugates was reported in wheat [14]. The cited authors also observed other conjugates, including DON-cysteine, as well as unknown conjugates formed in plants.
The microbiome of cereal grains, which is composed of various species of yeasts, bacteria, and filamentous fungi, is found on grain surfaces and in grain tissues, and it is much more abundant than toxin-producing fungi of the genus Fusarium [15,16]. The microbiome’s role in protecting plants against pathogens is intensively studied [15,17,18,19]. The microbiome can prevent the growth of Fusarium pathogens by antibiosis and competition during the growing season [20,21] and grain storage [22]. It also enhances host defense against pathogens [23,24]. Numerous studies have demonstrated that microorganisms can biotransform or biodegrade mycotoxins to compounds that are non-toxic or less toxic [25,26]. The aim of this review article was to summarize the existing knowledge regarding the use of selected microorganisms for inhibiting the development of Fusarium pathogens and lowering trichothecene concentrations in cereal grains and cereal-based feed.
2. Trichothecenes
Trichothecenes are one of the major groups of secondary metabolites produced by species of the genus Fusarium and covering more than 200 compounds [27,28]. The chemical structure and the possible biosynthesis pathways of trichothecenes are relatively well known [28]. Several questions regarding the toxicity mechanisms of trichothecenes remain unanswered, but some of these mechanisms have been identified in eukaryotes [29]. Trichothecenes are cyclic terpenoids which can be further subdivided into simple and macrocyclic forms. Simple trichothecenes are a family of tetracyclic or tricyclic sesquiterpenes with various substituents [28]. FHB is caused mainly by F. graminearum and F. culmorum which produce type B trichothecenes—DON with its acetylated derivatives, 3-acetyl-DON (3-Ac-DON) and 15-acetyl-DON (15-Ac-DON), and nivalenol (NIV). Fungi that are less frequently noted on cereal spikes include F. sporotrichioides and F. poae which produce type-A trichothecenes-T-2 toxin and HT-2 toxin [30].
Type-B trichothecenes suppress appetite, induce vomiting and weaken the immune system [27,31]. T-2 toxin and other type-A trichothecenes damage the skin and mucous membranes, promote vomiting and diarrhea, suppress appetite, lead to hemorrhaging and neurological disorders [32]. Trichothecenes often disrupt life processes in mammalian cells. These metabolites cause various human and animal diseases classified as mycotoxicoses. Goossens et al. [33] demonstrated that the intake of feed contaminated with 903 ± 85 μg kg−1 of T-2 toxin inhibited the activity of porcine liver enzymes within 14 days. At the cellular level, trichothecenes disrupt nucleic acid synthesis, mitochondrial function, membrane integrity, and cell division. Trichothecenes may also induce apoptosis in animal cells [29]. DON and T-2 toxin bind to the peptidyl transferase center of the ribosome and block protein synthesis [34].
The biosynthesis of trichothecenes begins with the cyclization of farnesyl pyrophosphate (FPP) to form trichodiene, which then undergoes a series of oxygenation, isomerization, cyclization, and esterification reactions to form T-2 toxin, NIV, or DON [35,36,37]. The transformation pathway was described in a previous study [38]. Proteins that participate in the biosynthesis of trichothecenes are encoded by TRI1-TRI16 genes grouped in three clusters. F. graminearum sensu stricto and F. sporotrichioides were the first model species where the main TRI cluster was characterized [39,40,41]. In both species, the main TRI cluster is composed of 12 genes responsible for the synthesis and modification of the trichothecene skeleton. The cluster can include seven genes (TRI8, TRI7, TRI3, TRI4, TRI5, TRI11, TRI13) encoding enzymes that catalyze 10 trichothecene biosynthetic reactions, two genes (TRI6 and TRI10) encoding transcriptional regulators, one gene (TRI12) encoding a transport protein, and two genes (TRI9 and TRI14) with uncertain functions. Both species have two additional TRI loci: the single gene TRI101 locus encoding C-3 acetyltransferase and a locus containing two genes—TRI1 encoding C-8 oxygenase and TRI16 encoding C-8 acetyltransferase. An analysis of the fungal genome revealed that those loci are positioned on different chromosomes [30,42]. The presence of the TRI5 gene, which encodes the enzyme participating in the isomerization and cyclization of farnesyl pyrophosphate to trichodiene in Fusarium isolates, determines their ability to produce trichothecenes. The chemotypes of Fusarium isolates can be determined by analyzing the polymorphism of TRI3, TRI7, TRI12, and TRI13 genes in the main TRI cluster and TRI1 in additional TRI loci containing genes responsible for the synthesis of various types of trichothecenes. For example, the development of the NIV chemotype of F. graminearum requires the expression of TRI7 and TRI13 genes, whereas their absence leads to the development of the DON chemotype [41,43]. TRI1 was verified to be responsible for the difference in hydroxylation at C-8 and allows to distinguish DON chemotypes from chemotypes defined as NX. The NX chemotypes due to the lack of a ketone group at C-8 are classified as type-A trichothecenes [44].
3. Inhibition of the Growth of Fusarium Pathogens and Trichothecene Production by Microorganisms
The above goal can be achieved through biological treatments where microorganisms are applied to plants during the growing season. Microorganisms adapt well to the surface of wheat spikes, they reduce symptoms of FHB and DON levels in kernels. Schisler et al. [20] demonstrated that the biocontrol agent Cryptococcus flavescens OH 182.9 delivers protective effects on wheat spikes (Table 1). DON can inhibit the activity of N-acetyl-beta-d-glucosaminidase, an enzyme which is produced by the antagonistic species Trichoderma atroviride and is capable of degrading the cell walls of pathogens [45]. Therefore, antagonistic microorganisms tolerant of the secondary metabolites produced by Fusarium pathogens have to be identified. In a study by Pan et al. [15] isolates of endophytic Bacillus megaterium (BM1) and Bacillus subtilis (BS43, BSM0, BSM2) bacteria inhibited the growth and spore formation in F. graminearum, thus decreasing DON levels in field-grown wheat by up to 89.3%. In an earlier study, Dunlap et al. [46] demonstrated that the above bacterial species produced fungicidal or fungistatic metabolites: surfactin, iturin and fengycin lipopeptides. Palazzini et al. [47] reported that antagonistic bacterium Bacillus velezensis RC 218 could significantly reduce FHB severity and DON accumulation under field conditions. Zhao et al. [48] demonstrated that a cell-free Lactobacillus rhamnosus culture supernatant can effectively inhibit in vitro growth of F. culmorum. Baffoni et al. [49] found that Lactobacillus plantarum isolated from silages and Bacillus amyloliquefaciens isolated from forest soil inhibited the growth of F. culmorum and F. graminearum in vitro. The cited authors also demonstrated that weekly biological treatments (applied from heading until anthesis) involving a mixture of two isolates of the above species were more effective in reducing the symptoms of FHB in T. durum than two treatments applied in the flowering stage.
Antagonistic bacteria and fungi can affect pathogens of the genus Fusarium via direct, mixed-path and indirect modes of action (Figure 1). Direct interactions include mycoparasitism and hyperparasitism (Table 1). They have been reported by Vujanovic and Goh [57] in a study evaluating the effectiveness of Sphaerodes mycoparasitica filamentous fungi against F. graminearum. The cited authors hypothesized that S. mycoparasitica absorbs aurofusarin from the attacked Fusarium cells by lysing the components of pathogenic cell membranes, such as chitin, and producing chitinase and chitosanase. Ferre and Santamarina [60] demonstrated that Trichoderma harzianum produces antifungal extracellular metabolites, including enzymes that degrade the wall of F. culmorum cells: chitinases, glucanases and proteases. The filamentous fungi Clonostachys rosea analyzed by Mamarabadi et al. [59] also produced enzymes that degrade cell membranes in F. culmorum.
