Role of Homologous Fc Fragment in the Potency and Efficacy of Anti‐Botulinum Antibody Preparations

 ,
,
Abstract
:1. Introduction
2. Results
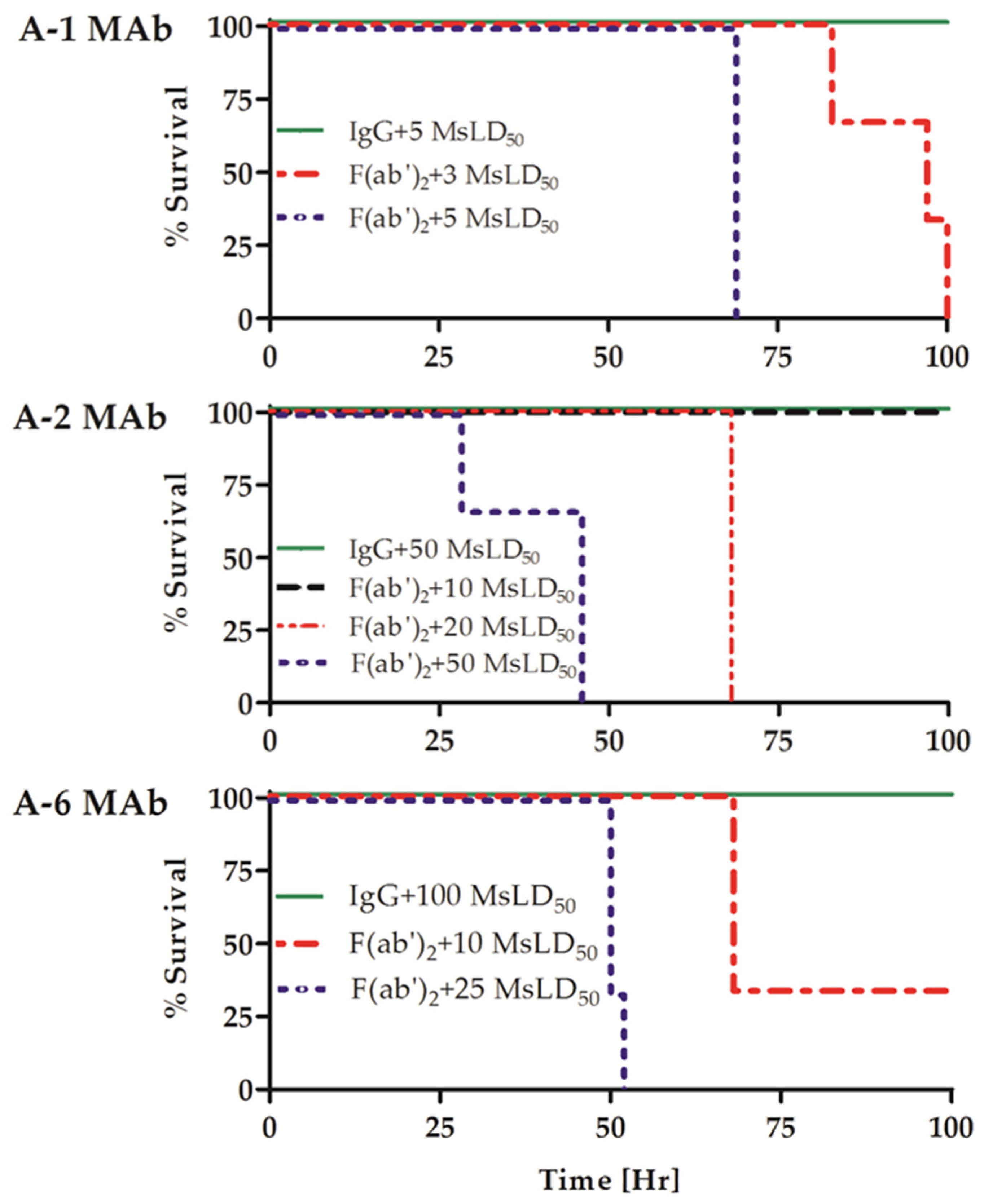
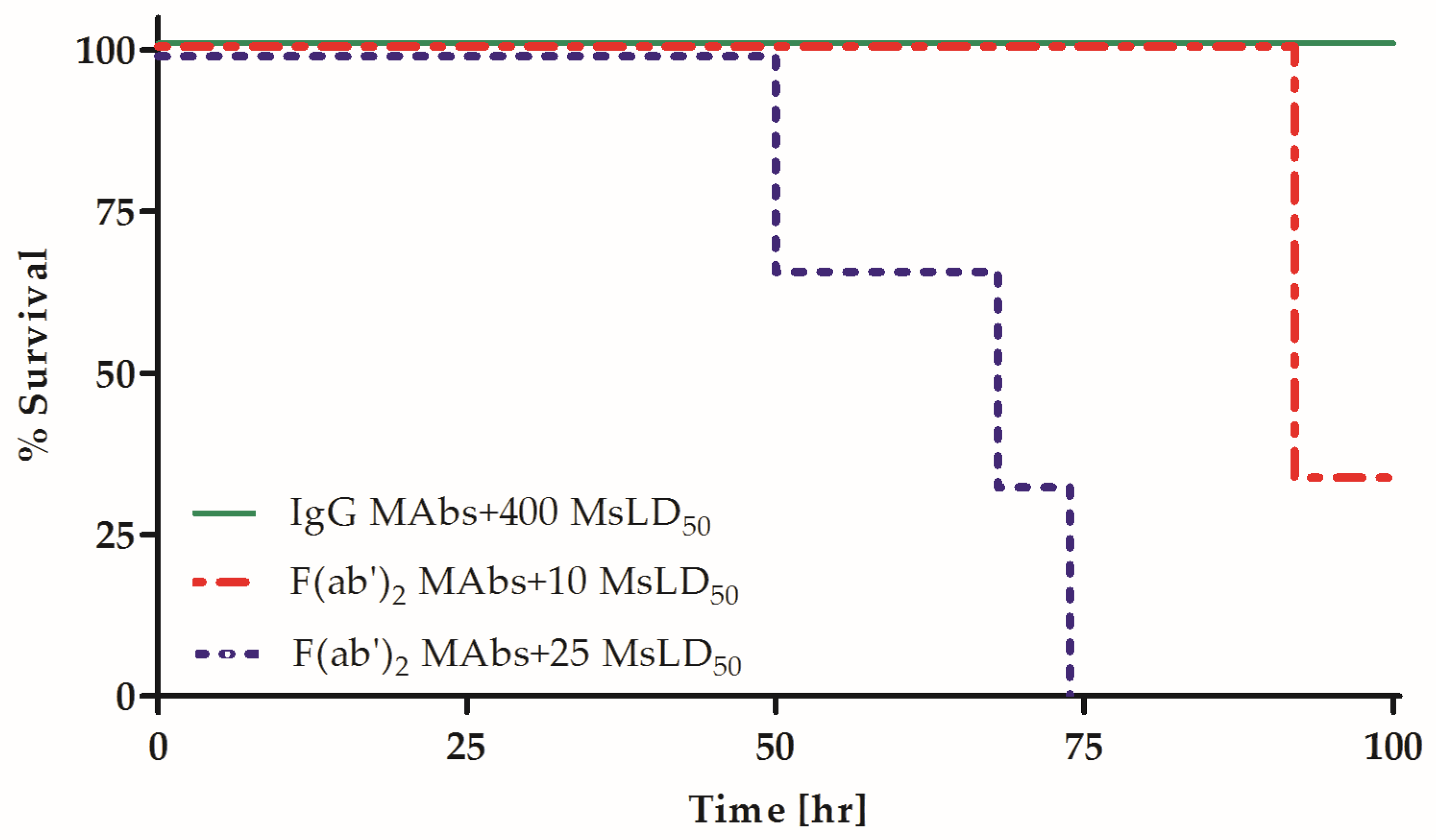
2.1. Individual Homologous Intact MAbs Exhibit Increased Neutralizing Activity Compared to Their F(ab′)2 Fragment Counterparts
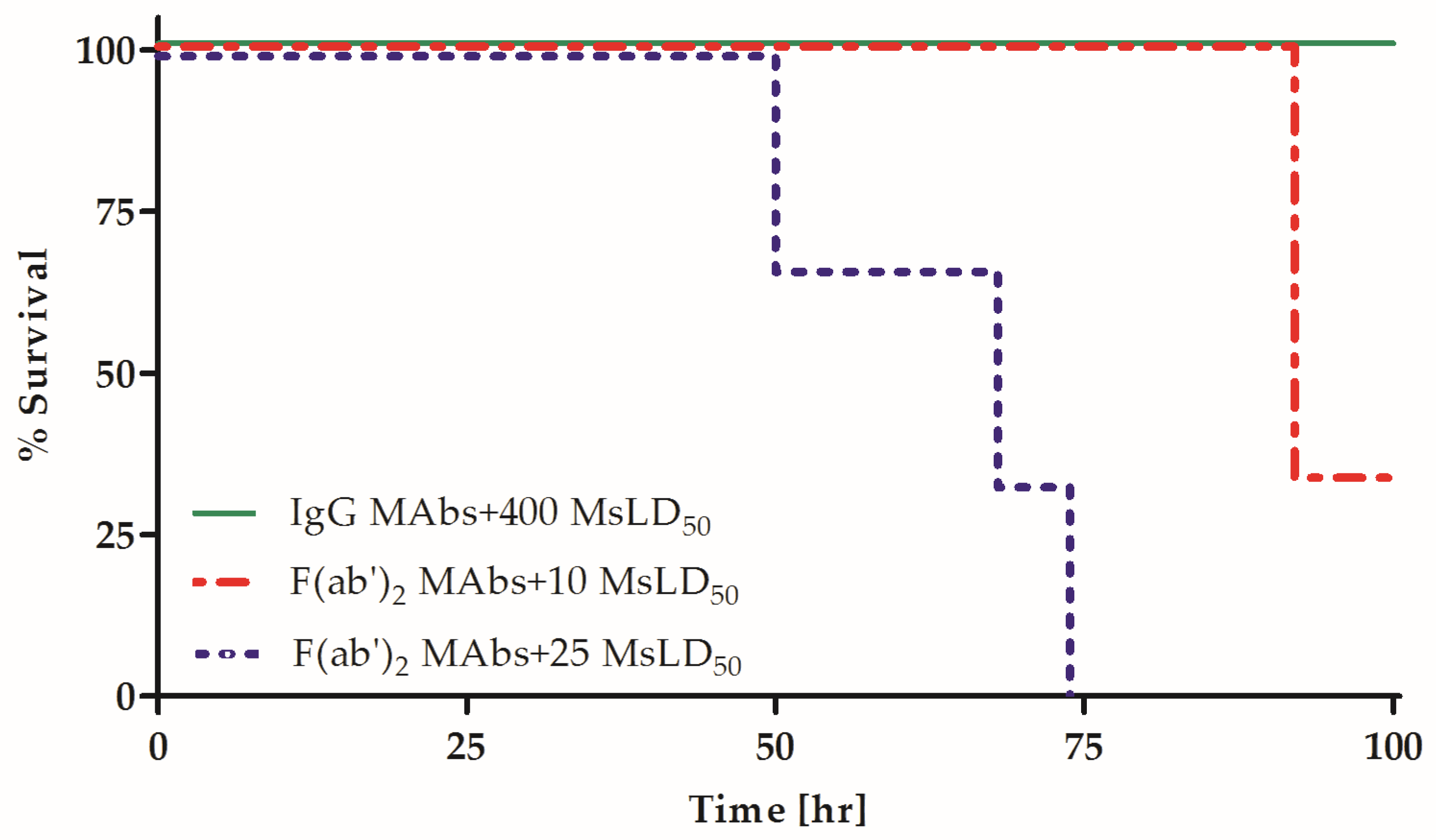
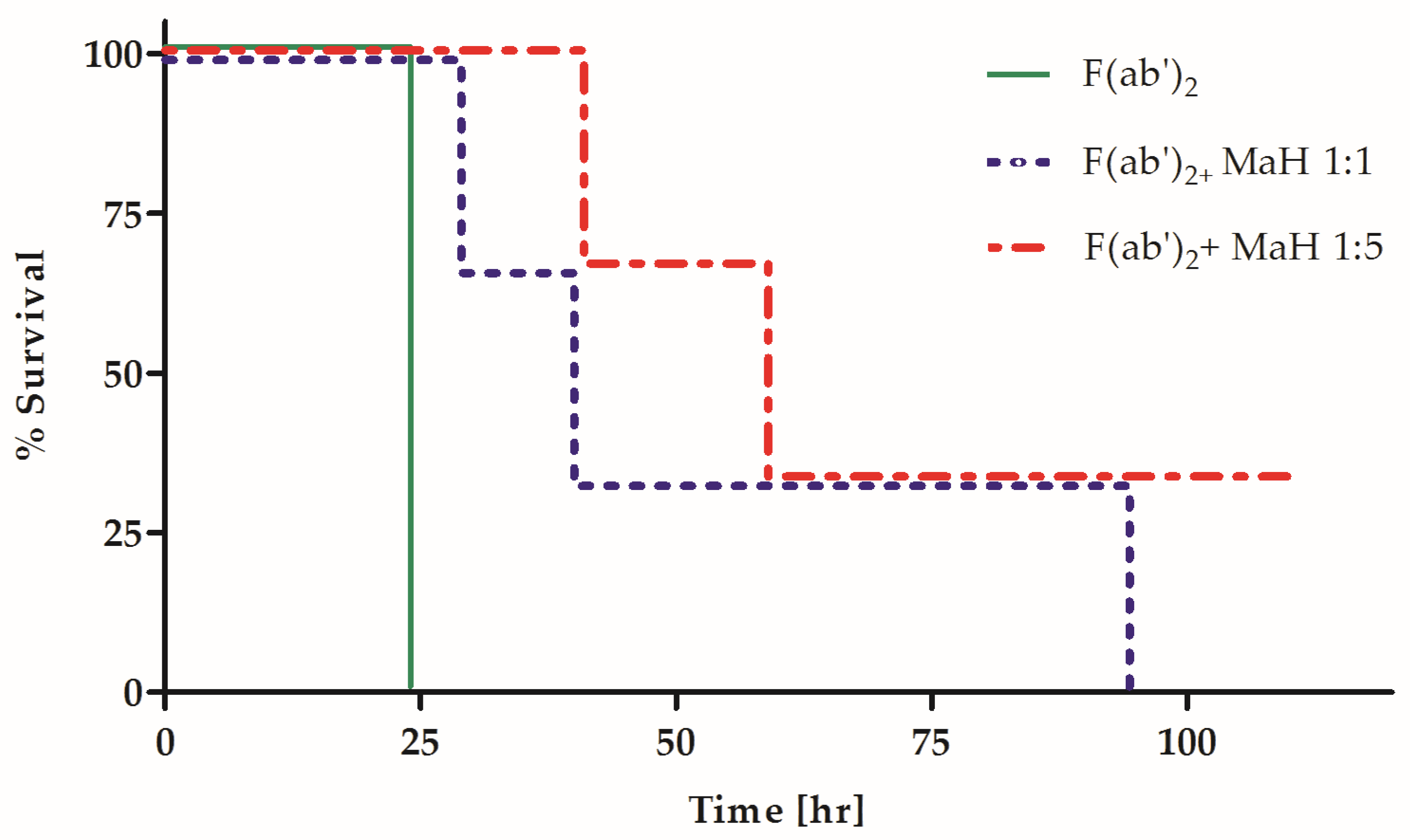
2.2. Fc Is Essential for the Synergistic Neutralizing Activity of Oligoclonal MAb Preparations
2.3. The Potency of a Homologous But Not a Heterologous Hyper-Immune, PAb Preparation Is Dependent on the Presence of Fc Fragments
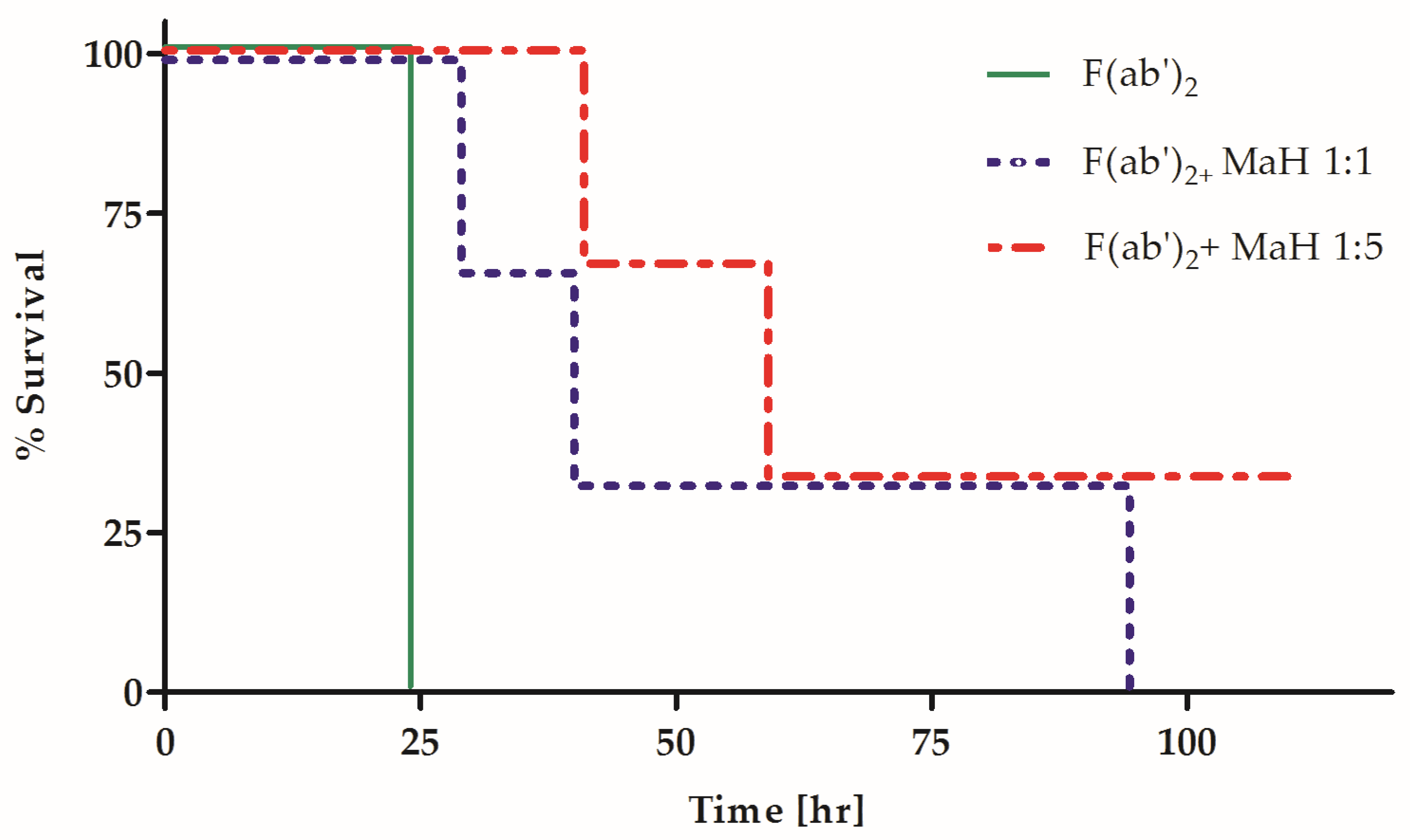
2.4. Homologous Fc Increases the Potency and Efficacy of a Heterologous Anti-BoNT PAb Preparation
2.5. The Contribution of Homologous Fc to Neutralization Potency Depends on In Vivo Interaction with Host-Expressed Components
3. Discussion
4. Materials and Methods
