
Prevalence and Genetic Characteristics of Staphylococcus aureus and Staphylococcus argenteus Isolates Harboring Panton-Valentine Leukocidin, Enterotoxins, and TSST-1 Genes from Food Handlers in Myanmar

 , and
, and
Abstract
:1. Introduction
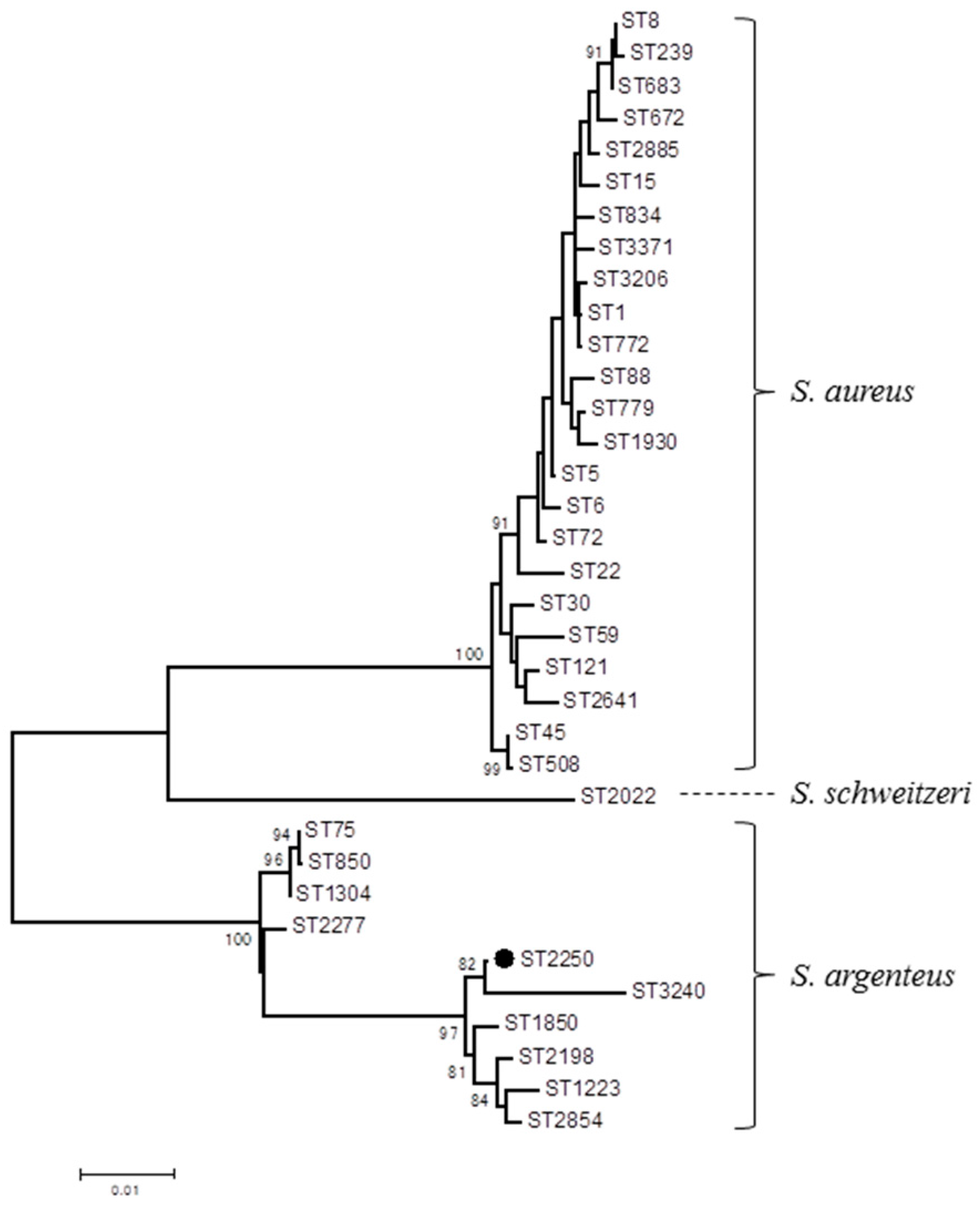
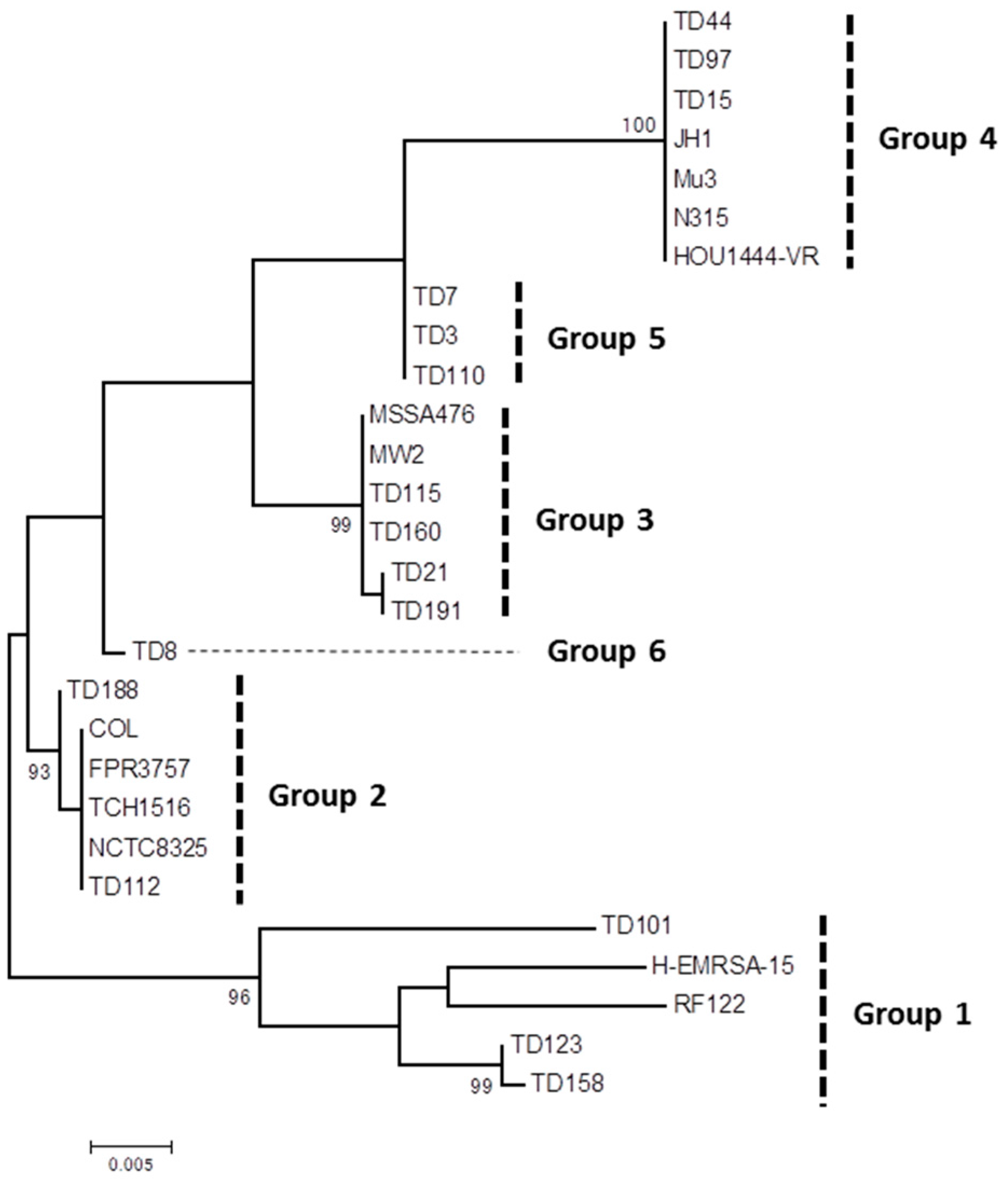
2. Results
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Study Subjects and Sample Collection, Bacterial Isolates
5.2. Antimicrobial Susceptibility Testing
5.3. Genotyping of S. aureus
5.4. Detection of SE(-Like)/TSST-1/Exfoliative Toxin Genes, Analysis of Selw
5.5. Sequence Analysis of S. argenteus
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Chambers, H.F.; Deleo, F.R. Waves of resistance: Staphylococcus aureus in the antibiotic era. Nat. Rev. Microbiol. 2009, 7, 629–641. [Google Scholar] [CrossRef] [PubMed]
- Shallcross, L.J.; Fragaszy, E.; Johnson, A.M.; Hayward, A.C. The role of the Panton-Valentine leucocidin toxin in staphylococcal disease: A systematic review and meta-analysis. Lancet Infect. Dis. 2013, 13, 43–54. [Google Scholar] [CrossRef]
- Argudín, M.Á.; Mendoza, M.C.; Rodicio, M.R. Food poisoning and Staphylococcus aureus enterotoxins. Toxins 2010, 2, 1751–1773. [Google Scholar] [CrossRef] [PubMed]
- Findlay, R.F.; Odom, R.B. Toxic shock syndrome. Int. J. Dermatol. 1982, 21, 117–121. [Google Scholar] [CrossRef] [PubMed]
- Jupin, C.; Anderson, S.; Damais, C.; Alouf, J.E.; Parant, M. Toxic shock syndrome toxin 1 as an inducer of human tumor necrosis factors and gamma interferon. J. Exp. Med. 1988, 167, 752–761. [Google Scholar] [CrossRef] [PubMed]
- Kulhankova, K.; King, J.; Salgado-Pabón, W. Staphylococcal toxic shock syndrome: Superantigen-mediated enhancement of endotoxin shock and adaptive immune suppression. Immunol. Res. 2014, 59, 182–187. [Google Scholar] [CrossRef] [PubMed]
- Ono, H.K.; Sato’o, Y.; Narita, K.; Naito, I.; Hirose, S.; Hisatsune, J.; Asano, K.; Hu, D.L.; Omoe, K.; Sugai, M.; et al. Identification and characterization of a novel staphylococcal emetic toxin. Appl. Environ. Microbiol. 2015, 81, 7034–7040. [Google Scholar] [CrossRef] [PubMed]
- Kluytmans, J.; van Belkum, A.; Verbrugh, H. Nasal carriage of Staphylococcus aureus: Epidemiology, underlying mechanisms, and associated risks. Clin. Microbiol. Rev. 1997, 10, 505–520. [Google Scholar] [PubMed]
- Van Belkum, A.; Verkaik, N.J.; de Vogel, C.P.; Boelens, H.A.; Verveer, J.; Nouwen, J.L.; Verbrugh, H.A.; Wertheim, H.F. Reclassification of Staphylococcus aureus nasal carriage types. J. Infect. Dis. 2009, 199, 1820–1826. [Google Scholar]
- Huang, S.S.; Diekema, D.J.; Warren, D.K.; Zuccotti, G.; Winokur, P.L.; Tendolkar, S.; Boyken, L.; Datta, R.; Jones, R.M.; Ward, M.A.; et al. Strain-relatedness of methicillin-resistant Staphylococcus aureus isolates recovered from patients with repeated infection. Clin. Infect. Dis. 2008, 46, 1241–1247. [Google Scholar] [CrossRef] [PubMed]
