Mucins and Pancreatic Cancer



INSERM, U837, Jean-Pierre Aubert Research Center, Team 5 "Mucins, epithelial differentiation and carcinogenesis", Lille, France
*
Author to whom correspondence should be addressed.
Cancers 2010, 2(4), 1794-1812; https://doi.org/10.3390/cancers2041794
Submission received: 17 September 2010
/
Revised: 14 October 2010
/
Accepted: 18 October 2010
/
Published: 25 October 2010
(This article belongs to the Special Issue Pancreatic Cancer)
{kind=link}
{kind=link}
Abstract
:Pancreatic cancer is characterized by an often dramatic outcome (five year survival < 5%) related to a late diagnosis and a lack of efficient therapy. Therefore, clinicians desperately need new biomarkers and new therapeutic tools to develop new efficient therapies. Mucins belong to an ever increasing family of O-glycoproteins. Secreted mucins are the main component of mucus protecting the epithelia whereas membrane-bound mucins are thought to play important biological roles in cell-cell and cell-matrix interactions, in cell signaling and in modulating biological properties of cancer cells. In this review, we will focus on the altered expression pattern of mucins in pancreatic cancer, from the early neoplastic lesion Pancreatic Intraepithelial Neoplasia (PanIN) to invasive pancreatic carcinomas, and the molecular mechanisms (including genetic and epigenetic regulation) and signaling pathways known to control their expression. Moreover, we will discuss the recent advances about the biology of both secreted and membrane-bound mucins and their key roles in pancreatic carcinogenesis and resistance to therapy. Finally, we will discuss exciting opportunities that mucins offer as potential therapeutic targets in pancreatic cancer.
1. Pancreatic Cancer
Pancreatic Ductal Adenocarcinoma (PDAC) is the fourth leading cause of death by cancer in the world. The survival rate is extremely low (3–5%) and the survival curve at five years is very short (six months) [1]. This dramatic outcome is related to a lack of efficient therapeutic tools and early diagnostic markers [2]. At the time of diagnosis, more than 80% of PDACs are already metastatic or locally advanced. Therefore, only about 10% to 15% of patients are considered eligible for surgical resection. For the majority of patients, palliative therapy with gemcitabine-based chemotherapy remains the only option. However, chemotherapeutic drug resistance is a common feature of pancreatic tumors. Chemotherapy produces benefits and symptom improvements in only 20–30% of patients. Gemcitabine resistance is not yet understood [3,4]. Understanding the mechanisms of that resistance may thus help overcome drug resistance and allow development of novel therapeutic approaches.
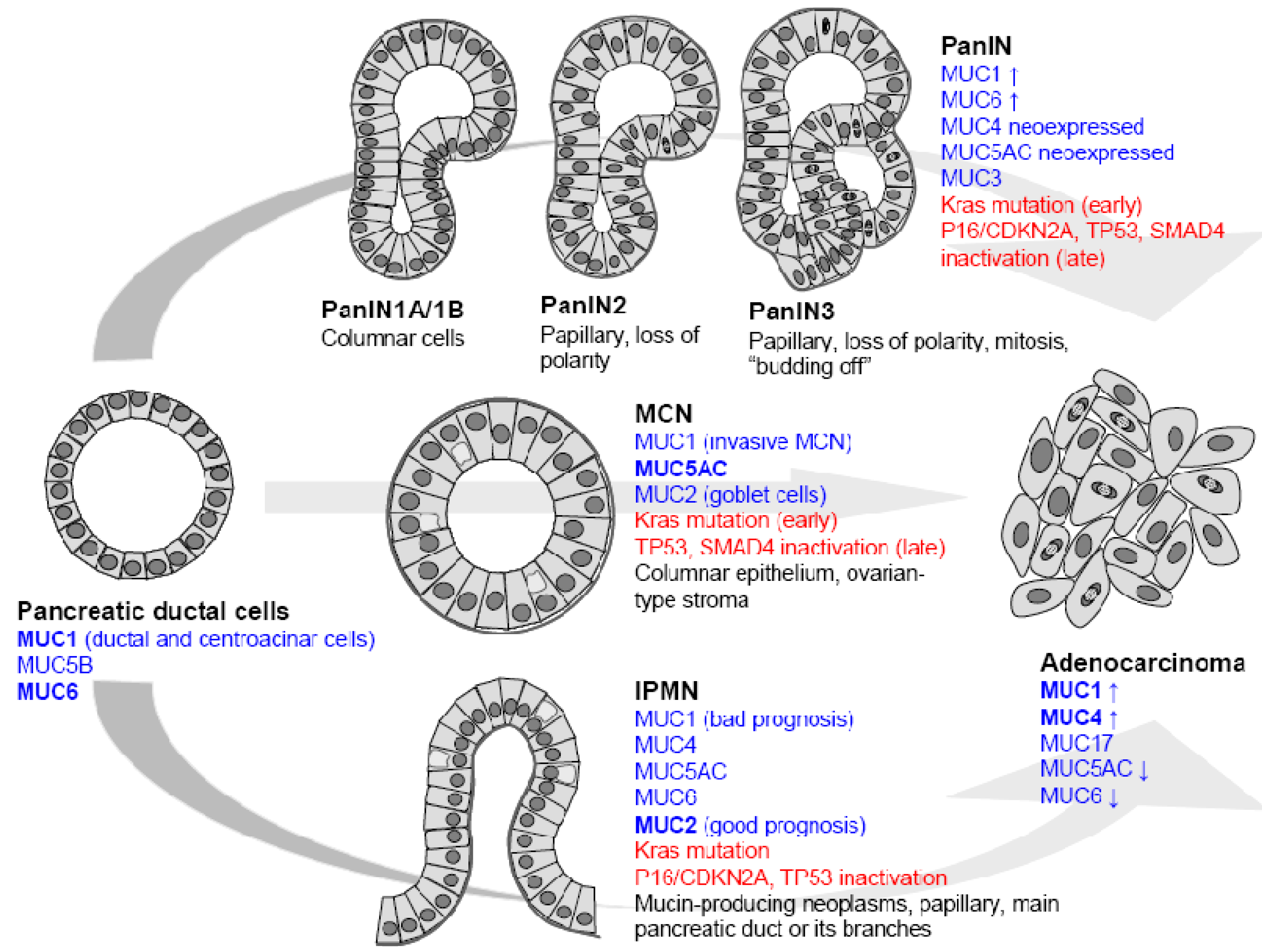
Clinical, pathological and genetics studies have identified three different preneoplastic lesions of the duct as precursors of pancreatic ductal adenocarcinoma (PDAC) [5,6]. These lesions are pancreatic intraepithelial neoplasia (PanIN), intraductal papillary mucinous neoplasm (IPMN) and mucinous cystic neoplasm (MCN). Among these lesions PanIN are the most frequent and best characterized both at the morphological and molecular levels (Figure 1).
Pancreatic carcinogenesis follows a hyperplasia/dysplasia/in situ carcinoma/invasive carcinoma progression. Hruban and colleagues have proposed a model with initial ductal proliferation toward invasive ductal carcinoma. 85% of PDAC develop from PanIN in which histologic, cytologic and genetic alterations (K-ras mutation, p16/CDKN2A, Smad4 deletion) are accumulated [6]. PanIN1A and PanIN1B are characterized by hyperplasia without dysplasia. PanIN2 lesions show variable dysplasia and PanIN3 represent in situ carcinoma. As an early event leading to PDAC, PanIN and genes expressed in PanIN are therefore potential therapeutic targets (Figure 1).
Intraductal Papillary Mucinous Neoplasms (IPMN) are thought to be another precursor of PDAC. They originate in the main pancreatic duct and are characterized by a massive dilatation of the ducts or cyst formation and hypersecretion of mucins. Usually, IPMN have a good clinical course but 10 to 20% of IPMN are invasive carcinomas with bad prognosis. The mutational spectrum of IPMN slightly differs from PDAC since Smad4 mutations are relatively uncommon. The overall five year survival for surgically resected patients with invasive adenocarcinoma arising from IPMN is 45% [5].
Mucinous cystic neoplasms (MCN) are defined as mucin-producing cyst forming epithelial neoplasms of the pancreas with a distinct ovarian type-stroma. Cysts of MCN are lined by a columnar mucin producing epithelium which can have a broad spectrum of dysplasia. Activation of Kras is an early event in the development of MCN whereas p53 and Smad4 gene mutation occurs at a later stage. Usually, MCN have a good prognosis since the five year survival is close to 100% for patients who undergo surgical resection without invasive carcinoma. When associated with invasive cancer, the mean five year survival is 50–60% [5].
Pancreatic endocrine carcinoma (PECA) or endocrine tumors account for less than 2% of pancreatic malignancies, although infrequent, their incidence has been increasing. Surgery is the only treatment modality with the potential to cure, but it is only likely to be effective in patients without diffuse metastatic disease [7].
These disastrous statistics for pancreatic cancer show that it is urgent to develop new diagnostic and/or prognostic tools for the clinicians in order to propose better healthcare management of the disease. Moreover, identification of new molecular targets by deciphering the molecular mechanisms underlying the disease will allow the development of new therapeutical approaches to treat the disease.
Figure 1.
Mucin expression in precursor lesions leading to pancreatic ductal adenocarcinoma. Pancreatic intraepithelial neoplasia (PanIN), intraductal papillary mucinous neoplasm (IPMN) and mucinous cystic neoplasm (MCN).
