Recently Identified Biomarkers That Promote Lymph Node Metastasis in Head and Neck Squamous Cell Carcinoma

Abstract
: Head and neck squamous cell carcinoma (HNSCC) is a heterogeneous cancer that arises in the upper aerodigestive tract. Despite advances in knowledge and treatment of this disease, the five-year survival rate after diagnosis of advanced (stage 3 and 4) HNSCC remains approximately 50%. One reason for the large degree of mortality associated with late stage HNSCC is the intrinsic ability of tumor cells to undergo locoregional invasion. Lymph nodes in the cervical region are the primary sites of metastasis for HNSCC, occurring before the formation of distant metastases. The presence of lymph node metastases is strongly associated with poor patient outcome, resulting in increased consideration being given to the development and implementation of anti-invasive strategies. In this review, we focus on select proteins that have been recently identified as promoters of lymph node metastasis in HNSCC. The discussed proteins are involved in a wide range of critical cellular functions, and offer a more comprehensive understanding of the factors involved in HNSCC metastasis while additionally providing increased options for consideration in the design of future therapeutic intervention strategies.1. Introduction
The main cause of cancer-related death is due to metastasis of primary tumors to secondary sites within the body. Advanced cases of head and neck squamous carcinoma (HNSCC) primarily spread locoregionally, where tumor cells infiltrate the lymphatic drainage and travel into cervical lymph nodes [1]. While incidence rates and overall disease-related deaths have dropped over recent years [2], the five-year survival rate for patients presenting with clinically advanced disease remains around 55% [3]. Predicting the inherent metastatic potential of primary HNSCC tumors would serve to aid in enhancing approaches to treatment that would improve patient management. However, current diagnosing strategies rely in part on histological analyses of biopsy samples, which have largely proven inadequate due to the high frequency of patients with recurrent disease [4]. The presence of lymph node metastasis in HNSCC patients has long been established as a poor prognostic indicator [5-7], making earlier detection of tumors with the propensity to invade and spread through local lymphatics an important step in patient management towards a more promising outcome. The stepwise model of carcinoma progression involves changes at the molecular level that ultimately provide normal epithelia cells with the ability to invade surrounding tissue [8]. In HNSCC, efforts at identifying molecules associated with and responsible for driving nodal metastasis has revealed many potential biomarkers for this process over the past decade [4,9-11]. Some of the more notable markers associated with nodal metastasis are cell cycle and proliferation regulators such as p53, epidermal growth factor receptor (EGFR), p16, and cyclin D1. The roles of these proteins in HNSCC development and progression are well documented [4,11-13,13]. More recently, expression profiling through DNA microarray technology has been useful in identifying genes previously unrecognized in the field that also contribute to or are associated with lymph node metastasis [14-16]. In this review, we focus on various studies conducted within the past four years that have linked overexpression of specific proteins in HNSCC to lymph node metastasis, highlighting several new potential candidates (Table 1) that could prove useful in the prediction, detection and treatment determination of metastatic disease.
2. Proteins Involved in Cell Cycle Regulation, Proliferation and Apoptosis
Regulation of the cell cycle requires the coordination of many protein classes, creating a system of checks and balances that when dysregulated results in either cell proliferation or death. Mutations or altered expression of proteins important for proper governing of cell cycle initiation and coordination can set the system off balance, providing tumor cells with means to bypass normal cell cycle check points, evade apoptosis and over-proliferate. Several proteins involved in cell cycle regulation have recently been identified as markers with increased expression in HNSCC that correspond with lymph node metastasis.
