Superoxide Enhances the Antitumor Combination of AdMnSOD Plus BCNU in Breast Cancer


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Culture
2.2. Reagents
2.3. Trypan Blue Dye Exclusion Assay
2.4. Adenovirus Infection
2.5. PhotofrinTM Photosensitization
2.6. Cell Homogenization and Protein Quantitation
2.7. Western Blot for MnSOD Protein
2.8. SOD Activity Gel Assay
2.9. Catalase Activity Gel Assay
2.10. GR Activity Gel Assay
2.11. GR Activity Assay
2.12. Detection of ROS by Plate Reader
2.13. Detection of ROS by Confocal Microscopy
2.14. Transfection of siRNA
2.15. RT-PCR Analysis
2.16. Statistics
3. Results
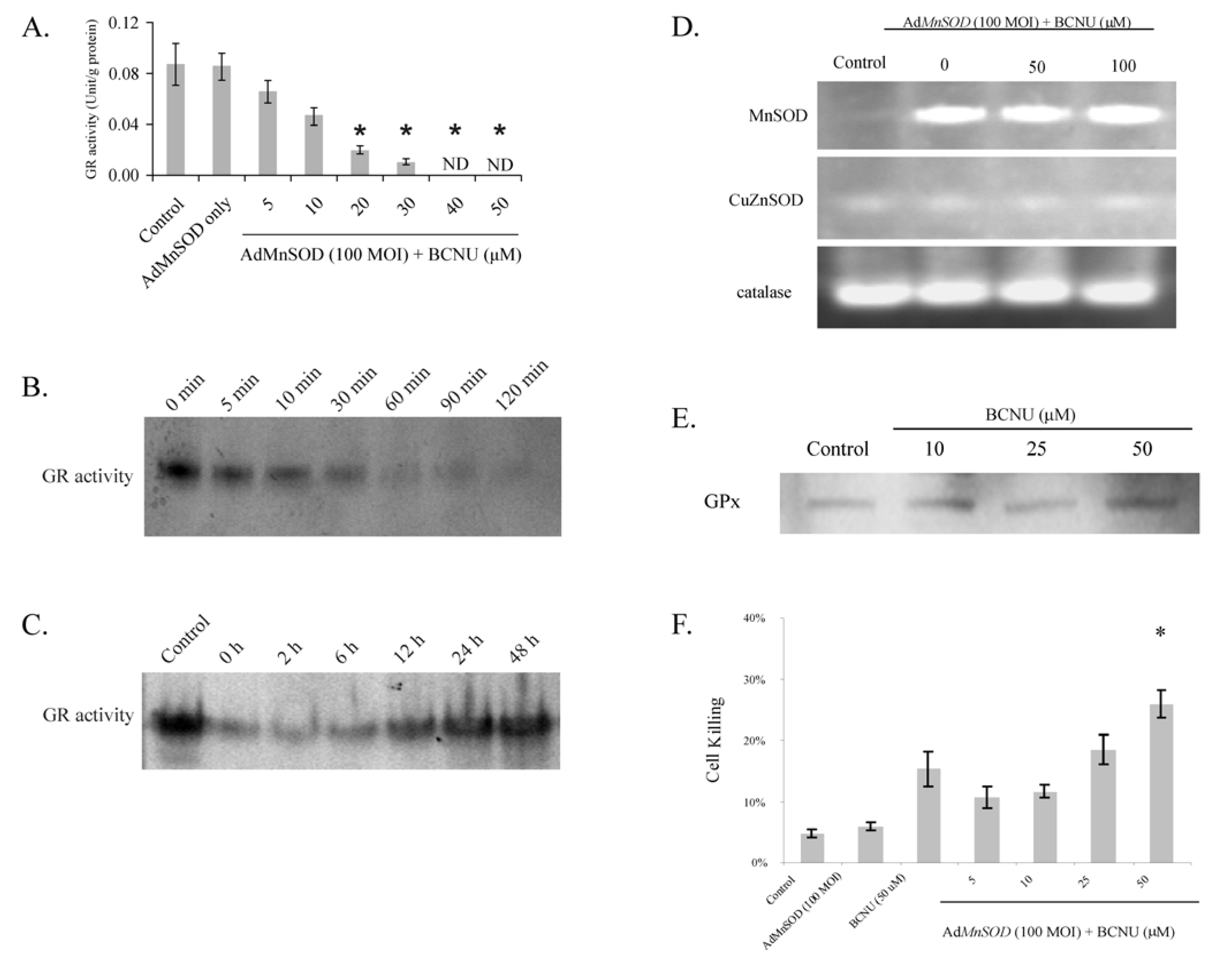
3.1. Determination of Effective BCNU Treatment Concentration

3.2. MnSOD Overexpression Sensitizes Cells to siRNA to GR-induced Cytotoxicity

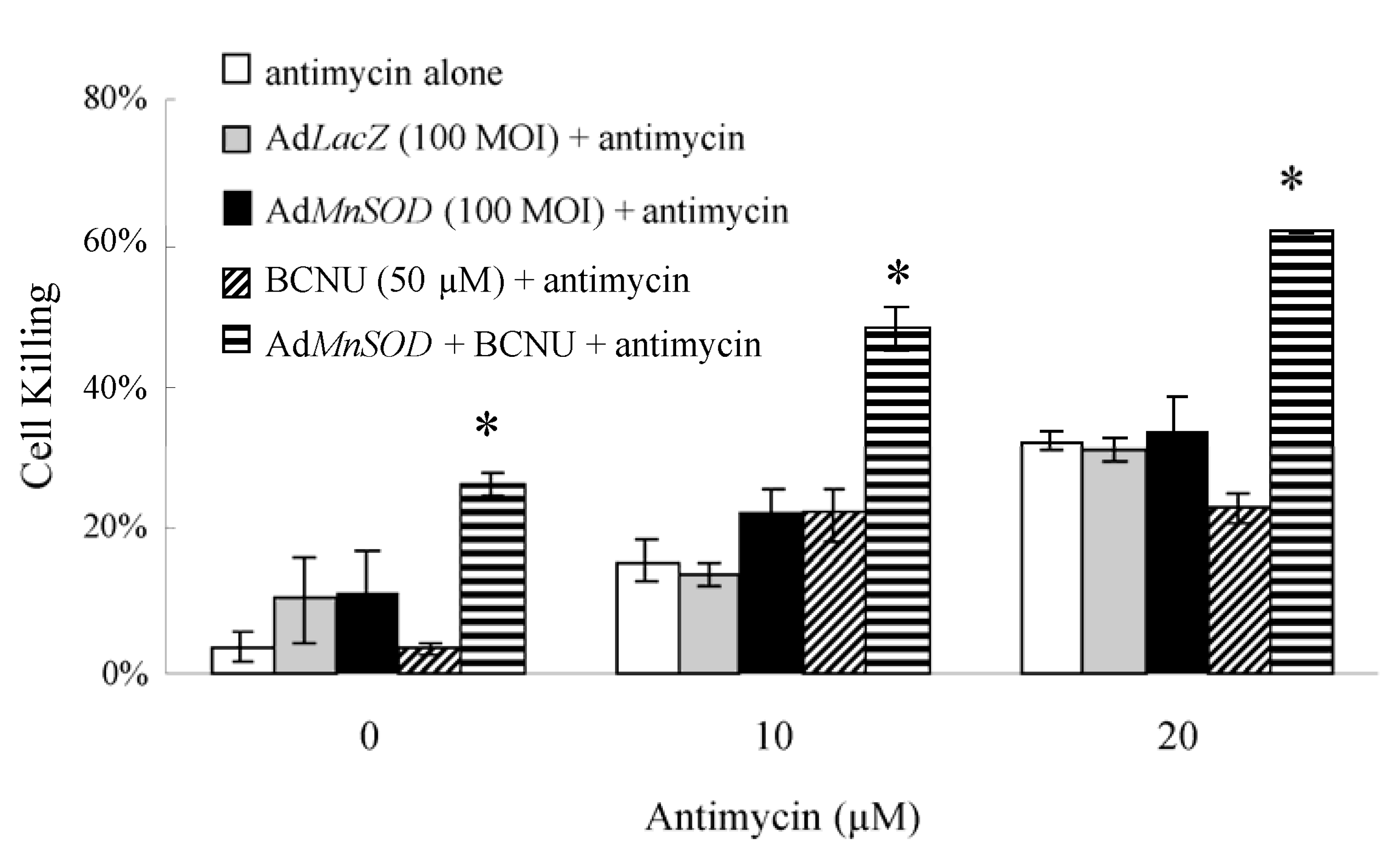
3.3. Increasing Superoxide Production Enhances Peroxide Induced Cell Toxicity: AdMnSOD Plus BCNU Sensitized Cells to Antimycin

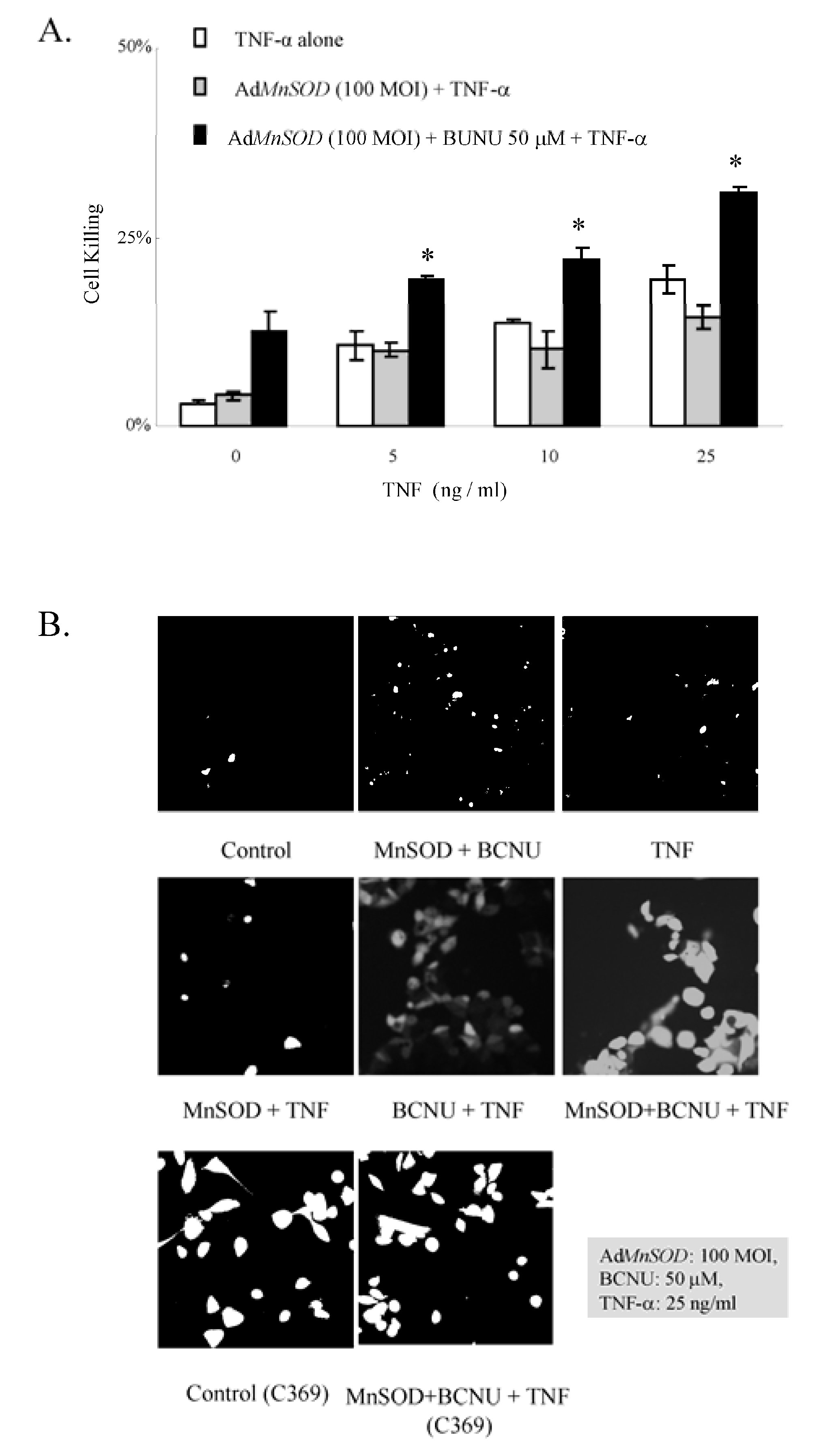
3.3.1. AdMnSOD plus BCNU Sensitized Cells to TNF-α

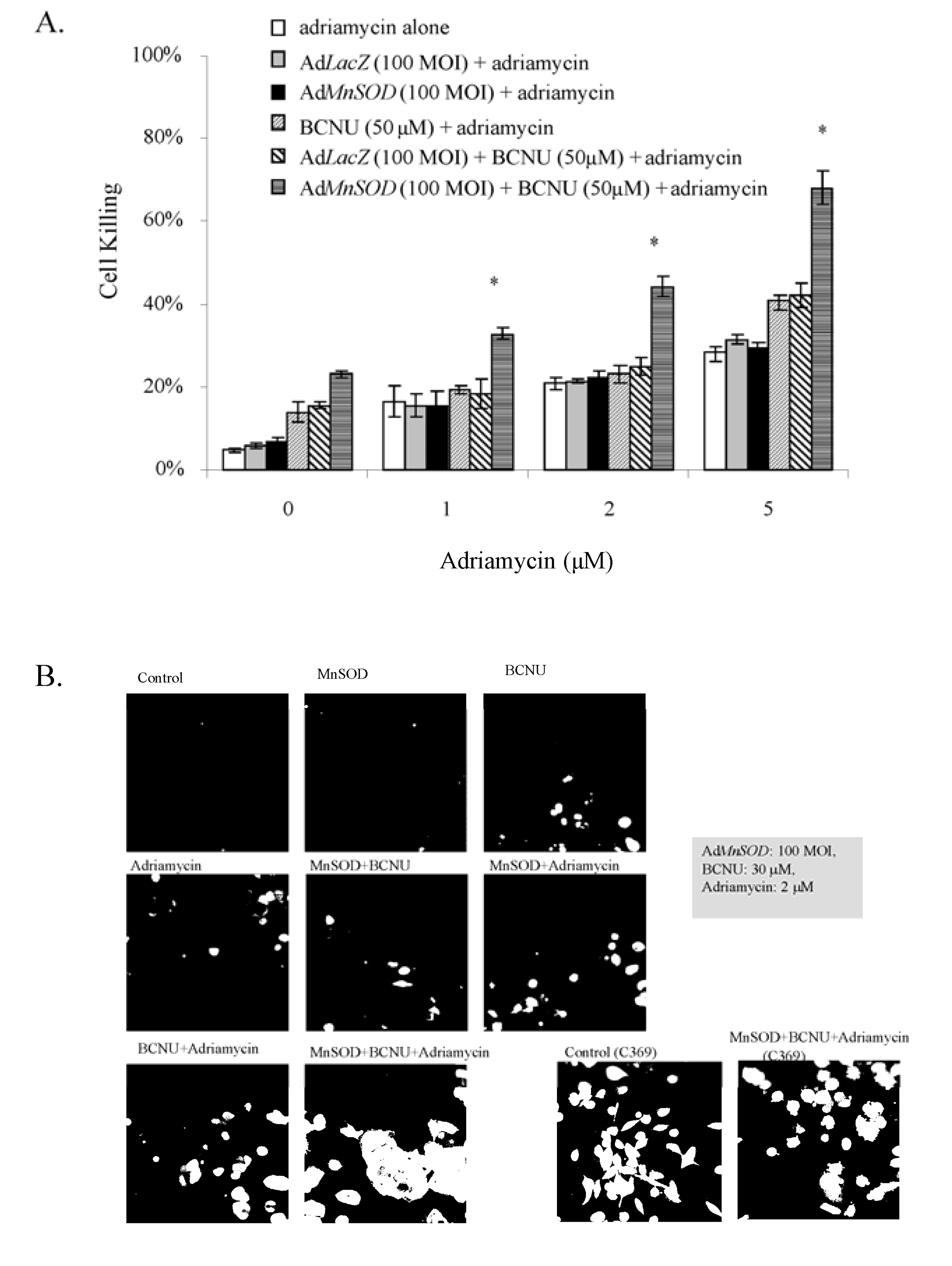
3.3.2. AdMnSOD plus BCNU Sensitized Cells to Adriamycin
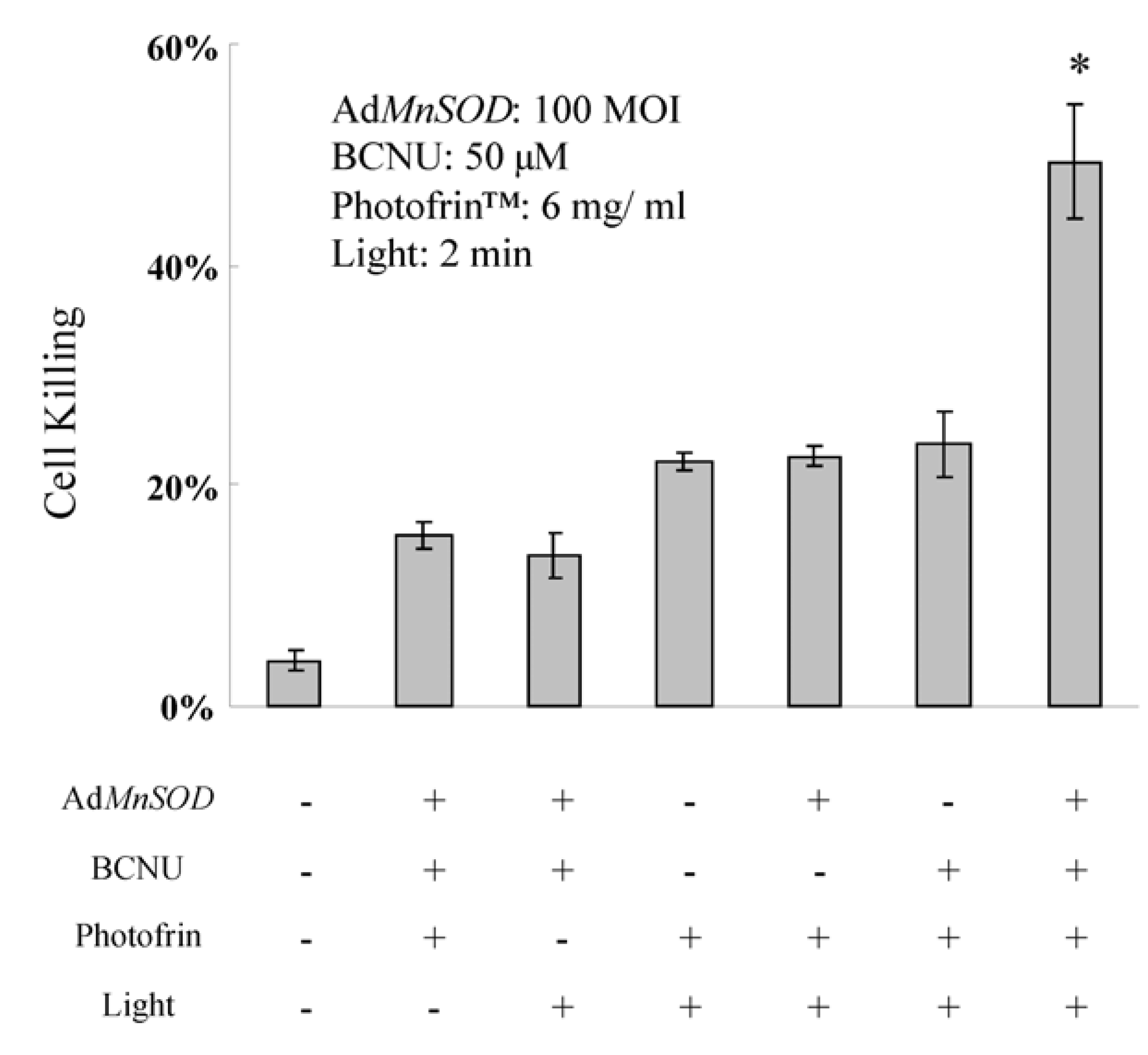
3.3.3. AdMnSOD Plus BCNU Sensitized Cells to Photodynamic Therapy (PDT)


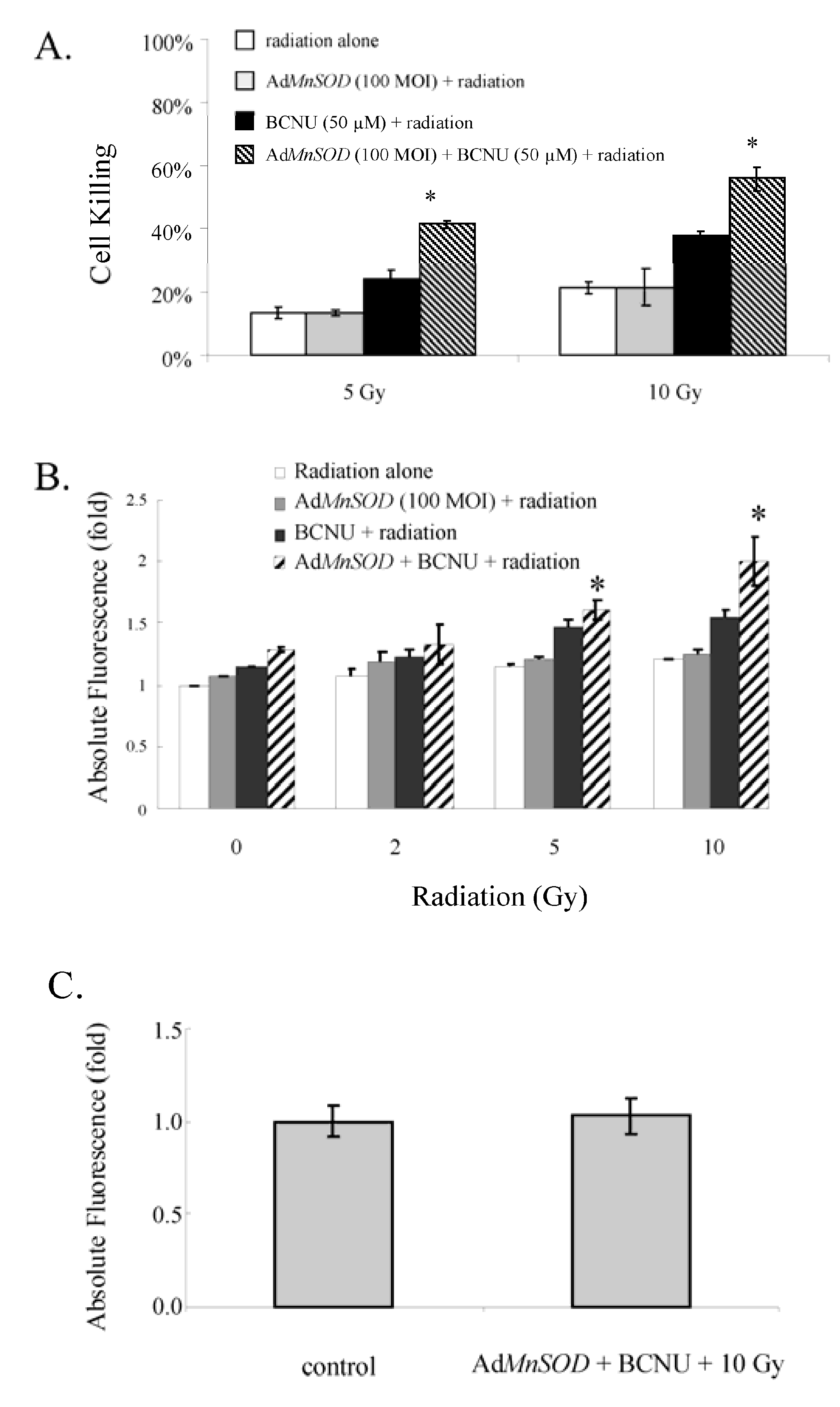
3.3.4. AdMnSOD Infection plus BCNU Sensitized Cells to Ionizing Radiation

4. Discussion
5. Conclusions
Acknowledgements
References
- Shibanuma, M.; Kuroki, T.; Nose, K. Induction of DNA replication and expression of proto-oncogenes c-myc and c-fos in quiescent Balb/3T3 cells by xanthine/xanthine oxidase. Oncogene 1988, 3, 17–21. [Google Scholar]