Figure 1.
Mucin expression in precursor lesions leading to pancreatic ductal adenocarcinoma. Pancreatic intraepithelial neoplasia (PanIN), intraductal papillary mucinous neoplasm (IPMN) and mucinous cystic neoplasm (MCN).

2. Mucins
Mucins belong to a heterogeneous family of large O-glycoproteins composed of a long peptidic chain called apomucin on which are linked hundreds of oligosaccharide side chains representing 50–80% of the total molecular weight (MW) of the molecules. Mucins were initially described as secreted by epithelia, able to form viscoelastic gels and responsible for rheological properties of mucus [8].
The family of secreted mucins, gel-forming components of viscoelastic mucus gels protecting the epithelia, includes mucins MUC2, MUC5AC, MUC5B, MUC6, and MUC19. They are all structurally related to the superfamily of cystine-knot growth factors [9]. Their main function is to participate in mucus formation by forming a tridimensional network via oligomerization domains to protect underlying epithelia against various injuries (inflammation, bacteria, virus, pollutants, pH, etc). MUC7 and MUC9 are smaller secreted mucins that do not oligomerize. Interestingly, four of the genes encoding gel-forming mucins, MUC2-MUC5AC-MUC5B-MUC6, are clustered on the p15 arm of chromosome 11. Among the secreted mucins, Yamazoe et al. showed that MUC5AC might have the potential to accelerate progression of PDAC since MUC5AC inhibition markedly reduced invasive and adhesive activities of SW1990 pancreatic cancer cells [10].
Membrane-bound mucins, which contain a transmembrane (TM) domain, include MUC1, MUC3A/B, MUC4, MUC12, MUC13, MUC15, MUC16, MUC17, MUC20 and MUC21 [11]. Among them, MUC1 and MUC4 are best characterized and are described in Section 2.1, Section 2.2 below, respectively. Based on their structure and localization they are thought to play important biological roles in cell-cell and cell-extracellular matrix interactions, in cell signaling and in biological properties of cancer cells [12]. Moreover, because of their specific pattern of expression during the different steps of tumor progression toward carcinoma, membrane-bound mucins remain under intense investigation as both potent new biomarkers and therapeutic targets in epithelial cancers [12,13].
2.1. MUC1
MUC1 was the first mucin gene characterized [14,15,16]. The gene encodes a large apomucin lacking EGF domains usually found in the membrane-bound mucins. The MUC1 mucin is also known as polymorphic epithelial mucin (PEM), episialin, DUPAN-2, DF3, human milk fat globule (HMFG), epithelial membrane antigen (EMA) and CD227. The MUC1 apomucin contains an O-glycosylated repeated central domain and a 72 amino acid cytoplasmic tail. Numerous reports have shown that the cytoplasmic tail of MUC1 which contains a SH2 interaction domain and several kinase phosphorylation sites, acts as a docking protein for cell signaling involving notably β-catenin, c-Src, glycogen-synthase kinase-3β (GSK-3β), protein kinase Cδ (PKCδ), and the four members of the ErbB receptor family. These interactions induce signaling pathways such as the "mitogen-activated protein kinase" (MAPK) [12,17].
MUC1 plays roles in cell signaling, differentiation and proliferation of epithelial cells [18]. The biological properties of MUC1, its altered expression and post-translational modifications also confer an important role to MUC1 in tumor progression and metastasis [19,20]. Notably, down-regulation of MUC1 expression using RNA interference decreases the metastatic potential of pancreatic adenocarcinoma in vivo [21].
2.2. MUC4
MUC4 gene was discovered and characterized in our laboratory [22,23,24]. The gene encodes a large apomucin composed of two sub-units MUC4α and MUC4β. MUC4α is an extracellular subunit featuring a typical hyperglycosylated region. MUC4β is a membrane-bound sub-unit containing epidermal growth factor (EGF)-like domains. Experimental evidence suggests that the EGF-like domains play a role in receptor-ligand interactions. The interaction of the MUC4 EGF1-like motif with ErbB2 is the best characterized, likely serving as a regulator in signaling related to growth, motility or differentiation properties of the cell [12,25]. Rat Muc4 and ErbB2 interaction leads to signaling pathway modulation. Muc4-ErbB2 complex induces differentiation via activation of the cell cycle inhibitor p27kip, whereas formation of a quaternary complex of Muc4-ErbB2-ErbB3-neuregulin activates PKB/Akt and MAPK pathways leading to proliferation and inhibition of apoptosis [26]. From these data, Carraway and colleagues have proposed Muc4 as a modulator of proliferation and differentiation [27].
MUC4 may also play important roles in the behavior of epithelial tumor cells. Indeed, invalidation of MUC4 expression by integration of MUC4 antisense RNA (siRNA) leads to reduced proliferation, motility and increased cellular aggregation possibly via modulation of ErbB2 expression [28]. Stable ectopic expression of a truncated form of MUC4 (lacking 90% of the tandem repeat (TR) region), resulted in increased growth, motility, and invasiveness of pancreatic cancer cells and in xenografted nude mice [29].
3. Mucins and the Normal Pancreas
MUC1 is the main membrane-bound mucin expressed in the normal pancreas (Figure 1). The labeling concerns acinar cells at the cytoplasmic level, and ductal cells at the apical level [30]. Therefore, MUC1 has been used as a marker of pancreatic ductal cells in several studies [31,32].
Despite controversial initial results, no MUC4 transcript or apomucin are found in normal pancreatic tissues [30]. However a number of non neoplastic duct lesions such as atrophic ducts, or ducts in the setting of an inflammatory reaction, show expression of MUC4.
Regarding secreted mucins, MUC2 is either absent or weakly expressed (33%) in ductal and acinar cells of the healthy pancreas [30,33]. MUC5B and MUC6 mRNA have been observed in intralobular ducts by in situ hybridization (ISH). In immunohistochemistry studies, MUC5B mucin is found expressed in both acinar (33%) and ductal cells (80%). Similarly, 12–27% of pancreatic ducts express MUC6 mucin. MUC5AC is rarely expressed in pancreatic ducts (2–4%).
4. Mucins and Pancreatic Cancer
Pancreatic carcinogenesis is characterized by an altered pattern of mucin expression at different stages of tumor progression (Figure 1). Membrane-bound mucin production is detected as early as PanIN1A stage and is characterized by neoexpression of MUC4 and increase of MUC1. The PanIN3 stage, previously named in situ carcinoma, is characterized by a strong expression of MUC4 and MUC1 and occurrence of MUC3 [13,34]. During carcinogenesis, gradual expression of MUC4 has been demonstrated by IHC: 17% of PanIN1A, 36% of PanIN2 and 85% of PanIN3 express MUC4 [35]. In true PDAC, the prevalence of MUC4 apomucin expression reaches 83 to 89% [35,36]. These results are in agreement with gene expression profile studies which showed that MUC4 belongs to the most differentially expressed genes in PDAC [37]. The MUC1 mucin is more frequently expressed by PanIN lesions (around 70% of the cases) compared to normal pancreas. Around 80% of PDAC samples still express MUC1 [33]. MUC17 was also shown to be overexpressed in pancreatic tumor cell lines and tumor tissues compared with the normal pancreas [38]. So far, there is no information about MUC17 in PanIN early precursors. PanIN lesions and PDAC express gastric-type mucin (MUC5AC and MUC6) but do not express intestinal type mucin MUC2. MUC5AC which is not expressed in healthy pancreas is neoexpressed in earliest stage PanIN1A (70% of cases) and its expression reaches 85% in PDAC [33]. MUC6 increase in PanIN1A has also been reported. MUC6 expression peaks in PanIN1A (74%) and then decreases during carcinogenetic progression (35% of PDAC).
IPMN expressing MUC1 correspond to invasive carcinoma with a short survival rate and bad prognosis whereas those that express MUC5AC correspond to slow-growing adenoma with good prognosis. The majority of IPMN express MUC2 (76–85% of cases) [39,40]. MUC1 expression is predominantly observed in pancreatobiliary-type papillae (44%) whereas MUC2 is expressed uniformly and diffusely in intestinal-type papillae (92%) but rarely and focally in the pancreatobiliary type (19%) [41]. Interestingly, MUC2+ MUC1− group of tumors (56%) is associated with an intermediate behavior [40], and is opposed to the data of Nakamura et al. who showed that MUC2+ tumors are correlated with a higher potential of malignancy than MUC2- tumors [42].
Because MCN are rare, no consensus about mucin expression has been established. Luttges et al. have shown that non invasive MCN are characterized by the expression of MUC5AC and lack of MUC1 whereas MUC2 is restricted to goblet cells scattered within the MCN epithelium. Invasive forms of MCN are MUC1 positive [43]. On the contrary, Terada et al. have reported that MUC1, MUC2, MUC3, and MUC5AC/MUC6 (both recognized by the A-HF antibody) apomucins were expressed in 7/8, 0/8, 2/8, and 3/8 cases, respectively [44].
5. Mucin Gene Regulation in Pancreatic Cancer
A better understanding of the molecular structure of regulatory regions as well as mechanisms governing mucin expression is also mandatory if one wants to assign direct roles to mucins in carcinogenesis and better understand their influence on the biological properties of the tumor cell. Studies aiming at deciphering the signaling pathways will allow identification of potential therapeutic targets.
MUC1 promoter has been extensively characterized mainly in breast cancer [13]. Despite being overexpressed in pancreatic cancer, no specific pancreatic transcriptional regulation has been published.
