Identification of Insulin-Like Growth Factor-I Receptor (IGF-IR) Gene Promoter-Binding Proteins in Estrogen Receptor (ER)-Positive and ER-Depleted Breast Cancer Cells


Abstract
:1. Introduction
2. Material and Methods
2.1. Cell Cultures
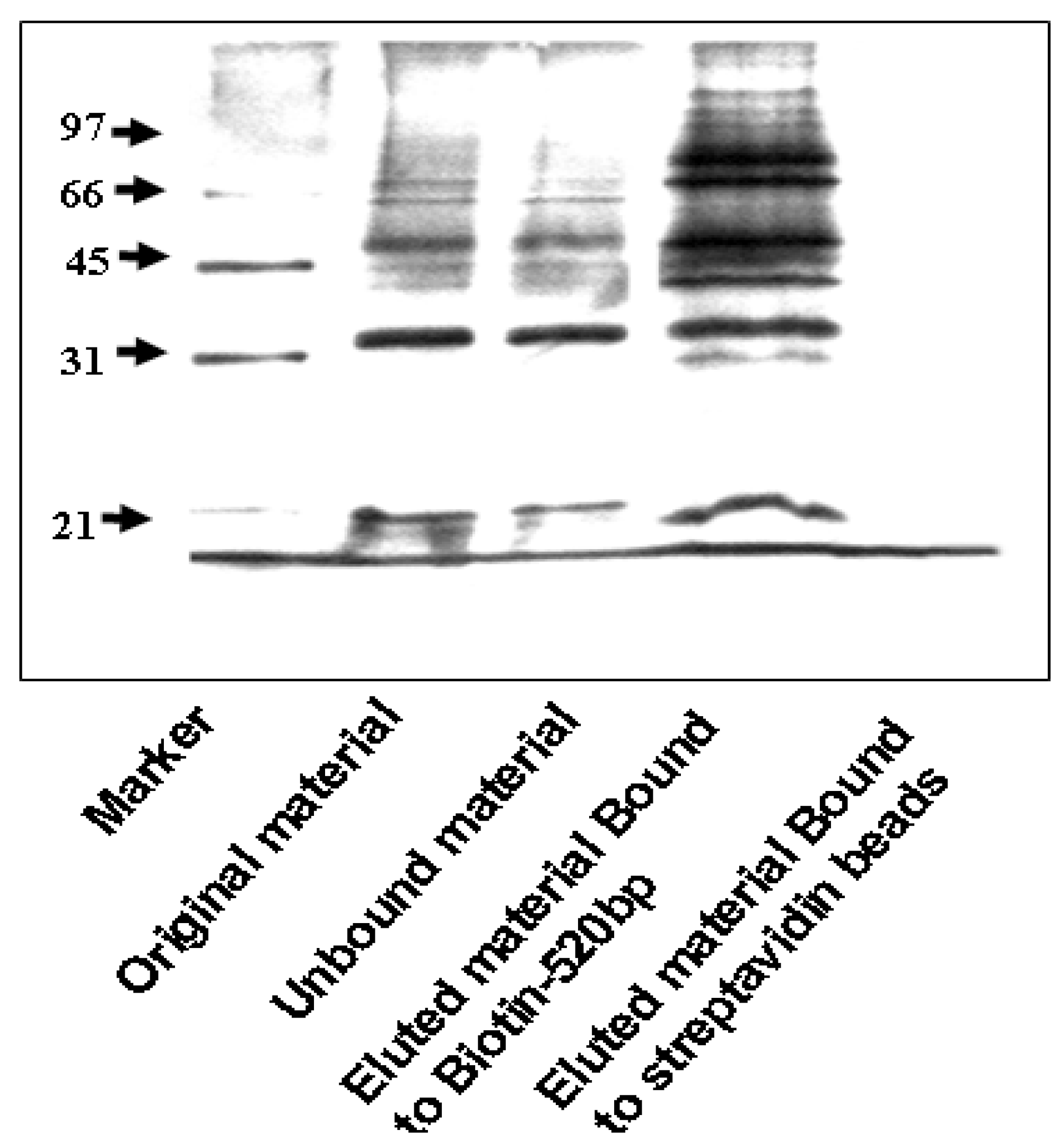
2.2. PCR and DNA Affinity Chromatography of the IGF-IR Promoter


2.3. Gel Slices Proteolysis and Mass Spectroscopy Analysis
2.4. Cell Fractionation
2.5. Western Immunoblots
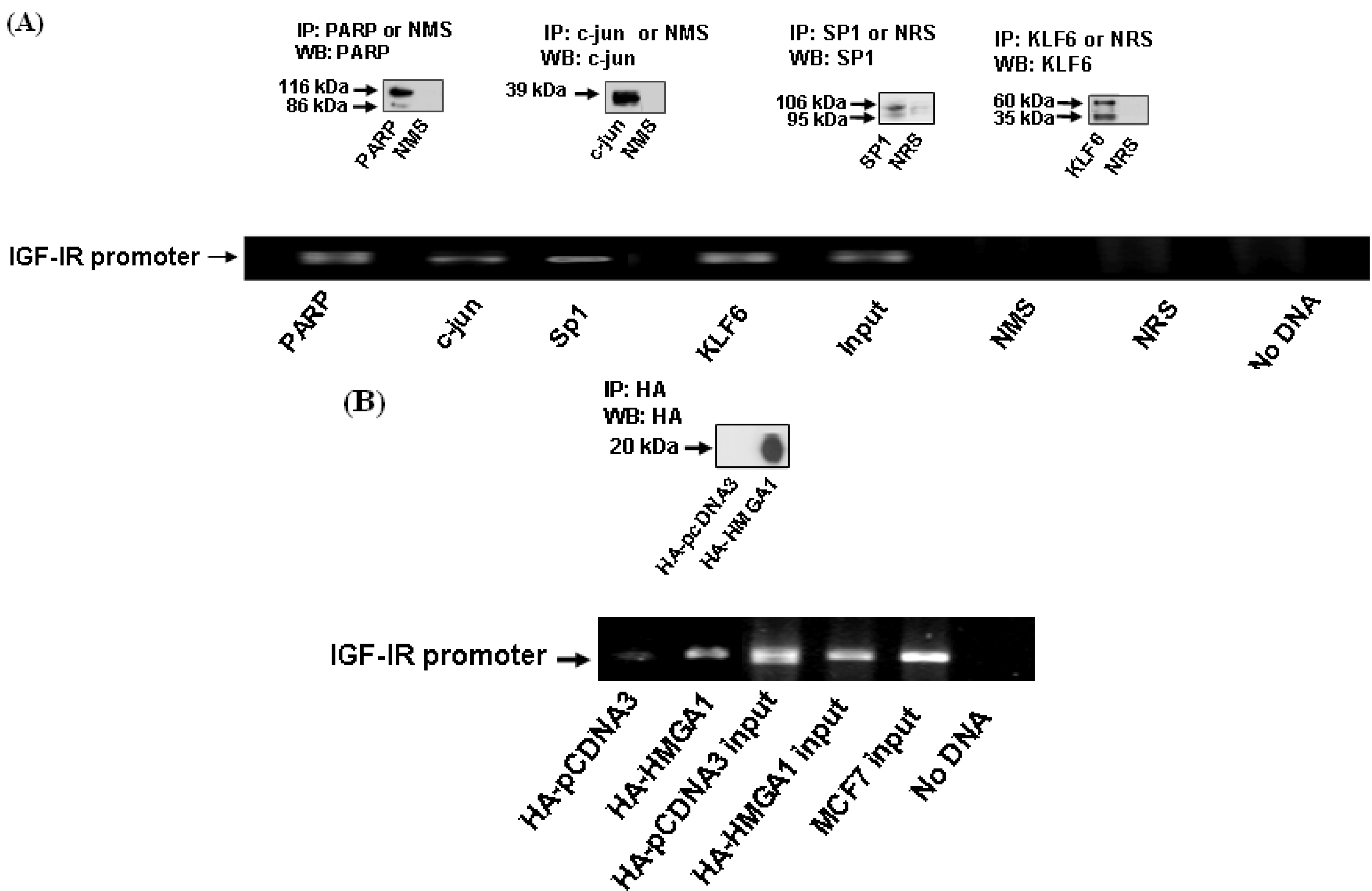
2.6. Chromatin Immunoprecipitation Assays
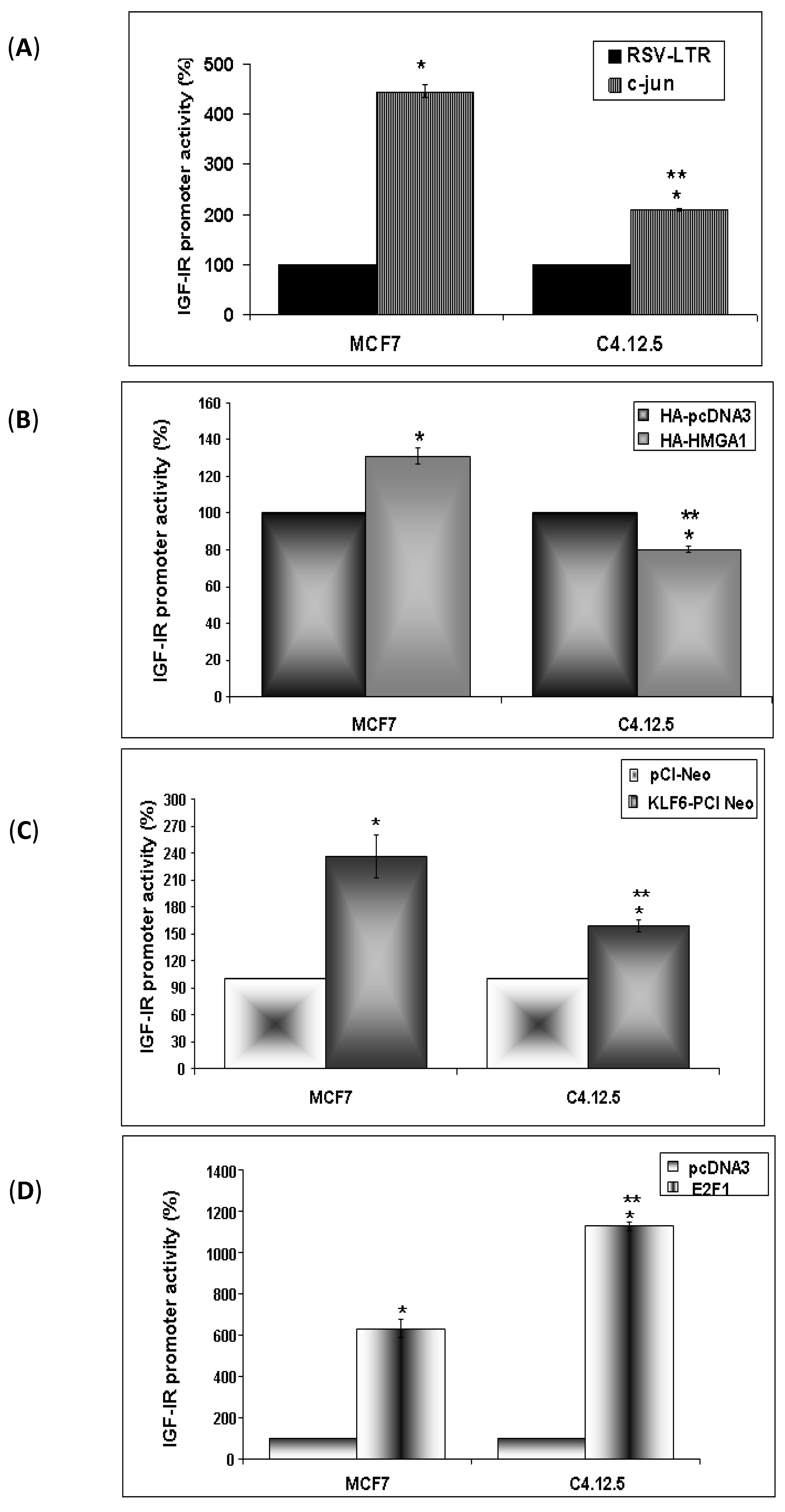
2.7. DNA Transfections
3. Results
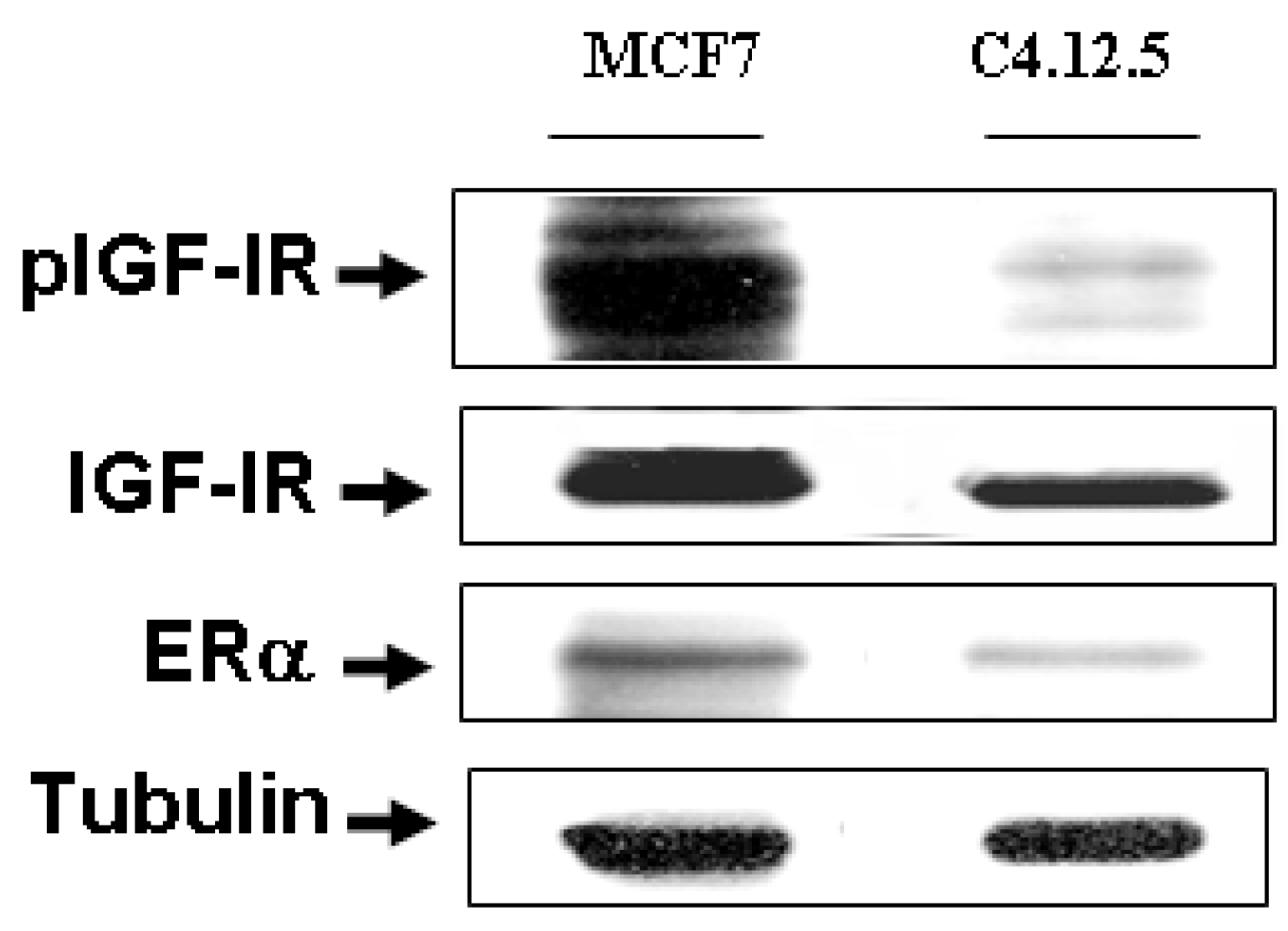
3.1. ER and IGF-IR Expression in MCF7 and C4.12.5 Cells

3.2. DNA Affinity Chromatography
3.3. MS Analysis

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Protein Name | Coverage | Coverage | Source | |
|---|---|---|---|---|
| MCF7 | C4.12.5 | |||
| 4760598 | Adenylate kinase isozyme 2 | 4 | - | Mus musculus |
| 61743954 | AHNAK nucleoprotein isoform 1 | 1 | 3 | Homo sapiens |
| 1061128 | Anpg | 25 | 21 | Homo sapiens |
| 5453541 | Anterior gradient 2 homolog | 26 | − | Homo sapiens |
| 6753086 | Apurinic/apyrimidinic endonuclease 1 | 8 | − | Mus musculus |
| 10947139 | Arginase, type I | − | 3 | Homo sapiens |
| 6680748 | ATP synthase, H+ transporting, mitochondrial F1 complex, alpha subunit, isoform 1 | 3 | 7 | Mus musculus |
| 10863945 | ATP-dependent DNA helicase II | 20 | 28 | Homo sapiens |
| 4503841 | ATP-dependent DNA helicase II, 70 kDa subunit | 22 | 28 | Homo sapiens |
| 1040689 | ATP-dependent DNA helicase Q1 (DNA-dependent ATPase Q1) | _ | 4 | Homo sapiens |
| 42740907 | Clusterin isoform 2 | − | 8 | Homo sapiens |
| 22208854 | Cytidine monophospho-N-acetylneuraminic acid synthetase | 3 | 3 | Mus musculus |
| 4557515 | Damage-specific DNA binding protein 2 (48kD) | 6 | 18 | Homo sapiens |
| 45476309 | Damage-specific DNA binding protein 2 splicing variant D4 | 7 | 13 | Homo sapiens |
| 4758138 | DEAD (Asp-Glu-Ala-Asp) box polypeptide 5 | 15 | 13 | Homo sapiens |
| 506756 | Desmoyokin | − | 6 | |
| 1706481 | DNA ligase III | 23 | 37 | Homo sapiens |
| 11225260 | DNA topoisomerase I | 10 | 11 | Homo sapiens |
| 347019 | DnaK-type molecular chaperone hsp72-ps1 | 8 | 10 | Rattus norvegicus |
| 1722884 | DNA-repair protein complementing XP-C cells | − | 6 | Drosophila melanogaster |
| 38201714 | ELAV-like 1 | 10 | − | Homo sapiens |
| 4503471 | Eukaryotic translation elongation factor 1 α 1 | 6 | 2 | Homo sapiens |
| 4503477 | Eukaryotic translation elongation factor 1 β 2 | 7 | − | Homo sapiens |
| 25453472 | Eukaryotic translation elongation factor 1 δ isoform 2 | 22 | − | Homo sapiens |
| 110625979 | Eukaryotic translation elongation factor 1 γ | 6 | − | Mus musculus |
| 37078458 | Far upstream element-binding protein 1 (FUSE-binding protein 1) | 11 | 13 | Mus musculus |
| 27806887 | Fusion (involved in t(12;16) in malignant liposarcoma) | 5 | _ | Bos taurus |
| 7669492 | Glyceraldehyde-3-phosphate dehydrogenase | 9 | − | Mus musculus |
| 74736618 | Grainyhead-like protein 2 homolog | − | 16 | Homo sapiens |
| 51316135 | GTP-binding nuclear protein Ran, testis-specific isoform | 10 | − | Rattus norvegicus |
| 4504517 | Heat shock 27kDa protein 1 | 33 | − | Homo sapiens |
| 2495339 | Heat shock 70 kDa protein 1B | 8 | 10 | Bos taurus |
| 8393547 | Heterogeneous nuclear ribonucleoprotein A1 | 13 | 11 | Rattus norvegicus |
| 14043072 | Heterogeneous nuclear ribonucleoprotein A2/B1 isoform B1 | 19 | 10 | Homo sapiens |
| 31559916 | Heterogeneous nuclear ribonucleoprotein A3 isoform a | 11 | 15 | Mus musculus |
| 55956919 | Heterogeneous nuclear ribonucleoprotein AB isoform a | 9 | 3 | Homo sapiens |
| 14249959 | Heterogeneous nuclear ribonucleoprotein C (C1/C2) | 10 | 2 | Homo sapiens |
| 870747 | Heterogeneous nuclear ribonucleoprotein D (hnRNP D) | 11 | 22 | Homo sapiens |
| 10946928 | Heterogeneous nuclear ribonucleoprotein H1 | 13 | 8 | Mus musculus |
| 14141157 | Heterogeneous nuclear ribonucleoprotein H3 isoform a | 11 | − | Homo sapiens |
| 13384620 | Heterogeneous nuclear ribonucleoprotein K | 23 | 10 | Mus musculus |
| 133274 | Heterogeneous nuclear ribonucleoprotein L (hnRNP L) | 4 | 4 | Homo sapiens |
| 14141152 | Heterogeneous nuclear ribonucleoprotein M isoform a | 15 | 18 | Homo sapiens |
| 5031755 | Heterogeneous nuclear ribonucleoprotein R isoform 2 | 6 | 6 | Homo sapiens |