- Allen, R.G.; Balon, A.K. Oxidative influence in development and differentiation: and overview of a free radical theory of development. Free Radic. Biol. Med. 1989, 6, 631–661. [Google Scholar] [CrossRef]
- Sun, Y.; Oberley, L.W. Redox regulation of transcriptional activators. Free Radic. Biol. Med. 1996, 21, 335–348. [Google Scholar] [CrossRef]
- Lo, Y.Y.C.; Wong, J.M.S.; Cruz, T.F. Reactive oxygen species mediate cytokine activation of c-Jun NH2– terminal kinases. J. Biol. Chem. 1996, 271, 16586–16590. [Google Scholar] [CrossRef]
- Keyse, S.M.; Emslie, E.A. Oxidative stress and heat shock induce a human gene encoding a protein-tyrosine phosphatase. Nature 1992, 359, 664–672. [Google Scholar] [CrossRef]
- Abe, J.; Kusuhara, M.; Ulevitch, R.J.; Berk, B.B.; Lee, J.D. Big mitogen-activated protein kinase 1 (BMK1) is a redox-sensitive kinase. J. Biol. Chem. 1996, 271, 15703–15707. [Google Scholar]
- Pombo, C.M.; Bonventre, J.V.; Molnar, A.; Kyriakis, J.; Force, T. Activation of a human Ste20-like kinase by oxidant stress defines a novel stress response pathway. EMBO J. 1996, 15, 4537–4546. [Google Scholar]
- Oberley, L.W.; Buettner, G.R. Role of superoxide dismutase in cancer: a review. Cancer Res. 1979, 39, 1141–1149. [Google Scholar]
- Church, S.L.; Grant, J.W.; Ridnour, L.A.; Oberley, L.W.; Swanson, P.E.; Meltzer, P.S.; Trent, J.M. Increased manganese superoxide dismutase expression suppresses the malignant phenotype of human melanoma cells. Proc. Natl. Acad. Sci. USA 1993, 90, 3113–3117. [Google Scholar]
- Liu, R.; Oberley, T.D.; Oberley, L.W. Transfection and expression of MnSOD cDNA decreases tumor malignancy of human oral squamous carcinoma SCC-25 cells. Hum. Gene Ther. 1997, 8, 585–595. [Google Scholar] [CrossRef]