- Kluytmans, J.; van Leeuwen, W.; Goessens, W.; Hollis, R.; Messer, S.; Herwaldt, L.; Bruining, H.; Heck, M.; Rost, J.; van Leeuwen, N.; et al. Food-initiated outbreak of methicillin-resistant Staphylococcus aureus analyzed by pheno- and genotyping. J. Clin. Microbiol. 1995, 33, 1121–1128. [Google Scholar] [PubMed]
- Sivaraman, K.; Venkataraman, N.; Cole, A.M. Staphylococcus aureus nasal carriage and its contributing factors. Future Microbiol. 2009, 4, 999–1008. [Google Scholar] [CrossRef] [PubMed]
- Castro, A.; Santos, C.; Meireles, H.; Silva, J.; Teixeira, P. Food handlers as potential sources of dissemination of virulent strains of Staphylococcus aureus in the community. J. Infect. Public Health 2016, 9, 153–160. [Google Scholar] [CrossRef] [PubMed]
- Castro, A.; Komora, N.; Ferreira, V.; Lira, A.; Mota, M.; Silva, J.; Teixeira, P. Prevalence of Staphylococcus aureus from nares and hands on health care professionals in a Portuguese Hospital. J. Appl. Microbiol. 2016, 121, 831–839. [Google Scholar] [CrossRef] [PubMed]
- Sdougkos, G.; Chini, V.; Papanastasiou, D.A.; Christodoulou, G.; Stamatakis, E.; Vris, A.; Christodoulidi, I.; Protopapadakis, G.; Spiliopoulou, I. Community-associated Staphylococcus aureus infections and nasal carriage among children: molecular microbial data and clinical characteristics. Clin. Microbiol. Infect. 2008, 14, 995–1001. [Google Scholar] [CrossRef] [PubMed]
- Miller, M.B.; Weber, D.J.; Goodrich, J.S.; Popowitch, E.B.; Poe, M.D.; Nyugen, V.; Shope, T.R.; Foster, D.T.; Miller, J.R.; Kotch, J. Prevalence and risk factor analysis for methicillin-resistant Staphylococcus aureus nasal colonization in children attending child care centers. J. Clin. Microbiol. 2011, 49, 1041–1047. [Google Scholar] [CrossRef] [PubMed]
- De Boeck, H.; Vandendriessche, S.; Hallin, M.; Batoko, B.; Alworonga, J.P.; Mapendo, B.; van Geet, C.; Dauly, N.; Denis, O.; Jacobs, J. Staphylococcus aureus nasal carriage among healthcare workers in Kisangani, the Democratic Republic of the Congo. Eur. J. Clin. Microbiol. Infect. Dis. 2015, 34, 1567–1572. [Google Scholar] [CrossRef] [PubMed]
- Hogan, B.; Rakotozandrindrainy, R.; Al-Emran, H.; Dekker, D.; Hahn, A.; Jaeger, A.; Poppert, S.; Frickmann, H.; Hagen, R.M.; Micheel, V.; et al. Prevalence of nasal colonisation by methicillin-sensitive and methicillin-resistant Staphylococcus aureus among healthcare workers and students in Madagascar. BMC Infect. Dis. 2016, 16, 420. [Google Scholar] [CrossRef] [PubMed]
- Ouedraogo, A.S.; Dunyach-Remy, C.; Kissou, A.; Sanou, S.; Poda, A.; Kyelem, C.G.; Solassol, J.; Bañuls, A.L.; Van De Perre, P.; Ouédraogo, R.; et al. High Nasal Carriage Rate of Staphylococcus aureus Containing Panton-Valentine leukocidin- and EDIN-Encoding Genes in Community and Hospital Settings in Burkina Faso. Front. Microbiol. 2016, 7, 1406. [Google Scholar] [CrossRef] [PubMed]
- Aung, M.S.; Zi, H.; New, K.M.; Maw, W.W.; Aung, M.T.; Min, W.W.; Nyein, N.; Kawaguchiya, M.; Urushibara, N.; Sumi, A.; et al. Drug resistance and genetic characteristics of clinical isolates of staphylococci in Myanmar: High prevalence of PVL among methicillin-susceptible Staphylococcus aureus belonging to various sequence types. New Microbes New Infect. 2016, 10, 58–65. [Google Scholar] [CrossRef] [PubMed]