| 8393536 | High mobility group box 2 | 14 | − | Rattus norvegicus |
| 1568557 | Histone H2B | 19 | 12 | Homo sapiens |
| 968888 | HMG-1 | 13 | − | Homo sapiens |
| 32358 | HnRNP U protein | 11 | 9 | Homo sapiens |
| 37183160 | HRPE773 | − | 29 | Homo sapiens |
| 1040689 | Human Diff6, H5, CDC10 homologue, KIAA0097 | 6 | − | Homo sapiens |
| 13385872 | Interleukin enhancer binding factor 2 | 8 | 13 | Mus musculus |
| 4504865 | KH-type splicing regulatory protein (FUSE binding protein 2) | 6 | 8 | Homo sapiens |
| 6453818 | Kinesin family member 22 | − | 20 | Homo sapiens |
| 2055427 | KSRP | 5 | 26 | Homo sapiens |
| 27436946 | Lamin A/C isoform 1 precursor | 20 | 15 | Homo sapiens |
| 5031877 | Lamin B1 | 8 | 21 | Homo sapiens |
| 14149645 | Methyl CpG binding protein 2 | 4 | 4 | Mus musculus |
| 1770458 | M-phase phosphoprotein 4 | 12 | 6 | Homo sapiens |
| 4038587 | Mucin | − | 2 | Homo sapiens |
| 55956788 | Nucleolin | 5 | 5 | Homo sapiens |
| 58037163 | Nucleoporin 35 | 6 | − | Mus musculus |
| 35119 | NuMA protein | − | 2 | Homo sapiens |
| 1483131 | p80 protein | 3 | 3 | Homo sapiens |
| 115497186 | Paraspeckle component 1 | 2 | 7 | Bos taurus |
| 156523968 | Poly (ADP-ribose)polymerase | 32 | 36 | Homo sapiens |
| 14141166 | Poly(rC)-binding protein 2 isoform b | 20 | 9 | Homo sapiens |
| 31543419 | Polynucleotide kinase 3 phosphatase | 13 | 11 | Homo sapiens |
| 4506243 | Polypyrimidine tract-binding protein 1 isoform a | 8 | 10 | Homo sapiens |
| 4502801 | Regulator of chromosome condensation (RCC1) | 12 | 10 | Homo sapiens |
| 4506583 | Replication protein A1, 70kDa | 18 | 26 | Homo sapiens |
| 4506585 | Replication protein A2, 32kDa | 7 | − | Homo sapiens |
| 4506439 | Retinoblastoma binding protein 7 | 3 | − | Homo sapiens |
| 15718687 | Ribosomal protein S3 | 23 | − | Homo sapiens |
| 7159943 | RNA and export factor binding protein 1-II | 24 | 9 | Mus musculus |
| 4757926 | RNA binding motif protein 39 isoform b | 3 | − | Homo sapiens |
| 66346679 | SERPINE1 mRNA binding protein 1 isoform 1 | 9 | 9 | Homo sapiens |
| 2440078 | SPIN protein | − | 4 | Homo sapiens |
| 4826998 | Splicing factor proline/glutamine rich | 9 | 11 | Homo sapiens |
| 5902076 | Splicing factor, arginine/serine-rich 1 isoform 1 | 4 | − | Homo sapiens |
| 39930517 | Sterile alpha motif domain containing 1 | 11 | 17 | Homo sapiens |
| 30088600 | Telomerase subunit EST1A | 1 | 1 | Homo sapiens |
| 4507555 | Thymopoietin isoform alpha | 8 | 17 | Homo sapiens |
| 4507401 | Transcription factor A, mitochondrial | 4 | - | Homo sapiens |
| 14389309 | Tubulin alpha 6 | 6 | 12 | Homo sapiens |
| 12963615 | Tubulin, beta 3 | 9 | 8 | Mus musculus |
| 21361322 | Tubulin, beta 4 | 9 | 9 | Homo sapiens |
| 1066004 | Typr II DNA topoisomerase beta isoform | − | 1 | Mus musculus |
| 28077011 | Tyrosyl-DNA phodphodiesterase 1 | 4 | 4 | Mus musculus |
| 4506387 | UV excision repair protein RAD23 homolog B | 4 | − | Homo sapiens |
| 2632123 | Xeroderma Pigmentosum Group E Complementing protein | 7 | 17 | Homo sapiens |
| 5454172 | X-ray repair cross complementing protein 1 | 9 | 12 | Homo sapiens |
| Gene | Subcellular fraction | Function |
|---|---|---|
| Cytoskeleton-associated proteins | ||
| KIAA0097 ; Human Diff6 | Cytoplasm Nucleus | Microtubule associated protein. |
| Kinesin family member 22 | Nucleus | Microtubule-dependent molecular motor. |
| NuMA protein | Nucleus | Component of the mitotic spindle matrix. |
| Cytoplasm Nucleus | Related to microtubules. |
| Factors involved in transcription, regulation of nucleobase, nucleoside, nucleotide and nucleic acid metabolism | ||
| Cytoplasm Nucleus | Involved in enzymatic delivery of aminoacyl tRNAs to the ribosome. |
| Poly(rC)-binding protein 2 isoform b | Cytoplasm Nucleus | RNA binding protein involved in regulation of nucleobase, nucleoside, nucleotide and nucleic acid metabolism. |
| Telomerase subunit EST1A Alternative name: Chromosome 17 open reading frame 31 Est1p like protein A Ever shorter telomeres 1 | Nucleus Nucleolus | Telomere regulation. |
| Nuclear stability, chromatin structure, cycle control, gene expression | ||
| Nucleus | Nuclear stability, chromatin structure and gene expression. |
| Proteins with a known function in DNA repair, breaking, replication, transcription and cell death | ||
| Apurinic/apyrimidinic endonuclease 1 | Nucleus | DNA repair and redox regulatory activities. |