- Yan, T.; Oberley, L.W.; Zhong, W.; St. Clair, D.K. Manganese-containing superoxide dismutase overexpression causes phenotypic reversion in SV40-transformed human lung fibroblasts. Cancer Res. 1996, 56, 2864–2871. [Google Scholar]
- Zhong, W.; Oberley, L.W.; Oberley, T.D.; Clair, D.K. Suppression of the malignant phenotype of human glioma cells by overexpression of manganese superoxide dismutase. Oncogene 1997, 14, 481–490. [Google Scholar]
- Li, J.J.; Oberley, L.W.; Fan, M.; Colburn, N.H. Inhibition of AP-1 and NF-κB by manganese-containing superoxide dismutase in human breast cancer cells. FASEB J. 1998, 12, 1713–1723. [Google Scholar]
- Manna, S.K.; Zhang, H.J.; Yan, T.; Oberley, L.W.; Aggarwal, B.B. Overexpression of manganese superoxide dismutase suppresses tumor necrosis factor-induced apoptosis and activation of nuclear transcription factor–κB and activated protein-1. J. Biol. Chem. 1998, 273, 13245–13254. [Google Scholar]
- Li, J.J.; Oberley, L.W.; St. Clair, D.K.; Ridnour, L.A.; Oberley, T.D. Phenotypic changes induced in human breast cancer cells by overexpression of manganese-containing superoxide dismutase. Oncogene 1995, 10, 1989–2000. [Google Scholar]
- Rodriguez, A.M.; Carrico, P.M.; Mazurkiewicz, J.E.; Melendez, J.A. Mitochondrial or cytosolic catalase reverses the MnSOD-dependent inhibition of proliferation by enhancing respiratory chain activity, net ATP production and decreasing the steady state levels of H2O2. Free Radic. Biol. Med. 2000, 29, 801–813. [Google Scholar] [CrossRef]