Mixed-path antagonism involves antibiotic secretion and the production of volatile antagonistic compounds. Chan et al. [50] demonstrated that culture filtrates of B. subtilis effectively reduced the severity of FHB (F. graminearum) in wheat and lowered DON levels in grains by up to 51% relative to control. Culture filtrates contained metabolites of the analyzed bacterium. They were identified as fengycin-like isoforms which are members of the family of membrane-active lipopeptide antibiotics against filamentous fungi. Shi et al. [51] analyzed the effectiveness of B. amyloliquefaciens in reducing DON concentrations in wheat grains. The experiment was conducted in a controlled environment, and the growth of F. graminearum mycelia in wheat kernels was observed in flasks containing bacterial cell-free culture supernatant. In the work of Nagaraja et al. [52], zones of inhibition were observed on Petri plates where Azotobacter nigricans was cultured in the presence of several Fusarium species. According to the authors, the analyzed bacterial species can be effectively used to protect wheat, maize and sorghum against toxin-producing fungi of the genus Fusarium. The tested strain of A. nigricans also stimulated root and shoot growth in the analyzed plants. Bacteria of the genera Bacillus, Pseudomonas and Microbacterium from wheat and barley rhizosphere [53], as well as B. cereus isolated from wheat [54] produced volatile antagonistic compounds that inhibited the development of F. culmorum and F. graminearum pathogens. In a study by Zou et al. [61], bacteria produced several volatile substances with antifungal activity, including acetamide, benzaldehyde, benzothiazole, 1-butanamine, methanamine, and phenylacetaldehyde.
Indirect interactions involve competition, namely the intrinsic ability to utilize the nutrients released by the host plant, as well as mechanisms responsible for the induction of host defense responses. Many species of rhizosphere or phyllosphere yeasts compete with Fusarium fungi for micronutrients and macronutrients [56]. Siderophores, the iron-chelating compounds with a high affinity for ferric iron, facilitate competition for Fe2+ ions in environments with neutral pH where the availability of Fe2+ ions is limited [62]. The A. pullulans HN6.2 isolate produced only fusigen, a hydroxamate siderophore [62], whereas B. subtilis CAS15 isolate produced the catechol siderophore 2,3-dihydroxybenzoate-glycine-threonine trimeric ester bacillibactin [63].
The use of microorganisms to activate immune responses to Fusarium pathogens in cereals has been rarely described in the literature. Petti et al. [24] demonstrated that the non-pathogenic bacterial strain of Pseudomonas fluorescens MKB158 elicited host defense responses and protected barley against F. culmorum. The bacterium significantly influenced the accumulation of 1203 transcripts, and primed 74 positive and 14 negative responses to the pathogen. Barley plants inoculated with P. fluorescens strain MKB158 improved grain filling, lignin deposition and oxidative responses, and lowered the content of protease inhibitors and proteins that play a key role in programmed cell death [24]. Méndez-Gómez et al. [55] demonstrated that Azospirillum brasilense cell wall components influenced superoxide (O2(-)) production in wheat roots. Roberti et al. [23] found that the Clonostachys rosea fungus promotes plant growth, lowers peroxidase expression and enhances chitinase isoforms that are putative cell wall disruptors. The analyzed antagonist also induced the expression of PR4 protein which probably plays an important role in plant defense responses against pathogens.
4. Trichothecene Degradation by Microorganisms
4.1. Trichothecene-Degrading Bacteria
The most common biologically-modified products of DON are DOM-1, 3-epi-DON, and 3-keto DON (Figure 2a). The modification products of T-2 toxin that are most frequently described in literature include HT-2 toxin, T-2 triol, T-2 tetraol, neosolaniol (NEO), 3-acetyl T-2 toxin, and 4-deoxy T-2 toxin (Figure 2b) [64,65,66,67]. Trichothecenes are modified by acetylation, deacetylation, de-epoxidation, epimerization, glucosylation, and oxidation. Trichothecene modification products are generally described as less toxic for humans and animals. In a study by Pierron et al. [26], bacterial de-epoxidation or epimerization of DON altered the interaction between the mycotoxin and the ribosome, which prevented the activation of MAPK (mitogen-activated protein kinase) and reduced toxicity. At the cellular and subcellular level, DON exerts toxic effects by binding to the ribosome, which inhibits the synthesis of proteins and nucleic acids, triggers ribotoxic stress, activates kinases, MAPKs, and their downstream signaling pathways [34]. The products of DON’s microbiological modification do not activate this pathway, which eliminates symptoms of acute or chronic poisoning in humans and animals. Mycotoxin glucosides in plants are referred to as masked mycotoxins because they are not always detected with the use of the existing analytical methods and may be hydrolyzed during digestion in animals or humans [11]. Analytical methods based on liquid chromatography coupled with mass spectrometry (LC-MS) are widely used for the determination of mycotoxins and their plant metabolites. Many plant biotransformation products of mycotoxins are difficult to detect by conventional analytical methods, and no regulatory limits have been set for them in food commodities [68]. Trichothecene glucosides have been isolated from Fusarium infected plant material [69]. The group of masked mycotoxins comprises not only some specific mycotoxins, such as DON and zearalenone, but also other Fusarium mycotoxins [70].
DON degradation is observed in aerobic and anaerobic organisms. Aerobic bacteria degrade the mycotoxin by oxidation to 3-keto DON with isomerization to 3-epi DON. Anaerobic bacteria [71,72] and aerobic bacteria [73] decompose DON by de-epoxidation to DOM-1 (Table 2). Soil from arable fields, water, cereal leaves and spikes can act as reservoirs of bacterial strains with the above properties. Bacteria isolated from soil where Fusarium-infected cereals were grown can rapidly degrade DON, but only on selected media and within a broad range of temperatures (12–40 °C) and pH (6.0–7.5), which suggests that those microorganisms could be applied in cereal production in temperate and warmer climates [73].
Bacteria of the genera Nocardioides and Devosia can degrade DON to 3-epi-DON and 3-keto DON [74]. After seven days of incubation, Nocardioides bacterial strains isolated from wheat rhizosphere degraded 90% of DON to 3-epi-DON and two unidentified compounds in seven days [66]. In a study by Sato et al. [74], 13 tested bacterial isolates of the genera Nocardioides and Devosia degraded DON at a concentration of 100 μg mL–1 to concentrations below the limit of detection (0.5 µg mL–1). According to the cited authors, the application of the tested isolates led to the production of various metabolites, which points to the cooperative catabolism of DON in the environment: the biodegradation products of one isolate were degraded by another isolate. According to Ito et al. [75], bacteria of the genus Marmoricola obtained from wheat spikes sprayed with DON or inoculated with F. graminearum effectively reduced DON contamination levels in wheat and barley grain. The cited studies demonstrated for the first time that bacteria of the genus Marmoricola are capable of degrading DON. They also revealed that Marmoricola strains can be effectively used to reduce DON levels in feeds containing wheat and barley grain. Cserháti et al. [25] reported that 32 isolates of Rhodococcus bacteria were capable of degrading a broad range of mycotoxins, including trichothecenes. The mechanism that was implicated, but not proven, in the degradation process was the high metabolic activity of bacteria containing large nucleoids and linear megaplasmids. These structures encode for oxidases and other enzymes which enable bacteria to compete for energy and carbon derived from organic compounds. In the above study, the concentrations of T-2 toxin were reduced already 24 h after the bacterial suspension had been introduced to the substrate. The trichothecene-degrading potential of bacteria from cropped soil can be utilized to protect crops or introduce genetic modifications to cereals to increase their resistance to pathogens of the genus Fusarium. The introduction of Biomin® BBSH 797 for detoxifying trichothecenes in animal feed was a commercial success on the EU market. This product contains bacterial strain Eubacterium BBSH 797 which is isolated from bovine rumen fluid [76] and has the ability to degrade several trichothecenes [77].
In recent years, bacterial consortia have been used to transform Fusarium mycotoxins. Ahad et al. [81] demonstrated that a bacterial consortium containing low levels of Gram-positive anaerobic bacteria, including Alkaliphilus crotonatoxidans, Bacillus spp. and Blautia coccoides, the aerobic species of Stenotrophomonas (Gram-negative), was highly effective in de-epoxidizing eleven trichothecene mycotoxins. They suggested that the soil-derived bacterial consortium contained more than 30% of unidentified bacterial species. The bacterial consortium was characterized by a rapid and stable ability to de-epoxidize DON after 48 h of incubation. Vanhoutte et al. [82] developed a biological test involving Lemna minor L. which is eliminated under exposure to DON concentrations higher than 1 mg L−1. When microorganisms derived from soil and activated sludge were incorporated into a macronutrient solution for plant growth, DON concentrations of 5 and 50 mg L−1 were effectively neutralized and modified. In addition, the metabolites 3-epi-DON and the epimer of de-epoxy-DON (3-epi-DOM-1) were found as biotransformation products of both consortia.