The MUC4 5’-flanking region (GenBank accession number: AF241535) has been characterized and contains two active promoters. A TATA-less proximal promoter, mainly composed of GC-rich domains that bind the transcription factor Sp1 as well as the CACCC box binding protein and a very high density of binding sites for factors known to initiate transcription in TATA-less promoter (Sp1, CACCC box, glucocorticoid receptor element, AP-1, polyomavirus Enhancer Activator-3 (PEA3) and Med-1). The distal promoter is characterized by a TATA box and contains numerous putative binding sites for Sp1, AP-1, AP-4, GATA and cyclic adenosine monophosphate (cAMP) responsive element binding protein (CREB) transcription factors [13,45].
Pancreatic cancer cells have been the major model for MUC4 regulation. In that model, MUC4 was shown to be regulated by a wide range of specific factors. Tumor suppressor AP-2 down-regulates MUC4 expression at both protein and transcriptional levels via two AP-2 cis-elements located in the proximal promoter [36]. Interestingly, AP2-α overexpression also leads to an increase of gemcitabine sensitivity [46]. The Ets family member PEA3 is implicated in a wide range of cellular processes such as differentiation, proliferation and transformation [47] and was shown to upregulate MUC4 expression in synergy with c-Jun and Sp1 whereas PEA3 represses the transcriptional activity of ErbB2 promoter [48]. IFN-γ inflammatory pathway increases MUC4 expression via STAT-1 upregulation [45,49].
In pancreatic carcinogenesis, TGF-β, which possesses both tumor-suppressive and oncogenic activities [50], is a strong activator of MUC4 expression [51,52]. TGF-β regulation involves a cooperation between Smad2 and Smad4 transcription factors, and Smad4 binding to seven Smad Binding Elements (SBE) located along proximal and distal promoters. When Smad4 is mutated and inactive as observed in 50% of PDAC, TGF-β1 is still able to activate MUC4 expression via mitogen activated protein kinase (MAPK), phosphoinositide-3 kinase (PI3K) and Protein Kinase A (PKA) signaling pathways [52]. Moreover, all-trans-retinoic acid (RA) treatment increases MUC4 expression via retinoic acid receptor-α (RAR-α). That upregulation is a consequence of the increase of expression of TGF-β2 regulating MUC4 expression in an autocrine manner [51]. IFN-γ and RA can synergize at the transcriptional level to affect MUC4 induction in pancreatic tumour cells [53]. MUC4 expression is negatively regulated by the “cystic fibrosis transmembrane conductance regulator” (CFTR) chloride channel that is implicated in multiple cellular functions. CFTR mutation carrier status is correlated with early-onset of pancreatic adenocarcinoma [54]. It was observed that CFTR was downregulated in pancreatic cancer cells and negatively correlated with MUC4 [55].
Very little is known about the regulation of the other membrane-bound mucins in pancreatic cancer. Only a strong enhancer was found in the MUC17 upstream region with a significant promoter activity in AsPC-1, and HPAF pancreatic cancer cell lines [38].
Despite an extremely intriguing upregulation in the earliest stage of pancreatic carcinogenesis, mechanisms that control atypically expressed secreted mucins remain to be deciphered. Transcriptional activity of the MUC5AC promoter region was analyzed in pancreatic cancer cell lines, SW1990 and HPAF. Sp1 and AP-1 were identified as key factors responsible for basal and PMA-induced MUC5AC transcription. PMA-induced upregulation is mediated by Sp1, PKC/ERK/AP-1 and PKC/JNK/AP-1 pathways [56]. Moreover, stimulation of adenylyl cyclase and the PKA pathway by forskolin and vasoactive intestinal peptide (VIP) increased MUC5AC antigen expression and release from pancreatic cancer cells suggesting that PKA may contribute to the up-regulation of MUC5AC expression seen in PanIN1A [57]. cDNA microarray analysis has enlightened the hypothesis that upregulation of gastric-type mucins (MUC5AC and MUC6) could be related, in a similar manner to the majority of foregut markers seen in early PanIN lesions, to the Hedgehog-mediated conversion of the normal pancreatic epithelium to a gastric epithelial differentiation program [58].
The correlation between mucins and foetal pancreatic development, that links the patterns of mucin expression found in PDAC, is not proved at this time. In vitro studies have shown so far that hepatocyte nuclear factors (HNF-1/-4), forkhead box A (FOXA1/A2), GATA-4/-5/-6, and caudal-related homeobox (CDX-1/-2) transcription factors which control cell differentiation of gut endoderm derived-tissues (including pancreas) during embryonic development, are potent regulators of human MUC2, human MUC4 and murine Muc5ac mucin genes [59,60,61,62]. Therefore, differentiation network involved in pancreatic development could also regulate mucin expression in a similar manner during carcinogenesis.
6. Epigenetic Regulation of Mucin Genes in Pancreatic Cancer
The term “epigenetics” refers to a heritable change in gene expression that is driven by mechanisms other than alteration of the nucleotide sequence [63]. Epigenetic changes may occur either at the transcriptional (methylation of DNA, methylation, acetylation and phosphorylation of histones) or post-transcriptional (miRNA) levels. These mechanisms are often altered in pathological processes [64]. Alterations of expression include both silencing and overexpression of mucin genes while promoters of mucin genes share a GC-rich structure and, for most of them, contain a CpG island, two structures characteristics in favor of epigenetic regulation [65,66].
MUC1 gene expression is regulated by a tightly related combination of DNA methylation and histone modification via histone H3 lysine 9 (H3-K9) in the 5'-flanking region of the MUC1 promoter in cancer cell lines including HPAFII (MUC1+), BxPC3 (MUC1+), PANC1 (MUC1+/–) pancreatic cell lines [67].
MUC4 proximal and distal promoters, which possess each a CpG island, are heavily methylated whether the gene is expressed or not in the cells. On the contrary, MUC4 5’-untranslated region (UTR) show a methylation profile correlated to the level of MUC4 expression in the cells. Moreover, this hypermethylation is correlated with a repressive histone code including histone deacetylation via HDAC3 and methylation of histone H3 via DNA methyltransferase (DNMT)-3A and DNMT3B [68]. Yamada and collaborators later confirmed the CpG methylation of MUC4 5’-UTR using the MassARRAY® compact system [69].
MUC2, MUC5AC, MUC5B and MUC6 genes belong to the 11p15 mucin gene cluster, which is located in a hot spot of abnormal methylation in cancer [70]. Epigenetic regulation of genes encoding secreted mucins has been mainly described in other tissues than pancreas [66].
Expression of MUC2 mRNA has been correlated with methylation of the proximal region of the promoter in pancreatic cancer cells whereas no direct link was found between methylation and MUC5AC expression. [71]. In PANC-1 cells, MUC2 expression is induced by 5-aza 2’-deoxycytidine (5-aza) and histone deacetylation. In a similar manner, 5-aza treatment leads to MUC5B increased mRNA level in CAPAN-1 pancreatic cancer cells. Chromatin immunoprecipitation (ChIP) assays indicated that, in PANC-1 cells, MUC2 and MUC5B repression was associated with histone H3 deacetylation and K9H3 methylation as well as with K27H3 trimethylation for MUC5B [72].
Recently, methylated CpG mapping of MUC2 and MUC5AC promoters have been determined in pancreatic cancer cell lines [73,74]. It was shown that promoter methylation acts in combination with associated H3 histone deacetylation [74,75].
So far, no data is in favor of MUC6 epigenetic regulation in pancreatic cancer.
miRNAs are differentially expressed during pancreatic carcinogenesis as in numerous organs. Interrogation of MiRbase and MiRanda databases revealed a large number of miRNAs that could potentially target the 3′-UTR of mucin genes including MUC1, MUC4, MUC17 or MUC2, MUC5AC, and MUC6 [66]. Very recent data indeed indicate that the MUC1 3’-UTR is directly regulated by miR-125b, miR-145 and miR-1226 in human breast cancer cell line [76,77,78].
7. Mucins as Biomarkers of Pancreatic Cancer
A major issue with PDAC is the dramatic lack of efficient biomarkers. Serum-based molecular markers include carbohydrate antigens CA 19-9 (sialyl Lewis blood antigen), CA125, and SC6-Ag, pyruvate kinase isoenzyme type 2 (M2-PK) and secreted proteins such as macrophage inhibitory cytokine 1 (MIC-1) [79]. Among available markers, those with a good sensitivity are also positive in patients with benign diseases and thus cannot be used routinely as screening or diagnostic markers. They still lack sensitivity, specificity or reproducibility. In clinical practice, the most commonly accepted use of CA 19-9 is to assess the prognosis and monitor the response to therapy.
Aberrant profiles of mucin expression offer promising options as biomarkers (see section 4). Notably, detection of MUC4 mucin could help distinguish reactive ductal epithelial cells from the cells of pancreatic adenocarcinoma in fine needle aspirate (FNA) samples guided by endoscopic ultrasound (EUS) [80]. Furthermore, the epitope born by the anti-MUC1 antibody (PAM4) is neoexpressed in early lesions and PDAC [81].
Generating multiple markers in combination with mucin typical pattern of expression may also provide a new approach for early detection of pancreatic cancer. For example, the detection of a glycan variant on MUC5AC mucin, using the lectin wheat-germ agglutinin, allows discrimination between MCN and IPMN from benign cystic lesions when used in combination with cyst fluid CA 19-9 with a better sensitivity and sensibility. These biomarkers were more efficient than cyst fluid carcinoembryonic antigen (CEA) [82].