- Li, S.; Yan, T.; Yang, J.Q.; Oberley, T.D.; Oberley, L.W. The role of the cellular glutathione peroxidase redox regulation in the suppression of tumor cell growth by manganese superoxide dismutase. Cancer Res. 2000, 60, 3927–3939. [Google Scholar]
- Halliwell, B.; Gutteridge, J.M.C. The chemistry of oxygen radicals and oxygen-derived species. In Free Radicals in Biology and Medicine; Clarendon Press: Oxford, England, 1985; pp. 20–66. [Google Scholar]
- Kelner, M.J.; Bagnell, R.; Montoya, M.; Estes, L.; Uglik, S.F.; Gerutti, P. Transfection with human Copper-Zine superoxide dismutase induces bi-directional alterations in other antioxidant enzymes, proteins, growth factor response, and paraquat resistance. Free Radic. Biol. Med. 1995, 18, 497–506. [Google Scholar]
- Zhong, W.; Oberley, L.W.; Oberley, T.D.; Tao, Y.; Domann, F.E.; St. Clair, D.K. Inhibition of cell growth and sensitization to oxidative damage by overexpression of manganese superoxide dismutase in rat glioma cells. Cell Growth Differ. 1996, 7, 1175–1186. [Google Scholar]
- Loo, T.L.; Dion, R.L. Colorimetric method for the determination of 1,3-1,3-bis-chloroethyl-1-nitrosourea (2-chloroethyl)-1-nitrosourea. J. Pharm. Sci. 1996, 54, 809–810. [Google Scholar]
- Arscott, L.D.; Gromer, S.; Schimer, R.H.; Becker, K.; Williams, C.H., Jr. The mechanism of thioredoxin reductase from human placenta is similar to the mechanisms of lipoamide dehydrogenase and glutathione reductase and is distinct from the mechanism of thioredoxin reductase from Escherichia coli. Proc. Natl. Acad. Sci. USA 1997, 94, 3621–3626. [Google Scholar]
- Babson, J.R.; Reed, D.J. Inactivation of glutathione reductase by 2-chloroethyl nitrosourea-derived isocyanates. Biochem. Biophys. Res. Commun. 1978, 83, 754–762. [Google Scholar] [CrossRef]