- Aung, M.S.; Urushibara, N.; Kawaguchiya, M.; Aung, T.S.; Mya, S.; San, T.; New, K.M.; Kobayashi, N. Virulence factors and genetic characteristics of methicillin-resistant and -susceptible Staphylococcus aureus isolates in Myanmar. Microb. Drug Resist. 2011, 17, 525–535. [Google Scholar] [CrossRef] [PubMed]
- Okumura, K.; Shimomura, Y.; Murayama, S.Y.; Yagi, J.; Ubukata, K.; Kirikae, T.; Miyoshi-Akiyama, T. Evolutionary paths of streptococcal and staphylococcal superantigens. BMC Genom. 2012, 13, 404. [Google Scholar] [CrossRef] [PubMed]
- Wilson, G.J.; Seo, K.S.; Cartwright, R.A.; Connelley, T.; Chuang-Smith, O.N.; Merriman, J.A.; Guinane, C.M.; Park, J.Y.; Bohach, G.A.; Schlievert, P.M.; et al. A novel core genome-encoded superantigen contributes to lethality of community-associated MRSA necrotizing pneumonia. PLoS Pathog. 2011, 7, e1002271. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tong, S.Y.; Schaumburg, F.; Ellington, M.J.; Corander, J.; Pichon, B.; Leendertz, F.; Bentley, S.D.; Parkhill, J.; Holt, D.C.; Peters, G.; et al. Novel staphylococcal species that form part of a Staphylococcus aureus-related complex: The non-pigmented Staphylococcus argenteus sp. nov. and the non-human primate-associated Staphylococcus schweitzeri sp. nov. Int. J. Syst. Evol. Microbiol. 2015, 65, 15–22. [Google Scholar] [CrossRef] [PubMed]
- Holt, D.C.; Holden, M.T.; Tong, S.Y.; Castillo-Ramirez, S.; Clarke, L.; Quail, M.A.; Currie, B.J.; Parkhill, J.; Bentley, S.D.; Feil, E.J.; et al. A very early-branching Staphylococcus aureus lineage lacking the carotenoid pigment staphyloxanthin. Genome Biol. Evol. 2011, 3, 881–895. [Google Scholar] [CrossRef] [PubMed]
- Zhang, D.F.; Xu, X.; Song, Q.; Bai, Y.; Zhang, Y.; Song, M.; Shi, C.; Shi, X. Identification of Staphylococcus argenteus in Eastern China based on a nonribosomal peptide synthetase (NRPS) gene. Future Microbiol. 2016, 11, 1113–1121. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.J.; Huang, Y.C. New epidemiology of Staphylococcus aureus infection in Asia. Clin. Microbiol. Infect. 2014, 20, 605–623. [Google Scholar] [CrossRef] [PubMed]
- Velasco, V.; Buyukcangaz, E.; Sherwood, J.S.; Stepan, R.M.; Koslofsky, R.J.; Logue, C.M. Characterization of Staphylococcus aureus from humans and a comparison with isolates of animal origin, in North Dakota, United States. PLoS ONE 2015, 10, e0140497. [Google Scholar] [CrossRef] [PubMed]
- Chen, B.; Dai, X.; He, B.; Pan, K.; Li, H.; Liu, X.; Bao, Y.; Lao, W.; Wu, X.; Yao, Y.; et al. Differences in Staphylococcus aureus nasal carriage and molecular characteristics among community residents and healthcare workers at Sun Yat-Sen University, Guangzhou, Southern China. BMC Infect. Dis. 2015, 15, 303. [Google Scholar] [CrossRef] [PubMed]