4.2. Fungi Capable of Degradation and Acetylation of Trichothecenes
Trichothecenes are toxic for eukaryotic fungal cells, therefore, fungi producing those mycotoxins have developed mechanisms that protect them against the harmful effects of trichothecenes [67]. In the cells of Fusarium pathogens, trichothecenes are biosynthesized by intermediate products of C-3 acetylation, where acetylation is controlled by TRI101 and TRI201 genes encoding trichothecene 3-O-acetyltransferase [35,83]. Acetylation of C-3 carbon (DON ⇒ 3-Ac-DON) reduces DON toxicity [84]. Under exposure to trichothecenes, the activity of trichothecene 3-O-acetyltransferase was also reported in the cells of fungi of the genera Aspergillus, Saccharomyces, Blastobotrys, and Trichomonascus which do not produce those compounds [65,83].
Yeasts belonging to the Trichomonascus clade can degrade T-2 toxin in three ways: by acetylating C-3 carbon, by hydrolyzing the isovaleric group, or by glycosylating C-3 carbon [35]. All three biotransformation products—3-acetyl T-2 toxin, NEO and T-2 toxin glucoside—are significantly less toxic than T-2 toxin [35]. Interestingly, in the group of the 23 analyzed species of yeasts capable of converting T-2 toxin to 3-acetyl T-2 toxin, only three species of Blastobotrys muscicola, B. robertii, B. peoriensis converted T-2 toxin to T-2 toxin 3-glucoside through glycosylation, whereas Trichomonascus petatosporus and four species of the genus Blastobotrys transformed T-2 toxin to two compounds: 3-acetyl T-2 toxin and NEO [35].
Filamentous fungi such as Alternaria alternata, Rhizopus microsporus var. rhizopodiformis and Aspergillus oryzae metabolized DON into hydrolyzable conjugated DON [79]. He et al. [71] demonstrated that Aspergillus tubingensis (strain NJA-1) was capable of hydrolyzing DON, but the metabolites participating in this process and the underlying mechanism of action have not been identified.
Kosawang et al. [21] studied the hyperparasitic fungus Clonostachys rosea IK726, which is resistant to DON, and demonstrated that the fungus produces several synergistic proteins to activate a number of defense mechanisms against the toxin. A more detailed study revealed that DON induces a transcriptome representing genes that encode metabolic enzymes in C. rosea cells, including cytochrome P450 oxidase, cytochrome c oxidase, and stress proteins. The ThiJ/PfpI protein family was most strongly induced in the group of proteins activated by DON. Genes encoding other stress proteins were also expressed under the influence of DON [21]. A recent study by Tian et al. [80] demonstrated that fungi of the genus Trichoderma have the same defense mechanisms as plants when exposed to F. graminearum, where DON is degraded to D3G.
5. Conclusions
Trichothecenes are the most ubiquitous mycotoxins in cereal grains and grain products. Fusarium pathogens are difficult to control with the use of chemical agents during the growing season. Organically-farmed cereals are characterized by a more varied microbiome, which contributes to the elimination of Fusarium fungi. The application of selected antagonistic microorganisms to cereal crops in the growing season can inhibit the proliferation of Fusarium fungi by way of hyperparasitism, competition or antibiosis. Such treatments reduce the severity of fungal infections in grains and indirectly protect plants against infections. Isolates of antagonistic microorganisms effectively reduce trichothecene concentrations in cereal grains by degrading them to non-toxic products or by binding to antagonistic cells. New mycotoxin detection methods and advanced techniques for selecting mycotoxin-degrading microorganisms have led to the development of a commercial product for the biodegradation of trichothecenes in grains and feeds.
Conflicts of Interest
The authors declare no conflict of interest.
Abbreviations
The following abbreviations are used in this manuscript:
| 15-Ac-DON | 15-Acetyl-deoxynivalenol |
| 3-Ac-DON | 3-Acetyl-deoxynivalenol |
| D3G | Deoxynivalenol-3-β-d-glucopyranoside |
| DOM-1 | De-epoxy-deoxynivalenol |
| DON | Deoxynivalenol |
| FHB | Fusarium head blight |
| FPP | Farnesyl pyrophosphate |
| MAPK | Mitogen-activated protein kinase |
| NEO | Neosolaniol |
| NIV | Nivalenol |
References
- Stoev, S.D. Foodborne mycotoxicoses, risk assessment and underestimated hazard of masked mycotoxins and joint mycotoxin effects or interaction. Environ. Toxicol. Pharmacol. 2015, 39, 794–809. [Google Scholar] [CrossRef] [PubMed]
- Pirgozliev, S.R.; Edwards, S.G.; Hare, M.C.; Jenkinson, P. Effect of dose rate of azoxystrobin and metconazole on the development of Fusarium Head Blight and the accumulation of deoxynivalenol (DON) in wheat grain. Eur. J. Plant Pathol. 2002, 108, 469–478. [Google Scholar] [CrossRef]
- European Commission. Commission Regulation (EC) No 1107/2009 of 21 October 2009. Concerning the Placing of Plant Protection Products on the Market. Available online: http://eur-lex.europa.eu/legal-content/EN/TXT/?uri=celex%3A32009R1107 (accessed on 2 December 2017).
- European Commission. Commission Regulation (EC) No 889/2008 of 5 September 2008 Laying down Detailed Rules for the Implementation of Council Regulation (EC) No 834/2007 on Organic Production and Labeling of Organic Products with Regard to Organic Production, Labeling and Control. Available online: http://eur-lex.europa.eu/legal-content/en/ALL/?uri=CELEX%3A32008R0889 (accessed on 2 December 2017).
- Commission Regulation EC No 1881/2006 of 19 December 2006 Setting Maximum Levels for Certain Contaminants for Foodstuffs. Available online: http://eur-lex.europa.eu/legal-content/EN/TXT?url=CELEX:02006R1881-20170728 (accessed on 2 December 2017).
- Lindblad, M.; Gidlund, A.; Sulyok, M.; Börjesson, T.; Krska, R.; Olsen, M.; Fredlund, E. Deoxynivalenol and other selected Fusarium toxins in Swedish wheat-occurrence and correlation to specific Fusarium species. Int. J. Food Microbiol. 2013, 167, 284–291. [Google Scholar] [CrossRef] [PubMed]
- Commission Recommendation of 27 March 2013 on the Presence T-2 and HT-2 Toxin in Cereals and Cereal Products. Available online: http://eur-lex.europa.eu/legal-content/EN/ALL/?uri=CELEX:32013H0165 (accessed on 2 December 2017).