- Weydert, C.J.D.; Smith, B.B.; Xu, L.; Kregel, K.C.; Ritchie, J.M.; Davis, C.S.; Oberley, L.W. Inhibition of oral cancer cell growth by adenovirusMnSOD plus BCNU treatment. Free Radic. Biol. Med. 2003, 34, 316–329. [Google Scholar] [CrossRef]
- Zhang, H.J.; Zhao, W.; Venkataraman, S.; Robbins, M.E.; Buettner, G.R.; Kregel, K.C.; Oberley, L.W. Activation of matrix metalloproteinase-2 by overexpression of manganese superoxide dismutase in human breast cancer MCF-7 cells involves reactive oxygen species. J. Biol. Chem. 2002, 277, 20919–20926. [Google Scholar]
- Beauchammp, C.; Fridovich, I. Superoxide dismutase: improved assays and an assay applicable to acrylamide gels. Anal. Biochem. 1971, 44, 276–287. [Google Scholar] [CrossRef]
- Sun, Y.; Elwell, J.H.; Oberley, L.W. A simultaneous visualization of the antioxidant enzymes glutathione peroxidase and catalase on polyacrylamide gels. Free Radic. Res. Commun. 1988, 5, 57–67. [Google Scholar]
- Mavis, R.D.; Stellwagen, E. Purification and subunit structure of glutathione reductase from bakers' yeast. J. Biol. Chem. 1968, 243, 809–814. [Google Scholar]
- Zhang, H.J.; Yan, T.; Oberley, T.D.; Oberley, L.W. Comparison of effects of two polymorphic variants of manganese superoxide dismutase on human breast MCF-7 cancer cell phenotype. Cancer Res. 1999, 59, 6276–6283. [Google Scholar]
- Paradies, G.; Petrosillo, G.; Pistolese, M.; Ruggiero, F.M. The effect of reactive oxygen species generated from the mitochondrial electron transport chain on the cytochrome c oxidase activity and on the cardiolipin content in bovine heart submitochondrial particles. FEBS Lett. 2000, 466, 323–326. [Google Scholar] [CrossRef]