- Mégevand, C.; Gervaix, A.; Heininger, U.; Berger, C.; Aebi, C.; Vaudaux, B.; Kind, C.; Gnehm, H.P.; Hitzler, M.; Renzi, G.; et al. Paediatric Infectious Disease Group Switzerland Staphylococcus aureus Study Group. Molecular epidemiology of the nasal colonization by methicillin-susceptible Staphylococcus aureus in Swiss children. Clin. Microbiol. Infect. 2010, 16, 1414–1420. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.; Sung, J.Y.; Kim, Y.M.; Oh, C.E.; Kim, H.B.; Choi, E.H.; Lee, H.J. Molecular characterization of methicillin-resistant Staphylococcus aureus obtained from the anterior nares of healthy Korean children attending daycare centers. Int. J. Infect. Dis. 2011, 15, e558–e563. [Google Scholar] [CrossRef] [PubMed]
- Santosaningsih, D.; Santoso, S.; Budayanti, N.S.; Kuntaman, K.; Lestari, E.S.; Farida, H.; Hapsari, R.; Hadi, P.; Winarto, W.; Milheiriço, C.; et al. Epidemiology of Staphylococcus aureus harboring the mecA or Panton-Valentine leukocidin genes in hospitals in Java and Bali, Indonesia. Am. J. Trop. Med. Hyg. 2014, 90, 728–734. [Google Scholar] [CrossRef] [PubMed]
- Sarkar, A.; Raji, A.; Garaween, G.; Soge, O.; Rey-Ladino, J.; Al-Kattan, W.; Shibl, A.; Senok, A. Antimicrobial resistance and virulence markers in methicillin sensitive Staphylococcus aureus isolates associated with nasal colonization. Microb. Pathog. 2016, 93, 8–12. [Google Scholar] [CrossRef] [PubMed]
- Severin, J.A.; Lestari, E.S.; Kuntaman, K.; Melles, D.C.; Pastink, M.; Peeters, J.K.; Snijders, S.V.; Hadi, U.; Duerink, D.O.; van Belkum, A.; et al. Antimicrobial Resistance in Indonesia, Prevalence and Prevention Study Group. Unusually high prevalence of panton-valentine leukocidin genes among methicillin-sensitive Staphylococcus aureus strains carried in the Indonesian population. J. Clin. Microbiol. 2008, 46, 1989–1995. [Google Scholar] [CrossRef] [PubMed]
- Schaumburg, F.; Ngoa, U.A.; Kösters, K.; Köck, R.; Adegnika, A.A.; Kremsner, P.G.; Lell, B.; Peters, G.; Mellmann, A.; Becker, K. Virulence factors and genotypes of Staphylococcus aureus from infection and carriage in Gabon. Clin. Microbiol. Infect. 2011, 17, 1507–1513. [Google Scholar] [CrossRef] [PubMed]
- Ruimy, R.; Maiga, A.; Armand-Lefevre, L.; Maiga, I.; Diallo, A.; Koumaré, A.K.; Ouattara, K.; Soumaré, S.; Gaillard, K.; Lucet, J.C.; et al. The carriage population of Staphylococcus aureus from Mali is composed of a combination of pandemic clones and the divergent Panton-Valentine leukocidin-positive genotype ST152. J. Bacteriol. 2008, 190, 3962–3968. [Google Scholar] [CrossRef] [PubMed]
- Ayepola, O.O.; Olasupo, N.A.; Egwari, L.O.; Becker, K.; Schaumburg, F. Molecular characterization and antimicrobial susceptibility of Staphylococcus aureus isolates from clinical infection and asymptomatic carriers in southwest Nigeria. PLoS ONE 2015, 10, e0137531. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fall, C.; Richard, V.; Dufougeray, A.; Biron, A.; Seck, A.; Laurent, F.; Breurec, S. Staphylococcus aureus nasal and pharyngeal carriage in Senegal. Clin. Microbiol. Infect. 2014, 20, O239–O241. [Google Scholar] [CrossRef] [PubMed]