8. Mucins as Therapeutic Tools in Pancreatic Carcinogenesis?
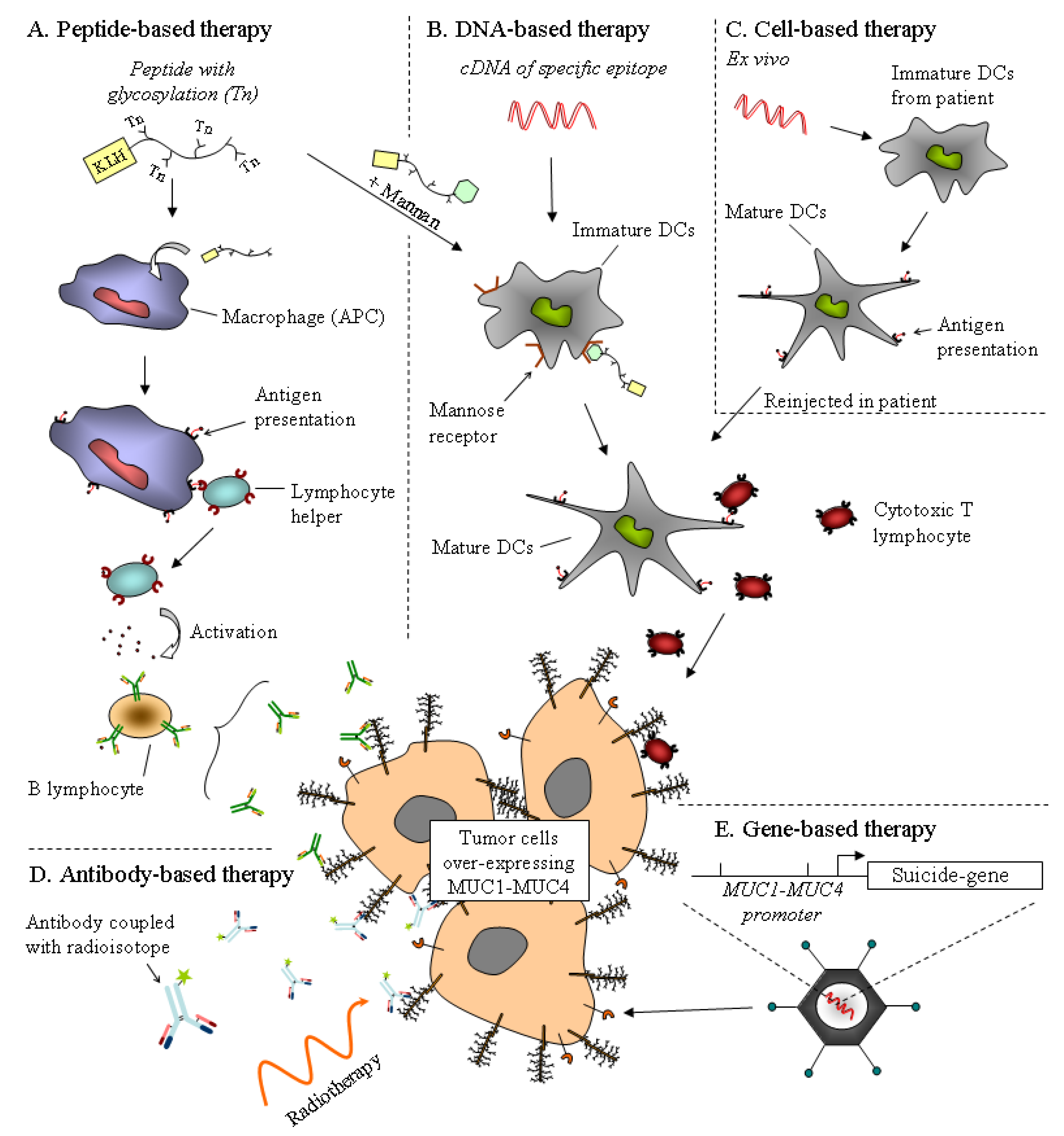
Mucins are thought to play important roles in the biological properties of tumor cells. One could consider mucins as promising therapeutic tools for gene therapy and immunotherapeutic approaches. Lack of effective therapeutic tools is a major reason for the dramatic outcome of PDAC. Using mucins as therapeutic targets could be a way to either potentialize existing therapy or develop new treatment for PDAC. So far, immuno-, gene based- and chemotherapy using mucins as therapeutic tools have been developed and hold promise (Figure 2).
8.1. Immunotherapy
MUC1 peptide core or glycopeptides have been used in immunotherapy assays mainly in breast cancer [12,83,84] that could be potentially useful in PDAC. Mab-PAM4 has been recently proposed as a promising anti-MUC1 antibody which reacts with 85% of pancreatic adenocarcinomas while showing no reactivity against normal pancreas or other tissues. Immuno- and radio-therapy can be combined to increase efficiency and tumor response. Combined chemotherapy with gemcitabine and low dose (131)I-PAM4 radioimmunotherapy may provide an improved alternative for the treatment of pancreatic cancer [81].
Recently, a peptide-based therapy using MUC1 protein transduction domain PTD4 (PMIP), which targets MUC1, EGFR and β-catenin interaction, displayed great efficiency in xenografted and transgenic mice models and inhibits cancer progression. This indicates a potential clinical implication in the treatment of MUC1+ pancreatic cancer [85].
8.2. Gene-Based Therapy
Promoters of mucin genes could also be used to drive cytotoxic agents (suicide genes) into the cells. Adenovirus with MUC1 promoter driving human somatostatin receptor subtype 2 (hSSTR2) in PANC-1 pancreatic cancer cells demonstrates a significant inhibition of cell proliferation in MUC1+ pancreatic carcinoma [86]. Since MUC4 is neoexpressed in early PanIN, its promoter could be used in a similar manner to target early pancreatic lesion.
Preliminary work has been conducted using MUC4 as a therapeutic target in DNA vaccine assays. Dendritic cell (DC)-based vaccine using MUC4 generated potent cytotoxic responses suggesting that PACRE/MUC4 DNA vaccine could also be a potential strategy for immunotherapy of MUC4+ tumors [87].
Figure 2.
Strategies for mucin-based therapies already used in phase trial. The aim of antigen-based therapy, by the use of a peptide or a cDNA derived from mucin epitopes, is to activate the immune system against tumor cells overexpressing the mucin. (A) The glycosylated mucin peptide, with KLH adjuvant, activates the humoral response by the activation of antigen-presenting cells (APC). The efficiency of this peptide may be increased with a mannan epitope recognized by a mannose receptor on dendritic cells (DCs), which in turn activates the cytotoxic response. (B) DNA vaccine encoding a specific mucin epitope is incorporated by DCs and activates the cytotoxic T lymphocyte. (C) Cell-based therapy is an alternative of DNA vaccine. Immature DCs extracted from the patient are incubated with DNA encoding the mucin epitope. Activated DCs exhibiting mucin antigen are injected into the patient. (D) Antibody-based therapy. Use of antibodies is promising for the detection and treatment of cancer. Antibodies against mucin epitopes are coupled with a radioisotope for radiotherapy ((131)I-PAM4) or a cytotoxic drug to target tumors. (E) Gene-based therapy. Promoter of mucin may be used to deliver a cytotoxic agent on tumor cells.
Figure 2.
Strategies for mucin-based therapies already used in phase trial. The aim of antigen-based therapy, by the use of a peptide or a cDNA derived from mucin epitopes, is to activate the immune system against tumor cells overexpressing the mucin. (A) The glycosylated mucin peptide, with KLH adjuvant, activates the humoral response by the activation of antigen-presenting cells (APC). The efficiency of this peptide may be increased with a mannan epitope recognized by a mannose receptor on dendritic cells (DCs), which in turn activates the cytotoxic response. (B) DNA vaccine encoding a specific mucin epitope is incorporated by DCs and activates the cytotoxic T lymphocyte. (C) Cell-based therapy is an alternative of DNA vaccine. Immature DCs extracted from the patient are incubated with DNA encoding the mucin epitope. Activated DCs exhibiting mucin antigen are injected into the patient. (D) Antibody-based therapy. Use of antibodies is promising for the detection and treatment of cancer. Antibodies against mucin epitopes are coupled with a radioisotope for radiotherapy ((131)I-PAM4) or a cytotoxic drug to target tumors. (E) Gene-based therapy. Promoter of mucin may be used to deliver a cytotoxic agent on tumor cells.

8.3. Chemotherapy
Gemcitabine is a 2′,2′-difluoro-2′-deoxycytidine analog which bioactivity results in cell apoptosis. For the majority of PDAC patients, resistance to gemcitabine-based chemotherapy is a common feature. Thus, chemotherapy produces benefits and symptom improvements only in 20–30% of patients. Understanding these mechanisms may help overcome drug resistance. MUC4 invalidation blocks activation of intrinsic apoptosis pathway induced by gemcitabine treatment. MUC4 promotes sequestration of Bad proteins in the cytoplasm and deactivation of BclXL via MUC4-ErbB2-ERK mediated pathway and therefore might contribute to gemcitabine resistance of total population and CD133+ cells [88,89]. Kalra and collaborators also showed that in pancreatic tumor cells, O-glycosylation inhibitor benzyl-α-GalNAc promotes the effectiveness of 5-fluorouracil (5-FU) [90]. Thus, intracellular uptake, antineoplastic and antitumor drug effects of 5-FU is enhanced in a reduced extracellular matrix environment with limited amount of mucins induced by intratumoural (IT) injections of benzyl-alpha-GalNAc [91].