- Schmid, D.S.; Hornung, R.; McGrath, K.M.; Paul, N.; Ruddle, N.H. Target cell DNA fragmentation is mediated by lymphotoxin and tumor necrosis factor. Lymphokine Res. 1987, 6, 195–202. [Google Scholar]
- Zimmerman, R.J.; Chan, A.; Leadon, S.A. Oxidative damage in murine tumor cells treated in vitro by recombinant human tumor necrosis factor. Cancer Res. 1989, 49, 1144–1148. [Google Scholar]
- Yamauchi, N.; Kuriyama, H.; Watanabe, N.; Neda, H.; Maeda, M.; Niitsu, Y. Intracellular hydroxyl radical production induced by recombinant human tumor necrosis factor and its implication in the killing of tumor cells in vitro. Cancer Res. 1989, 49, 1671–1675. [Google Scholar]
- Wong, G.H.; Elwell, J.H.; Oberley, L.W.; Goeddel, D.V. Manganous superoxide dismutase is essential for cellular resistance to cytotoxicity of tumor necrosis factor. Cell 1999, 58, 923–931. [Google Scholar]
- Gewirtz, D.A. A critical evaluation of the mechanisms of action proposed for the antitumor effects of the anthracycline antibiotics adriamycin and daunorubicin. Biochem. Pharm. 1999, 57, 727–741. [Google Scholar] [CrossRef]
- Kalyanaraman, B.; Morehouse, K.M.; Mason, R.P. An electron paramagnetic resonance study of the interactions between the adriamycin semi-quinone, hydrogen peroxide, iron chelators, and radical scavengers. Arch. Biochem. Biophys. 1991, 286, 164–170. [Google Scholar] [CrossRef]
- Wyss, P.; Schwarz, V.; Dobler-Girdziunaite, D.; Hornung, R.; Walt, H.; Degen, A.; Fehr, M. Photodynamic therapy of locoregional breast cancer recurrences using a chlorin-type photosensitizer. Int. J. Cancer 2001, 93, 720–724. [Google Scholar] [CrossRef]