- Panel, E.C. Scientific opinion on risks for animal and public health related to the presence of nivalenol in food and feed. EFSA J. 2013, 11, 3262. [Google Scholar]
- Okubara, P.A.; Blechl, A.E.; McCormick, S.P.; Alexander, N.J.; Dill-Macky, R.; Hohn, T.M. Engineering deoxynivalenol metabolism in wheat through the expression of a fungal trichothecene acetyltransferase gene. Theor. Appl. Genet. 2002, 106, 74–83. [Google Scholar] [CrossRef] [PubMed]
- Ohsato, S.; Ochiai-Fukuda, T.; Nishiuchi, T.; Takahashi-Ando, N.; Koizumi, S.; Hamamoto, H.; Kudo, T.; Yamaguchi, I.; Kimura, M. Transgenic rice plants expressing trichothecene 3-O-acetyltransferase show resistance to the Fusarium phytotoxin deoxynivalenol. Plant Cell Rep. 2007, 26, 531–538. [Google Scholar] [CrossRef] [PubMed]
- Berthiller, F.; Crews, C.; Dall’Asta, C.; Saeger, S.D.; Haesaert, G.; Karlovsky, P.; Oswald, I.P.; Seefelder, W.; Speijers, G.; Stroka, J. Masked mycotoxins: A review. Mol. Nutr. Food Res. 2013, 57, 165–186. [Google Scholar] [PubMed]
- Rychlik, M.; Humpf, H.U.; Marko, D.; Dänicke, S.; Mally, A.; Berthiller, F.; Klaffke, H.; Lorenz, N. Proposal of a comprehensive definition of modified and other forms of mycotoxins including “masked” mycotoxins. Mycotoxin Res. 2014, 30, 197–205. [Google Scholar] [PubMed]
- Wiwart, M.; Suchowilska, E.; Kandler, W.; Sulyok, M.; Groenwald, P.; Krska, R. Can Polish wheat (Triticum polonicum L.) be an interesting gene source for breeding wheat cultivars with increased resistance to Fusarium head blight? Genet. Resour. Crop Evol. 2013, 60, 2359–2373. [Google Scholar] [CrossRef]
- Kluger, B.; Bueschl, C.; Lemmens, M.; Berthiller, F.; Häubl, G.; Jaunecker, G.; Adam, G.; Krska, R.; Schuhmacher, R. Stable isotopic labelling-assisted untargeted metabolic profiling reveals novel conjugates of the mycotoxin deoxynivalenol in wheat. Anal. Bioanal. Chem. 2013, 405, 5031–5036. [Google Scholar] [PubMed]
- Pan, D.; Mionetto, A.; Tiscornia, S.; Bettucci, L. Endophytic bacteria from wheat grain as biocontrol agents of Fusarium graminearum and deoxynivalenol production in wheat. Mycotoxin Res. 2015, 31, 137–143. [Google Scholar] [CrossRef] [PubMed]
- Turner, T.R.; James, E.K.; Poole, P.S. The plant microbiome. Genome Biol. 2013, 14, 209–219. [Google Scholar] [CrossRef] [PubMed]
- Gdanetz, K.; Frances Trail, F. The Wheat Microbiome under Four Management Strategies, and Potential for Endophytes in Disease Protection. Phytobiomes 2017, 1, 158–168. [Google Scholar] [CrossRef]
- Doty, S.L. Functional Importance of the Plant Microbiome: Implications for Agriculture, Forestry and Bioenergy; Springer: Berlin, Germany, 2017; p. 111. [Google Scholar]
- Wachowska, U.; Głowacka, K. Antagonistic interactions between Aureobasidium pullulans and Fusarium culmorum, a fungal pathogen of winter wheat. BioControl 2014, 59, 635–645. [Google Scholar] [CrossRef]
- Schisler, D.A.; Core, A.B.; Boehm, M.J.; Horst, L.; Krause, C.; Dunlap, C.A.; Rooney, A.P. Population dynamics of the Fusarium head blight biocontrol agent Cryptococcus flavescens OH 182.9 on wheat anthers and heads. Biol. Control 2014, 70, 17–27. [Google Scholar] [CrossRef]
- Kosawang, C.; Karlsson, M.; Jensen, D.F.; Dilokpimol, A.; Collinge, D.B. Transcriptomic profiling to identify genes involved in Fusarium mycotoxin deoxynivalenol and zearalenone tolerance in the mycoparasitic fungus Clonostachys rosea. BMC Genom. 2014, 15, 55. [Google Scholar] [CrossRef] [PubMed]
- Druefors, U.A.; Schnürer, J. Mold-inhibitory activity of different yeast species during airtight storage of wheat grain. FEMS Yeast Res. 2005, 5, 373–378. [Google Scholar]
- Roberti, R.; Veronesi, A.R.; Cesari, A.; Cascone, A.; Berardino, I.D.; Bertini, L.; Caruso, C. Induction of PR proteins and resistance by the biocontrol agent Clonostachys rosea in wheat plants infected with Fusarium culmorum. Plant Sci. 2008, 175, 339–347. [Google Scholar] [CrossRef]
- Petti, C.; Khan, M.; Doohan, F. Lipid transfer proteins and protease inhibitors as key factors in the priming of barley responses to Fusarium head blight disease by a biocontrol strain of Pseudomonas fluorescens. Funct. Integr. Genom. 2010, 10, 619–627. [Google Scholar] [CrossRef] [PubMed]
- Cserháti, M.; Kriszt, B.; Krifaton, C.; Szoboszlay, S.; Hahn, J.; Toth, S.; Nagy, I.; Kukolya, J. Mycotoxin-degradation profile of Rhodococcus strains. Int. J. Food Microbiol. 2013, 166, 176–185. [Google Scholar] [CrossRef] [PubMed]
- Pierron, A.; Mimoun, S.; Murate, L.S.; Loiseau, N.; Lippi, Y.; Bracarense, A.P.F.L.; Schatzmayr, G.; He, J.W.; Zhou, T.; Moll, W.D.; et al. Microbial biotransformation of DON: Molecular basis for reduced toxicity. Sci. Rep. 2016, 6, 29105. [Google Scholar] [CrossRef] [PubMed]
- Grove, J.F. The Trichothecenes and Their Biosynthesis. In Progress in the Chemistry of Organic Natural Products; Herz, W., Falk, H., Kirby, G.W., Eds.; Springer: Vienna, Austria, 2007; pp. 63–130. ISBN 978-3-211-20688-1. [Google Scholar]