- Wardyn, S.E.; Forshey, B.M.; Smith, T.C. High prevalence of Panton-Valentine leukocidin among methicillin-sensitive Staphylococcus aureus colonization isolates in rural Iowa. Microb. Drug Resist. 2012, 18, 427–433. [Google Scholar] [CrossRef] [PubMed]
- Abimanyu, N.; Murugesan, S.; Krishnan, P. High prevalence of exfoliative toxins among carrier isolates of Staphylococcus aureus from healthy individuals from various communities in Chennai, South India. Indian J. Microbiol. 2013, 53, 288–290. [Google Scholar] [CrossRef] [PubMed]
- Netsvyetayeva, I.; Fraczek, M.; Piskorska, K.; Golas, M.; Sikora, M.; Mlynarczyk, A.; Swoboda-Kopec, E.; Marusza, W.; Palmieri, B.; Iannitti, T. Staphylococcus aureus nasal carriage in Ukraine: antibacterial resistance and virulence factor encoding genes. BMC Infect. Dis. 2014, 14, 128. [Google Scholar] [CrossRef] [PubMed]
- Paul, S.K.; Ghosh, S.; Kawaguchiya, M.; Urushibara, N.; Hossain, M.A.; Ahmed, S.; Mahmud, C.; Jilani, M.S.; Haq, J.A.; Ahmed, A.A.; et al. Detection and genetic characterization of PVL-positive ST8-MRSA-IVa and exfoliative toxin D-positive European CA-MRSA-Like ST1931 (CC80) MRSA-IVa strains in Bangladesh. Microb. Drug Resist. 2014, 20, 325–336. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, S.; Kawaguchiya, M.; Ghosh, S.; Paul, S.K.; Urushibara, N.; Mahmud, C.; Nahar, K.; Hossain, M.A.; Kobayashi, N. Drug resistance and molecular epidemiology of aerobic bacteria isolated from puerperal infections in Bangladesh. Microb. Drug Resist. 2015, 21, 297–306. [Google Scholar] [CrossRef] [PubMed]
- Tenover, F.C.; Goering, R.V. Methicillin-resistant Staphylococcus aureus strain USA300: Origin and epidemiology. J. Antimicrob. Chemother. 2009, 64, 441–446. [Google Scholar] [CrossRef] [PubMed]
- Ho, J.; Boost, M.; O’Donoghue, M. Prevalence of enterotoxin genes in Staphylococcus aureus colonising food handlers: does nasal carriage status matter? Eur. J. Clin. Microbiol. Infect. Dis. 2015, 34, 2177–2181. [Google Scholar] [CrossRef] [PubMed]
- Roetzer, A.; Haller, G.; Beyerly, J.; Geier, C.B.; Wolf, H.M.; Gruener, C.S.; Model, N.; Eibl, M.M. Genotypic and phenotypic analysis of clinical isolates of Staphylococcus aureus revealed production patterns and hemolytic potentials unlinked to gene profiles and source. BMC Microbiol. 2016, 16, 13. [Google Scholar] [CrossRef] [PubMed]
- Johler, S.; Sihto, H.M.; Macori, G.; Stephan, R. Sequence Variability in staphylococcal enterotoxin genes seb, sec, and sed. Toxins 2016, 8, 169. [Google Scholar] [CrossRef] [PubMed]
- Thaipadungpanit, J.; Amornchai, P.; Nickerson, E.K.; Wongsuvan, G.; Wuthiekanun, V.; Limmathurotsakul, D.; Peacock, S.J. Clinical and molecular epidemiology of Staphylococcus argenteus infections in Thailand. J. Clin. Microbiol. 2015, 53, 1005–1008. [Google Scholar] [CrossRef] [PubMed]
