Phosphorylation of Sox2 at Threonine 116 is a Potential Marker to Identify a Subset of Breast Cancer Cells with High Tumorigenecity and Stem-Like Features

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Lines and Reagents
2.2. Primary Breast Cancer Cells, Lentiviral Transfection and Fluorescence-Activated Cell Sorting (FACS)
2.3. Mammosphere Formation Assay and Limiting Dilution Mammosphere Assay and Luciferase Reporter Assay
2.4. Mass-Spectrometry Analysis and Database Search
2.5. Antibody Production and Purification
2.6. Immunoprecipitation, Peptide Dot Blotting and Western Blotting
2.7. Immunohistochemistry
2.8. Statistical Analysis
3. Results
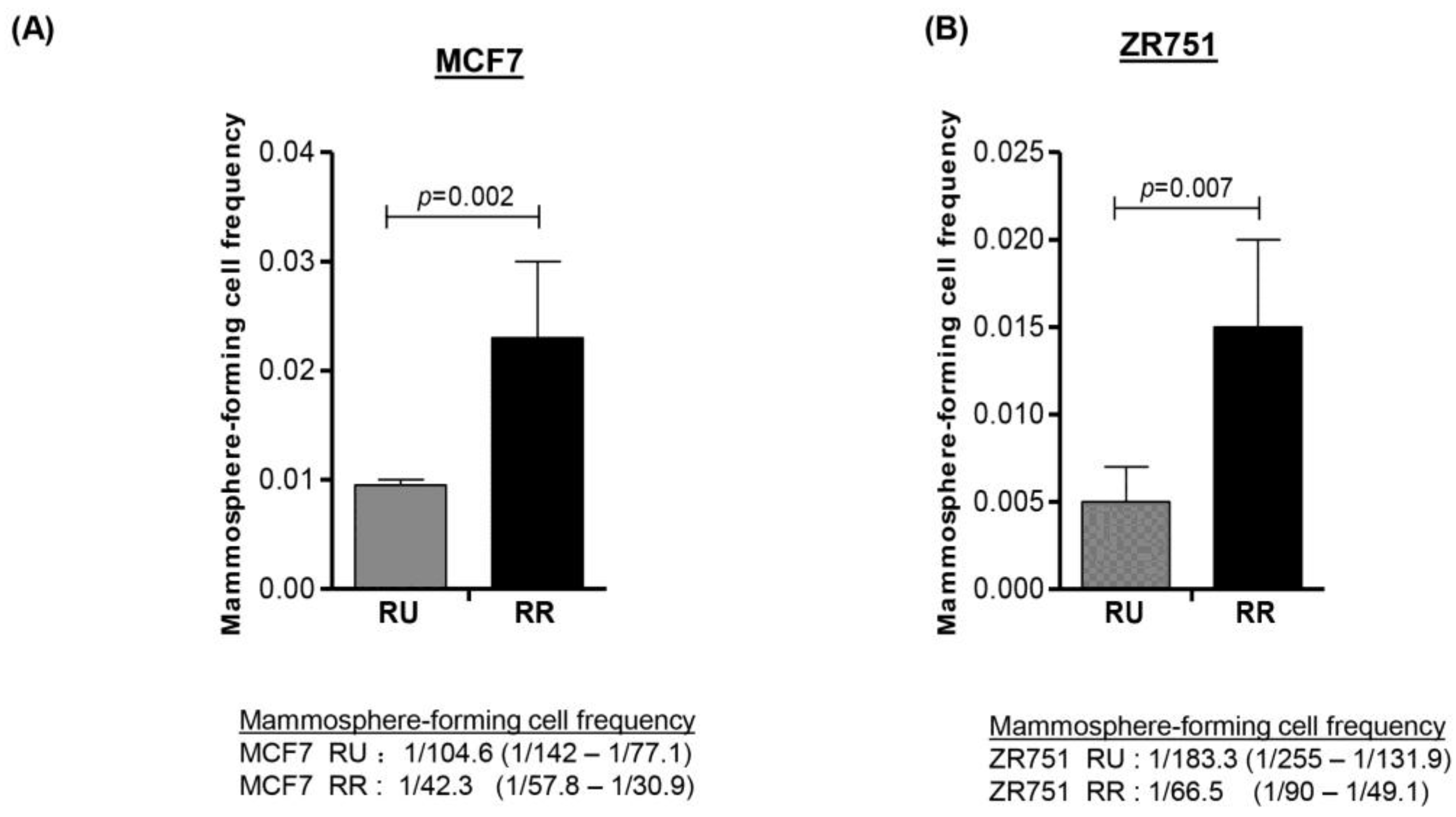
3.1. RR Cells Are More Stem-Like than RU Cells
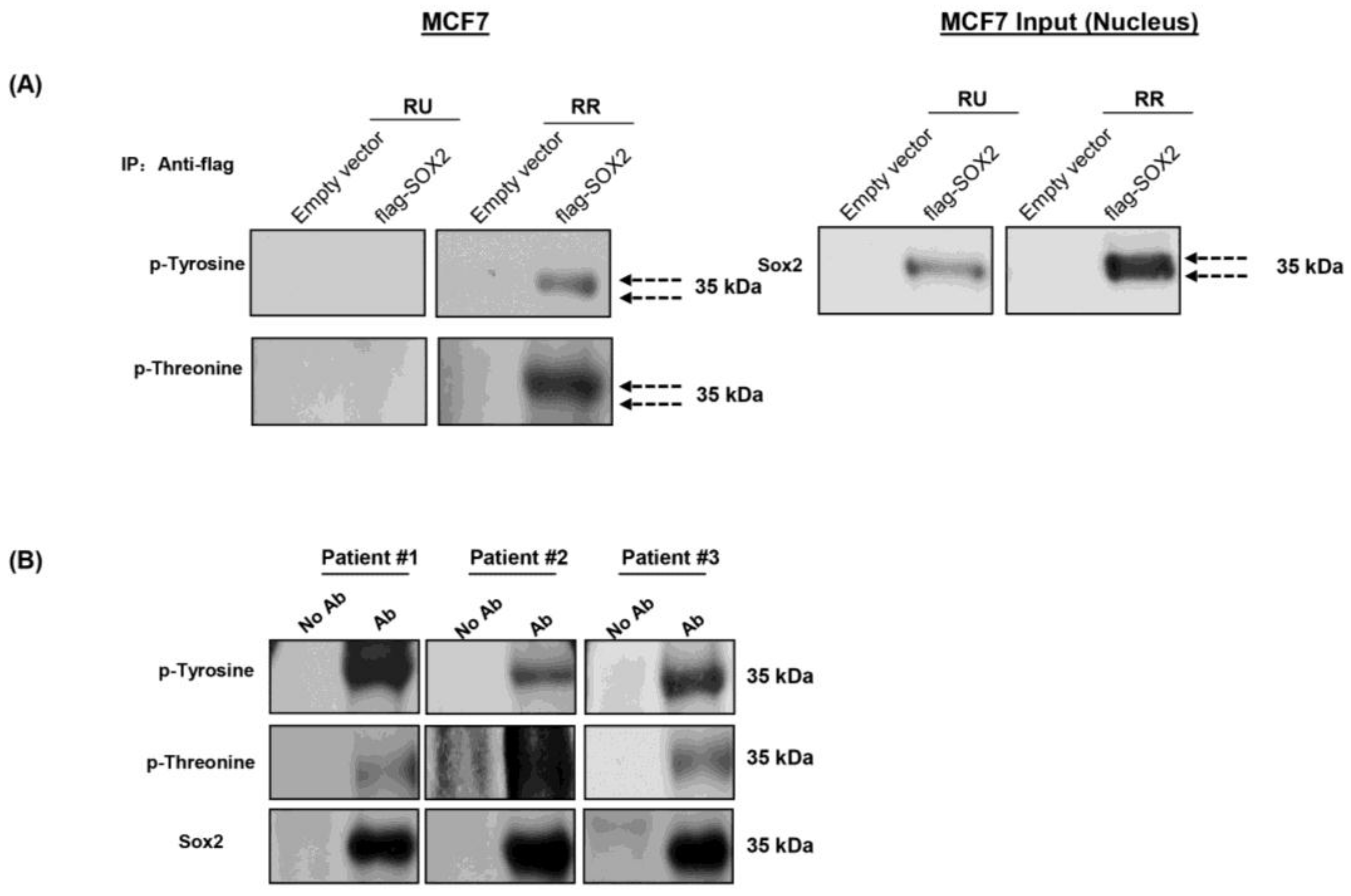
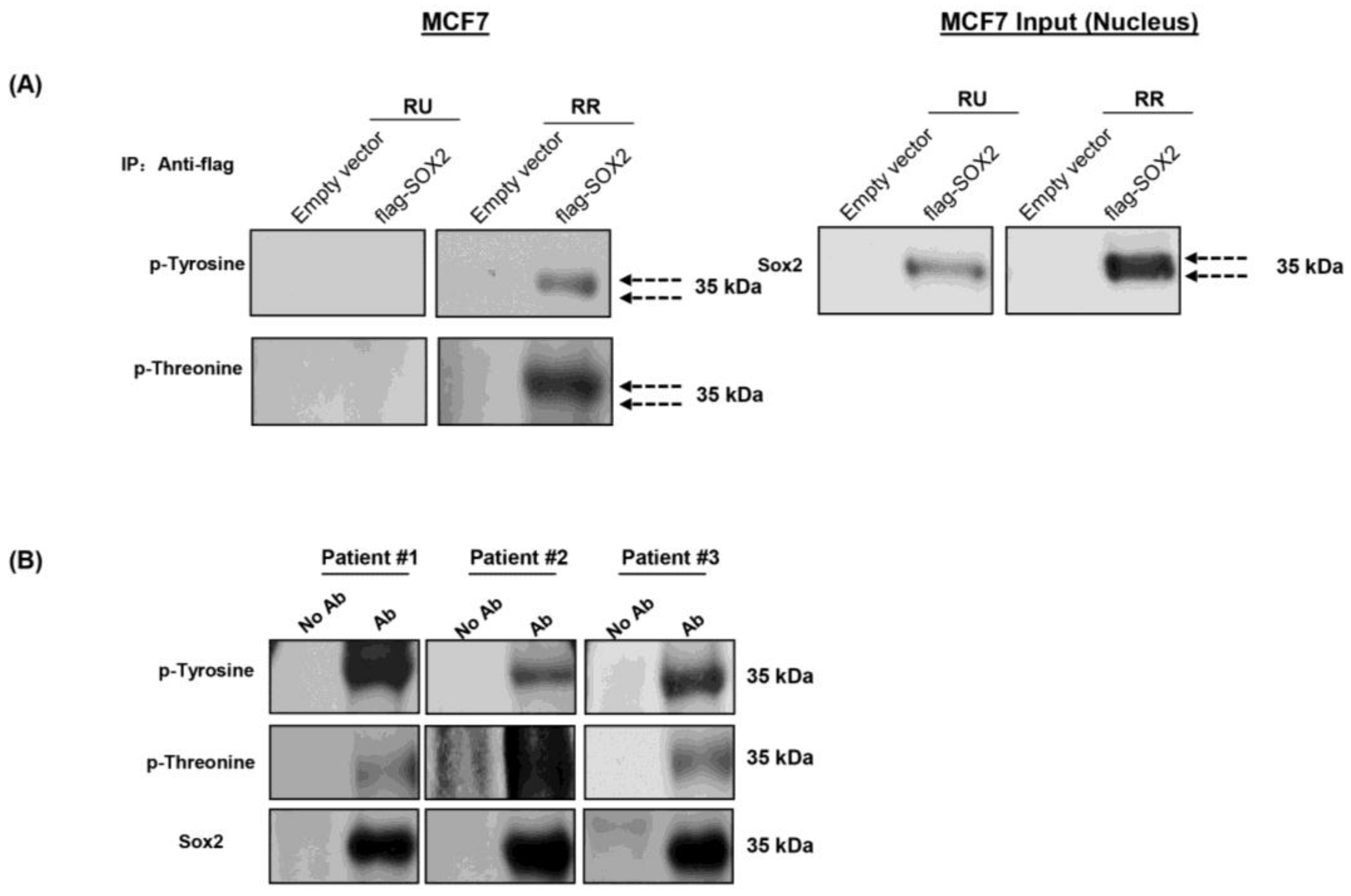
3.2. Phosphorylation of Sox2 Is Substantially Higher in RR Cells
3.3. Sox2 Is Phosphorylated at T116 in RR But Not RU Cells
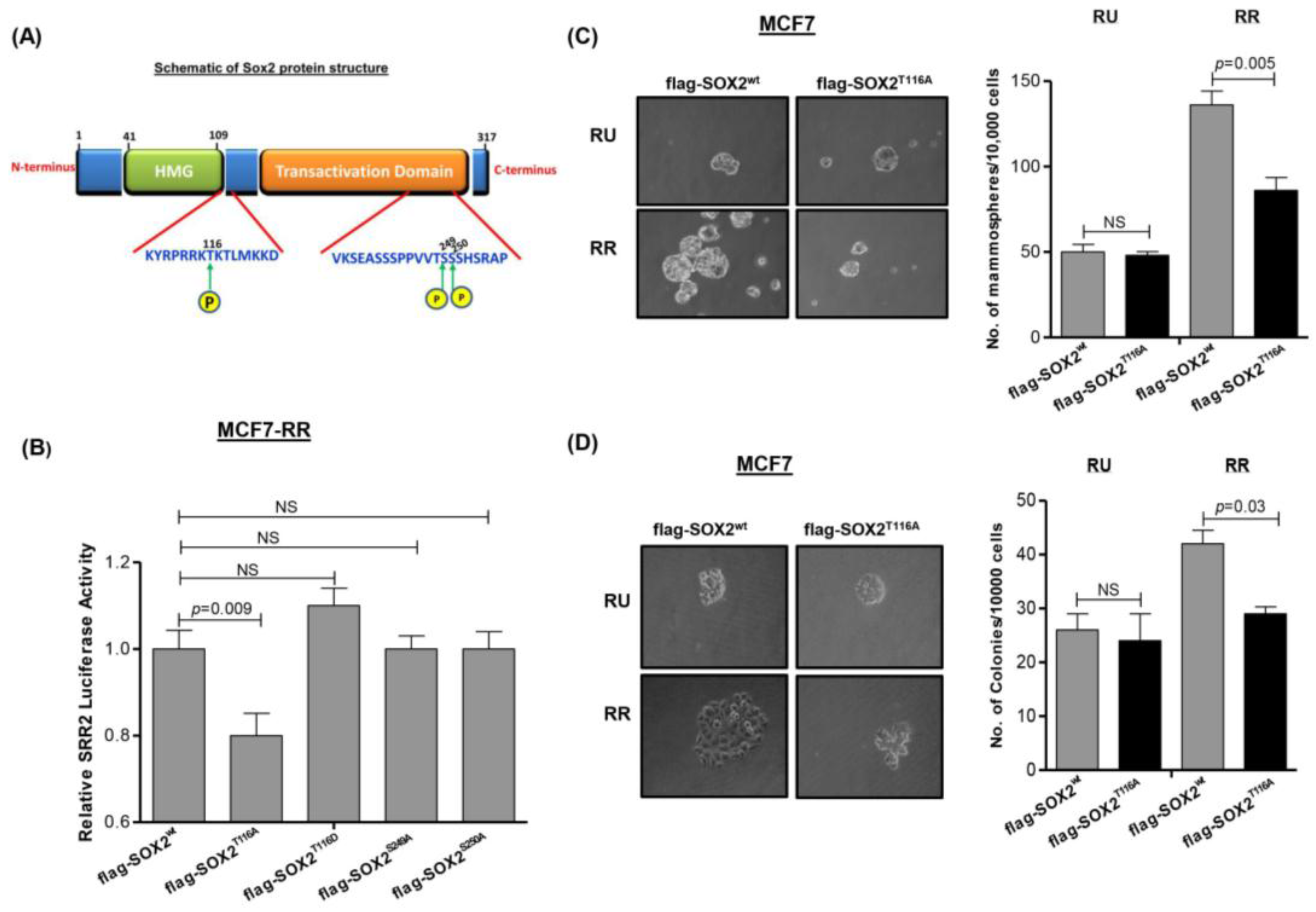
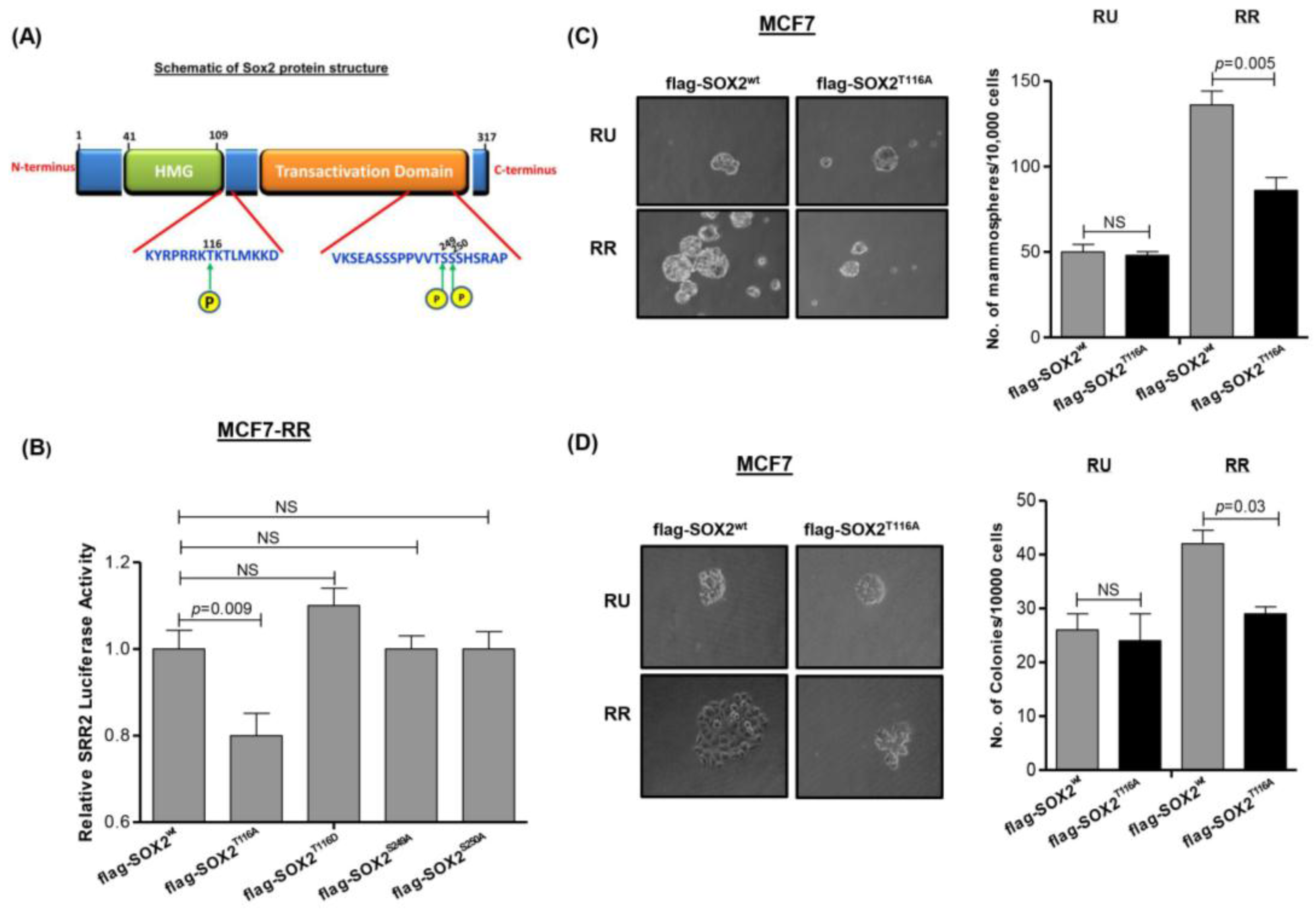
3.4. Phosphorylation of Sox2T116 Is Important in Conferring SRR2 Reporter Activity and Stem-Like Features
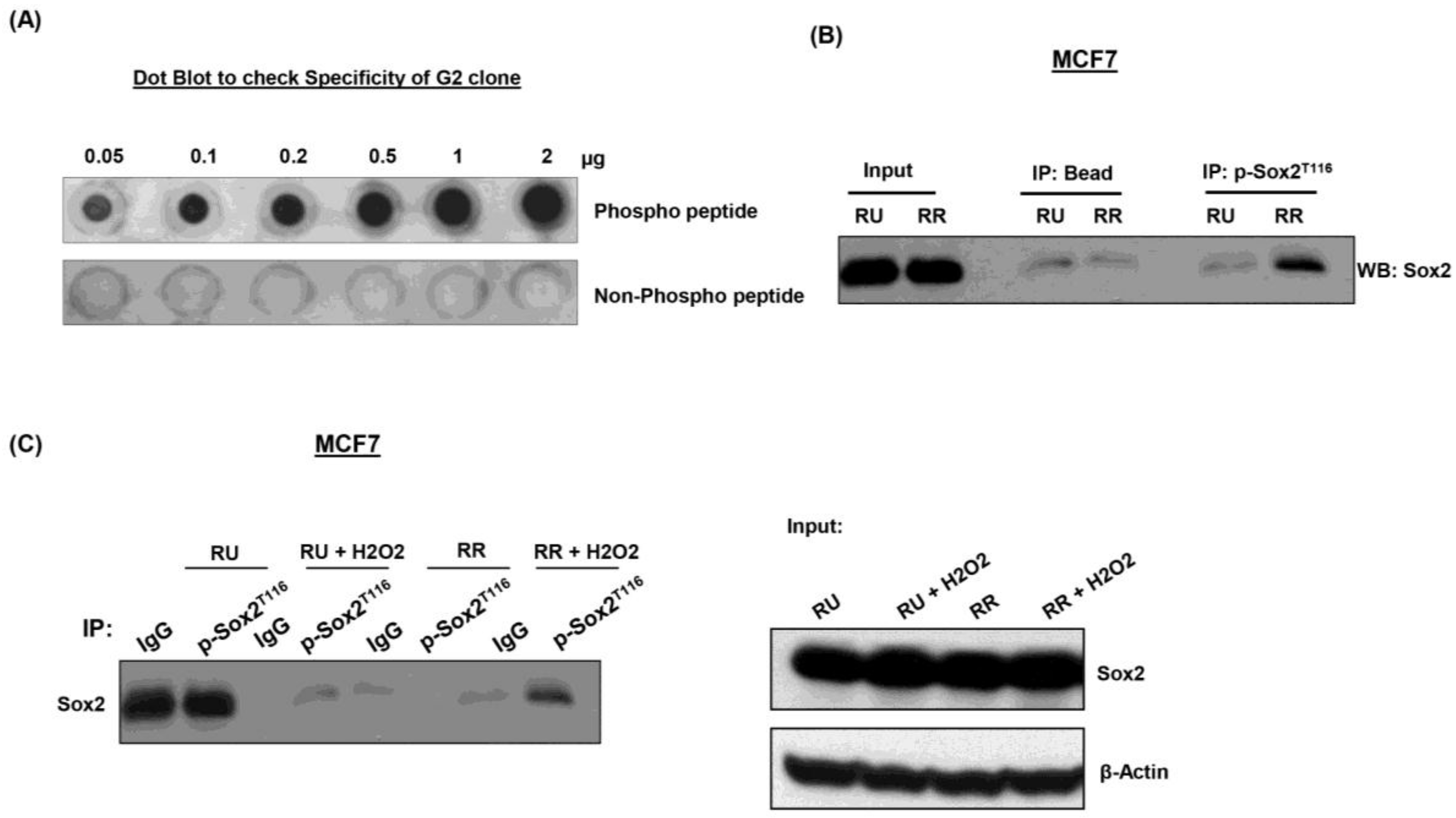
3.5. Generation and Characterization of Anti-pSox2T116 Antibody
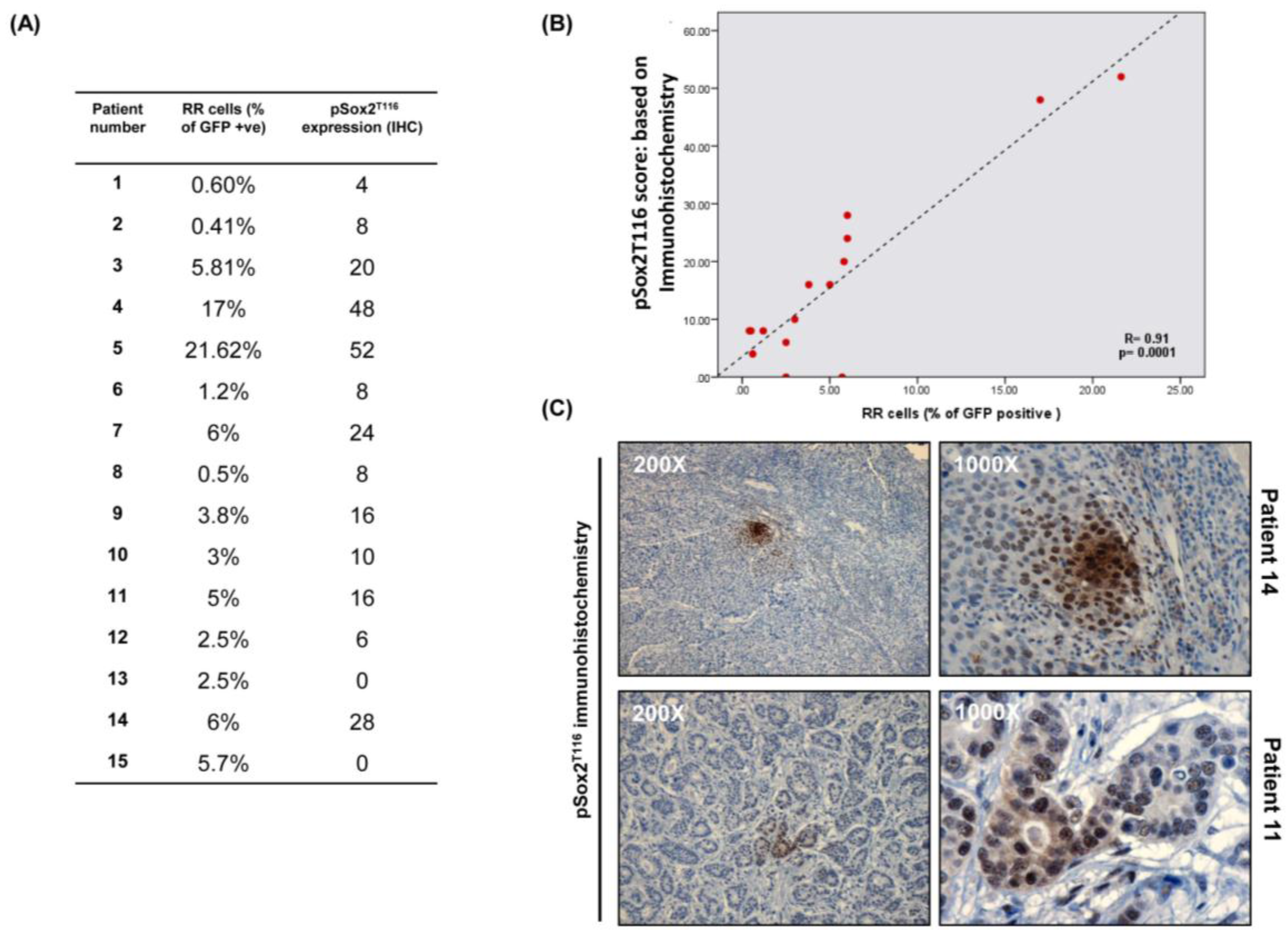
3.6. pSox2T116 Is Acquired during the RU/RR Conversion Upon Oxidative Stress
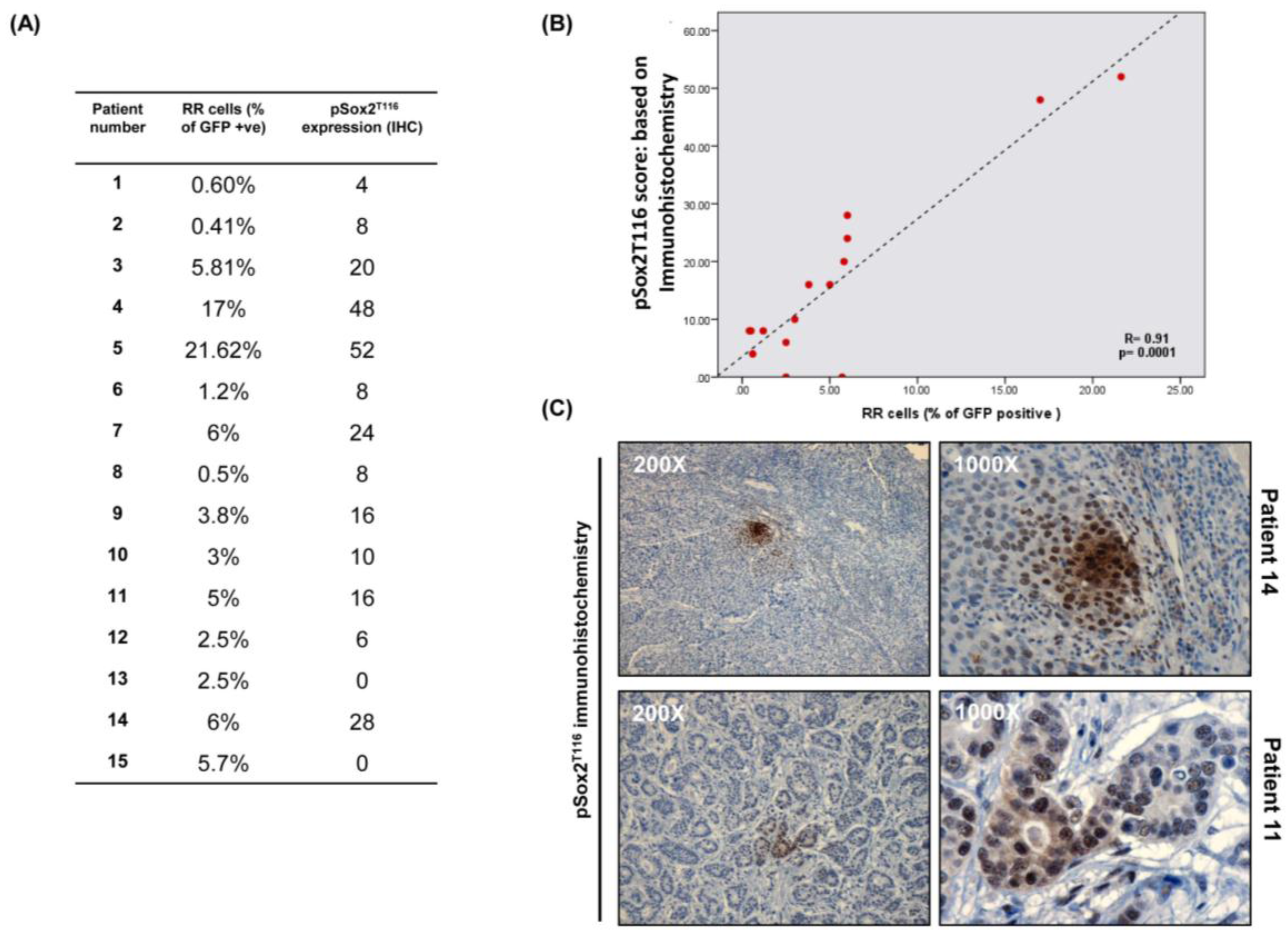
3.7. Immunohistochemical Studies of pSox2T116 in Primary BC Tumors
4. Discussion
5. Conclusions
Supplementary Materials
Acknowledgement
Author Contributions
Conflicts of Interest
References
- Yang, F.; Xu, J.; Tang, L.; Guan, X. Breast cancer stem cell: The roles and therapeutic implications. Cell. Mol. Life Sci. 2017, 74, 951–956. [Google Scholar] [CrossRef] [PubMed]