- McCormick, S.P.; Stanley, A.M.; Stover, N.A.; Alexander, N.J. Trichothecenes: From Simple to Complex Mycotoxins. Toxins 2011, 3, 802–814. [Google Scholar] [CrossRef] [PubMed]
- Arunachalam, C.; Doohan, F.M. Trichothecene toxicity in eukaryotes: Cellular and molecular mechanisms in plants and animals. Toxicol. Lett. 2013, 217, 149–158. [Google Scholar] [CrossRef] [PubMed]
- Stępień, Ł. The use of Fusarium secondary metabolite biosynthetic genes in chemotypic and phylogenetic studies. Crit. Rev. Microbiol. 2014, 40, 176–185. [Google Scholar] [CrossRef] [PubMed]
- Pestka, J. Toxicological mechanisms and potential health effects of deoxynivalenol and nivalenol. World Mycotoxin J. 2010, 3, 323–347. [Google Scholar] [CrossRef]
- Li, Y.; Wang, Z.; Beier, R.C.; Shen, J.; Smet, D.; Saeger, S.; Zhang, S. T-2 toxin, a trichothecene mycotoxins: Review of toxicity, metabolism, and analytical methods. J. Agric. Food Chem. 2011, 8, 3441–3453. [Google Scholar] [CrossRef] [PubMed]
- Goossens, J.; Devreese, M.; Pasmans, F.; Osselaere, A.; De Baere, S.; Verbrugghe, E.; Haesebrouck, F.; De Backer, P.; Croubels, S. Chronic exposure to the mycotoxin T-2 promotes oral absorption of chlortetracycline in pigs. J. Vet. Pharmacol. Ther. 2013, 36, 621–624. [Google Scholar] [CrossRef] [PubMed]
- Garreau de Loubresse, N.; Prokhorova, I.; Holtkamp, W.; Rodnina, M.V.; Yusupova, G.; Yusupov, M. Structural basis for the inhibition of the eukaryotic ribosome. Nature 2014, 513, 517–522. [Google Scholar] [CrossRef] [PubMed]
- McCormick, S.P.; Price, N.P.J.; Kurtzman, C.P. Glucosylation and other biotransformations of T-2 toxin by yeasts of the Trichomonascus clade. Appl. Environ. Microbiol. 2012, 78, 8694–8702. [Google Scholar] [CrossRef] [PubMed]
- Lee, T.; Lee, S.H.; Shin, J.Y.; Kim, H.K.; Yun, S.H.; Kim, H.Y.; Lee, S.; Ryu, J.G. Comparison of Trichothecene Biosynthetic Gene Expression between Fusarium graminearum and Fusarium asiaticum. Plant Pathol. J. 2014, 30, 33–42. [Google Scholar] [CrossRef] [PubMed]
- Cardoza, R.E.; Malmierca, M.G.; Hermosa, M.R.; Alexander, N.J.; McCormick, S.P.; Proctor, R.H.; Tijerino, A.M.; Rumbero, A.; Monte, E.; Gutierrez, S. Identification of Loci and Functional Characterization of Trichothecene Biosynthesis Genes in Filamentous Fungi of the Genus Trichoderma. Appl. Environ. Microbiol. 2011, 77, 4867–4877. [Google Scholar] [CrossRef] [PubMed]
- Kimura, M.; Tokai, T.; Takahashi-Ando, N.; Ohsato, S.; Fujimura, M. Molecular and genetic studies of Fusarium trichothecene biosynthesis: Pathways, genes and evolution. Biosci. Biotechnol. Biochem. 2007, 71, 2105–2123. [Google Scholar] [CrossRef] [PubMed]
- Brown, D.W.; McCormick, S.P.; Alexander, N.J.; Proctor, R.H.; Desjardins, A.E. A genetic and biochemical approach to study trichothecene diversity in Fusarium sporotrichioides and Fusarium graminearum. Fungal Genet. Biol. 2001, 32, 121–133. [Google Scholar] [CrossRef] [PubMed]
- Brown, D.W.; McCormick, S.P.; Alexander, N.J.; Proctor, R.H.; Desjardins, A.E. Inactivation of a cytochrome P-450 is a determinant of trichothecene diversity in Fusarium species. Fungal Genet. Biol. 2002, 36, 224–233. [Google Scholar] [CrossRef]
- Lee, T.; Han, Y.K.; Kim, K.H.; Yun, S.H.; Lee, Y.W. Tri13 and Tri17 determine deoxynivalenol- and nivalenol-producing chemotypes of Gibberella zeae. Appl. Environ. Microbiol. 2002, 68, 2148–2154. [Google Scholar] [CrossRef] [PubMed]
- Moretti, A.; Susca, A.; Mule, G.; Logrieco, A.F.; Proctor, R.H. Molecular biodiversity of mycotoxigenic fungi that threaten food safety. Int. J. Food Microbiol. 2013, 167, 57–66. [Google Scholar] [CrossRef] [PubMed]
- Lia, H.P.; Wu, A.B.; Zhao, C.S.; Scholten, O.; Löffler, H.; Lia, Y.C. Development of a generic PCR detection of deoxynivalenol- and nivalenol-chemotypes of Fusarium graminearum. FEMS Microbiol. Lett. 2005, 243, 505–511. [Google Scholar] [CrossRef] [PubMed]
- Varga, E.; Wiesenberger, G.; Hametner, C.; Ward, T.J.; Dong, Y.; Schöfbeck, D.; McCormick, S.; Broz, K.; Stückler, R.; Schuhmacher, R.; et al. New tricks an old enemy: Isolates of Fusarium graminearum produce a type A trichothecene mycotoxin. Environ. Microbiol. 2015, 17, 2588–2600. [Google Scholar] [CrossRef] [PubMed]
- Lutz, M.P.; Feichtinger, G.; Defago, G.; Duffy, B. Mycotoxigenic Fusarium and deoxynivalenol production repress chitinase gene expression in the biocontrol agent Trichoderma atroviride P1. Appl. Environ. Microbiol. 2003, 69, 3077–3084. [Google Scholar] [CrossRef] [PubMed]
- Dunlap, C.A.; Schisler, D.A.; Price, N.P.; Vaughn, S.F. Cyclic lipopeptide profile of three Bacillus subtilis strains; antagonists of Fusarium head blight. J. Microbiol. 2011, 49, 603–609. [Google Scholar] [CrossRef] [PubMed]
- Palazzini, J.M.; Dunlap, C.A.; Bowman, M.J.; Sofía, N.; Chulze, S.N. Bacillus velezensis RC 218 as a biocontrol agent to reduce Fusarium head blight and deoxynivalenol accumulation: Genome sequencing and secondary metabolite cluster profiles. Microbiol. Res. 2016, 192, 30–36. [Google Scholar] [CrossRef] [PubMed]