- Allison, R.; Mang, T.; Hewson, G.; Snider, W.; Dougherty, D. Photodynamic therapy for chest wall progression from breast carcinoma is an underutilized treatment modality. Cancer 2001, 91, 1–8. [Google Scholar] [CrossRef]
- Sharman, W.M.; Allen, C.M.; van Lier, J.E. Role of activated oxygen species in photodynamic therapy. Meth. Enzymol. 2000, 319, 376–400. [Google Scholar] [CrossRef]
- Salet, C.; Moreno, G.; Ricchelli, F. Effects of Photofrin photodynamic action on mitochondrial respiration and superoxide radical generation. Free Radic. Res. 1997, 26, 201–208. [Google Scholar] [CrossRef]
- Athar, M.; Mukhtar, H.; Elmets, C.A.; Zaim, M.T.; Lloyd, J.R.; Bickers, D.R. In situ evidence for the involvement of superoxide anions in cutaneous porphyrin photosensitization. Biochem. Biophys. Res. Commun. 1988, 151, 1054–1059. [Google Scholar] [CrossRef]
- Golab, J.; Nowis, D.; Skrzycki, M.; Czeczot, H.; Baranczyk-Kuzma, A.; Wilczynski, G.M.; Makowski, M.; Mroz, P.; Kozar, K.; Kaminski, R.; Jalili, A.; Kopec’, M.; Grzela, T.; Jakobisiak, M. Antitumor Effects of Photodynamic Therapy Are Potentiated by 2-Methoxyestradiol, a Superoxide Dismutase Inhibitor. J. Biol. Chem. 2003, 278, 407–414. [Google Scholar]
- Nathan, C.F.; Cohn, ZA. Antitumor effects of hydrogen peroxide in vivo. J. Exp. Med. 1981, 154, 1539–1553. [Google Scholar] [CrossRef]
- Oberley, L.W.; St. Clair, D.K.; Autor, A.P.; Oberley, T.D. Increase in manganese superoxide dismutase activity in the mouse heart after X-irradiation. Arch. Biochem. Biophys. 1987, 254, 69–80. [Google Scholar] [CrossRef]