- Al-Hajj, M.; Wicha, M.S.; Benito-Hernandez, A.; Morrison, S.J.; Clarke, M.F. Prospective identification of tumorigenic breast cancer cells. Proc. Natl. Acad. Sci. USA 2003, 100, 3983–3988. [Google Scholar] [CrossRef] [PubMed]
- Singh, S.K.; Hawkins, C.; Clarke, I.D.; Squire, J.A.; Bayani, J.; Hide, T.; Henkelman, R.M.; Cusimano, M.D.; Dirks, P.B. Identification of human brain tumour initiating cells. Nature 2004, 432, 396–401. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z.F.; Ho, D.W.; Ng, M.N.; Lau, C.K.; Yu, W.C.; Ngai, P.; Chu, P.W.K.; Lam, C.T.; Poon, R.T.P.; Fan, S.T. Significance of CD90+ Cancer Stem Cells in Human Liver Cancer. Cancer Cell 2008, 13, 153–166. [Google Scholar] [CrossRef] [PubMed]
- Ginestier, C.; Hur, M.H.; Charafe-Jauffret, E.; Monville, F.; Dutcher, J.; Brown, M.; Jacquemier, J.; Viens, P.; Kleer, C.; Liu, S.; et al. ALDH1 is a marker of normal and malignant human mammary stem cells and a predictor of poor clinical outcome. Cell Stem Cell 2007, 1, 555–567. [Google Scholar] [CrossRef] [PubMed]
- Liang, S.; Furuhashi, M.; Nakane, R.; Nakazawa, S.; Goudarzi, H.; Hamada, J.-I.; Iizasa, H. Isolation and characterization of human breast cancer cells with Sox2 promoter activity. Biochem. Biophys. Res. Commun. 2013, 437, 205–211. [Google Scholar] [CrossRef] [PubMed]
- Iglesias, J.M.; Leis, O.; Pérez Ruiz, E.; Gumuzio Barrie, J.; Garcia-Garcia, F.; Aduriz, A.; Beloqui, I.; Hernandez-Garcia, S.; Lopez-Mato, M.P.; Dopazo, J.; et al. The Activation of the Sox2 RR2 Pluripotency Transcriptional Reporter in Human Breast Cancer Cell Lines is Dynamic and Labels Cells with Higher Tumorigenic Potential. Front. Oncol. 2014, 4, 308. [Google Scholar] [CrossRef] [PubMed]
- Wu, F.; Zhang, J.; Wang, P.; Ye, X.; Jung, K.; Bone, K.M.; Pearson, J.D.; Ingham, R.J.; McMullen, T.P.; Ma, Y.; et al. Identification of two novel phenotypically distinct breast cancer cell subsets based on Sox2 transcription activity. Cell. Signal. 2012, 24, 1989–1998. [Google Scholar] [CrossRef] [PubMed]