4.1. Ethics Statement
4.2. Materials
4.3. Toxins
4.4. IgG Purification and Generation of F(ab′)2 Fragments
4.5. IgG and F(ab′)2 Fragment Quantification Assay
4.6. Determination of Anti-BoNT IgG and F(ab′)2 Fragment Titers by ELISA
4.7. MAb In Vivo Potency Assay
4.8. PAbs In Vivo Potency Assay
4.9. In Vitro Neutralization Assay
4.10. UPLC Analysis
4.11. Statistical Analysis
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Ecker, D.M.; Jones, S.D.; Levine, H.L. The therapeutic monoclonal antibody market. mAbs 2015, 7, 9–14. [Google Scholar] [CrossRef] [PubMed]
- Mullard, A. FDA approves antitoxin antibody. Nat. Rev. Drug Discov. 2016, 15, 811. [Google Scholar] [CrossRef] [PubMed]
- Reichert, J.M.; Dewitz, M.C. Anti-infective monoclonal antibodies: Perils and promise of development. Nat. Rev. Drug Discov. 2006, 5, 191–195. [Google Scholar] [CrossRef] [PubMed]
- Consroe, P.; Egen, N.B.; Russell, F.E.; Gerrish, K.; Smith, D.C.; Sidki, A.; Landon, J.T. Comparison of a new ovine antigen binding fragment (Fab) antivenin for united states crotalidae with the commercial antivenin for protection against venom-induced lethality in mice. Am. J. Trop. Med. Hyg. 1995, 53, 507–510. [Google Scholar] [CrossRef] [PubMed]
- Black, R.E.; Gunn, R.A. Hypersensitivity reactions associated with botulinal antitoxin. Am. J. Med. 1980, 69, 567–570. [Google Scholar] [CrossRef]
- Hibbs, R.G.; Weber, J.T.; Corwin, A.; Allos, B.M.; Abd el Rehim, M.S.; Sharkawy, S.E.; Sarn, J.E.; McKee, K.T., Jr. Experience with the use of an investigational F(ab′)2 heptavalent botulism immune globulin of equine origin during an outbreak of type E botulism in egypt. Clin. Infect. Dis. 1996, 23, 337–340. [Google Scholar] [CrossRef] [PubMed]
- Diamant, E.; Torgeman, A.; Ozeri, E.; Zichel, R. Monoclonal antibody combinations that present synergistic neutralizing activity: A platform for next-generation anti-toxin drugs. Toxins 2015, 7, 1854–1881. [Google Scholar] [CrossRef] [PubMed]