- Guo, H.; Epperly, M.W.; Bernarding, M.; Nie, S.; Gretton, J.; Jefferson, M.; Greenberger, J.S. Manganese superoxide dismutase-plasmid/liposome (MnSOD-PL) intratracheal gene therapy reduction of irradiation-induced inflammatory cytokines does not protect orthotopic Lewis lung carcinomas. In Vivo 2003, 17, 13–21. [Google Scholar]
- Liang, L.B.; Ma, Y.W.; Zhao, Q.Z.; Zhou, X.S.; Yang, J.; Li, Y.C.; Liu, Y.Y.; Wang, Z.; Zhang, Y.P. MnSOD gene regulated by aminopeptidase N promoter specifically protects bone marrow from radiation. Ai Zheng 2002, 21, 939–943. [Google Scholar]
- Sun, W.; Kalen, A.L.; Smith, B.J.; Cullen, J.J.; Oberley, L.W. Enhancing the antitumor activity of adriamycin and ionizing radiation. Cancer Res. 2009, 69, 4294–4300. [Google Scholar] [CrossRef]
© 2010 by the authors; Molecular Diversity Preservation International, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Sun, W.G.; Weydert, C.J.; Zhang, Y.; Yu, L.; Liu, J.; Spitz, D.R.; Cullen, J.J.; Oberley, L.W. Superoxide Enhances the Antitumor Combination of AdMnSOD Plus BCNU in Breast Cancer. Cancers 2010, 2, 68-87. https://doi.org/10.3390/cancers2010068
Sun WG, Weydert CJ, Zhang Y, Yu L, Liu J, Spitz DR, Cullen JJ, Oberley LW. Superoxide Enhances the Antitumor Combination of AdMnSOD Plus BCNU in Breast Cancer. Cancers. 2010; 2(1):68-87. https://doi.org/10.3390/cancers2010068
Chicago/Turabian StyleSun, Wenqing G., Christine J. Weydert, Yuping Zhang, Lei Yu, Jingru Liu, Douglas R. Spitz, Joseph J. Cullen, and Larry W. Oberley. 2010. "Superoxide Enhances the Antitumor Combination of AdMnSOD Plus BCNU in Breast Cancer" Cancers 2, no. 1: 68-87. https://doi.org/10.3390/cancers2010068