- Zhao, H.; Vegi, A.; Wolf-Hall, C. Screening of lactic acid bacteria for anti-Fusarium activity and optimization of incubation conditions. J. Food Prot. 2017, 80, 1648–1656. [Google Scholar] [CrossRef] [PubMed]
- Baffoni, L.; Gaggia, F.; Dalanaj, N.; Prodi, A.; Nipoti, P.; Pisi, A.; Biavati, B.; Di Gioia, D. Microbial inoculants for the biocontrol of Fusarium spp. in durum wheat. BMC Microbiol. 2015, 15, 242. [Google Scholar] [CrossRef] [PubMed]
- Chan, Y.K.; Savard, M.E.; Reid, L.M.; Cyr, T.; McCormick, W.A.; Seguin, C. Identification of lipopeptide antibiotics of a Bacillus subtilis isolate and their control of Fusarium graminearum diseases in maize and wheat. BioControl 2009, 54, 567–574. [Google Scholar] [CrossRef]
- Shi, C.; Yan, P.; Li, J.; Wu, H.; Li, Q.; Guan, S. Biocontrol of Fusarium graminearum growth and deoxynivalenol production in wheat kernels with bacterial antagonists. Int. J. Environ. Res. Public Health 2014, 11, 1094–1105. [Google Scholar] [CrossRef]
- Nagaraja, H.; Chennappa, G.; Rakesh, S.; Naik, M.K.; Amaresh, Y.S.; Sreenivasa, M.Y. Antifungal activity of Azotobacter nigricans against trichothecene-producing Fusarium species associated with cereals. Food Sci. Biotechnol. 2016, 25, 1197–1204. [Google Scholar] [CrossRef]
- Mnasri, N.; Chennaoui, C.; Gargouri, S.; Mhamdi, R.; Hessini, K.; Elkahoui, S.; Djébali, N. Efficacy of some rhizospheric and endophytic bacteria in vitro and as seed coating for the control of Fusarium culmorum infecting durum wheat in Tunisia. Eur. J. Plant Pathol. 2017, 147, 501–515. [Google Scholar] [CrossRef]
- Alimi, M.; Soleimani, M.J.; Darzi, M.T. Characterization and application of microbial antagonists for control of Fusarium head blight of wheat caused by Fusarium graminearum using single and mixture strain of antagonistic bacteria on resistance and susceptible cultivars. Afr. J. Microbiol. Res. 2012, 6, 326–334. [Google Scholar]
- Méndez-Gómez, M.; Castro-Mercado, E.; Alexandre, G.; García-Pineda, E. Oxidative and antioxidative responses in the wheat-Azospirillum brasilense interaction. Protoplasma 2016, 253, 477–486. [Google Scholar] [CrossRef] [PubMed]
- Hilber-Bodmer, M.; Schmid, M.; Ahrens, C.H.; Freimoser, F.M. Competition assays and physiological experiments of soil and phyllosphere yeasts identify Candida subhashii as a novel antagonist of filamentous fungi. BMC Microbiol. 2017, 17, 4. [Google Scholar] [CrossRef] [PubMed]
- Vujanovic, V.; Goh, Y.K. Sphaerodes mycoparasitica biotrophic mycoparasite of 3-acetyldeoxynivalenol- and 15-acetyldeoxynivalenol-producing toxigenic Fusarium graminearum chemotypes. FEMS Microbiol. Lett. 2011, 316, 136–143. [Google Scholar] [CrossRef] [PubMed]
- Mahmoud, A.F. Genetic Variation and Biological Control of Fusarium graminearum Isolated from Wheat in Assiut-Egypt. Plant Pathol. J. 2016, 32, 145–156. [Google Scholar] [CrossRef] [PubMed]
- Mamarabadi, M.; Dan Funck, J.; Mette, L. An N-acetyl-β-d-glucosaminidase gene, cr-nag1, from the biocontrol agent Clonostachys rosea is up-regulated in antagonistic interactions with Fusarium culmorum. Mycol. Res. 2009, 1, 33–43. [Google Scholar] [CrossRef] [PubMed]
- Ferre, F.S.; Santamarina, M.P. Efficacy of Trichoderma harzianum in suppression of Fusarium culmorum. Ann. Microbiol. 2010, 60, 335–340. [Google Scholar] [CrossRef]
- Zou, C.S.; Mo, M.H.; Gu, Y.Q.; Zhou, J.P.; Zhang, K.Q. Possible contributions of volatile-producing bacteria to soil fungistases. Soil Biol. Biochem. 2007, 39, 2371–2379. [Google Scholar] [CrossRef]
- Chi, Z.; Wang, X.X.; Ma, Z.C.; Buzdar, M.A.; Chiet, Z.M. The unique role of siderophore in marine-derived Aureobasidium pullulans HN6.2. Biometals 2012, 25, 219–230. [Google Scholar] [CrossRef] [PubMed]
- Yu, X.; Ai, C.; Xin, L.; Zhou, G. The siderophore-producing bacterium, Bacillus subtilis CAS15, has a biocontrol effect on Fusarium wilt and promotes the growth of pepper. Eur. J. Soil Biol. 2011, 47, 138–145. [Google Scholar] [CrossRef]
- Alexander, N.J.; McCormick, S.P.; Hohn, T.M. The identification of the Saccharomyces cerevisiae gene AYT1(ORF-YLL063c) encoding an acetyltransferase. Yeast 2002, 19, 1425–1430. [Google Scholar] [CrossRef] [PubMed]
- Beeton, S.; Bull, A.T. Biotransformation and detoxification of T-2 toxin by soil and freshwater bacteria. Appl. Environ. Microbiol. 1989, 55, 190–197. [Google Scholar] [PubMed]
- Ikunaga, Y.; Sato, I.; Grond, S.; Numaziri, N.; Yoshida, S.; Yamaya, H.; Hiradate, S.; Hasegawa, M.; Toshima, H.; Koitabashi, M.; et al. Nocardioides sp. strain WSN05-2, isolated from a wheat field, degrades deoxynivalenol, producing the novel intermediate 3-epi-deoxynivalenol. Appl. Microbiol. Biotechnol. 2011, 89, 419–427. [Google Scholar] [CrossRef] [PubMed]
- McCormick, S.P. Microbial Detoxification of Mycotoxins. J. Chem. Ecol. 2013, 39, 907–918. [Google Scholar] [CrossRef] [PubMed]
- Meng-Reiterer, J.; Buesch, C.; Rechhaler, J.; Berthiller, F.; Lemmens, M.; Schuhmacher, R. Metabolism of HT-2 toxin and T-2-toxin in oats. Toxins 2016, 8, 364. [Google Scholar] [CrossRef] [PubMed]