2.1. c-Met
The receptor tyrosine kinase c-Met is normally active during embryonic development and wound healing [17]. Activated c-Met promotes cellular proliferation by organizing an “invasive growth” program by which normal cells migrate to new sites to form polarized cells and functional 3D structures. [18]. The ligand for c-Met is hepatocyte growth factor (HGF). HGF is secreted by mesenchymal cells and upon binding to c-Met causes receptor homodimerization and phosphorylation/activation of the catalytic site, resulting in downstream signaling events that lead to cell transformation and invasion [17,19]. Expression of c-Met on epithelial cells enables them to receive signals from surrounding stromal cells through stromal cell HGF secretion. In transformed cells, c-Met activity is enhanced through several mechanisms, including increased ligand-based stimulation via elevated autocrine or paracrine HGF secretion, receptor overexpression or mutational activation of the c-Met kinase domain [17,19]. One notable function of c-Met is its interaction with the oncogenic tyrosine kinase c-Src, creating a mechanism to bypass inhibited EGFR signaling in breast cancer cell lines [20] as well as increasing resistance to c-Src inhibitory drugs in HNSCC [21]. Both c-Src and EGFR are overexpressed in HNSCC and are important in head and neck cancer development and progression [22]. A retrospective analysis of 61 surgically treated cases of HNSCC showed frequent expression of c-Met in tumors with higher T-stage classification [23]. Patients with lymph node metastasis have a significant increase in c-Met expression when compared with indolent cases lacking metastasis. While HGF was found at elevated levels in over 60% of the cases, it was not a significant factor in this study; however when combined with c-Met overexpression there was a correlation with lymph node metastasis. Being a paracrine factor secreted by cells of mesenchymal origin, HGF may be more relevant as a serum biomarker, as shown in previous studies [24-26]. In these prospective studies, serum cytokine levels were measured in patients before and after treatment. High levels of serum HGF were found in patients before treatment, decreased after treatment, and were found to increase again with recurrence. These results suggest monitoring HGF levels may prove useful in determining response to treatment and recurrence. The HGF/c-Met pathway has been implicated in invasion and metastasis in HNSCC and other cancers, both in vitro and in vivo [27-31] and has been linked to resistance of EGFR inhibitors and cisplatin [20,32-35], making both c-Met and HGF attractive drug targets as well as determinants of treatment. Currently there are several clinical trials involving drugs that target c-Met or HGF specifically, although these trials have not gone past phase I/II [36].
2.2. CEP55 (FLJ10540)
The cytokinesis regulator CEP55, also known as FLJ10540, is a 55 kDa protein that localizes to the centrosome of chromosomes in interphase and the midbody during cytokinesis, where it mediates the final stages of mitotic division into two daughter cells [37]. CEP55 is a recently identified downstream target of the oncogene FOXM1, which has been shown to be upregulated in pre-malignant HNSCC lesions [38]. Subsequently, CEP55 overexpression has been directly correlated with an increase in tumor aggressiveness in oral squamous cell carcinoma (OSCC) [39]. Retrospective immunohistochemistry (IHC) analysis revealed overexpression in patient tumor samples, which was linked to tumor and nodal stage as well as a poor prognosis [39]. There was also significantly higher expression in patients with advanced T stage (3 and 4) with lymph node metastasis when compared with node negative and stage 1-2 tumors. In vitro work has linked CEP55 expression to increased cell motility and invasion through regulation of FOXM1 and MMP-2 [39]. In another report, while CEP55 was shown to be significantly upregulated in dysplasias and HNSCC, upregulation within lymph node metastases was not significant, which the authors cite as being due to tissue heterogeneity [40]. Taken together, these results suggest CEP55 may prove useful in predicting disease progression. As cytokinesis is of obvious importance to highly proliferative cells, overexpression of CEP55 is therefore a logical candidate for potential use as an HNSCC metastatic biomarker in clinical settings.