- Jung, K.; Wang, P.; Gupta, N.; Gopal, K.; Wu, F.; Ye, X.; Alshareef, A.; Bigras, G.; McMullen, T.P.; Abdulkarim, B.S.; et al. Profiling gene promoter occupancy of Sox2 in two phenotypically distinct breast cancer cell subsets using chromatin immunoprecipitation and genome-wide promoter microarrays. Breast Cancer Res. 2014, 16, 470. [Google Scholar] [CrossRef] [PubMed]
- Jung, K.; Gupta, N.; Wang, P.; Lewis, J.T.; Gopal, K.; Wu, F.; Ye, X.; Alshareef, A.; Abdulkarim, B.S.; Douglas, D.N.; et al. Triple negative breast cancers comprise a highly tumorigenic cell subpopulation detectable by its high responsiveness to a Sox2 regulatory region 2 (SRR2) reporter. Oncotarget 2015, 6, 10366–10373. [Google Scholar] [CrossRef] [PubMed]
- Gupta, N.; Jung, K.; Wu, C.; Alshareef, A.; Alqahtani, H.; Damaraju, S.; Mackey, J.R.; Ghosh, S.; Sabri, S.; Abdulkarim, B.S.; et al. High Myc expression and transcription activity underlies intra-tumoral heterogeneity in triple-negative breast cancer. Oncotarget 2017, 8, 28101–28115. [Google Scholar] [CrossRef] [PubMed]
- Calcagno, A.M.; Salcido, C.D.; Gillet, J.P.; Wu, C.P.; Fostel, J.M.; Mumau, M.D.; Gottesman, M.M.; Varticovski, L.; Ambudkar, S.V. Prolonged drug selection of breast cancer cells and enrichment of cancer stem cell characteristics. J. Natl. Cancer Inst. 2010, 102, 1637–1652. [Google Scholar] [CrossRef] [PubMed]
- Rota, L.M.; Lazzarino, D.A.; Ziegler, A.N.; LeRoith, D.; Wood, T.L. Determining mammosphere-forming potential: Application of the limiting dilution analysis. J. Mammary Gland Biol. Neoplasia 2012, 17, 119–123. [Google Scholar] [CrossRef] [PubMed]