9. Perspectives
So far, evaluation of the biological roles of mucins in pancreatic cancer has been carried out in vitro using cancer cell lines or in vivo using xenograft approaches. The development of new animal models by generating specific knockout (KO) mice or transgenic mice overexpressing mucins should provide new information about their role in vivo.
Very few mouse models (Muc1/Muc2/Muc16) have been published so far. MUC1 transgenic mice (MUC1.Tg), which express MUC1 in a similar pattern and level as observed in humans, have been crossed with LStopL-KrasG12D pancreatic cancer developing mice. The presence of MUC1 in these transgenic mice enhanced PanIN progression and development of pancreatic adenocarcinoma via an immunosuppressive effect due to a higher level of cyclooxygenase-2 or indoleamine 2,3-dioxygenase [92].
Mouse models have previously provided evidence of a direct role of MUC1 in tumor progression in other pathological models despite the fact that Muc1 KO mice develop normally, are healthy and fertile. These mice, however, showed a significantly slower growth rate of primary breast tumors [93]. Using the MUC1.Tg model, Lalani et al. showed a reduction in tumor incidence at low inocula and a delay in tumor growth at higher inocula [94]. This indicated for the first time that Muc1 participates in tumor progression. As for MUC1, future mucin KO mice could be crossed with models such as LStopL-KrasG12D, Trp53R172H, Ink4α or mice deficient in TGF-β/Smad pathway components. These models develop spontaneously pancreatic adenocarcinoma and faithfully recapitulate the PanIN sequence and therefore provide outstanding pathological study models [95,96,97,98].
Muc2 deficient mice spontaneously develop colonic adenoma that may degenerate into colorectal tumors [99]. Muc2 is thus involved in the suppression of colorectal cancer but the question “cause or consequence” of Muc2 role in tumor suppression remains to be answered. A Muc2 deficient mouse could be combined with a murine model of IPMN that remains to be characterized in order to assess the direct role of this secreted mucin in IPMN development.
Finally, Muc16 homozygous mutant mice are viable, fertile, and develop normally [100]. This KO mouse model provides a unique platform for future studies to identify the role of CA125/MUC16 in ovarian cancer. However, the potential use for pancreatic cancer remains to be demonstrated.
In the future, development of mouse models for MUC4, MUC5AC and MUC6 mucins that are overexpressed during pancreatic carcinogenesis would be of an extremely valuable interest and would greatly help the scientific community demonstrate the potential of these mucins as therapeutic tools in this disastrous cancer.
Acknowledgments
Nicolas Jonckheere is the recipient of a post-doctoral fellowship from la Ligue Nationale Contre le Cancer (LNCC). Nicolas Skrypek is the recipient of a PhD fellowship from Centre Hospitalier Régional Universitaire (CHRU) de Lille- région Nord-Pas de Calais. This work is funded by la Ligue Nationale Contre le Cancer (IVS, Equipe Labellisée LIGUE 2008).
References
- Bardeesy, N.; DePinho, R.A. Pancreatic cancer biology and genetics. Nat. Rev. Cancer 2002, 2, 897–909. [Google Scholar] [CrossRef]
- Schneider, G.; Siveke, J.T.; Eckel, F.; Schmid, R.M. Pancreatic cancer: basic and clinical aspects. Gastroenterology 2005, 128, 1606–1625. [Google Scholar] [CrossRef]
- El Maalouf, G.; Le Tourneau, C.; Batty, G.N.; Faivre, S.; Raymond, E. Markers involved in resistance to cytotoxics and targeted therapeutics in pancreatic cancer. Cancer Treat. Rev. 2009, 35, 167–174. [Google Scholar] [CrossRef]
- Kang, S.P.; Saif, M.W. Pharmacogenomics and pancreatic cancer treatment. Optimizing current therapy and individualizing future therapy. JOP: J. Pancreas 2008, 9, 251–266. [Google Scholar]
- Hruban, R.H.; Maitra, A.; Kern, S.E.; Goggins, M. Precursors to pancreatic cancer. Gastroenterol. Clin. North. Am. 2007, 36, 831–849. [Google Scholar] [CrossRef]
- Hruban, R.H.; Wilentz, R.E.; Kern, S.E. Genetic progression in the pancreatic ducts. Am. J. Pathol. 2000, 156, 1821–1825. [Google Scholar] [CrossRef]
- Figueiredo, F.A.; Giovannini, M.; Monges, G.; Charfi, S.; Bories, E.; Pesenti, C.; Caillol, F.; Delpero, J.R. Pancreatic endocrine tumors: a large single-center experience. Pancreas 2009, 38, 936–940. [Google Scholar]
- Dekker, J.; Rossen, J.W.; Buller, H.A.; Einerhand, A.W. The MUC family: an obituary. Trends Biochem. Sci. 2002, 27, 126–131. [Google Scholar] [CrossRef]
- Desseyn, J.L.; Gouyer, V.; Tetaert, D. Architecture of the gel-forming mucins. In The Epithelial Mucins: Structure/Function. Roles in Cancer and Inflammatory Diseases; Van Seuningen, I., Ed.; Research Signpost: Kerala, India, 2008; pp. 1–16. [Google Scholar]
- Yamazoe, S.; Tanaka, H.; Sawada, T.; Amano, R.; Yamada, N.; Ohira, M.; Hirakawa, K. RNA interference suppression of mucin 5AC (MUC5AC) reduces the adhesive and invasive capacity of human pancreatic cancer cells. J. Exp. Clin. Cancer Res. 2010, 29, 53. [Google Scholar] [CrossRef]
- Jonckheere, N.; Van Seuningen, I. The ever growing family of membrane-bound mucins. In The Epithelial Mucins: Structure/Function. Roles in Cancer and Inflammatory Diseases; Van Seuningen, I., Ed.; Research Signpost: Kerala, India, 2008; pp. 17–38. [Google Scholar]
- Jonckheere, N.; Van Seuningen, I. The membrane-bound mucins: how large O-glycoproteins play key roles in epithelial cancers and hold promise as biological tools for gene-based and immunotherapies. Crit. Rev. Oncog. 2008, 14, 177–196. [Google Scholar] [CrossRef]
- Jonckheere, N.; Van Seuningen, I. The membrane-bound mucins: From cell signalling to transcriptional regulation and expression in epithelial cancers. Biochimie 2010, 92, 1–11. [Google Scholar] [CrossRef]
- Lan, M.S.; Batra, S.K.; Qi, W.N.; Metzgar, R.S.; Hollingsworth, M.A. Cloning and sequencing of a human pancreatic tumor mucin cDNA. J. Biol. Chem. 1990, 265, 15294–15299. [Google Scholar]
- Ligtenberg, M.J.; Vos, H.L.; Gennissen, A.M.; Hilkens, J. Episialin, a carcinoma-associated mucin, is generated by a polymorphic gene encoding splice variants with alternative amino termini. J. Biol. Chem. 1990, 265, 5573–5578. [Google Scholar]
- Swallow, D.M.; Gendler, S.; Griffiths, B.; Corney, G.; Taylor-Papadimitriou, J.; Bramwell, M.E. The human tumour-associated epithelial mucins are coded by an expressed hypervariable gene locus PUM. Nature 1987, 328, 82–84. [Google Scholar] [CrossRef]
- Hollingsworth, M.A.; Swanson, B.J. Mucins in cancer: protection and control of the cell surface. Nat. Rev. Cancer. 2004, 4, 45–60. [Google Scholar] [CrossRef]
- Meerzaman, D.; Shapiro, P.S.; Kim, K.C. Involvement of the MAP kinase ERK2 in MUC1 mucin signaling. Am. J. Physiol. Lung Cell Mol. Physiol. 2001, 281, L86–91. [Google Scholar]
- Ho, J.J.; Kim, Y.S. Serological pancreatic tumor markers and the MUC1 apomucin. Pancreas 1994, 9, 674–691. [Google Scholar] [CrossRef]
- Schroeder, J.A.; Adriance, M.C.; Thompson, M.C.; Camenisch, T.D.; Gendler, S.J. MUC1 alters beta-catenin-dependent tumor formation and promotes cellular invasion. Oncogene 2003, 22, 1324–1332. [Google Scholar] [CrossRef]