2.3. NBS1
Nijmegen breakage syndrome (NBS) is a syndrome characterized by growth retardation, immunodeficiencies and predisposition to malignancies [41]. The only gene associated with this syndrome is NBS1, and its gene product plays an important cell cycle checkpoint role in double strand DNA break repair [41]. NBS1 is part of a complex including Mre11 and Rad5 (MRN complex) that is central to detection of DNA breakage, coordinating response programs for and catalyzing repair mechanisms of double-strand breaks [42]. A study analyzing OSCC samples revealed an increase in NBS1 mRNA expression that correlated to increased protein expression [43]. NBS1 overexpression was associated with advanced disease and recurrence/metastasis in OSCC, while non-oral HNSCC samples with the same levels of expression were only associated with recurrence. An increase in NBS1 expression in HNSCC regardless of origin site was additionally associated with lymph node involvement [43]. In this same study, NBS1 was found to be a prognostic marker even with samples divided into subgroups based on tumor and nodal stage or treatment type. Earlier studies by the authors had linked NBS1 overexpression to more aggressive disease and worse prognosis in advanced HNSCC [44], and to lymph node and distant metastasis [45]. These studies also determined NBS1 expression to be involved in cellular transformation through activation of the PI3K/Akt pathway and induction of EMT [44,45]. One explanation for this association could be due to single nucleotide polymorphisms (SNPs) within the NBS1 gene. Several studies have linked genetic variations to development of cancers of the breast, lung, esophagus, non-Hodgkin's lymphoma and upper aerodigestive tract [46-48]. Identifying high risk patients through detection of NBS1 SNPs may be a useful tool in predicting patient outcome in OSCC and other HNSCC subtypes.
2.4. Survivin
An inhibitor of apoptosis (IAP) family member, survivin, suppresses apoptosis by directly binding to and inhibiting caspase family members, typically caspase 3 and caspase 7, or by indirectly suppressing apoptosis through activation of caspase-associated cofactors [49,50]. Survivin overexpression has been identified in different cancer types, suggesting it may be a tumor marker and possible drug target [49,50]. There have been multiple studies linking survivin expression to HNSCC progression in recent years. In a retrospective study of 42 OSCC cases, individuals with lymph node metastasis had significantly high survivin expression compared to non-metastatic cases [51]. A correlation between expression and low survival rate was also concluded in this study. A significantly higher expression of survivin was found in another study of OSCC cases [52]. Premalignant lesions were also included, and survivin expression was elevated in these regions when compared to normal tissue. Survivin has been found to play different roles depending on location within the cell [49]. In the nucleus, survivin controls cell division by functioning as a subunit of the chromosomal passenger complex (CPC), while cytoplasmic survivin is cytoprotective, providing the cell with protection from cell death induced by radiation or chemotherapeutic drugs [53]. An examination of normal and HNSCC tissues by IHC showed nuclear and cytoplasmic staining, both which were significantly correlated to poor differentiation and lymph node metastasis [54]. There have been several other studies that investigated the prognostic role of cytoplasmic and nuclear survivin. Although no general consensus was found based on subcellular location, survivin expression has consistently been linked to unfavorable outcome and reduced disease-free survival. [55-57]. The anti-apoptotic effects of survivin may be linked to treatment failure as some in vitro studies have suggested [58-60]. In one particular model, HNSCC cells that escaped cellular senescence after treatment with the chemotherapeutic drug camptothecin were unable to escape senescence upon knockdown of survivin [61]. Another retrospective study analyzed OSCC cases and correlated survivin mRNA expression to tumor differentiation, stage and lymph node involvement. This study also found that down-regulation of survivin increased sensitivity of HNSCC cells to cisplatin [62]. While more studies are needed to firmly link survivin overexpression to lymph node metastasis, its role as a prognostic and drug resistance marker warrants further investigation as a potential therapeutic target.
2.5. RSK2
p90 ribosomal S6 kinase 2 (RSK2) is a serine/threonine kinase activated downstream in the MAPK pathway [63,64]. Numerous substrates have been identified for RSK2, including GSK3β, c-Fos, p27kip1, e1F4B, and p65, a subunit of NF-κB. These substrates link RSK2 activity in mediating pathways central to cell proliferation, transcriptional and translational regulation, survival and apoptosis [63,64]. Retrospective IHC analysis has identified RSK2 overexpression in HNSCC tumor and lymph node patient samples [65]. Primary tumors from patients with lymph node metastases and matched lymph node specimens had a significantly higher expression of RSK2 than patients with non-metastatic primary HNSCC. Manipulation of RSK2 levels by RNA interference demonstrates a clear dependence of RSK2 expression levels in modulating invasion in Matrigel transwell assays. Similar results were obtained in a xenograft mouse model, where cells with stable knockdown of RSK2 had less metastatic potential than controls [65]. Previous in vitro work identified RSK2 as a critical regulator in cellular transformation [66] and though not extensively studied in head and neck cancer, the versatility of RSK2 makes it a worthwhile target for additional investigation. Multiple small molecule inhibitors specific to RSK family members have been identified [67] that may serve as viable platforms for further development of potential targeted anti-RSK2 therapeutic compounds.