| ATP-dependent DNA helicase II, 70 kDa subunit ATP-dependent (80 kda) | Nucleus | Repair of nonhomologous DNA ends. |
| ATP-dependent DNA helicase Q1 (DNA-dependent ATPase | Nucleus | Repair of UV-irradiated DNA damage or other mutagens. |
| Cytoplasm Nucleus | Repair of UV-irradiated DNA damage. |
| Cytoplasm Nucleus Nucleoplasm | Type I topoisomerase breaks single DNA strands, whereas Type II topoisimerase breaks double stranded DNA. |
| Nucleus | DNA repair, recombination, replication, and transcription. |
| Histone 1,2 and 3 | Nucleus | Play a central role in transcription regulation, DNA repair, DNA replication and chromosomal stability. |
| Poly (ADP-ribosylation) polymerase I-PARP-1 | Nucleus | DNA repair, replication, transcription and cell death. |
| Polynucleotide kinase 3 phosphatase | Nucleus | DNA repair following ionizing radiation or oxidative damage. |
| Replication protein A1, 70kDa Replication Protein A2, 32-KD; RPA2 | Nucleus | Replication, recombination and DNA repair. |
| Ribosomal Protein S3; RPS3 | Cytoplasm Nucleus | DNA repair and apoptosis. |
| Xeroderma Pigmentosum Group E Complementing protein | Cytoplasm Nucleus | Involved in DNA repair. |
| X-ray repair cross complementing protein 1 | Nucleus | Repair of single-strand DNA breaks. |
| RNA slipicing, RNA processing, Transcription, Translation and other functions | ||
| Nucleus | Implicated in cellular processes involving alteration of RNA secondary structure. |
| ELAV-like 1 | Cytoplasm Nucleus | RNA-binding proteins. |
| Fusion (involved in t(12;16) in malignant liposarcoma) | Nucleus | Plays a role in maintenance of genomic integrity. |
| hnRNP U protein | Cell surface Nucleus | Binds to pre-mRNA. |
| Interleukin enhancer binding factor 2 variant | Nucleus Nucleolus | Regulates transcription of the IL2 gene during T-cell activation. |
| Nucleolin | Nucleus Nucleolus | The major nucleolar protein of growing eukaryotic cells. |
| RNA and export factor binding protein 1 | Cytoplasm Nucleus | Nuclear mRNA export pathway. |
| RNA binding motif protein 39 isoform b | Nucleus | Transcriptional coactivator for steroid nuclear receptors ESR1/ER α and ESR2/ER-β, and JUN/AP-1. |
| Cytoplasm Nucleus | Prevents exon skipping. |
| Others | ||
| Adenylate kinase isozyme 2 | Membrane Microsomal Mitochondria Nucleus | Small ubiquitous enzyme. |
| Alkyladenine DNA glycosylase (AAG), also known as 3-methyladenine DNA glycosylase, | Nucleus | Catalyzes the first step in base excision repair. |
| Arginase, type I | Mitochondria | Plays a role in the regulation of extra-urea cycle arginine metabolism. |
| ATP synthase, H+ transporting, mitochondrial F1 complex, α subunit, isoform 1 | Mitochondria Cytoplasm Nucleus | Produces ATP from ADP in the presence of a proton gradient across the membrane. |
| BBF2H7/FUS protein | Nucleus | Binds both single-stranded and double-stranded DNA and promotes ATP-independent annealing of complementary single-stranded DNAs and d-loop formation in superhelical double-stranded. |
| Clusterin | Cytoplasm Nucleus | Undefined function, being described as being both pro-apoptotic and anti- apoptotic. |
| Cytidine monophospho-N-acetylneuraminic acid synthetase | Cytoplasm Nucleus | Involved in the biosynthesis of sialo-glycoproteins and gangliosides. |
| Desmoyokin/AHNAK | Plasma Membrane Cytoplasm Nucleus | Involved in the formation of cytoskeletal structure, calcium homeostasis, and muscle regeneration. |
| DNA ligase III α and β | Nucleus | Interacts with DNA-repair protein XRCC1 and corrects defective DNA strand-break repair and sister chromatid exchange. |
| Cytosol Perinuclear Mitochondria Endoplasmatic Reticulum Nucleus | Chaperone activity, thermotolerance, inhibition of apoptosis, regulation of cell development and cell differentiation. |
| DNA-repair protein complementing | Nucleus | Nucleotide excision repair (NER) of DNA. |
| Nucleus (Probable). | Regulates MYC expression by binding to a far-upstream element (FUSE) upstream of the MYC promoter. |
| Grainyhead-like protein 2 homolog (Brother of mammalian grainyhead) (Transcription factor CP2-like 3) | Nucleus | Transcription factor involved in epidermal differentiation. |
| GTP-binding nuclear protein Ran, testis-specific isoform | Cytoplasm Nucleus | Nuclear transport pathways, required for the import of protein into the nucleus and also for RNA export. |