- Hu, Y.; Smyth, G.K. ELDA: Extreme limiting dilution analysis for comparing depleted and enriched populations in stem cell and other assays. J. Immunol. Methods 2009, 347, 70–78. [Google Scholar] [CrossRef] [PubMed]
- Ye, X.; Wu, F.; Wu, C.; Wang, P.; Jung, K.; Gopal, K.; Ma, Y.; Li, L.; Lai, R. β-Catenin, a Sox2 binding partner, regulates the DNA binding and transcriptional activity of Sox2 in breast cancer cells. Cell. Signal. 2014, 26, 492–501. [Google Scholar] [CrossRef] [PubMed]
- Wu, F.; Wang, P.; Zhang, J.; Young, L.C.; Lai, R.; Li, L. Studies of Phosphoproteomic Changes Induced by Nucleophosmin-Anaplastic Lymphoma Kinase (ALK) Highlight Deregulation of Tumor Necrosis Factor (TNF)/Fas/TNF-related Apoptosis-induced Ligand Signaling Pathway in ALK-positive Anaplastic Large Cell Lymphoma. Mol. Cell. Proteom. 2010, 9, 1616–1632. [Google Scholar] [CrossRef] [PubMed]
- Wang, N.; Xie, C.; Young, J.B.; Li, L. Off-Line Two-Dimensional Liquid Chromatography with Maximized Sample Loading to Reversed-Phase Liquid Chromatography-Electrospray Ionization Tandem Mass Spectrometry for Shotgun Proteome Analysis. Anal. Chem. 2009, 81, 1049–1060. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.-S.; Rejtar, T.; Andreev, V.; Moskovets, E.; Karger, B.L. Enhanced Characterization of Complex Proteomic Samples Using LC−MALDI MS/MS: Exclusion of Redundant Peptides from MS/MS Analysis in Replicate Runs. Anal. Chem. 2005, 77, 7816–7825. [Google Scholar] [CrossRef] [PubMed]
- Alqahtani, H.; Gopal, K.; Gupta, N.; Jung, K.; Alshareef, A.; Ye, X.; Wu, F.; Li, L.; Lai, R. DDX17 (P72), a Sox2 binding partner, promotes stem-like features conferred by Sox2 in a small cell population in estrogen receptor-positive breast cancer. Cell. Signal. 2016, 28, 42–50. [Google Scholar] [CrossRef] [PubMed]
- Gelebart, P.; Anand, M.; Armanious, H.; Peters, A.C.; Dien Bard, J.; Amin, H.M.; Lai, R. Constitutive activation of the Wnt canonical pathway in mantle cell lymphoma. Blood 2008, 112, 5171–5179. [Google Scholar] [CrossRef] [PubMed]