3. Cell Motility, Adhesion and Extracellular Matrix Degradation
Without the ability to modify their surroundings and move to new areas, tumor cells would remain in their primary location, making them more manageable by conventional surgical and radiation-based treatment regimes. Overexpression of proteins involved in cell motility and extracellular matrix remodeling equips primary HNSCC cells with the ability to degrade and escape an encapsulating extracellular matrix and penetrate through the surrounding stroma, ultimately resulting in lymphatic intravasation and spread into regional lymph nodes.
3.1. Cortactin
Cortactin is an actin binding protein that plays roles in cell motility and invasion by promoting Arp2/3 complex actin nucleation and by stabilizing the newly formed actin branchpoints [68]. Located at chromosomal region 11q13, an area frequently amplified in HNSCC, cortactin has consistently been associated with more aggressive and invasive tumors, lymph node metastasis and poor clinical outcome in HNSCC [69-73]. Yamada et al. analyzed a series of OSCC patient biopsy samples, finding overexpression of cortactin more often in OSCC than in normal epithelium, as well as localization of cortactin at the invasive front [74]. Cortactin overexpression was also found more frequently in tumors with high T and N classification and significantly correlated to regional invasion in these patients [74]. A separate study looked at the relationship between CTTN gene amplification status, mRNA and protein expression in patients with pharyngeal or laryngeal SCC [75]. This study found that gene amplification correlated significantly to mRNA and protein expression, with cases containing strong cortactin staining significantly associated with lymph node metastasis. This group conducted an analysis of CTTN gene amplification, comparing amplification within epithelial hyperplastic/dysplastic lesions and high-grade dysplasias/carcinoma in situ, to determine timing of amplification. Consistent with studies of 11q13 gene amplification being a later stage event [4], CTTN was only detected in the higher grade lesions [75]. Cortactin expression was also studied along with EGFR status in a series of HNSCC samples of different origins [76]. Cortactin overexpression correlated to higher TNM stage, histologic grade and was associated with decreased overall survival and increased local recurrence. However, patients that had both cortactin and EGFR overexpression did not have a different survival rate than those with cortactin only. This is surprising due to evidence that cortactin overexpression contributes to sustained EGFR surface expression by preventing ligand-mediated receptor downregulation [77] suggesting cortactin overexpression can be uncoupled from EGFR as reported in a subsequent study [78]. Cortactin overexpression in HNSCC does enhance c-Met surface expression, providing an additional mechanism for sustained c-Met signaling [77]. Another study focusing on laryngeal SCC found cortactin expression to be linked to both lymph node and distant metastases, and was identified as a predictor of poor prognosis in this HNSCC subtype [79]. The collective results from these recent studies strongly indicate that cortactin gene amplification and/or protein overexpression increases HNSCC aggressiveness. Although a late stage player in HNSCC progression, monitoring cortactin expression in HNSCC may also be useful in predicting invasive carcinoma and tumor recurrence.