| Gyceraldehyde-3-phosphate dehydrogenase | Membrane Cytoplasm Nucleus | Carbohydrate metabolism and membrane trafficking in the early secretory pathway. |
| Methyl CpG binding protein 2 | Nucleus | Nuclear proteins containing a methyl-binding domain (MBD) |
| M-phase phosphoprotein 4 | Nucleolus | Regulation of cell cycle, cell communication and signal transduction. |
| Mucin | Cytoplasm Mitochondria Nucleus | Activation of Wnt target genes. |
| Nucleoporin 35 [Nuclear pore glycoprotein-210 (gp210)] | Membrane Cytoplasm Nucleus | Trafficking regulator in the eukaryotic nuclear pore complex. |
| p80 protein | Cytoplasm Nucleus | Part of the nuclear coiled bodies (CBS) involved in assembly/disassembly of nucleoplasmic snRNPs. |
| Paraspeckle component 1 | Cytoplasm Nucleus Nucleolus | Regulates cooperatively with NONO and SFPQ, androgen receptor-mediated gene transcription activity. |
| PREDICTED: similar to RAN protein | Cytoplasm Nucleus | Inhibits GTP exchange on Ran. |
| Regulator of chromosome condensation (RCC1) | Cytoplasm Nucleus | Promotes the exchange of Ran-bound GDP by GTP. |
| Retinoblastoma binding protein 7 | Nucleus | Modulates the functions ascribed to BRCA1 in transcriptional regulation, DNA repair, and/or cell cycle checkpoint control. |
| SERPINE1 mRNA binding protein 1 isoform 1 | Cytoplasm Nucleus | Regulates mRNA stability. |
| SPIN | Nucleus | Regulation of cell cycle |
| Sterile alpha motif domain containing 1 | Nucleus | Widespread domain in signalling and nuclear proteins. |
| Thymopoietin γ and β ( TMPOs) | Nucleus | Directs the assembly of the nuclear lamina. |
| Transcription factor A, mitochondrial | Mitochondria | Involved in mitochondrial transcription regulation. |
| UV excision repair protein RAD23 homolog B | Cytoplasm Nucleus | Plays a role both in proteosomal degradation of misfolded proteins and DNA repair. |
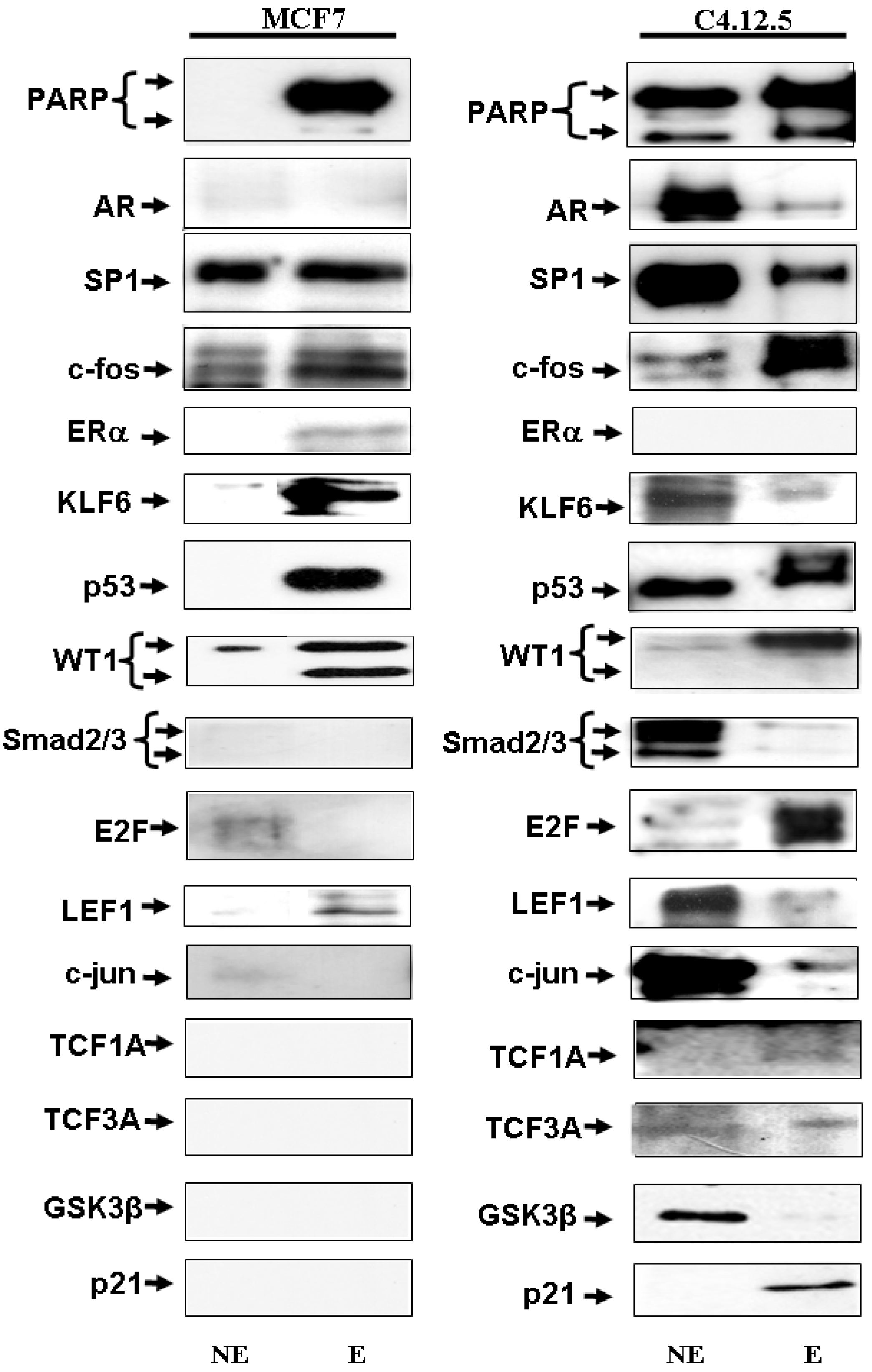
3.4. Identification of IGF-IR Promoter-Binding Transcription Factors by Western Blots

| Gene | MCF7 | C4.12.5 | Nomenclature |
|---|---|---|---|
| BRCA1 | 1 | 0.3 | Breast and ovarian cancer type I |
| TOPOII | 1 | 0 | Type II topoisomerase |
| IRS-I | 0 | 0 | Insulin receptor substrate-1 |
| PARP | 1 | 2 | Poly ADP ribose polymerase |
| MET | 1 | 0 | Met tyrosine kinase |
| p120 | 1 | 1.39 | Zinc finger protein 120 |
| AR | 1 | 0.64 | Androgen receptor |
| Sp1 | 1 | 0.79 | Trans-acting transcription factor 1 |
| Foxo1 | 1 | 0.5 | Forkhead Box A1 |
| ER | 1 | 0.59 | Estrogen receptor |
| WT1 | 1 | 0.8 | Wilms' tumor suppressor gene-1 |
| KLF6 | 1 | 1.2 | Kruppel-like factor 6 |
| LaminA/C | 1 | 2.4 | Nuclear protein |
| Smad 2 | 1 | 1.12 | Smad 2/3 family |
| Smad 3 | 0 | 2.6 | |
| pSmad2/3 | 1 | 1.13 | |
| c-Myc | 1 | 1.48 | Myelocytomatosis oncogene |
| E2F1 | 1 | 0.88 | E2F transcription factor 1 |
| TCF1 | 1 | 0.717 | T-cell factor 1 |
| TCF3A | 1 | 0.002 | High Mobility Group box transcription factor 3 |
| LEF1 | 1 | 1.15 | Lymphoid enhancer binding factor 1 |
| p53 | 1 | 0.96 | p53-related tumor suppressors |
| p63 | 1 | 1.12 | |
| p73 | 1 | 0.03 | |
| p21 | 1 | 0.91 | Waf1/Cip1 |
| GSK3α | 1 | 1.05 | Glycogen Synthase Kinase-3 |
| c-fos | 1 | 0.92 | c-fos/c-jun family of transcription factors |
| c-jun | 1 | 10 | |
| pc-jun | 1 | 0.5 | |
| Cyclin G | 1 | 0.13 | Regulatory subunits of Cdc2 p34 |
| Cyclin D | 1 | 0 | |
| Histone1 | 1 | 0.74 | Nuclear protein |
| Gene | Function |
|---|---|
| BRCA1 | Tumor suppressor involved in DNA damage repair, cell growth and apoptosis, and gene transcription. |
| TOPOII | Type II topoisomerase, breaks double strands of duplex DNA. |
| IRIS-I | Mediates insulin related activities. |
| PARP | DNA repair, replication, transcription and cell death. |
| MET | Receptor for HGF/SF. |
| p120 | Prototypical member of a subfamily of armadillo (Arm)-domain proteins involved in intercellular adhesion and in nuclei. |
| AR | Androgen receptor (AR), a zinc finger transcription factor of the nuclear receptor superfamily. |
| SP1 | Transcription factor of the zinc finger family. |
| Foxo1 | Transcription factor of the forkhead family. |
| ER | Estrogen receptor (ER), a DNA binding transcription factor of the nuclear receptor superfamily. |
| WT1 | Wilm’s tumor suppressor protein-1. |
| KLF6 | Kruppel-like zinc finger transcription factor-6. |
| LaminA/C | Involved in nuclear stability, chromatin structure and gene expression. |
| pSmad 2/3 | Smad transcription factors mediate the effect of TGF-β. |