- Gopal, K.; Gupta, N.; Zhang, H.; Alshareef, A.; Alqahtani, H.; Bigras, G.; Lewis, J.; Douglas, D.; Kneteman, N.; Lavasanifar, A.; et al. Oxidative stress induces the acquisition of cancer stem-like phenotype in breast cancer detectable by using a Sox2 regulatory region-2 (SRR2) reporter. Oncotarget 2016, 7, 3111–3127. [Google Scholar] [CrossRef] [PubMed]
- Leis, A.; Eguiara, E.; Lopez-Arribillaga, M.J.; Alberdi, S.; Hernandez-Garcia, K.; Elorriaga, A.; Pandiella, R.; Rezola, A.G. Martin, Sox2 expression in breast tumours and activation in breast cancer stem cells. Oncogene 2012, 31, 1354–1365. [Google Scholar] [CrossRef] [PubMed]
- Tian, T.; Zhang, Y.; Wang, S.; Zhou, J.; Xu, S. Sox2 enhances the tumorigenicity and chemoresistance of cancer stem-like cells derived from gastric cancer. J. Biomed. Res. 2012, 26, 336–345. [Google Scholar] [CrossRef] [PubMed]
- Bareiss, P.M.; Paczulla, A.; Wang, H.; Schairer, R.; Wiehr, S.; Kohlhofer, U.; Rothfuss, O.C.; Fischer, A.; Perner, S.; Staebler, A.; et al. Sox2 Expression Associates with Stem Cell State in Human Ovarian Carcinoma. Cancer Res. 2013, 73, 5544–5555. [Google Scholar] [CrossRef] [PubMed]
- Rybak, A.P.; Tang, D. Sox2 plays a critical role in EGFR-mediated self-renewal of human prostate cancer stem-like cells. Cell. Signal. 2013, 25, 2734–2742. [Google Scholar] [CrossRef] [PubMed]
- Xu, C.; Xie, D.; Yu, S.-C.; Yang, X.-J.; He, L.-R.; Yang, J.; Ping, Y.-F.; Wang, B.; Yang, L.; Xu, S.-L.; et al. β-Catenin/POU5F1/Sox2 Transcription Factor Complex Mediates IGF-I Receptor Signaling and Predicts Poor Prognosis in Lung Adenocarcinoma. Cancer Res. 2013, 73, 3181–3189. [Google Scholar] [CrossRef] [PubMed]