- Tsutsumida, H.; Swanson, B.J.; Singh, P.K.; Caffrey, T.C.; Kitajima, S.; Goto, M.; Yonezawa, S.; Hollingsworth, M.A. RNA interference suppression of MUC1 reduces the growth rate and metastatic phenotype of human pancreatic cancer cells. Clin. Cancer Res. 2006, 12, 2976–2987. [Google Scholar] [CrossRef]
- Moniaux, N.; Nollet, S.; Porchet, N.; Degand, P.; Laine, A.; Aubert, J.P. Complete sequence of the human mucin MUC4: a putative cell membrane-associated mucin. Biochem. J. 1999, 338 (Pt 2), 325–333. [Google Scholar]
- Nollet, S.; Moniaux, N.; Maury, J.; Petitprez, D.; Degand, P.; Laine, A.; Porchet, N.; Aubert, J.P. Human mucin gene MUC4: organization of its 5'-region and polymorphism of its central tandem repeat array. Biochem. J. 1998, 332 (Pt 3), 739–748. [Google Scholar]
- Porchet, N.; Nguyen, V.C.; Dufosse, J.; Audie, J.P.; Guyonnet-Duperat, V.; Gross, M.S.; Denis, C.; Degand, P.; Bernheim, A.; Aubert, J.P. Molecular cloning and chromosomal localization of a novel human tracheo-bronchial mucin cDNA containing tandemly repeated sequences of 48 base pairs. Biochem. Biophys. Res. Commun. 1991, 175, 414–422. [Google Scholar] [CrossRef]
- Carraway, K.L.; Theodoropoulos, G.; Kozloski, G.A.; Carothers Carraway, C.A. Muc4/MUC4 functions and regulation in cancer. Future Oncol. 2009, 5, 1631–1640. [Google Scholar] [CrossRef]
- Jepson, S.; Komatsu, M.; Haq, B.; Arango, M.E.; Huang, D.; Carraway, C.A.; Carraway, K.L. Muc4/sialomucin complex, the intramembrane ErbB2 ligand, induces specific phosphorylation of ErbB2 and enhances expression of p27(kip), but does not activate mitogen-activated kinase or protein kinaseB/Akt pathways. Oncogene 2002, 21, 7524–7532. [Google Scholar] [CrossRef]
- Carraway, K.L.; Ramsauer, V.P.; Haq, B.; Carothers Carraway, C.A. Cell signaling through membrane mucins. Bioessays 2003, 25, 66–71. [Google Scholar] [CrossRef]
- Chaturvedi, P.; Singh, A.P.; Chakraborty, S.; Chauhan, S.C.; Bafna, S.; Meza, J.L.; Singh, P.K.; Hollingsworth, M.A.; Mehta, P.P.; Batra, S.K. MUC4 mucin interacts with and stabilizes the HER2 oncoprotein in human pancreatic cancer cells. Cancer Res. 2008, 68, 2065–2070. [Google Scholar] [CrossRef]
- Moniaux, N.; Chaturvedi, P.; Varshney, G.C.; Meza, J.L.; Rodriguez-Sierra, J.F.; Aubert, J.P.; Batra, S.K. Human MUC4 mucin induces ultra-structural changes and tumorigenicity in pancreatic cancer cells. Br. J. Cancer 2007, 97, 345–357. [Google Scholar] [CrossRef]
- Balague, C.; Gambus, G.; Carrato, C.; Porchet, N.; Aubert, J.P.; Kim, Y.S.; Real, F.X. Altered expression of MUC2, MUC4, and MUC5 mucin genes in pancreas tissues and cancer cell lines. Gastroenterology 1994, 106, 1054–1061. [Google Scholar]
- Cano, D.A.; Murcia, N.S.; Pazour, G.J.; Hebrok, M. Orpk mouse model of polycystic kidney disease reveals essential role of primary cilia in pancreatic tissue organization. Development 2004, 131, 3457–3467. [Google Scholar] [CrossRef]
- Pierreux, C.E.; Poll, A.V.; Kemp, C.R.; Clotman, F.; Maestro, M.A.; Cordi, S.; Ferrer, J.; Leyns, L.; Rousseau, G.G.; Lemaigre, F.P. The transcription factor hepatocyte nuclear factor-6 controls the development of pancreatic ducts in the mouse. Gastroenterology 2006, 130, 532–541. [Google Scholar] [CrossRef]
- Kim, G.E.; Bae, H.I.; Park, H.U.; Kuan, S.F.; Crawley, S.C.; Ho, J.J.; Kim, Y.S. Aberrant expression of MUC5AC and MUC6 gastric mucins and sialyl Tn antigen in intraepithelial neoplasms of the pancreas. Gastroenterology 2002, 123, 1052–1060. [Google Scholar] [CrossRef]
- Van Seuningen, I.; Leteurtre, E.; Pigny, P. Mucins in pancreas and hepato-biliary tract. Expression, regulation, biomarkers and therapy. In The Epithelial Mucins: Structure/Function. Roles in Cancer and Inflammatory Diseases; Van Seuningen, I., Ed.; Research Signpost: Kerala, India, 2008; pp. 233–248. [Google Scholar]
- Swartz, M.J.; Batra, S.K.; Varshney, G.C.; Hollingsworth, M.A.; Yeo, C.J.; Cameron, J.L.; Wilentz, R.E.; Hruban, R.H.; Argani, P. MUC4 expression increases progressively in pancreatic intraepithelial neoplasia. Am. J. Clin. Pathol. 2002, 117, 791–796. [Google Scholar]
- Fauquette, V.; Aubert, S.; Groux-Degroote, S.; Hemon, B.; Porchet, N.; Van Seuningen, I.; Pigny, P. Transcription factor AP-2alpha represses both the mucin MUC4 expression and pancreatic cancer cell proliferation. Carcinogenesis 2007, 28, 2305–2312. [Google Scholar] [CrossRef]
- Iacobuzio-Donahue, C.A.; Ashfaq, R.; Maitra, A.; Adsay, N.V.; Shen-Ong, G.L.; Berg, K.; Hollingsworth, M.A.; Cameron, J.L.; Yeo, C.J.; Kern, S.E.; Goggins, M.; Hruban, R.H. Highly expressed genes in pancreatic ductal adenocarcinomas: a comprehensive characterization and comparison of the transcription profiles obtained from three major technologies. Cancer Res. 2003, 63, 8614–8622. [Google Scholar]
- Moniaux, N.; Junker, W.M.; Singh, A.P.; Jones, A.M.; Batra, S.K. Characterization of human mucin MUC17. Complete coding sequence and organization. J. Biol. Chem. 2006, 281, 23676–23685. [Google Scholar] [CrossRef]
- Adsay, N.V. Role of MUC genes and mucins in pancreatic neoplasia. Am. J. Gastroenterol. 2006, 101, 2330–2332. [Google Scholar] [CrossRef]
- Ito, H.; Endo, T.; Oka, T.; Matumoto, T.; Abe, T.; Toyota, M.; Imai, K.; Satoh, M.; Maguchi, H.; Shinohara, T. Mucin expression profile is related to biological and clinical characteristics of intraductal papillary-mucinous tumors of the pancreas. Pancreas 2005, 30, e96–102. [Google Scholar] [CrossRef]
- Adsay, N.V.; Merati, K.; Basturk, O.; Iacobuzio-Donahue, C.; Levi, E.; Cheng, J.D.; Sarkar, F.H.; Hruban, R.H.; Klimstra, D.S. Pathologically and biologically distinct types of epithelium in intraductal papillary mucinous neoplasms: delineation of an "intestinal" pathway of carcinogenesis in the pancreas. Am. J. Surg. Pathol. 2004, 28, 839–848. [Google Scholar] [CrossRef]
- Nakamura, A.; Horinouchi, M.; Goto, M.; Nagata, K.; Sakoda, K.; Takao, S.; Imai, K.; Kim, Y.S.; Sato, E.; Yonezawa, S. New classification of pancreatic intraductal papillary-mucinous tumour by mucin expression: its relationship with potential for malignancy. J. Pathol. 2002, 197, 201–210. [Google Scholar] [CrossRef]
- Luttges, J.; Feyerabend, B.; Buchelt, T.; Pacena, M.; Kloppel, G. The mucin profile of noninvasive and invasive mucinous cystic neoplasms of the pancreas. Am. J. Surg. Pathol. 2002, 26, 466–471. [Google Scholar] [CrossRef]
- Terada, T.; Ohta, T.; Sasaki, M.; Nakanuma, Y.; Kim, Y.S. Expression of MUC apomucins in normal pancreas and pancreatic tumours. J. Pathol. 1996, 180, 160–165. [Google Scholar] [CrossRef]
- Perrais, M.; Pigny, P.; Ducourouble, M.P.; Petitprez, D.; Porchet, N.; Aubert, J.P.; Van Seuningen, I. Characterization of human mucin gene MUC4 promoter: importance of growth factors and proinflammatory cytokines for its regulation in pancreatic cancer cells. J. Biol. Chem. 2001, 276, 30923–30933. [Google Scholar] [CrossRef]
- Jonckheere, N.; Fauquette, V.; Stechly, L.; Saint-Laurent, N.; Aubert, S.; Susini, C.; Huet, G.; Porchet, N.; Van Seuningen, I.; Pigny, P. Tumour growth and resistance to gemcitabine of pancreatic cancer cells are decreased by AP-2alpha overexpression. Br. J. Cancer 2009, 101, 637–644. [Google Scholar] [CrossRef]
- Oikawa, T.; Yamada, T. Molecular biology of the Ets family of transcription factors. Gene 2003, 303, 11–34. [Google Scholar] [CrossRef]