| Smad 2/3 | |
| c-Myc | Nuclear proteins involved in cell proliferation, differentiation and neoplastic disease. |
| E2F1 | Involved in cell cycle progression, apoptosis and oncogenesis. |
| LEF1 | Members of the high mobility group (HMG) DNA binding protein family of transcription factors. |
| TCFA1 | |
| TCF3A | |
| P53 | Tumor suppressor p53, p63 and p73. |
| p63 | |
| p73 | |
| GSK3α | Involved in protein synthesis, cell adhesion, proliferation, and differentiation. |
| c-fos | Fos and Jun protooncogenes. |
| c-junp | |
| c-jun | |
| Cyclin G | Regulatory subunits of Cdc2 p34 and related cyclin-dependent kinases (Cdks). |
| Cyclin D | |
| Histone1 | Basic and water soluble nuclear proteins. |
| p21 | Tumor suppressor protein p21/Waf1/Cip1 acts as an inhibitor of cell cycle progression. |
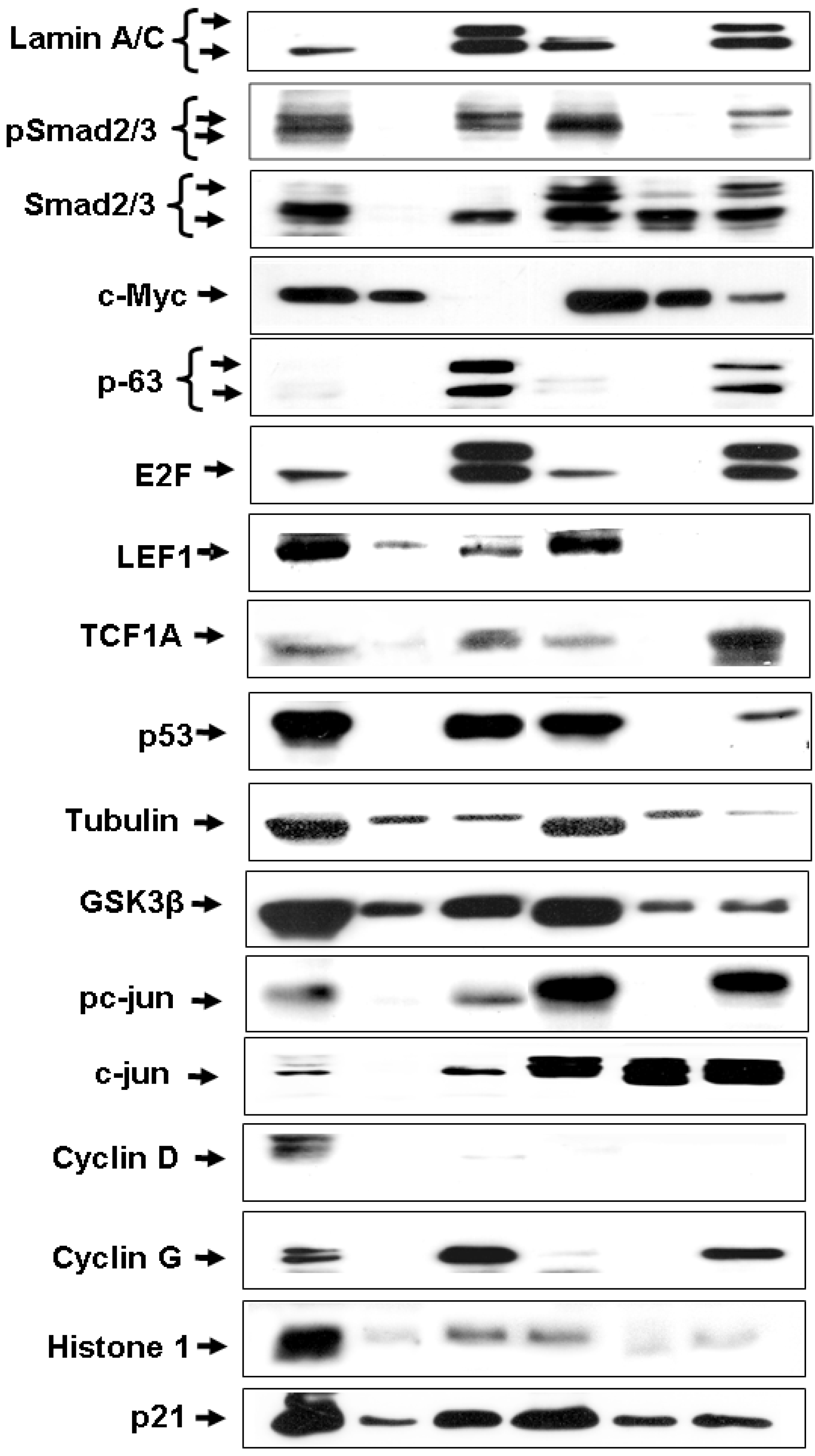
3.5. Cellular Distribution of IGF-IR Promoter-Binding Proteins

3.6. ChIP Assays

3.7. Regulation of IGF-IR Promoter Activity by c-jun, HMGA1, KLF6 and E2F1

4. Discussion
5. Conclusions
Abbreviations:
| IGF-IR: | Insulin-like growth factor I receptor |
| ER: | Estrogen receptor |
Acknowledgements
References
- Werner, H.; LeRoith, D. The role of the insulin-like growth factor system in human cancer. Adv. Cancer Res. 1996, 68, 183–223. [Google Scholar] [CrossRef]
- Yee, D.; Paik, S.; Lebovic, G.S.; Marcus, R.R.; Favoni, R.E.; Cullen, K.J.; Lippman, M.E.; Rosen, N. Analysis of insulin-like growth factor I gene expression in malignancy: evidence for a paracrine role in human breast cancer. Mol. Endocrinol. 1989, 3, 509–517. [Google Scholar] [CrossRef]
- Schnarr, B.; Strunz, K.; Ohsam, J.; Benner, A.; Wacker, J.; Mayer, D. Down-regulation of insulin-like growth factor-I receptor and insulin receptor substrate-1 expression in advanced human breast cancer. Int. J. Cancer 2000, 89, 506–513. [Google Scholar] [CrossRef]
- Surmacz, E. Function of the IGF-I receptor in breast cancer. J. Mammary Gland Biol. Neoplasia 2000, 5, 95–105. [Google Scholar] [CrossRef]
- LeRoith, D.; Werner, H.; Beitner-Johnson, D.; Roberts, C.T., Jr. Molecular and cellular aspects of the insulin-like growth factor I receptor. Endocr. Rev. 1995, 16, 143–163. [Google Scholar]
- Sell, C.; Rubini, M.; Rubin, R.; Liu, J.-P.; Efstratiadis, A.; Baserga, R. Simian virus 40 large tumor antigen is unable to transform mouse embryonic fibroblasts lacking type 1 insulin-like growth factor receptor. Proc. Natl. Acad. Sci. USA 1993, 90, 11217–11221. [Google Scholar] [CrossRef]
- Sell, C.; Dumenil, G.; Deveaud, C.; Miura, M.; Coppola, D.; DeAngelis, T.; Rubin, R.; Efstratiadis, A.; Baserga, R. Effect of a null mutation of the insulin-like growth factor I receptor gene on growth and transformation of mouse embryo fibroblasts. Mol. Cell. Biol. 1994, 14, 3604–3612. [Google Scholar]
- Arteaga, C.L.; Kitten, L.J.; Coronado, E.B.; Jacobs, S.; Kull, F.C., Jr.; Allred, D.C.; Osborne, C.K. Blockade of the type I somatomedin receptor inhibits growth of human breast cancer cells in athymic mice. J. Clin. Invest. 1989, 84, 1418–1423. [Google Scholar] [CrossRef]
- Surmacz, E.; Guvakova, M.; Nolan, M.; Nicosia, R.; Sciacca, L. Type I insulin-like growth factor receptor function in breast cancer. Breast Cancer Res. Treat. 1998, 47, 255–267. [Google Scholar] [CrossRef]
- Van der Burg, B.; Isbrucker, L.; Van Selm-Miltenburg, A.J.; de Laat, S.W.; Van Zoelen, E.J. Role of estrogen-induced insulin-like growth factors in the proliferation of human breast cancer cells. Cancer Res. 1990, 50, 7770–7774. [Google Scholar]
- Cullen, K.I.; Yee, D.; Sly, W.S.; Perdue, J.; Hampton, B.; Lippman, M.E.; Rosen, N. Insulin-like growth factor receptor expression and function in human breast cancer. Cancer Res. 1990, 50, 48–53. [Google Scholar]
- Wu, J.D.; Odman, A.; Higgins, L.M.; Haugk, K.; Vessella, R.; Ludwig, D.L.; Plymate, S.R. In vivo effects of the human type I insulin-like growth factor receptor antibody A12 on androgen-dependent and androgen-independent xenograft human prostate tumors. Clin. Cancer Res. 2005, 11, 3065–3074. [Google Scholar] [CrossRef]
- Ji, Q.S.; Mulvihill, M.J.; Rosenfeld-Franklin, M.; Cooke, A.; Feng, L.; Mak, G.; O'Connor, M.; Yao, Y.; Pirritt, C.; Buck, E.; Eyzaguirre, A.; Arnold, L.D.; Gibson, N.W.; Pachter, J.A. A novel, potent, and selective insulin-like growth factor-I receptor kinase inhibitor blocks insulin-like growth factor-I receptor signaling in vitro and inhibits insulin-like growth factor-I receptor dependent tumor growth in vivo. Mol. Cancer Ther. 2007, 6, 2158–2167. [Google Scholar] [CrossRef]