- Xiang, R.; Liao, D.; Cheng, T.; Zhou, H.; Shi, Q.; Chuang, T.S.; Markowitz, D.; Reisfeld, R.A.; Luo, Y. Downregulation of transcription factor Sox2 in cancer stem cells suppresses growth and metastasis of lung cancer. Br. J. Cancer 2011, 104, 1410–1417. [Google Scholar] [CrossRef] [PubMed]
- Gangemi, R.M.R.; Griffero, F.; Marubbi, D.; Perera, M.; Capra, M.C.; Malatesta, P.; Ravetti, G.L.; Zona, G.L.; Daga, A.; Corte, G. Sox2 Silencing in Glioblastoma Tumor-Initiating Cells Causes Stop of Proliferation and Loss of Tumorigenicity. Stem Cells 2009, 27, 40–48. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.H.; Oh, S.Y.; Do, S.I.; Lee, H.J.; Kang, H.J.; Rho, Y.S.; Bae, W.J.; Lim, Y.C. Sox2 regulates self-renewal and tumorigenicity of stem-like cells of head and neck squamous cell carcinoma. Br. J. Cancer 2014, 111, 2122–2130. [Google Scholar] [CrossRef] [PubMed]
- Finicelli, M.; Benedetti, G.; Squillaro, T.; Pistilli, B.; Marcellusi, A.; Mariani, P.; Santinelli, A.; Latini, L.; Galderisi, U.; Giordano, A. Expression of stemness genes in primary breast cancer tissues: The role of Sox2 as a prognostic marker for detection of early recurrence. Oncotarget 2014, 5, 9678–9688. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.H.; Luo, M.H.; Ni, Y.B.; Tsang, J.Y.S.; Chan, S.K.; Lui, P.C.W.; Yu, A.M.C.; Tan, P.H.; Tse, G.M. Increased Sox2 expression in less differentiated breast carcinomas and their lymph node metastases. Histopathology 2014, 64, 494–503. [Google Scholar] [CrossRef] [PubMed]
- Lengerke, C.; Fehm, T.; Kurth, R.; Neubauer, H.; Scheble, V.; Muller, F.; Schneider, F.; Petersen, K.; Wallwiener, D.; Kanz, L.; et al. Expression of the Embryonic Stem Cell Marker Sox2 in Early-Stage Breast Carcinoma. BMC Cancer 2011, 11, 42. [Google Scholar] [CrossRef] [PubMed]