- Mc Cormick, S.P.; Kato, T.; Maragos, C.M.; Busman, M.; Lattanzio, V.M.T.; Galaverna, G.; Dall-Asta, C.; Crich, D.; Price, N.P.J.; Kurtzman, C.P. Anomericity of T-2 toxin-glucoside: Masked mycotoxin in cereal crops. J. Agric. Food Chem. 2015, 63, 731–738. [Google Scholar] [CrossRef] [PubMed]
- Nakagawa, H. Research on mycotoxin glucosides (masked mycotoxins). JSM Mycotoxins 2016, 66, 21–25. [Google Scholar] [CrossRef]
- He, C.; Fan, Y.; Liu, G.; Zhang, H. Isolation and identification of a strain of Aspergillus tubingensis with deoxynivalenol biotransformation capability. Int. J. Mol. Sci. 2008, 9, 2366–2375. [Google Scholar] [CrossRef] [PubMed]
- Yu, H.; Zhou, T.; Gong, J.; Young, C.; Su, X.; Li, X.Z.; Zhu, H.; Tsao, R.; Yang, R. Isolation of deoxynivalenol-transforming bacteria from the chicken intestines using the approach of PCR-DGGE guided microbial selection. BMC Microbiol. 2010, 10, 182. [Google Scholar] [CrossRef] [PubMed]
- Islam, R.; Zhou, T.; Young, J.C.; Goodwin, P.H.; Pauls, K.P. Aerobic and anaerobic de-epoxidation of mycotoxin deoxynivalenol by bacteria originating from agricultural soil. World J. Microbiol. Biotechnol. 2012, 28, 7–13. [Google Scholar] [CrossRef] [PubMed]
- Sato, I.; Ito, M.; Ishizaka, M.; Ikunaga, Y.; Sato, Y.; Yoshida, S.; Koitabashi, M.; Tsushima, S. Thirteen novel deoxynivalenol-degrading bacteria are classified within two genera with distinct degradation mechanisms. FEMS Microbiol. Lett. 2012, 327, 110–117. [Google Scholar] [CrossRef] [PubMed]
- Ito, M.; Sato, I.; Koitabashi, M.; Yoshida, S.; Imai, M.; Tsushima, S. A novel actinomycete derived from wheat heads degrades deoxynivalenol in the grain of wheat and barley affected by Fusarium head blight. Appl. Microbiol. Biotechnol. 2012, 96, 1059–1070. [Google Scholar] [CrossRef] [PubMed]
- Fuchs, E.; Binder, E.M.; Heidler, D.; Krska, R. Structural characterization of metabolites after the microbial degradation of A- and B-trichothecenes by the bacterial strain BBSH 797. Mycotoxin Res. 2002, 16, 66–69. [Google Scholar] [CrossRef] [PubMed]
- Vanhoutte, I.; Audenaert, K.; De Gelder, L. Biodegradation of Mycotoxins: Tales from Known and Unexplored Worlds. Front. Microbiol. 2016, 7, 561. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ueno, Y.; Nakayama, K.; Ishii, K.; Tashiro, F.; Minoda, Y.; Omori, T.; Komagata, K. Metabolism of T-2 toxin in Curtobacterium sp. strain 114-2. Appl. Environ. Microbiol. 1983, 46, 120–127. [Google Scholar] [PubMed]
- Tran, S.T.; Smith, T.K. Conjugation of deoxynivalenol by Alternaria alternata (54028 NRRL), Rhizopus microsporus var. rhizopodiformis (54029 NRRL) and Aspergillus oryzae (5509 NRRL). Mycotoxin Res. 2014, 28, 7–13. [Google Scholar] [CrossRef] [PubMed]
- Tian, Y.; Tan, Y.; Liu, N.; Yan, Z.; Liao, Y.; Chen, J.; de Saeger, S. Detoxification of Deoxynivalenol via Glycosylation Represents Novel Insights on Antagonistic Activities of Trichoderma when Confronted with Fusarium graminearum. Toxins 2016, 8, 335. [Google Scholar] [CrossRef] [PubMed]
- Ahad, R.; Zhou, T.; Lepp, D.; Pauls, K.P. Microbial detoxification of eleven food and feed contaminating trichothecene mycotoxins. BMC Biotechnol. 2017, 17, 30. [Google Scholar] [CrossRef] [PubMed]
- Vanhoutte, I.; De Mets, L.; De Boevre, M.; Uka, V.; Di Mavungu, J.D.; De Saeger, S.; De Gelder, L.; Audenaert, K. Microbial detoxification of deoxynivalenol (DON), assessed via a Lemna minor L. bioassay, through biotransformation to 3-epi-DON and 3-epi-DOM-1. Toxins 2017, 9, 63. [Google Scholar] [CrossRef] [PubMed]
- Khatibi, P.A.; Newmister, S.A.; Rayment, I.; McCormick, S.P.; Alexander, N.J.; Schmale, D.G. Bioprospecting for trichothecene-3-O-acetyltransferases in the fungal genus Fusarium yields functional enzymes that vary in their ability to modify the mycotoxin deoxynivalenol. Appl. Environ. Microbiol. 2011, 77, 1162–1170. [Google Scholar] [CrossRef] [PubMed]
- Eriksen, G.S.; Pettersson, H.; Lundh, T. Comparative cytotoxicity of deoxynivalenol, nivalenol, their acetylated derivatives and de-epoxy metabolites. Food Chem. Toxicol. 2004, 42, 619–624. [Google Scholar] [CrossRef] [PubMed]
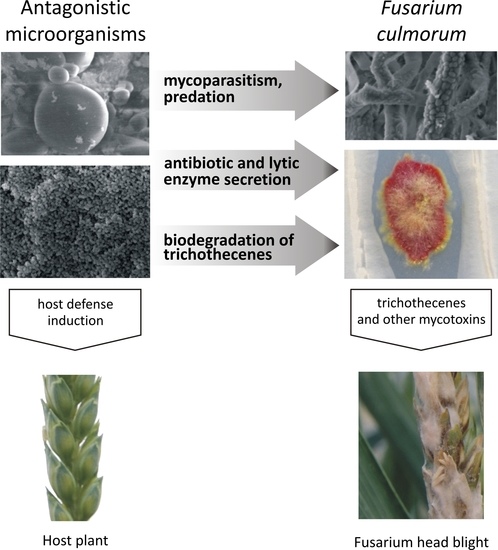
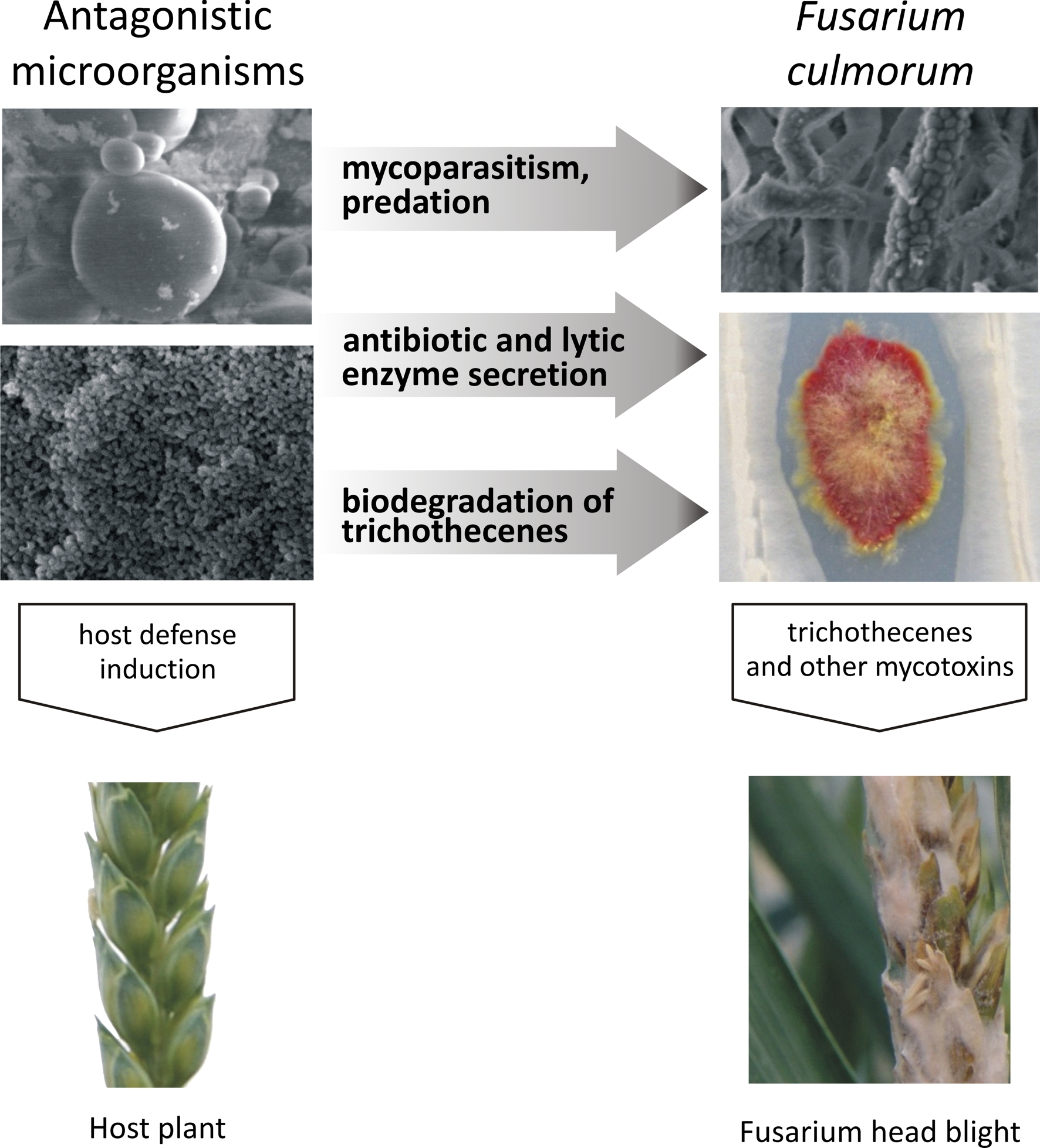
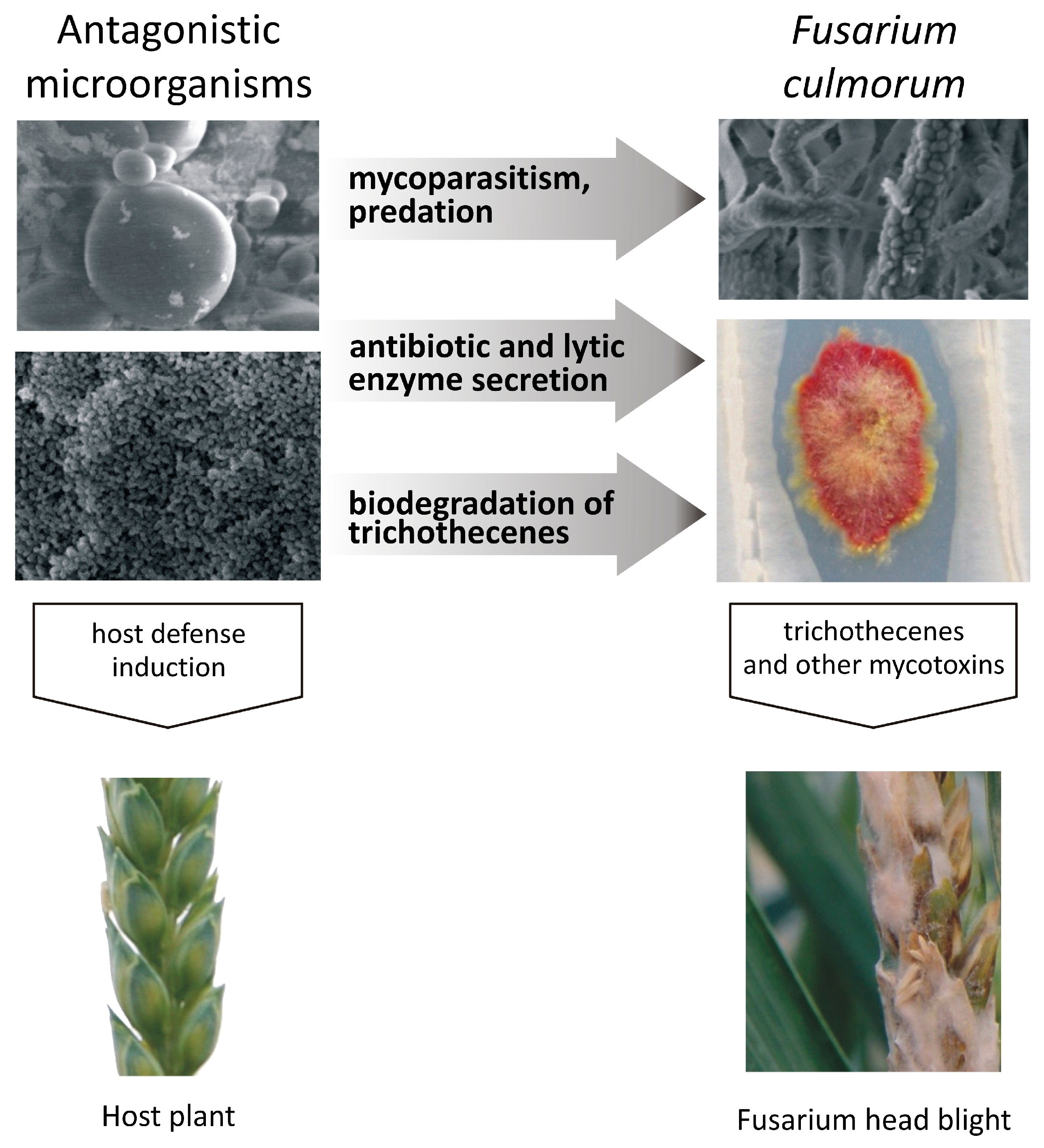
Figure 1.
Inhibition of Fusarium pathogens by antagonistic microorganisms: direct inhibition (mycoparasitism, predation), mixed-path antagonism (antibiotic and lytic enzyme secretion, biological modification of trichothecenes), and indirect inhibition involving interactions with the host plant (induction of host defense responses).
Figure 1.
Inhibition of Fusarium pathogens by antagonistic microorganisms: direct inhibition (mycoparasitism, predation), mixed-path antagonism (antibiotic and lytic enzyme secretion, biological modification of trichothecenes), and indirect inhibition involving interactions with the host plant (induction of host defense responses).
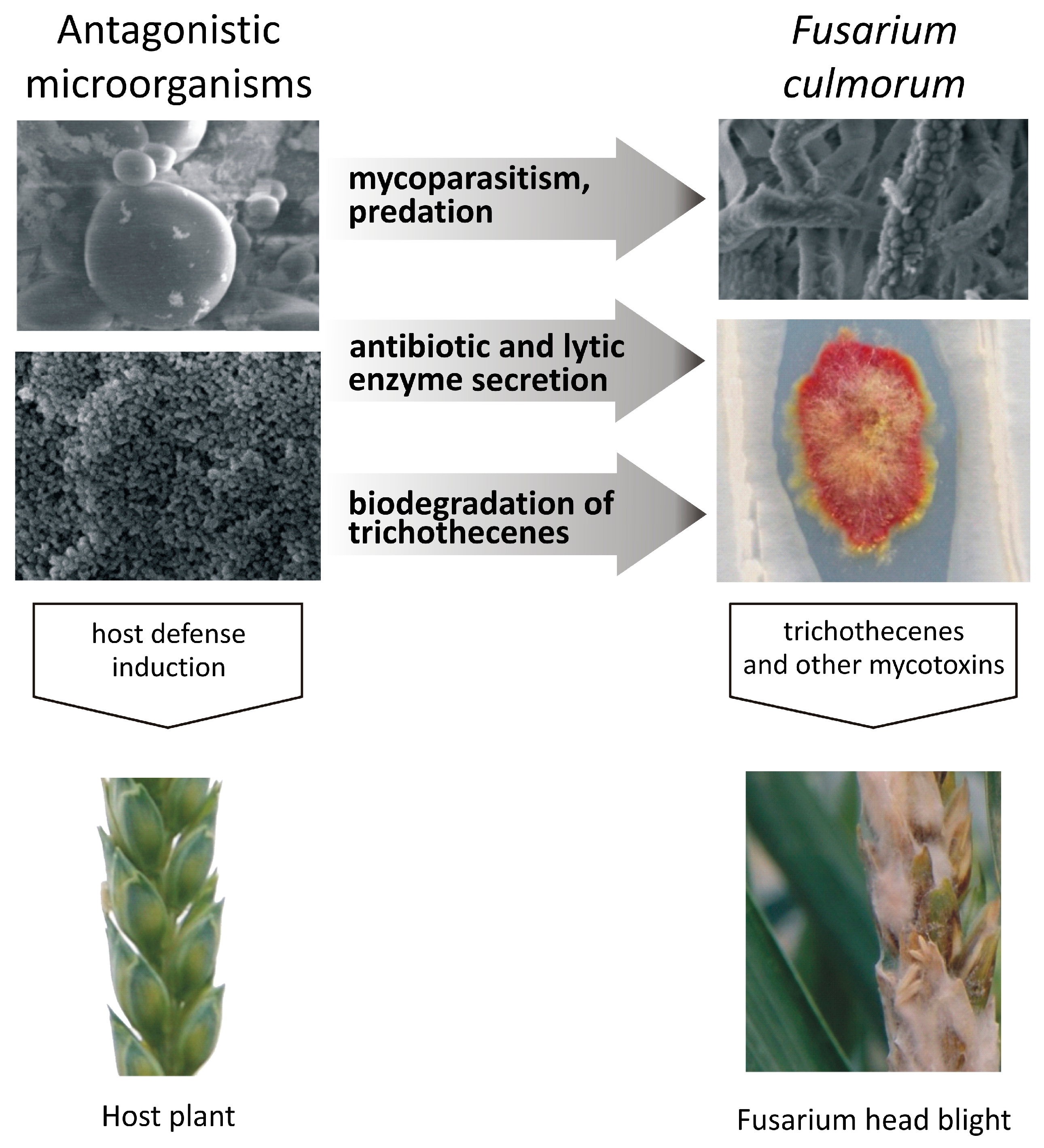
Figure 2.