- Liu, W.; Bloom, D.A.; Cance, W.G.; Kurenova, E.V.; Golubovskaya, V.M.; Hochwald, S.N. FAK and IGF-IR interact to provide survival signals in human pancreatic adenocarcinoma cells. Carcinogenesis 2008, 29, 1096–1107. [Google Scholar] [CrossRef]
- Hankinson, S.E.; Willett, W.C.; Colditz, G.A.; Hunter, D.J.; Michaud, D.S.; Deroo, B.; Rosner, B.; Speizer, F.E.; Pollak, M. Circulating concentrations of insulin-like growth factor-I and risk of breast cancer. Lancet 1998, 351, 1393–1396. [Google Scholar] [CrossRef]
- Renehan, A.G.; Zwahlen, M.; C, M.; O'Dwyer, S.T.; Shalet, S.M.; Egger, M. Insulin-like growth factor-I, IGF binding protein-3, and cancer risk: systematic review and meta-regression analysis. Lancet 2004, 363, 1346–1353. [Google Scholar] [CrossRef]
- Beitner-Johnson, D.; Werner, H.; Roberts, C.T.Jr.; LeRoith, D. Regulation of insulin-like growth factor I receptor gene expression by Sp1: Physical and functional interactions of Sp1 at GC boxes and at a CT element. Mol. Endocrinol. 1995, 9, 1147–1156. [Google Scholar] [CrossRef]
- Rubinstein, M.; Idelman, G.; Plymate, S.R.; Narla, G.; Friedman, S.L.; Werner, H. Transcriptional activation of the IGF-I receptor gene by the Kruppel-like factor-6 (KLF6) tumor suppressor protein: potential interactions between KLF6 and p53. Endocrinology 2004, 145, 3769–3777. [Google Scholar] [CrossRef]
- Abramovitch, S.; Glaser, T.; Ouchi, T.; Werner, H. BRCA1-Sp1 interactions in transcriptional regulation of the IGF-IR gene. FEBS Lett. 2003, 541, 149–154. [Google Scholar] [CrossRef]
- Idelman, G.; Glaser, T.; Roberts, C.T., Jr.; Werner, H. WT1-p53 interactions in IGF-I receptor gene regulation. J. Biol. Chem. 2003, 278, 3474–3482. [Google Scholar]
- Werner, H.; Karnieli, E.; Rauscher, F.J., III; LeRoith, D. Wild type and mutant p53 differentially regulate transcription of the insulin-like growth factor I receptor gene. Proc. Natl. Acad. Sci. USA 1996, 93, 8318–8323. [Google Scholar] [CrossRef]
- Werner, H.; Re, G.G.; Drummond, I.A.; Sukhatme, V.P.; Rauscher, F.J., III; Sens, D.A.; Garvin, A.J.; LeRoith, D.; Roberts, C.T., Jr. Increased expression of the insulin-like growth factor-I receptor gene, IGFIR, in Wilms’ tumor is correlated with modulation of IGFIR promoter activity by the WT1 Wilms’ tumor gene product. Proc. Natl. Acad. Sci. USA 1993, 90, 5828–5832. [Google Scholar] [CrossRef]
- Yuen, J.S.P.; Cockman, M.E.; Sullivan, M.; Protheroe, A.; Turner, G.D.H.; Roberts, I.S.; Pugh, C.W.; Werner, H.; Macaulay, V.M. The VHL tumor suppressor inhibits expression of the IGF1R and its loss induces IGF1R upregulation in human clear cell renal carcinoma. Oncogene 2007, 26, 6499–6508. [Google Scholar] [CrossRef]
- Nahor, I.; Abramovitch, S.; Engeland, K.; Werner, H. The p53-family members p63 and p73 inhibit insulin-like growth factor-I receptor gene expression in colon cancer cells. Growth Hormone IGF Res. 2005, 15, 388–396. [Google Scholar] [CrossRef]
- Lee, A.V.; Jackson, J.G.; Gooch, J.L.; Hilsenbeck, S.G.; Coronado-Heinsohn, E.; Osborne, C.K.; Yee, D. Enhancement of insulin-like growth factor signaling in human breast cancer: estrogen regulation of insulin receptor substrate-1 expression in vitro and in vivo. Mol. Endocrinol. 1999, 13, 787–796. [Google Scholar] [CrossRef]
- Umayahara, Y.; Kawamori, R.; Watada, H.; Imano, E.; Iwama, N.; Morishima, T.; Yamasaki, Y.; Kajimoto, Y.; Kamada, T. Estrogen regulation of the insulin-like growth factor I gene transcription involves an AP-1 enhancer. J. Biol. Chem. 1994, 269, 16433–16442. [Google Scholar]
- Osborne, C.K.; Coronado, E.B.; Kitten, L.J.; Arteaga, C.I.; Fuqua, S.A.W.; Ramasharma, K.; Marshall, M.; Li, C.H. Insulin-like growth factor-II (IGF-II): a potential autocrine/paracrine growth factor for human breast cancer acting via the IGF-I receptor. Mol. Endocrinol. 1989, 3, 1701–1709. [Google Scholar] [CrossRef]
- Stewart, A.J.; Johnson, M.D.; May, F.E.B.; Westley, B.R. Role of insulin-like growth factors and the type I insulin-like growth factor receptor in the estrogen stimulated proliferation of human breast cancer cells. J. Biol. Chem. 1990, 265, 21172–21178. [Google Scholar]
- Salerno, M.; Sisci, D.; Mauro, L.; Guvakova, M.A.; Ando, S.; Surmacz, E. Insulin receptor substrate 1 is a target for the pure antiestrogen ICI 182,780 in breast cancer cells. Int. J. Cancer 1999, 81, 299–304. [Google Scholar] [CrossRef]