- Jeong, C.H.; Cho, Y.Y.; Kim, M.O.; Kim, S.H.; Cho, E.J.; Lee, S.Y.; Jeon, Y.J.; Lee, K.Y.; Yao, K.; Keum, Y.S.; et al. Phosphorylation of Sox2 Cooperates in Reprogramming to Pluripotent Stem Cells. Stem Cells 2010, 28, 2141–2150. [Google Scholar] [CrossRef] [PubMed]
- Schaefer, T.; Wang, H.; Mir, P.; Konantz, M.; Pereboom, T.C.; Paczulla, A.M.; Merz, B.; Fehm, T.; Perner, S.; Rothfuss, O.C.; et al. Molecular and Functional Interactions between Akt and Sox2 in Breast Carcinoma. Oncotarget 2015, 6, 43540–43556. [Google Scholar] [CrossRef] [PubMed]
- Van Hoof, D.; Muñoz, J.; Braam, S.R.; Pinkse, M.W.H.; Linding, R.; Heck, A.J.R.; Mummery, C.L.; Krijgsveld, J. Phosphorylation Dynamics during Early Differentiation of Human Embryonic Stem Cells. Cell Stem Cell 2009, 5, 214–226. [Google Scholar] [CrossRef] [PubMed]
- Justilien, V.; Walsh, M.P.; Ali, S.A.; Thompson, E.A.; Murray, N.R.; Fields, A.P. The PRKCI and Sox2 Oncogenes are Co-amplified and Cooperate to Activate Hedgehog Signaling in Lung Squamous Cell Carcinoma. Cancer Cell 2014, 25, 139–151. [Google Scholar] [CrossRef] [PubMed]
- Fang, L.; Zhang, L.; Wei, W.; Jin, X.; Wang, P.; Tong, Y.; Li, J.; Du, J.X.; Wong, J. A Methylation-Phosphorylation Switch Determines Sox2 Stability and Function in Esc Maintenance or Differentiation. Mol. Cell 2014, 55, 537–551. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.C.; Cui, F.J.; Kim, Y. Hydrogen Peroxide Promotes Epithelial to Mesenchymal Transition and Stemness in Human Malignant Mesothelioma Cells. Asian Pac. J. Cancer Prev. 2013, 14, 3625–3630. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Characteristics | pSox2T116 Expression | |||
|---|---|---|---|---|
| Low Expression n (%) | High Expression n (%) | Total n | p Value | |
| Age | 0.691 | |||
| <50 | 5 (55.6%) | 4 (44.4%) | 9 | |
| ≥50 | 17 (65.4%) | 9 (34.6%) | 26 | |
| Mitotic | ||||
| 1 | 12 (75%) | 4 (25%) | 16 | 0.293 |
| 2 | 1 (33.3%) | 2 (66.7%) | 3 | |