- Arnon, S.S.; Schechter, R.; Inglesby, T.V.; Henderson, D.A.; Bartlett, J.G.; Ascher, M.S.; Eitzen, E.; Fine, A.D.; Hauer, J.; Layton, M.; et al. Botulinum toxin as a biological weapon: Medical and public health management. JAMA 2001, 285, 1059–1070. [Google Scholar] [CrossRef] [PubMed]
- Gill, D.M. Bacterial toxins: A table of lethal amounts. Microbiol. Rev. 1982, 46, 86–94. [Google Scholar] [PubMed]
- Pirazzini, M.; Rossetto, O.; Eleopra, R.; Montecucco, C. Botulinum neurotoxins: Biology, pharmacology, and toxicology. Pharmacol. Rev. 2017, 69, 200–235. [Google Scholar] [CrossRef] [PubMed]
- Rossetto, O.; Pirazzini, M.; Montecucco, C. Botulinum neurotoxins: Genetic, structural and mechanistic insights. Nat. Rev. Microbiol. 2014, 12, 535–549. [Google Scholar] [CrossRef] [PubMed]
- Atassi, M.Z.; Oshima, M. Structure, activity, and immune (T and B cell) recognition of botulinum neurotoxins. Crit. Rev. Immunol. 1999, 19, 219–260. [Google Scholar] [PubMed]
- The Medical Letter. In brief: Heptavalent botulism antitoxin. In The Medical Letter on Drugs and Therapeutics, 2014/05/30 ed.; The Medical Letter, Inc.: New Rochelle, NY, USA, 2014; Volume 56, p. 44. [Google Scholar]
- Centers for Disease Control and Prevention (CDC). Investigational heptavalent botulinum antitoxin (HBAT) to replace licensed botulinum antitoxin AB and investigational botulinum antitoxin E. MMWR Morb. Mortal. Wkly. Rep. 2010, 59, 299. [Google Scholar]
- Arnon, S.S.; Schechter, R.; Maslanka, S.E.; Jewell, N.P.; Hatheway, C.L. Human botulism immune globulin for the treatment of infant botulism. N. Engl. J. Med. 2006, 354, 462–471. [Google Scholar] [CrossRef] [PubMed]
- Public Health Notifiable Disease Management Guidelines. Botulism. Available online: http://www.health.alberta.ca/professionals/notifiable-diseases-guide.html (accessed on 28 May 2017).