- Dupieux, C.; Blondé, R.; Bouchiat, C.; Meugnier, H.; Bes, M.; Laurent, S.; Vandenesch, F.; Laurent, F.; Tristan, A. Community-acquired infections due to Staphylococcus argenteus lineage isolates harbouring the Panton-Valentine leucocidin, France, 2014. Euro Surveill. 2015, 20, 21154. [Google Scholar] [CrossRef] [PubMed]
- Argudín, M.A.; Dodémont, M.; Vandendriessche, S.; Rottiers, S.; Tribes, C.; Roisin, S.; de Mendonça, R.; Nonhoff, C.; Deplano, A.; Denis, O. Low occurrence of the new species Staphylococcus argenteus in a Staphylococcus aureus collection of human isolates from Belgium. Eur. J. Clin. Microbiol. Infect. Dis. 2016, 35, 1017–1022. [Google Scholar] [CrossRef] [PubMed]
- Zhang, K.; McClure, J.A.; Elsayed, S.; Louie, T.; Conly, J.M. Novel multiplex PCR assay for simultaneous identification of community-associated methicillin-resistant Staphylococcus aureus strains USA300 and USA400 and detection of mecA and panton-valentine leukocidin genes, with discrimination of Staphylococcus aureus from coagulase-negative staphylococci. J. Clin. Microbiol. 2008, 46, 1118–1122. [Google Scholar] [PubMed]
- Hirose, M.; Kobayashi, N.; Ghosh, S.; Paul, S.K.; Shen, T.; Urushibara, N.; Kawaguchiya, M.; Shinagawa, M.; Watanabe, N. Identification of staphylocoagulase genotypes I-X and discrimination of type IV and V subtypes by multiplex PCR assay for clinical isolates of Staphylococcus aureus. Jpn. J. Infect. Dis. 2010, 63, 257–263. [Google Scholar] [PubMed]
- Strommenger, B.; Cuny, C.; Werner, G.; Witte, W. Obvious lack of association between dynamics of epidemic methicillin-resistant Staphylococcus aureus in central Europe and agr specificity groups. Eur. J. Clin. Microbiol. Infect. Dis. 2004, 23, 15–19. [Google Scholar] [PubMed]
- Enright, M.C.; Day, N.P.; Davies, C.E.; Peacock, S.J.; Spratt, B.G. Multilocus sequence typing for characterization of methicillin-resistant and methicillin-susceptible clones of Staphylococcus aureus. J. Clin. Microbiol. 2000, 38, 1008–1015. [Google Scholar] [PubMed]
- Ma, X.X.; Ito, T.; Kondo, Y.; Cho, M.; Yoshizawa, Y.; Kaneko, J.; Katai, A.; Higashiide, M.; Li, S.; Hiramatsu, K. Two different Panton-Valentine leukocidin phage lineages predominate in Japan. J. Clin. Microbiol. 2008, 46, 3246–3258. [Google Scholar] [CrossRef] [PubMed]
- Boakes, E.; Kearns, A.M.; Ganner, M.; Perry, C.; Hill, R.L.; Ellington, M.J. Distinct bacteriophages encoding Panton-Valentine leukocidin (PVL) among international methicillin-resistant Staphylococcus aureus clones harboring PVL. J. Clin. Microbiol. 2011, 49, 684–692. [Google Scholar] [CrossRef] [PubMed]
- Kawaguchiya, M.; Urushibara, N.; Kuwahara, O.; Ito, M.; Mise, K.; Kobayashi, N. Molecular characteristics of community-acquired methicillin-resistant Staphylococcus aureus in Hokkaido, northern main island of Japan: Identification of sequence types 6 and 59 Panton-Valentine leucocidin-positive community-acquired methicillin-resistant Staphylococcus aureus. Microb. Drug Resist. 2011, 17, 241–250. [Google Scholar] [PubMed]
- Molecular Evolutionary Genetics Analysis. Available online: http://www.megasoftware.net/ (accessed on 3 August 2017).