- Fauquette, V.; Perrais, M.; Cerulis, S.; Jonckheere, N.; Ducourouble, M.P.; Aubert, J.P.; Pigny, P.; Van Seuningen, I. The antagonistic regulation of human MUC4 and ErbB-2 genes by the Ets protein PEA3 in pancreatic cancer cells: implications for the proliferation/differentiation balance in the cells. Biochem. J. 2005, 386, 35–45. [Google Scholar] [CrossRef]
- Andrianifahanana, M.; Singh, A.P.; Nemos, C.; Ponnusamy, M.P.; Moniaux, N.; Mehta, P.P.; Varshney, G.C.; Batra, S.K. IFN-gamma-induced expression of MUC4 in pancreatic cancer cells is mediated by STAT-1 upregulation: a novel mechanism for IFN-gamma response. Oncogene 2007, 26, 7251–7261. [Google Scholar] [CrossRef]
- Akhurst, R.J.; Derynck, R. TGF-beta signaling in cancer--a double-edged sword. Trends Cell. Biol. 2001, 11, S44–51. [Google Scholar]
- Choudhury, A.; Singh, R.K.; Moniaux, N.; El-Metwally, T.H.; Aubert, J.P.; Batra, S.K. Retinoic acid-dependent transforming growth factor-beta 2-mediated induction of MUC4 mucin expression in human pancreatic tumor cells follows retinoic acid receptor-alpha signaling pathway. J. Biol. Chem. 2000, 275, 33929–33936. [Google Scholar]
- Jonckheere, N.; Perrais, M.; Mariette, C.; Batra, S.K.; Aubert, J.P.; Pigny, P.; Van Seuningen, I. A role for human MUC4 mucin gene, the ErbB2 ligand, as a target of TGF-beta in pancreatic carcinogenesis. Oncogene 2004, 23, 5729–5738. [Google Scholar] [CrossRef]
- Andrianifahanana, M.; Agrawal, A.; Singh, A.P.; Moniaux, N.; van Seuningen, I.; Aubert, J.P.; Meza, J.; Batra, S.K. Synergistic induction of the MUC4 mucin gene by interferon-gamma and retinoic acid in human pancreatic tumour cells involves a reprogramming of signalling pathways. Oncogene 2005, 24, 6143–6154. [Google Scholar] [CrossRef]
- McWilliams, R.; Highsmith, W.E.; Rabe, K.G.; de Andrade, M.; Tordsen, L.A.; Holtegaard, L.M.; Petersen, G.M. Cystic fibrosis transmembrane regulator gene carrier status is a risk factor for young onset pancreatic adenocarcinoma. Gut 2005, 54, 1661–1662. [Google Scholar] [CrossRef]
- Singh, A.P.; Chauhan, S.C.; Andrianifahanana, M.; Moniaux, N.; Meza, J.L.; Copin, M.C.; van Seuningen, I.; Hollingsworth, M.A.; Aubert, J.P.; Batra, S.K. MUC4 expression is regulated by cystic fibrosis transmembrane conductance regulator in pancreatic adenocarcinoma cells via transcriptional and post-translational mechanisms. Oncogene 2007, 26, 30–41. [Google Scholar] [CrossRef]
- Kato, S.; Hokari, R.; Crawley, S.; Gum, J.; Ahn, D.H.; Kim, J.W.; Kwon, S.W.; Miura, S.; Basbaum, C.B.; Kim, Y.S. MUC5AC mucin gene regulation in pancreatic cancer cells. Int. J. Oncol. 2006, 29, 33–40. [Google Scholar]
- Ho, J.J.; Crawley, S.; Pan, P.L.; Farrelly, E.R.; Kim, Y.S. Secretion of MUC5AC mucin from pancreatic cancer cells in response to forskolin and VIP. Biochem. Biophys. Res. Commun. 2002, 294, 680–686. [Google Scholar] [CrossRef]
- Prasad, N.B.; Biankin, A.V.; Fukushima, N.; Maitra, A.; Dhara, S.; Elkahloun, A.G.; Hruban, R.H.; Goggins, M.; Leach, S.D. Gene expression profiles in pancreatic intraepithelial neoplasia reflect the effects of Hedgehog signaling on pancreatic ductal epithelial cells. Cancer Res. 2005, 65, 1619–1626. [Google Scholar] [CrossRef]
- Jonckheere, N.; Van Der Sluis, M.; Velghe, A.; Buisine, M.P.; Sutmuller, M.; Ducourouble, M.P.; Pigny, P.; Buller, H.A.; Aubert, J.P.; Einerhand, A.W.; Van Seuningen, I. Transcriptional activation of the murine Muc5ac mucin gene in epithelial cancer cells by TGF-beta/Smad4 signalling pathway is potentiated by Sp1. Biochem. J. 2004, 377, 797–808. [Google Scholar] [CrossRef]
- Jonckheere, N.; Vincent, A.; Perrais, M.; Ducourouble, M.P.; Male, A.K.; Aubert, J.P.; Pigny, P.; Carraway, K.L.; Freund, J.N.; Renes, I.B.; Van Seuningen, I. The human mucin MUC4 is transcriptionally regulated by caudal-related homeobox, hepatocyte nuclear factors, forkhead box A, and GATA endodermal transcription factors in epithelial cancer cells. J. Biol. Chem. 2007, 282, 22638–22650. [Google Scholar] [CrossRef]
- van der Sluis, M.; Melis, M.H.; Jonckheere, N.; Ducourouble, M.P.; Buller, H.A.; Renes, I.; Einerhand, A.W.; Van Seuningen, I. The murine Muc2 mucin gene is transcriptionally regulated by the zinc-finger GATA-4 transcription factor in intestinal cells. Biochem. Biophys. Res. Commun. 2004, 325, 952–960. [Google Scholar] [CrossRef]
- van der Sluis, M.; Vincent, A.; Bouma, J.; Korteland-Van Male, A.; van Goudoever, J.B.; Renes, I.B.; Van Seuningen, I. Forkhead box transcription factors Foxa1 and Foxa2 are important regulators of Muc2 mucin expression in intestinal epithelial cells. Biochem. Biophys. Res. Commun. 2008, 369, 1108–1113. [Google Scholar] [CrossRef]
- Russo, V.E.A.; Martienssen, R.A.; Riggs, A.D. Introduction. In Epigenetic Mechanisms of Gene Regulation; Cold Spring Harbor Laboratory Press: New York, NY, USA, 1996; pp. 1–4. [Google Scholar]
- Van Seuningen, I. What is epigenetics? Strategies for research and application to cancer. Ann. Pathol. 2009, 29 (Spec No 1), S28–30. [Google Scholar] [CrossRef]
- Vincent, A.; Van Seuningen, I. Mucins, epigenetics and cancer. In The Epithelial Mucins: Structure/Function. Roles in Cancer and Inflammatory Diseases; Van Seuningen, I., Ed.; Research Signpost: Kerala, India, 2008; pp. 95–108. [Google Scholar]
- Van Seuningen, I.; Vincent, A. Mucins: A new family of epigenetic biomarkers in epithelial cancers. Expert Opinion on Medical Diagnostics 2009, 3, 411–427. [Google Scholar] [CrossRef]
- Yamada, N.; Nishida, Y.; Tsutsumida, H.; Hamada, T.; Goto, M.; Higashi, M.; Nomoto, M.; Yonezawa, S. MUC1 expression is regulated by DNA methylation and histone H3 lysine 9 modification in cancer cells. Cancer Res. 2008, 68, 2708–2716. [Google Scholar] [CrossRef]
- Vincent, A.; Ducourouble, M.P.; Van Seuningen, I. Epigenetic regulation of the human mucin gene MUC4 in epithelial cancer cell lines involves both DNA methylation and histone modifications mediated by DNA methyltransferases and histone deacetylases. Faseb. J. 2008, 22, 3035–3045. [Google Scholar] [CrossRef]
- Yamada, N.; Nishida, Y.; Tsutsumida, H.; Goto, M.; Higashi, M.; Nomoto, M.; Yonezawa, S. Promoter CpG methylation in cancer cells contributes to the regulation of MUC4. Br. J. Cancer 2009, 100, 344–351. [Google Scholar] [CrossRef]
- de Bustros, A.; Nelkin, B.D.; Silverman, A.; Ehrlich, G.; Poiesz, B.; Baylin, S.B. The short arm of chromosome 11 is a "hot spot" for hypermethylation in human neoplasia. Proc. Natl. Acad. Sci. USA 1988, 85, 5693–5697. [Google Scholar]
- Ho, J.J.; Han, S.W.; Pan, P.L.; Deng, G.; Kuan, S.F.; Kim, Y.S. Methylation status of promoters and expression of MUC2 and MUC5AC mucins in pancreatic cancer cells. Int. J. Oncol. 2003, 22, 273–279. [Google Scholar]
- Vincent, A.; Perrais, M.; Desseyn, J.L.; Aubert, J.P.; Pigny, P.; Van Seuningen, I. Epigenetic regulation (DNA methylation, histone modifications) of the 11p15 mucin genes (MUC2, MUC5AC, MUC5B, MUC6) in epithelial cancer cells. Oncogene 2007, 26, 6566–6576. [Google Scholar] [CrossRef]
- Hamada, T.; Goto, M.; Tsutsumida, H.; Nomoto, M.; Higashi, M.; Sugai, T.; Nakamura, S.; Yonezawa, S. Mapping of the methylation pattern of the MUC2 promoter in pancreatic cancer cell lines, using bisulfite genomic sequencing. Cancer Lett. 2005, 227, 175–184. [Google Scholar] [CrossRef]