Degradation products of DON (a) and T-2 toxin (b).


{kind=link}
{kind=link}
{kind=link}
Table 1.
Mechanisms of action by which bacteria and fungi degrade Fusarium pathogens.
| Microorganisms | Microbial Mechanisms of Action | Pathogens | Reduction in DON Content | Reference |
|---|---|---|---|---|
| Bacteria | ||||
| Bacillus megaterium, Bacillus subtilis | Not studied | Fusarium graminearum | 50–89% | [15] |
| Bacillus subtilis | Fungicidal or fungistatic metabolites: surfactin, iturin and fengycin lipopeptides | F. graminearum | not studied | [46] |
| Bacillus subtilis | Antifungal lipopeptides, fengycin-like isoforms | F. graminearum | 51% | [50] |
| Bacillus amyloliquefaciens | Production of antibiotics or competition for nutrients | F. graminearum | 16–90% | [51] |
| Azotobacter nigricans | Antifungal activity | F. sporotrichioides, F. poae, F. crookwellense, F. equiseti, F. graminearum, F. sambucinum, F. culmorum | not studied | [52] |
| Bacillus, Pseudomonas, Microbacterium | Promotion of plant growth | F. culmorum | not studied | [53] |
| Bacillus cereus | Volatile antagonistic compounds | F. graminearum | not studied | [54] |
| Pseudomonas fluorescens | Elicit host defense responses | F. culmorum | not studied | [24] |
| Azospirillium brasilense | Elicit host defense responses | root pathogens | not studied | [55] |
| Yeasts | ||||
| Cryptococcus flavescens | Competitive inhibition via aggressive colonization of plant | F. graminearum | 15–18% | [20] |
| Metschnikowia pulcherrima, Hanseniaspora sp., Cyberlindnera sargentensis, Aureobasidium pullulans, Candida subhashii, Pichia kluyveri | Strong competition for micronutrients and/or macronutrients | F. poae, F. langsethiae, F. graminearum, F. culmorum, F. crookwellense, F. oxysporum | not studied | [56] |
| Sphaerodes mycoparasitica | Hyperparasitism | F. graminearum | not studied | [57] |
| Trichoderma harzianum | Antagonistic activity | F. graminearum | not studied | [58] |
| Trichoderma atroviride | Hyperparasitism | F. graminearum | not studied | [45] |
| Clonostachys rosea | Cell wall degrading enzymes: chitinases, glucanases and proteases | F. culmorum | not studied | [59] |
| Trichoderma harzianum | Hyperparasitism elicit host defense responses | F. culmorum | not studied | [60] |
| Clonostachys rosea | F. culmorum | not studied | [23] |
DON-deoxynivalenol.
Table 2.
Trichothecene-degrading microorganisms.
| Microorganisms | Origin of Isolates | Mycotoxins | Efficiency of Biodegradation | Biodegradation Products | Reference |
|---|---|---|---|---|---|
| Bacteria | |||||
| Lactobacillus plantarum and other lactic acid bacteria | Culture collection | DON, T-2 toxin | 28–35% | Physical adsorption | [61] |
| Nocardioides sp. | Soil | DON | 90% | 3-epi-DON | [66] |
| Marmoricola sp. | Wheat spikes | DON | 100% | Not studied | [75] |
| Serratia, Clostridium, Citrobacter, Enterococcus, Stenotrophomonas, Streptomyces | Soil | DON | 100% | DOM-1 | [73] |
| Rhodococcus erythropolis, Rh. coprophilus, Rh. rhodochrous, Rh. globerulus | Soil/Soil contaminated with oil | T-2 toxin | 90% | Not studied | [25] |
| Pseudomonas sp., Blastobacter sp., Arthrobacter sp., Rhizobiaceae | Leaves, soil, water | T-2 toxin | 100% | NEO, T-2 tetraol, T-2 triol | [65] |
| Curtobacterium sp. | No data | T-2 toxin | 100% | T-2 triol | [78] |
| Devosia sp., Nocardioides sp. | Soil, wheat leaves | DON | 100% | 3-epi DON and other unidentified compounds | [74] |
| Bacillus, Anaerofilum, Collinsella, Clostridiales | Chicken intestines | DON | 32–100% | Not studied | [72] |
| Eubacterium sp. | Rumen | DON | DOM-1 | [76] | |
| Yeasts | |||||
| Blastobotrys muscicola, B. robertii, B. peoriensis | Culture collection | T-2 toxin | T-2 toxin 3-β-d-glucoside | [35] | |
| B. capitulata, B. adeninivorans, B. mokoenaii, B. malaysiensis, B. raffinofermentas, Trichomonascus petasosporum, B. indianensis | 48–100% | NEO | |||
| T. petasosporun, T. ciferri, B. indianensis, B. adeninivorans, B. raffinofermentas, B. mokoenaii, B. malaysiensis, B. nivea, B. terestis, B. arbuscula, B. attinorum, B. parvus, B. serpentis, B. illinoisensis | 3-acetyl T-2 toxin | ||||
| Filamentous fungi | |||||
| Alternaria alternata | Culture collection | DON | 91–97% | DON glutathione conjugates | [79] |
| Rhizopus microsporus var. rhizopodiformis | 93–96% | ||||
| Aspergillus oryzae | 84–97% | ||||
| Aspergillus tubingenesis | Soil | DON | 94% | Hydrolysis | [71] |
| Trichoderma harzianum, T. koningii, T. longibranchiatum, T. atroviride, T. asperellum, T. virens | Culture collection | DON | 70–90% | D3G | [80] |
DON: deoxynivalenol, D3G: deoxynivalenol-3-β-d-glucopyranoside, NIV: nivalenol, 3-Ac-DON: 3-acetyl-deoxynivalenol, 15-Ac-DON: 15-acetyldeoxynivalenol, DOM-1: deepoxydeoxynivalenol, NEO: neosolaniol.
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Wachowska, U.; Packa, D.; Wiwart, M. Microbial Inhibition of Fusarium Pathogens and Biological Modification of Trichothecenes in Cereal Grains. Toxins 2017, 9, 408. https://doi.org/10.3390/toxins9120408
AMA Style
Wachowska U, Packa D, Wiwart M. Microbial Inhibition of Fusarium Pathogens and Biological Modification of Trichothecenes in Cereal Grains. Toxins. 2017; 9(12):408. https://doi.org/10.3390/toxins9120408
Chicago/Turabian StyleWachowska, Urszula, Danuta Packa, and Marian Wiwart. 2017. "Microbial Inhibition of Fusarium Pathogens and Biological Modification of Trichothecenes in Cereal Grains" Toxins 9, no. 12: 408. https://doi.org/10.3390/toxins9120408
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.