{kind=link}
{kind=link}
| Toxin Genes | coa-Genotype (No. of Isolates) | Total n = 144 (%) | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Ia (7) | IIa (3) | IIIa (15) | IVb (11) | Va (10) | Vb (9) | VIa (22) | VIc (1) | VIIa (18) | VIIb (24) | VIIIa (1) | Xa (18) | XIb (5) | ||
| lukS-PV-lukF-PV (PVL) | 0 | 0 | 2 | 0 | 1 | 0 | 12 | 0 | 2 | 0 | 0 | 0 | 1 | 18 (12.5%) |
| tst-1 (TSST-1) | 0 | 2 | 2 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 5 (3.5%) |
| Enterotoxin (-like) genes *1 | ||||||||||||||
| sea | 0 | 0 | 0 | 9 | 1 | 0 | 16 | 0 | 2 | 3 | 0 | 0 | 0 | 31 (22%) |
| seb | 0 | 1 | 5 | 0 | 2 | 0 | 1 | 0 | 0 | 10 | 0 | 1 | 0 | 20 (14%) |
| sec | 0 | 1 | 0 | 0 | 1 | 2 | 13 | 0 | 1 | 4 | 0 | 9 | 0 | 31 (22%) |
| seg | 6 | 1 | 0 | 0 | 5 | 0 | 3 | 0 | 0 | 9 | 1 | 1 | 0 | 26 (18%) |
| seh | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 2 | 1 | 0 | 2 | 0 | 5 (3%) |
| sei | 6 | 2 | 0 | 0 | 8 | 0 | 8 | 0 | 1 | 11 | 1 | 4 | 0 | 41 (28%) |
| selj | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 5 | 0 | 5 (3%) |
| sek | 0 | 0 | 3 | 1 | 1 | 0 | 0 | 0 | 4 | 11 | 0 | 0 | 0 | 20 (14%) |
| sel | 1 | 0 | 1 | 0 | 0 | 4 | 10 | 0 | 2 | 6 | 1 | 9 | 0 | 34 (24%) |
| sem | 6 | 2 | 1 | 0 | 9 | 0 | 4 | 0 | 0 | 10 | 1 | 4 | 0 | 37 (26%) |
| sen | 6 | 1 | 0 | 0 | 8 | 0 | 1 | 0 | 0 | 10 | 0 | 2 | 0 | 28 (19%) |
| seo | 6 | 2 | 0 | 0 | 8 | 0 | 3 | 0 | 0 | 9 | 1 | 1 | 0 | 30 (21%) |
| sep | 0 | 0 | 7 | 0 | 1 | 0 | 0 | 0 | 0 | 5 | 0 | 0 | 0 | 13 (9%) |
| seq | 0 | 0 | 3 | 1 | 1 | 0 | 0 | 0 | 3 | 8 | 0 | 0 | 0 | 16 (11%) |
| ser | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 4 | 0 | 4 (3%) |
| ses | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 4 | 0 | 4 (3%) |
| set | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 4 | 0 | 4 (3%) |
| selu | 6 | 1 | 0 | 0 | 5 | 0 | 3 | 0 | 0 | 6 | 1 | 1 | 0 | 23 (16%) |
| selw | 7 | 3 | 15 | 11 | 10 | 9 | 22 | 1 | 18 | 24 | 1 | 18 | 2 | 141 (98%) |
| selx | 7 | 3 | 15 | 11 | 10 | 9 | 22 | 1 | 18 | 23 | 1 | 18 | 1 | 139 (97%) |
| sely | 2 | 0 | 1 | 1 | 0 | 0 | 1 | 0 | 11 | 11 | 0 | 9 | 4 | 40 (28%) |
| Enterotoxin genes clusters *2/combinations (initials of genes) | ||||||||||||||
| g, i, m, n, o, u | 6 | 1 | 1 | 0 | 4 | 0 | 1 | 0 | 0 | 5 | 0 | 1 | 0 | 19 (13%) |
| g, i, m, n, o | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 3 | 0 | 0 | 0 | 3 (2%) |
| j, r, s, t | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 4 | 0 | 4 (3%) |
| k, q | 0 | 0 | 3 | 1 | 1 | 0 | 0 | 0 | 3 | 8 | 0 | 0 | 0 | 16 (11%) |
| Strain ID a | Age/Sex | Specimen | coa Type | ST | CC | agr Type | PVL Phage | Enterotoxin (-Like)/TSST-1 Genes b | Drug Resistance c |
|---|---|---|---|---|---|---|---|---|---|
| TD8 | 32/M | Nasal swab | IIIa | ST88 | CC88 | III | ΦPVL | selx, selw | |
| TD188 | 32/F | Nasal swab | IIIa | ST683 | CC8 | I | ΦPVL | seg, sei, sem, sen, seo, seu, sep, selx, selw | |
| TD158 | 45/F | Hand swab | Va | ST45 | CC45 | IV | ΦPVL | sea, seg, sei, sem, sen, seo, seu, selx, selw | |
| TD1 | 28/M | Nasal swab | VIa | ST1930 | CC96 | III | ΦPVL | sea, sec, sei, sel, selx, selw | |
| TD3 | 25/F | Nasal swab | VIa | ST1930 | CC96 | III | ΦSa2mw | sea, sec, sei, selx, selw | |
| TD7 | 29/M | Nasal swab | VIa | ST1930 | CC96 | III | ΦPVL | sea, sec, selx, sely, selw | |
| TD19 | 22/M | Nasal swab | VIa | ST1930 | CC96 | III | ΦPVL | sea, sec, sei, sel, selx, selw | |
| TD25 | 18/M | Nasal swab | VIa | ST1930 | CC96 | III | ΦPVL | sea, sec, sel, selx, selw | |
| TD31 | 27/M | Nasal swab | VIa | ST1930 | CC96 | III | ΦPVL | sea, sec, sel, selx, selw | |