- McGuire, W.; Jackson, J.G.; Figueroa, J.A.; Shimasaki, S.A.; Powell, D.R.; Yee, D. Regulation of IGFBP expression by breast cancer cells: use of IGFBP-1 as an inhibitor of insulin-like growth factor action. J. Natl. Cancer Inst. 1992, 84, 1336–1341. [Google Scholar] [CrossRef]
- Mathieu, M.; Vignon, F.; Capony, F.; Rochefort, H. Estradiol down-regulates the mannose-6-phosphate/insulin-like growth factor-II receptor gene and induces cathepsin-D in breast cancer cells: a receptor saturation mechanism to increase the secretion of lysosomal proenzymes. Mol. Endocrinol. 1991, 5, 815–822. [Google Scholar] [CrossRef]
- Kahlert, S.; Nuedling, S.; van Eickels, M.; Vetter, H.; Meyer, R.; Grohe, C. Estrogen receptor α rapidly activates the IGF-I receptor pathway. J. Biol. Chem. 2000, 275, 18447–18453. [Google Scholar]
- Kato, S.; Endoh, H.; Masuhiro, Y.; Kitamoto, T.; Uchiyama, S.; Sasaki, H.; Masushige, S.; Gotoh, Y.; Nishida, E.; Kawashima, H.; Metzger, D.; Chambon, P. Activation of the estrogen receptor through phosphorylation by mitogen-activated protein kinase. Science 1995, 270, 1491–1494. [Google Scholar]
- Martin, M.B.; Franke, T.F.; Stoica, G.E.; Chambon, P.; Katzenellenbogen, B.S.; Stoica, B.A.; McLemore, M.S.; Olivo, S.E.; Stoica, A. A role for Akt in mediating the estrogenic functions of epidermal growth factor and insulin-like growth factor I. Endocrinology 2000, 141, 4503–4511. [Google Scholar] [CrossRef]
- Feng, W.; Webb, P.; Nguyen, P.; Liu, X.; Li, J.; Karin, M.; Kushner, P.J. Potentiation of estrogen receptor activation function 1 (AF-1) by Src/JNK through a serine 118-independent pathway. Mol. Endocrinol. 2001, 15, 32–45. [Google Scholar] [CrossRef]
- Joel, P.B.; Smith, J.; Sturgill, T.W.; Fisher, T.L.; Blenis, J.; Lannigan, D.A. pp90rsk1 regulates estrogen receptor-mediated transcription through phosphorylation of Ser-167. Mol. Cell Biol. 1998, 18, 1978–1984. [Google Scholar]
- Oesterreich, S.; Zhang, P.; Guler, R.L.; Sun, X.; Curran, E.M.; Welshons, W.V.; Osborne, C.K.; Lee, A.V. Re-expression of estrogen receptor α in estrogen receptor α-negative MCF-7 cells restores both estrogen and insulin-like growth factor-mediated signaling and growth. Cancer Res. 2001, 61, 5771–5777. [Google Scholar]
- Cooke, D.W.; Bankert, L.A.; Roberts, C.T., Jr; LeRoith, D.; Casella, S.J. Analysis of the human type I insulin-like growth factor receptor promoter region. Biochem. Biophys. Res. Comm. 1991, 177, 1113–1120. [Google Scholar] [CrossRef]
- Beer, I.; Barnea, E.; Ziv, T.; Admon, A. Improving large-scale proteomics by clustering of mass spectrometry data. Proteomics 2004, 4, 950–960. [Google Scholar] [CrossRef]
- Maor, S.; Mayer, D.; Yarden, R.I.; Lee, A.V.; Sarfstein, R.; Werner, H.; Papa, M.Z. Estrogen receptor regulates insulin-like growth factor-I receptor gene expression in breast tumor cells: involvement of transcription factor Sp1. J. Endocrinol. 2006, 191, 605–612. [Google Scholar] [CrossRef]
- Werner, H.; Bach, M.A.; Stannard, B.; Roberts, C.T., Jr; LeRoith, D. Structural and functional analysis of the insulin-like growth factor I receptor gene promoter. Mol. Endocrinol. 1992, 6, 1545–1558. [Google Scholar] [CrossRef]
- Werner, H.; Rauscher, F.J., III; Sukhatme, V.P.; Drummond, I.A.; Roberts, C.T., Jr.; LeRoith, D. Transcriptional repression of the insulin-like growth factor I receptor (IGF-I-R) gene by the tumor suppressor WT1 involves binding to sequences both upstream and downstream of the IGF-I-R gene transcription start site. J. Biol. Chem. 1994, 269, 12577–12582. [Google Scholar]
- Sgarra, R.; Lee, J.; Tessari, M.A.; Altamura, S.; Spolaore, B.; Giancotti, V.; Bedford, M.T.; Manfioletti, G. The AT-hook of the chromatin architectural transcription factor high mobility group A1a is arginine-methylated by protein arginine methyltransferase 6. J. Biol. Chem. 2006, 281, 3764–3772. [Google Scholar] [CrossRef]
- Berkovich, E.; Ginsberg, D. ATM is a target for positive regulation by E2F1. Oncogene 2003, 22, 161–167. [Google Scholar] [CrossRef]
- Reizner, N.; Maor, S.; Sarfstein, R.; Abramovitch, S.; Welshons, W.V.; Curran, E.; Lee, A.V.; Werner, H. The WT1 Wilms' tumor suppressor gene product interacts with estrogen receptor-alpha and regulates IGF-I receptor gene transcription in breast cancer cells. J. Mol. Endocrinol. 2005, 35, 135–144. [Google Scholar] [CrossRef]