3.2. CD44
Cell adhesion molecules (CAMs) are necessary for cell-cell or cell-extracellular matrix (ECM) contacts. The transmembrane glycoprotein CD44 is a CAM that binds hyaluronan (HA), a glycosaminoglycan component of the ECM and the primary ligand for CD44. The CD44 protein family consists of various isoforms that are the result of alternative splicing of exons 6-14 [80]. CD44 variants are often overexpressed in different cancer types, with overexpression correlating to poor patient outcome [80]. Standard CD44 (CD44s) and three variants, v3, v6 and v10, are overexpressed in HNSCC [81]. CD44s was overexpressed at a higher frequency in supraglottic laryngeal carcinomas of patients that were lymph node positive, although no significant differences were found between tumor stages and differentiation and CD44 expression [82]. The CD44 variants v3, v6 and v10 were identified in HNSCC samples from the oral cavity, oropharynx or larynx, with strong staining by IHC in both primary and lymph node metastases [81]. CD44 v3 and v6 were both associated with advanced T stage, while a strong v3 expression within primary tumors was related to lymph node metastasis and v10 expression related to distant metastases [81]. The same study also found CD44 mediated sensitivity to cisplatin in vitro and CD44 knockdown increased HNSCC cell death. Another interesting study analyzed blood for CD44 mRNA from patients with advanced HNSCC who had been treated with chemotherapy and radiotherapy to determine if there the presence of CD44 mRNA correlated with prognosis during the patient follow up period [83]. Quantitative RT-PCR detected mRNA in peripheral blood from patients and healthy volunteers, and elevated levels of CD44 mRNA in HNSCC patients correlated with the degree of lymph node involvement and recurrence. IHC analysis of tumor samples confirmed CD44 protein expression for all patients. These studies suggest a role for CD44 and specific splice variant isoforms in the regional and distant spread of HNSCC, with evidence for pre-treatment screening of CD44 being beneficial to determining prognosis and drug response.
3.3. Matrix Metalloproteinases
Invasion and metastasis of tumor cells requires proteolysis of the basement membrane and surrounding ECM. Matrix metalloproteinases (MMPs) provide cells with a mechanism to modulate the microenvironment through the breakdown of ECM molecules present in basement membranes and stroma [84]. Twenty three different MMPs are expressed in humans, with MMP-9 and MMP-14 (MT1-MMP) accepted as playing critical roles in HNSCC invasion and metastasis. MMP-9 is a secreted proteinase that utilizes CD44 as a docking site, allowing for its retention on the cell surface [84,85]. CD44s expression in supraglottic laryngeal carcinoma samples correlates with MMP-9 expression in lymph node positive patients [82]. Similar results were obtained in a second study of OSCC, linking MMP-9 expression to lymph node metastasis [86]. Expression of MMP-9 was also found to co-localize to the invasive front with CD44 of HNSCC patient samples, while normal mucosa showed little to no MMP-9 labeling [87]. A prospective study investigated whether serum MMP-9 levels of 161 patients with OSCC could be correlated to clinicopathological parameters [88]. Pre-treatment serum MMP-9 levels correlated to clinical stage and were also significantly higher in patients with lymph node metastasis, while a significant decrease in levels was seen after surgery. Presurgery levels of MMP-9 in patients who died during the study were found to be significantly higher than those who survived, linking serum MMP-9 to patient outcome [88]. The results of this study are comparable to an earlier study from 2005, which also investigated serum levels of MMP-9 in HNSCC patients before surgery [89]. Transmembrane MMPs are also important for the activation of secreted MMPs through the cleavage of secreted MMP proforms to generate a functional extracellular enzyme. Studies on supraglottic HNSCC patients demonstrate increased MT1-MMP expression compared to normal tissue. In this study, the level of MT1-MMP overexpression correlated to the depth of invasion, presence of lymph node metastasis and advanced clinical stage [90]. Patients with high MT1-MMP expression also had a poor prognosis. Interestingly, cortactin overexpression has been reported to enhance MMP-9 secretion and promote MT1-MMP surface expression in HNSCC cell lines, resulting in enhanced ECM degradation at plasma membrane structures known as invadopodia [91,92]. The pro-invasive function of combined MMP activity in HNSCC and other tumor types makes them useful as prognostic markers as well as attractive and important therapeutic targets. Despite previous failed attempts at targeted drug therapy [93,94], development of new generation MMP inhibitors is being pursued, where improved drug and trial design may yet result in the production of selective and effective anti-invasive drugs [94].