- Dembek, Z.F.; Smith, L.A.; Rusnak, J.M. Botulism: Cause, effects, diagnosis, clinical and laboratory identification, and treatment modalities. Disaster Med. Public Health Prep. 2007, 1, 122–134. [Google Scholar] [CrossRef] [PubMed]
- Meng, Q.; Li, M.; Silberg, M.A.; Conrad, F.; Bettencourt, J.; To, R.; Huang, C.; Ma, J.; Meyer, K.; Shimizu, R.; et al. Domain-based assays of individual antibody concentrations in an oligoclonal combination targeting a single protein. Anal. Biochem. 2012, 421, 351–361. [Google Scholar] [CrossRef] [PubMed]
- Miethe, S.; Rasetti-Escargueil, C.; Avril, A.; Liu, Y.; Chahboun, S.; Korkeala, H.; Mazuet, C.; Popoff, M.R.; Pelat, T.; Thullier, P.; et al. Development of human-like scFv-Fc neutralizing botulinum neurotoxin E. PLoS ONE 2015, 10, e139905. [Google Scholar] [CrossRef] [PubMed]
- Nayak, S.U.; Griffiss, J.M.; McKenzie, R.; Fuchs, E.J.; Jurao, R.A.; An, A.T.; Ahene, A.; Tomic, M.; Hendrix, C.W.; Zenilman, J.M. Safety and pharmacokinetics of XOMA 3AB, a novel mixture of three monoclonal antibodies against botulinum toxin A. Antimicrob. Agents Chemother. 2014, 58, 5047–5053. [Google Scholar] [CrossRef] [PubMed]
- Bournazos, S.; DiLillo, D.J.; Ravetch, J.V. The role of Fc-fcgammaR interactions in IgG-mediated microbial neutralization. J. Exp. Med. 2015, 212, 1361–1369. [Google Scholar] [CrossRef] [PubMed]
- Cheng, L.W.; Stanker, L.H.; Henderson, T.D., 2nd; Lou, J.; Marks, J.D. Antibody protection against botulinum neurotoxin intoxication in mice. Infect. Immun. 2009, 77, 4305–4313. [Google Scholar] [CrossRef] [PubMed]
- Diamant, E.; Lachmi, B.E.; Keren, A.; Barnea, A.; Marcus, H.; Cohen, S.; David, A.B.; Zichel, R. Evaluating the synergistic neutralizing effect of anti-botulinum oligoclonal antibody preparations. PLoS ONE 2014, 9, e87089. [Google Scholar] [CrossRef] [PubMed]
- Ober, R.J.; Radu, C.G.; Ghetie, V.; Ward, E.S. Differences in promiscuity for antibody-FcRn interactions across species: Implications for therapeutic antibodies. Int. Immunol. 2001, 13, 1551–1559. [Google Scholar] [CrossRef] [PubMed]
- Park, H.I.; Yoon, H.W.; Jung, S.T. The highly evolvable antibody Fc domain. Trends Biotechnol. 2016, 34, 895–908. [Google Scholar] [CrossRef] [PubMed]
- Ravichandran, E.; Gong, Y.; Al Saleem, F.H.; Ancharski, D.M.; Joshi, S.G.; Simpson, L.L. An initial assessment of the systemic pharmacokinetics of botulinum toxin. J. Pharmacol. Exp. Ther. 2006, 318, 1343–1351. [Google Scholar] [CrossRef] [PubMed]
- Sepulveda, J.; Mukherjee, J.; Tzipori, S.; Simpson, L.L.; Shoemaker, C.B. Efficient serum clearance of botulinum neurotoxin achieved using a pool of small antitoxin binding agents. Infect. Immun. 2010, 78, 756–763. [Google Scholar] [CrossRef] [PubMed]
- Mukherjee, J.; Tremblay, J.M.; Leysath, C.E.; Ofori, K.; Baldwin, K.; Feng, X.; Bedenice, D.; Webb, R.P.; Wright, P.M.; Smith, L.A.; et al. A novel strategy for development of recombinant antitoxin therapeutics tested in a mouse botulism model. PLoS ONE 2012, 7, e29941. [Google Scholar] [CrossRef] [PubMed]
- Mazuet, C.; Dano, J.; Popoff, M.R.; Creminon, C.; Volland, H. Characterization of botulinum neurotoxin type A neutralizing monoclonal antibodies and influence of their half-lives on therapeutic activity. PLoS ONE 2010, 5, e12416. [Google Scholar] [CrossRef] [PubMed]
- Nowakowski, A.; Wang, C.; Powers, D.B.; Amersdorfer, P.; Smith, T.J.; Montgomery, V.A.; Sheridan, R.; Blake, R.; Smith, L.A.; Marks, J.D. Potent neutralization of botulinum neurotoxin by recombinant oligoclonal antibody. Proc. Natl. Acad. Sci. USA 2002, 99, 11346–11350. [Google Scholar] [CrossRef] [PubMed]
- Rosen, O.; Ozeri, E.; Barnea, A.; David, A.B.; Zichel, R. Development of an innovative in vitro potency assay for anti-botulinum antitoxins. Toxins 2016, 8, 276. [Google Scholar] [CrossRef] [PubMed]
- Pincus, S.H.; Das, A.; Song, K.; Maresh, G.A.; Corti, M.; Berry, J. Role of Fc in antibody-mediated protection from ricin toxin. Toxins 2014, 6, 1512–1525. [Google Scholar] [CrossRef] [PubMed]