- Sarfstein, R.; Maor, S.; Reizner, N.; Abramovitch, S.; Werner, H. Transcriptional regulation of the insulin-like growth factor-1 receptor in breast cancer. Mol. Cell. Endocrinol. 2006, 252, 241–246. [Google Scholar] [CrossRef]
- Mak, H.H.; Peschard, P.; Lin, T.; Naujokas, M.A.; Zuo, D.; Park, M. Oncogenic activation of the Met receptor tyrosine kinase fusion protein, Tpr-Met, involves exclusion from the endocytic degradative pathway. Oncogene 2007, 26, 7213–7221. [Google Scholar] [CrossRef]
- Barginear, M.F.; Bradley, T.; Shapira, I.; Budman, D.R. Implications of applied research for prognosis and therapy of breast cancer. Crit. Rev. Oncol. Hematol. 2008, 65, 223–234. [Google Scholar] [CrossRef]
- Happerfield, L.C.; Miles, D.W.; Barnes, D.M.; Thomsen, L.L.; Smith, P.; Hanby, A. The localization of the insulin-like growth factor receptor 1 (IGFR-1) in benign and malignant breast tissue. J. Pathol. 1997, 183, 412–417. [Google Scholar] [CrossRef]
- Peyrat, J.P.; Bonneterre, J. Type 1 IGF receptor in human breast diseases. Breast Cancer Res. Treat. 1992, 22, 59–67. [Google Scholar] [CrossRef]
- Papa, V.; Russo, P.; Gliozzo, B.; Goldfine, I.D.; Vigneri, R.; Pezzino, V. An intact and functional soluble form of the insulin receptor is secreted by cultured cells. Endocrinology 1993, 133, 1369–1376. [Google Scholar] [CrossRef]
- Yee, D. The insulin-like growth factor system as a target in breast cancer. Breast Cancer Res. Treat. 1994, 32, 85–95. [Google Scholar] [CrossRef]
- Turner, B.C.; Haffty, B.G.; Narayanan, L.; Yuan, J.; Havre, P.A.; Gumbs, A.A.; Kaplan, L.; Burgaud, J.-L.; Carter, D.; Baserga, R.; Glazer, P.M. Insulin-like growth factor-I receptor overexpression mediates cellular radioresistance and local breast cancer recurrence after lumpectomy and radiation. Cancer Res. 1997, 57, 3079–3083. [Google Scholar]
- Lee, A.V.; Hilsenbeck, S.G.; Yee, D. IGF system components as prognostic markers in breast cancer. Breast Cancer Res. Treat. 1998, 47, 295–302. [Google Scholar] [CrossRef]
- Karnieli, E.; Werner, H.; Rauscher, F.J., III; Benjamin, L.E.; LeRoith, D. The IGF-I receptor gene promoter is a molecular target for the Ewings' sarcoma-Wilms' tumor 1 fusion protein. J. Biol. Chem. 1996, 271, 19304–19309. [Google Scholar]
- Bentov, I.; Narla, G.; Schayek, H.; Akita, K.; Plymate, S.R.; LeRoith, D.; Friedman, S.L.; Werner, H. Insulin-like growth factor-i regulates Kruppel-like factor-6 gene expression in a p53-dependent manner. Endocrinology 2008, 149, 1890–1897. [Google Scholar] [CrossRef]
- Shahrabani-Gargir, L.; Pandita, T.K.; Werner, H. Ataxia-telangiectasia mutated gene controls insulin-like growth factor I receptor gene expression in a deoxyribonucleic acid damage response pathway via mechanisms involving zinc-finger transcription factors Sp1 and WT1. Endocrinology 2004, 145, 5679–5687. [Google Scholar] [CrossRef]
- Werner, H.; Roberts, C.T., Jr. The IGF-I receptor gene: a molecular target for disrupted transcription factors. Genes Chromosomes Cancer 2003, 36, 113–120. [Google Scholar] [CrossRef]
- Kadonaga, J.T.; Tjian, R. Affinity purification of sequence-specific DNA binding proteins. Proc. Natl. Acad. Sci. USA 1986, 83, 5889–5893. [Google Scholar] [CrossRef]
- Maor, S.; Yosepovich, A.; Papa, M.Z.; Yarden, R.I.; Mayer, D.; Friedman, E.; Werner, H. Elevated insulin-like growth factor-I receptor (IGF-IR) levels in primary breast tumors associated with BRCA1 mutations. Cancer Lett. 2007, 257, 236–243. [Google Scholar] [CrossRef]
© 2010 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Sarfstein, R.; Belfiore, A.; Werner, H. Identification of Insulin-Like Growth Factor-I Receptor (IGF-IR) Gene Promoter-Binding Proteins in Estrogen Receptor (ER)-Positive and ER-Depleted Breast Cancer Cells. Cancers 2010, 2, 233-261. https://doi.org/10.3390/cancers2020233
Sarfstein R, Belfiore A, Werner H. Identification of Insulin-Like Growth Factor-I Receptor (IGF-IR) Gene Promoter-Binding Proteins in Estrogen Receptor (ER)-Positive and ER-Depleted Breast Cancer Cells. Cancers. 2010; 2(2):233-261. https://doi.org/10.3390/cancers2020233
Chicago/Turabian StyleSarfstein, Rive, Antonino Belfiore, and Haim Werner. 2010. "Identification of Insulin-Like Growth Factor-I Receptor (IGF-IR) Gene Promoter-Binding Proteins in Estrogen Receptor (ER)-Positive and ER-Depleted Breast Cancer Cells" Cancers 2, no. 2: 233-261. https://doi.org/10.3390/cancers2020233