4. Tumor Microenvironment and Angiogenesis
HNSCC and other tumor types frequently exploit signals generated from cellular and non-cellular ECM components to promote tumor growth and dissemination. The process of new blood vessel formation (angiogenesis) is initiated in order to provide tumors with the means to supply nutrients to facilitate their growth, as well providing avenues for eventual metastasis through the vascular system.
4.1. Chemokines
Chemokines are small peptides that upon receptor binding act as chemoattractants, homing leukocytes to areas of inflammation [95,96]. Their roles in cell trafficking and angiogenesis help promote tumor growth, as evidenced by their overexpression in several different human cancers. The receptor CXCR4 and its ligand CXCL12 (also called SDF-1) is one chemokine pathway exploited by metastatic HNSCC [97]. A retrospective analysis by IHC of 30 patients with laryngeal and hypopharyngeal SCC showed a significant increase in CXCR4 expression in patients with positive lymph node and distant metastases compared to patients lacking metastatic disease [98]. This study also looked at CXCL12 expression and while higher in patients with metastasis, it was not statistically significant. Two other studies assessing the prognostic value of CXCR4 in OSCC also drew similar conclusions, finding a significant association of expression with lymph node metastasis [86,99]. One study examined expression of CXCR4 and another chemokine receptor, CCR7, which has been shown to activate the PI3K/Akt pathway in HNSCC, a pathway involved in cell growth, differentiation and survival [100]. Both chemokines were expressed at significantly higher levels in those patients with positive lymph node involvement compared with lymph node negative cases [100]. One notable difference was that CCR7 expression correlated to cases with advanced tumor stage, while CXCR4 was significantly higher in patients with distant metastases. A similar study also significantly associated CCR7 expression with lymph node metastasis, while CXCR4 expression was associated but not statistically significant [101]. CCR7 also positively associated with lymph node metastasis in patients with tonsillar SCC [102] and OSCC [103]. The influence of autocrine/paracrine activation of CCR7 was examined in another retrospective study of HNSCC and found higher mRNA expression of CCR7 and its ligands CCL19 and CCL21 in metastatic lymph nodes [104]. This study further concluded from additional in vitro and orthotopic mouse model studies that blockage of CCR7 impaired tumor cell proliferation and decreased resistance to cisplatin-induced apoptosis. The impact of chemokines and their receptors on HNSCC may be explained in part by their influence on MMPs [95,105]. CXCR4 increased HNSCC cellular invasion in vitro by upregulating expression of MMP-9 and MMP-13 [98,106]. High CXCR4 levels also correlate with high MMP-9 in OSCC patient samples [86]. CCR7 is typically involved in directing dendritic cells to peripheral lymph nodes [105]. The frequent expression of CCR7 in HNSCC and association with lymph node metastasis suggests that CCR7 expression provides tumor cells with a mechanism for direct lymph node infiltration [95,96,107,108]. These results suggest overlapping roles for chemokine receptors in HNSCC progression to metastatic disease, a consideration that may need to be taken into account for any future therapeutic targeting strategies.