- Sharma, R.; Zhao, H.; Al-Saleem, F.H.; Ubaid, A.S.; Puligedda, R.D.; Segan, A.T.; Lindorfer, M.A.; Bermudez, R.; Elias, M.; Adekar, S.P.; et al. Mechanisms of enhanced neutralization of botulinum neurotoxin by monoclonal antibodies conjugated to antibodies specific for the erythrocyte complement receptor. Mol. Immunol. 2014, 57, 247–254. [Google Scholar] [CrossRef] [PubMed]
- Chow, S.K.; Smith, C.; MacCarthy, T.; Pohl, M.A.; Bergman, A.; Casadevall, A. Disease-enhancing antibodies improve the efficacy of bacterial toxin-neutralizing antibodies. Cell Host Microbe 2013, 13, 417–428. [Google Scholar] [CrossRef] [PubMed]
- Adekar, S.P.; Segan, A.T.; Chen, C.; Bermudez, R.; Elias, M.D.; Selling, B.H.; Kapadnis, B.P.; Simpson, L.L.; Simon, P.M.; Dessain, S.K. Enhanced neutralization potency of botulinum neurotoxin antibodies using a red blood cell-targeting fusion protein. PLoS ONE 2011, 6, e17491. [Google Scholar] [CrossRef] [PubMed]
- Bournazos, S.; Chow, S.K.; Abboud, N.; Casadevall, A.; Ravetch, J.V. Human IgG Fc domain engineering enhances antitoxin neutralizing antibody activity. J. Clin. Investig. 2014, 124, 725–729. [Google Scholar] [CrossRef] [PubMed]
- Smith, P.; DiLillo, D.J.; Bournazos, S.; Li, F.; Ravetch, J.V. Mouse model recapitulating human Fcgamma receptor structural and functional diversity. Proc. Natl. Acad. Sci. USA 2012, 109, 6181–6186. [Google Scholar] [CrossRef] [PubMed]
- Jones, R.G.; Alsop, T.A.; Hull, R.; Tierney, R.; Rigsby, P.; Holley, J.; Sesardic, D. Botulinum type A toxin neutralisation by specific IgG and its fragments: A comparison of mouse systemic toxicity and local flaccid paralysis assays. Toxicon 2006, 48, 246–254. [Google Scholar] [CrossRef] [PubMed]
- Derman, Y.; Selby, K.; Miethe, S.; Frenzel, A.; Liu, Y.; Rasetti-Escargueil, C.; Avril, A.; Pelat, T.; Urbain, R.; Fontayne, A.; et al. Neutralization of botulinum neurotoxin type E by a humanized antibody. Toxins 2016, 8, 257. [Google Scholar] [CrossRef] [PubMed]
- Francisco, A.M.; Arnon, S.S. Clinical mimics of infant botulism. Pediatrics 2007, 119, 826–828. [Google Scholar] [CrossRef] [PubMed]
- Rasetti-Escargueil, C.; Avril, A.; Chahboun, S.; Tierney, R.; Bak, N.; Miethe, S.; Mazuet, C.; Popoff, M.R.; Thullier, P.; Hust, M.; et al. Development of human-like scFv-Fc antibodies neutralizing botulinum toxin serotype B. mAbs 2015, 7, 1161–1177. [Google Scholar] [CrossRef] [PubMed]
- Miethe, S.; Mazuet, C.; Liu, Y.; Tierney, R.; Rasetti-Escargueil, C.; Avril, A.; Frenzel, A.; Thullier, P.; Pelat, T.; Urbain, R.; et al. Development of germline-humanized antibodies neutralizing botulinum neurotoxin a and b. PLoS ONE 2016, 11, e161446. [Google Scholar] [CrossRef] [PubMed]
- Ben David, A.; Diamant, E.; Barnea, A.; Rosen, O.; Torgeman, A.; Zichel, R. The receptor binding domain of botulinum neurotoxin serotype A (BoNT/A) inhibits BoNT/A and bont/e intoxications in vivo. Clin. Vaccine Immunol. 2013, 20, 1266–1273. [Google Scholar] [CrossRef] [PubMed]
- Ben David, A.; Torgeman, A.; Barnea, A.; Zichel, R. Expression, purification and characterization of the receptor-binding domain of botulinum neurotoxin serotype B as a vaccine candidate. Protein Expr. Purif. 2015, 110, 122–129. [Google Scholar] [CrossRef] [PubMed]
- Zichel, R.; Mimran, A.; Keren, A.; Barnea, A.; Steinberger-Levy, I.; Marcus, D.; Turgeman, A.; Reuveny, S. Efficacy of a potential trivalent vaccine based on Hc fragments of botulinum toxins A, B, and E produced in a cell-free expression system. Clin. Vaccine Immunol. 2010, 17, 784–792. [Google Scholar] [CrossRef] [PubMed]
- Binz, T.; Kurazono, H.; Wille, M.; Frevert, J.; Wernars, K.; Niemann, H. The complete sequence of botulinum neurotoxin type A and comparison with other clostridial neurotoxins. J. Biol. Chem. 1990, 265, 9153–9158. [Google Scholar] [PubMed]