- Yamada, N.; Nishida, Y.; Yokoyama, S.; Tsutsumida, H.; Houjou, I.; Kitamoto, S.; Goto, M.; Higashi, M.; Yonezawa, S. Expression of MUC5AC, an early marker of pancreatobiliary cancer, is regulated by DNA methylation in the distal promoter region in cancer cells. J. Hepatobiliary. Pancreat. Sci. 2010, 17, 844–854. [Google Scholar] [CrossRef]
- Yamada, N.; Hamada, T.; Goto, M.; Tsutsumida, H.; Higashi, M.; Nomoto, M.; Yonezawa, S. MUC2 expression is regulated by histone H3 modification and DNA methylation in pancreatic cancer. Int. J. Cancer 2006, 119, 1850–1857. [Google Scholar] [CrossRef]
- Jin, C.; Rajabi, H.; Kufe, D. miR-1226 targets expression of the mucin 1 oncoprotein and induces cell death. Int. J. Oncol. 2010, 37, 61–69. [Google Scholar]
- Rajabi, H.; Jin, C.; Ahmad, R.; McClary, C.; Joshi, M.D.; Kufe, D. Mucin 1 Oncoprotein Expression Is Suppressed by the miR-125b Oncomir. Genes Cancer 2010, 1, 62–68. [Google Scholar] [CrossRef]
- Sachdeva, M.; Mo, Y.Y. MicroRNA-145 suppresses cell invasion and metastasis by directly targeting mucin 1. Cancer Res. 2010, 70, 378–387. [Google Scholar] [CrossRef]
- Fry, L.C.; Monkemuller, K.; Malfertheiner, P. Molecular markers of pancreatic cancer: development and clinical relevance. Langenbecks Arch. Surg. 2008, 393, 883–890. [Google Scholar] [CrossRef]
- Jhala, N.; Jhala, D.; Vickers, S.M.; Eltoum, I.; Batra, S.K.; Manne, U.; Eloubeidi, M.; Jones, J.J.; Grizzle, W.E. Biomarkers in Diagnosis of pancreatic carcinoma in fine-needle aspirates. Am. J. Clin. Pathol. 2006, 126, 572–579. [Google Scholar] [CrossRef]
- Gold, D.V.; Karanjawala, Z.; Modrak, D.E.; Goldenberg, D.M.; Hruban, R.H. PAM4-reactive MUC1 is a biomarker for early pancreatic adenocarcinoma. Clin. Cancer Res. 2007, 13, 7380–7387. [Google Scholar] [CrossRef]
- Haab, B.B.; Porter, A.; Yue, T.; Li, L.; Scheiman, J.; Anderson, M.A.; Barnes, D.; Schmidt, C.M.; Feng, Z.; Simeone, D.M. Glycosylation variants of mucins and CEACAMs as candidate biomarkers for the diagnosis of pancreatic cystic neoplasms. Ann. Surg. 2010, 251, 937–945. [Google Scholar] [CrossRef]
- Tang, C.K.; Apostolopoulos, V. Strategies used for MUC1 immunotherapy: preclinical studies. Expert Rev. Vaccines 2008, 7, 951–962. [Google Scholar] [CrossRef]
- Tang, C.K.; Katsara, M.; Apostolopoulos, V. Strategies used for MUC1 immunotherapy: human clinical studies. Expert Rev. Vaccines 2008, 7, 963–975. [Google Scholar] [CrossRef]
- Bitler, B.G.; Menzl, I.; Huerta, C.L.; Sands, B.; Knowlton, W.; Chang, A.; Schroeder, J.A. Intracellular MUC1 peptides inhibit cancer progression. Clin. Cancer Res. 2009, 15, 100–109. [Google Scholar] [CrossRef]
- Chen, L.; Liu, Q.; Qin, R.; Le, H.; Xia, R.; Li, W.; Kumar, M. Amplification and functional characterization of MUC1 promoter and gene-virotherapy via a targeting adenoviral vector expressing hSSTR2 gene in MUC1-positive Panc-1 pancreatic cancer cells in vitro. Int. J. Mol. Med. 2005, 15, 617–626. [Google Scholar]
- Wei, J.; Gao, W.; Wu, J.; Meng, K.; Zhang, J.; Chen, J.; Miao, Y. Dendritic cells expressing a combined PADRE/MUC4-derived polyepitope DNA vaccine induce multiple cytotoxic T-cell responses. Cancer Biother. Radiopharm. 2008, 23, 121–128. [Google Scholar] [CrossRef]
- Bafna, S.; Kaur, S.; Momi, N.; Batra, S.K. Pancreatic cancer cells resistance to gemcitabine: the role of MUC4 mucin. Br. J. Cancer 2009, 101, 1155–1161. [Google Scholar] [CrossRef]
- Mimeault, M.; Johansson, S.L.; Senapati, S.; Momi, N.; Chakraborty, S.; Batra, S.K. MUC4 down-regulation reverses chemoresistance of pancreatic cancer stem/progenitor cells and their progenies. Cancer Lett. 2010, in press. [Google Scholar]
- Kalra, A.V.; Campbell, R.B. Mucin impedes cytotoxic effect of 5-FU against growth of human pancreatic cancer cells: overcoming cellular barriers for therapeutic gain. Br. J. Cancer 2007, 97, 910–918. [Google Scholar] [CrossRef]
- Kalra, A.V.; Campbell, R.B. Mucin overexpression limits the effectiveness of 5-FU by reducing intracellular drug uptake and antineoplastic drug effects in pancreatic tumours. Eur. J. Cancer 2008, 45, 164–173. [Google Scholar] [CrossRef]
- Tinder, T.L.; Subramani, D.B.; Basu, G.D.; Bradley, J.M.; Schettini, J.; Million, A.; Skaar, T.; Mukherjee, P. MUC1 enhances tumor progression and contributes toward immunosuppression in a mouse model of spontaneous pancreatic adenocarcinoma. J. Immunol. 2008, 181, 3116–3125. [Google Scholar]
- Spicer, A.P.; Duhig, T.; Chilton, B.S.; Gendler, S.J. Analysis of mammalian MUC1 genes reveals potential functionally important domains. Mamm. Genome 1995, 6, 885–888. [Google Scholar] [CrossRef]
- Lalani, E.N.; Berdichevsky, F.; Boshell, M.; Shearer, M.; Wilson, D.; Stauss, H.; Gendler, S.J.; Taylor-Papadimitriou, J. Expression of the gene coding for a human mucin in mouse mammary tumor cells can affect their tumorigenicity. J. Biol. Chem. 1991, 266, 15420–15426. [Google Scholar]
- Aguirre, A.J.; Bardeesy, N.; Sinha, M.; Lopez, L.; Tuveson, D.A.; Horner, J.; Redston, M.S.; DePinho, R.A. Activated Kras and Ink4a/Arf deficiency cooperate to produce metastatic pancreatic ductal adenocarcinoma. Genes Dev. 2003, 17, 3112–3126. [Google Scholar] [CrossRef]
- Hingorani, S.R.; Petricoin, E.F.; Maitra, A.; Rajapakse, V.; King, C.; Jacobetz, M.A.; Ross, S.; Conrads, T.P.; Veenstra, T.D.; Hitt, B.A.; Kawaguchi, Y.; Johann, D.; Liotta, L.A.; Crawford, H.C.; Putt, M.E.; Jacks, T.; Wright, C.V.; Hruban, R.H.; Lowy, A.M.; Tuveson, D.A. Preinvasive and invasive ductal pancreatic cancer and its early detection in the mouse. Cancer Cell 2003, 4, 437–450. [Google Scholar] [CrossRef]
- Hingorani, S.R.; Wang, L.; Multani, A.S.; Combs, C.; Deramaudt, T.B.; Hruban, R.H.; Rustgi, A.K.; Chang, S.; Tuveson, D.A. Trp53R172H and KrasG12D cooperate to promote chromosomal instability and widely metastatic pancreatic ductal adenocarcinoma in mice. Cancer Cell 2005, 7, 469–483. [Google Scholar] [CrossRef]
- Izeradjene, K.; Combs, C.; Best, M.; Gopinathan, A.; Wagner, A.; Grady, W.M.; Deng, C.X.; Hruban, R.H.; Adsay, N.V.; Tuveson, D.A.; Hingorani, S.R. Kras(G12D) and Smad4/Dpc4 haploinsufficiency cooperate to induce mucinous cystic neoplasms and invasive adenocarcinoma of the pancreas. Cancer Cell 2007, 11, 229–243. [Google Scholar] [CrossRef]
- Velcich, A.; Yang, W.; Heyer, J.; Fragale, A.; Nicholas, C.; Viani, S.; Kucherlapati, R.; Lipkin, M.; Yang, K.; Augenlicht, L. Colorectal cancer in mice genetically deficient in the mucin Muc2. Science 2002, 295, 1726–1729. [Google Scholar] [CrossRef]
- Cheon, D.J.; Wang, Y.; Deng, J.M.; Lu, Z.; Xiao, L.; Chen, C.M.; Bast, R.C.; Behringer, R.R. CA125/MUC16 is dispensable for mouse development and reproduction. PLoS One 2009, 4, e4675. [Google Scholar]
© 2010 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
MDPI and ACS Style
Jonckheere, N.; Skrypek, N.; Van Seuningen, I. Mucins and Pancreatic Cancer. Cancers 2010, 2, 1794-1812. https://doi.org/10.3390/cancers2041794
AMA Style
Jonckheere N, Skrypek N, Van Seuningen I. Mucins and Pancreatic Cancer. Cancers. 2010; 2(4):1794-1812. https://doi.org/10.3390/cancers2041794
Chicago/Turabian StyleJonckheere, Nicolas, Nicolas Skrypek, and Isabelle Van Seuningen. 2010. "Mucins and Pancreatic Cancer" Cancers 2, no. 4: 1794-1812. https://doi.org/10.3390/cancers2041794