| 3 | 9 (56.2%) | 7 (43.8%) | 16 | |
| Histological grade | ||||
| Grade 1 | 8 (72.7%) | 3 (27.3%) | 11 | 0.006 |
| Grade 2 | 7 (100%) | 0 | 7 | |
| Grade 3 | 7 (41.2%) | 10 (58.8%) | 17 | |
| Blood/Invasion | ||||
| No | 19 (86.4%) | 3 (13.6%) | 22 | 0.001 |
| Yes | 3 (23%) | 10 (76.9%) | 13 | |
| pN | ||||
| 1 | 2 (66.7%) | 1 (33.3%) | 3 | 0.332 |
| 2 | 12 (75%) | 4 (25%) | 16 | |
| 3 | 8 (50%) | 8 (50%) | 16 | |
| Architecture | ||||
| 1 | 2 (100%) | 0 | 2 | 0.371 |
| 2 | 5 (62.5%) | 3 (37.5%) | 8 | |
| 3 | 15 (60%) | 10 (40%) | 25 | |
| ER status | ||||
| Negative | 14 (82.4%) | 3 (17.6%) | 17 | 0.032 |
| Positive | 8 (44.4%) | 10 (55.6%) | 18 | |
| PR status | ||||
| Negative | 8 (61.5%) | 5 (38.5%) | 13 | 0.591 |
| Positive | 14 (63.6%) | 8 (36.4%) | 22 | |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gupta, N.; Gopal, K.; Wu, C.; Alshareef, A.; Chow, A.; Wu, F.; Wang, P.; Ye, X.; Bigras, G.; Lai, R. Phosphorylation of Sox2 at Threonine 116 is a Potential Marker to Identify a Subset of Breast Cancer Cells with High Tumorigenecity and Stem-Like Features. Cancers 2018, 10, 41. https://doi.org/10.3390/cancers10020041
Gupta N, Gopal K, Wu C, Alshareef A, Chow A, Wu F, Wang P, Ye X, Bigras G, Lai R. Phosphorylation of Sox2 at Threonine 116 is a Potential Marker to Identify a Subset of Breast Cancer Cells with High Tumorigenecity and Stem-Like Features. Cancers. 2018; 10(2):41. https://doi.org/10.3390/cancers10020041
Chicago/Turabian StyleGupta, Nidhi, Keshav Gopal, Chengsheng Wu, Abdulraheem Alshareef, Alexandra Chow, Fang Wu, Peng Wang, Xiaoxia Ye, Gilbert Bigras, and Raymond Lai. 2018. "Phosphorylation of Sox2 at Threonine 116 is a Potential Marker to Identify a Subset of Breast Cancer Cells with High Tumorigenecity and Stem-Like Features" Cancers 10, no. 2: 41. https://doi.org/10.3390/cancers10020041