| TD47 | 25/M | Nasal swab | VIa | ST1930 | CC96 | III | ΦPVL | sea, sec, sei, sem, selx, selw | |
| TD110 | 28/M | Nasal swab | VIa | ST1930 | CC96 | III | ΦPVL | sea, sec, sel, selx, selw | |
| TD157 | 28/M | Hand swab | VIa | ST1930 | CC96 | III | ΦPVL | sea, sec, sel, selx, selw | |
| TD149 | 24/M | Nasal swab | VIa | ST3206 | CC1 | II | ΦPVL | sea, seg, sei, sem, seo, seu, selx, selw | |
| TD161 | 23/M | Nasal swab | VIa | ST1930 | CC96 | III | ΦPVL | sea, sec, sel, selx, selw | |
| TD170 | 29/F | Nasal swab | VIa | ST1930 | CC96 | III | ΦPVL | sea, sec, sel, selx, selw | |
| TD115 | 24/F | Nasal swab | VIIa | ST1 | CC1 | III | ΦPVL | seh, sek, selx, selw | |
| TD160 | 24/F | Hand swab | VIIa | ST1 | CC1 | III | ΦPVL | sek, seq, selx, selw | |
| TD162 | 23/F | Nasal swab | XIb | ST2250 d | CC2250 | NT e | ΦPVL | sely | |
| TD97 | 27/M | Hand swab | Ia | ST672 | CC672 | I | (-) | seg, sei, sem, sen, seo, seu, selx, sely, selw | LVX, ERY |
| TD15 | 27/M | Nasal swab | IVb | ST6 | CC6 | I | (-) | sea, selx, selw | |
| TD40 | 27/M | Nasal swab | Va | ST2641 | II | (-) | sei, sem, sen, seo, selx, sew | ||
| TD101 | 32/M | Nasal swab | VIIb | ST2885 | I | (-) | seb, sek, seq, selx, sely, selw | GEN, LVX, CLI, ERY | |
| TD104 | 29/M | Hand swab | VIIb | ST59 | CC59 | I | (-) | seb, sek, sep, seq, selx, sely, selw | GEN, LVX, CLI, ERY |
| TD21 | 25/F | Nasal swab | Xa | ST15 | CC15 | II | (-) | sej, ses, set, ser, selx, selw | |
| TD44 | 28/M | Nasal swab | IIa | ST5 | CC5 | II | (-) | seg, sei, sem, sen, seo, seu, selx, selw, tst-1 | CLI, ERY |
| TD191 | 31/M | Nasal swab | IIa | ST779 | CC779 | III | (-) | seb, selx, selw, tst-1 | ERY |
| TD112 | 23/F | Nasal swab | IIIa | ST8 | CC8 | I | (-) | sek, sel, seq, selx, selw, tst-1 | |
| TD159 | 23/F | Hand swab | IIIa | ST8 | CC8 | I | (-) | sek, sem, seq, selx, selw, tst-1 | |
| TD123 | 25/M | Nasal swab | VIIb | ST508 | CC45 | I | (-) | sec, seg, sei, sel, sem, sen, seu, selx, selw, tst-1 | |
| TD20 | 22/M | Hand swab | VIa | ST1930 | CC96 | III | (-) | sea, sec, sei, sel, selx, selw | |
| TD99 | 25/F | Nasal swab | VIa | ST3371 | I | (-) | selx, selw | ||
| TD153 | 31/M | Nasal swab | VIa | ST3206 | CC1 | II | (-) | seg, sei, sem, sen, seo, seu, selx, selw | |
| TD171 | 25/M | Nasal swab | XIb | ST2250 d | NT e | (-) | sely |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Aung, M.S.; San, T.; Aye, M.M.; Mya, S.; Maw, W.W.; Zan, K.N.; Htut, W.H.W.; Kawaguchiya, M.; Urushibara, N.; Kobayashi, N. Prevalence and Genetic Characteristics of Staphylococcus aureus and Staphylococcus argenteus Isolates Harboring Panton-Valentine Leukocidin, Enterotoxins, and TSST-1 Genes from Food Handlers in Myanmar. Toxins 2017, 9, 241. https://doi.org/10.3390/toxins9080241
Aung MS, San T, Aye MM, Mya S, Maw WW, Zan KN, Htut WHW, Kawaguchiya M, Urushibara N, Kobayashi N. Prevalence and Genetic Characteristics of Staphylococcus aureus and Staphylococcus argenteus Isolates Harboring Panton-Valentine Leukocidin, Enterotoxins, and TSST-1 Genes from Food Handlers in Myanmar. Toxins. 2017; 9(8):241. https://doi.org/10.3390/toxins9080241
Chicago/Turabian StyleAung, Meiji Soe, Thida San, Mya Mya Aye, San Mya, Win Win Maw, Khin Nyein Zan, Wut Hmone Win Htut, Mitsuyo Kawaguchiya, Noriko Urushibara, and Nobumichi Kobayashi. 2017. "Prevalence and Genetic Characteristics of Staphylococcus aureus and Staphylococcus argenteus Isolates Harboring Panton-Valentine Leukocidin, Enterotoxins, and TSST-1 Genes from Food Handlers in Myanmar" Toxins 9, no. 8: 241. https://doi.org/10.3390/toxins9080241