- Whelan, S.M.; Elmore, M.J.; Bodsworth, N.J.; Brehm, J.K.; Atkinson, T.; Minton, N.P. Molecular cloning of the Clostridium botulinum structural gene encoding the type B neurotoxin and determination of its entire nucleotide sequence. Appl. Environ. Microbiol. 1992, 58, 2345–2354. [Google Scholar] [PubMed]
- Malizio, C.J.; Goodnough, M.C.; Johnson, E.A. Purification of Clostridium botulinum type A neurotoxin. Methods Mol. Biol. 2000, 145, 27–39. [Google Scholar] [PubMed]
- Irwin, J.O.; Cheeseman, E.A. On an approximate method of determining the median effective dose and its error, in the case of a quantal response. J. Hyg. 1939, 39, 574–580. [Google Scholar] [CrossRef] [PubMed]
- European Directorate for the Quality of Medicines and Healthcare. Botulinum antitoxin. In European Pharmacopoeia, 8th ed.; EDQM Council of Europe: Strasbourg, France, 2014; p. 1029. [Google Scholar]
- Barr, J.R.; Moura, H.; Boyer, A.E.; Woolfitt, A.R.; Kalb, S.R.; Pavlopoulos, A.; McWilliams, L.G.; Schmidt, J.G.; Martinez, R.A.; Ashley, D.L. Botulinum neurotoxin detection and differentiation by mass spectrometry. Emerg. Infect. Dis. 2005, 11, 1578–1583. [Google Scholar] [CrossRef] [PubMed]
- Kalb, S.R.; Moura, H.; Boyer, A.E.; McWilliams, L.G.; Pirkle, J.L.; Barr, J.R. The use of Endopep-MS for the detection of botulinum toxins A, B, E, and F in serum and stool samples. Anal. Biochem. 2006, 351, 84–92. [Google Scholar] [CrossRef] [PubMed]
- Rosen, O.; Feldberg, L.; Gura, S.; Zichel, R. Improved detection of botulinum type E by rational design of a new peptide substrate for endopeptidase-mass spectrometry assay. Anal. Biochem. 2014, 456, 50–52. [Google Scholar] [CrossRef] [PubMed]
- Rosen, O.; Feldberg, L.; Gura, S.; Zichel, R. A new peptide substrate for enhanced botulinum neurotoxin type B detection by endopeptidase-liquid chromatography-tandem mass spectrometry/multiple reaction monitoring assay. Anal. Biochem. 2015, 473, 7–10. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; Baudys, J.; Krilich, J.; Smith, T.J.; Barr, J.R.; Kalb, S.R. A two-stage multiplex method for quantitative analysis of botulinum neurotoxins type A, B, E, and F by MALDI-TOF mass spectrometry. Anal. Biochem. 2014, 86, 10847–10854. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
{kind=link}
| Antibody | Normalized ELISA Titer per 1 µM Antibody | Neutralizing Activity a | ||||
|---|---|---|---|---|---|---|
| IgG | F(ab′)2 | IgG/F(ab′)2 Ratio | IgG [MsLD50] | F(ab′)2 [MsLD50] | IgG/F(ab′)2 Ratio | |
| A-6 | 3.6 × 104 | 4.4 × 104 | 0.8 | 100 | <10 | >10 |
| A-2 | 2.2 × 105 | 2.2 × 105 | 1.0 | 50 | 10 | 5 |
| A-1 | 3.1 × 104 | 3.5 × 104 | 0.9 | 5 | <3 | >1.7 |
| A-1 + A-2 + A-6 MAb Combination | Expected Neutralization [MsLD50] a | Measured Neutralization [MsLD50] | Synergism b |
|---|---|---|---|
| IgGs | 15.5 | 400 | ~25 |
| F(ab′)2 | 2.3 | <10 | <4 |
| PAb Source | Neutralizing Activity in Vivo | ||
|---|---|---|---|
| IgG [IU/nmole] | F(ab′)2 [IU/nmole] | IgG/F(ab′)2 Ratio | |
| Mouse | 8 | <1 | <8.00 |
| Horse | 15 | 14 | 1.07 |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Torgeman, A.; Ozeri, E.; Ben David, A.; Diamant, E.; Rosen, O.; Schwartz, A.; Barnea, A.; Makovitzki, A.; Mimran, A.; Zichel, R. Role of Homologous Fc Fragment in the Potency and Efficacy of Anti‐Botulinum Antibody Preparations. Toxins 2017, 9, 180. https://doi.org/10.3390/toxins9060180
Torgeman A, Ozeri E, Ben David A, Diamant E, Rosen O, Schwartz A, Barnea A, Makovitzki A, Mimran A, Zichel R. Role of Homologous Fc Fragment in the Potency and Efficacy of Anti‐Botulinum Antibody Preparations. Toxins. 2017; 9(6):180. https://doi.org/10.3390/toxins9060180
Chicago/Turabian StyleTorgeman, Amram, Eyal Ozeri, Alon Ben David, Eran Diamant, Osnat Rosen, Arieh Schwartz, Ada Barnea, Arik Makovitzki, Avishai Mimran, and Ran Zichel. 2017. "Role of Homologous Fc Fragment in the Potency and Efficacy of Anti‐Botulinum Antibody Preparations" Toxins 9, no. 6: 180. https://doi.org/10.3390/toxins9060180