4.2. VEGF/R
Vascular endothelial growth factor (VEGF) is a cytokine expressed by tumors that plays a key role in angiogenesis. VEGF performs its function through binding to one of the VEGF receptor family members, with VEGFR2 serving as the major receptor subtype in several different neoplasms [109]. The importance of VEGF in cancer progression has been well documented [110-114], and recent studies further demonstrate VEGF as a valuable prognostic marker for HNSCC. As a growth factor, circulating VEGF can be a useful marker for detecting advanced disease since circulating serum levels of VEGF in HNSCC patients before treatment was significantly higher when compared with non-cancerous individuals [115]. Patients with advanced T stage, lymph node metastasis and advanced disease stage also had significantly higher serum VEGF levels [115]. Another study involving the prognostic value of serum VEGF levels in nasopharyngeal carcinoma found a significant relationship between higher levels of VEGF and several clinicopathologic parameters, including T and N stage and distant metastasis [116]. Similar results were obtained in another prospective serum analysis of OSCC patients before and after treatment [88]. In a separate retrospective study, OSCC cases were analyzed for VEGF-C and VEGF-D expression by IHC [117], where increased staining intensity for these ligands significantly corresponded with lymph node involvement. In addition, lymphatic vessel density (LVD) was also evaluated in this report, where high LVD correlated with VEGF-C/VEGF-D expression. Another study evaluated the relationship between lymph node metastasis and VEGF. Positive IHC expression of VEGF and Notch1, a receptor capable of promoting transcription of genes involved in cellular proliferation [118] was observed in patients with early SCC of the tongue. High Notch1 expression was also more frequent in patients with lymph node involvement. Another analyzed variable in this study was the distance of tumor cell invasion from the surface mucosa. A greater invasion depth was found in patients with elevated VEGF expression compared with cases containing normal VEGF levels [110]. In a separate study focusing on the role of angiogenesis in early SCC of the tongue, VEGF expression was found in 74% of analyzed patient samples, where it correlated with increased tumor size, disease stage, lymph node invasion, tumor recurrence and distant metastases [119]. Retrospective mRNA analysis of VEGF-C and VEGFR3 in locoregionally relapsed HNSCC revealed a significant association of high mRNA levels and relapse beyond the primary tumor [120]. While the majority of studies on VEGF expression supports a pro-metastatic role, there have been some reports where such a correlation between VEGF expression and advanced tumor stage is not evident [121,122]. However, only laryngeal SCC was analyzed in one of these studies, with the data suggesting patients with advanced disease and lower VEGF expression would benefit the most from induction chemotherapy [121]. In a separate report, VEGF expression did not predict metastasis in early (T1 or 2) stage OSCC [122]. Collectively these studies suggest VEGF and/or VEGFR status is useful as a pro-metastatic marker in HNSCC, and may also serve to predict relapse and treatment response.
5. Transcription Factors
Transcription factors are required for normal cellular homeostasis. Dysregulation of transcription factor expression is a major contributor in initiating cancer and driving tumor progression. Several transcription factors have been found to regulate expression of target genes involved in promoting HNSCC lymph node metastasis. The pleiotropic effect of these transcriptional regulators on diverse signaling pathways makes them rational targets for therapeutic intervention in HNSCC and other cancer types.
5.1. NF-κB
Nuclear factor-kappa B (NF-κB) is part of a family of transcription factors that regulate genes needed for most aspects of neoplastic transformation [123,124]. Inflammation has been linked to cancer progression [125], and as a proinflammatory transcription factor NF-κB is often found expressed in most tumor types [124]. Specific to HNSCC, analysis of tumors from varying primary sites as well as matched lymph node metastases showed positive nuclear NF-κB expression, and was found with a greater significant frequency in primary tumors with metastasis [126]. NF-κB expression levels were highest in the nodal metastases evaluated in this study. A study of laryngeal cancer patients also reported a connection between NF-κB expression and lymph node metastasis, as well as T stage and overall survival [127]. A retrospective study of early-stage laryngeal cancer correlated NF-κB expression to local recurrence in patients resistant to radiotherapy [128]. In addition, patients with recurrence and positive NF-κB expression in pretreatment tumors showed enhanced expression in recurrent tumors, while those with recurrence but without expression before treatment became NF-κB-positive. NF-κB has been shown to regulate other proteins involved in HNSCC cellular proliferation and metastasis, such as survivin [55], Twist1 [129], Snail [130], VEGF and MMP-9 [126], as well as other targets in HSNCC [123,124]. It has also been demonstrated that NF-κB expression can be regulated by chemokines or interact with other transcriptional regulators (Hif-1α) [131-133], making NF-κB a central player in the development and spread of HNSCC. Therapeutic targeting of NF-κB would therefore potentially disrupt multiple pathways important in HSNCC.
5.2. Regulators of EMT
Epithelial-mesenchymal transition (EMT) is a process in which cells lose epithelial traits and obtain a mesenchymal phenotype. While a normal part of embryonic development, EMT is a major mechanism that drives cancer development and progression. Several proteins involved in the induction of EMT in cancer have been identified [134,135]. Among these are the transcription factors Twist and Snail. Twist belongs to the basic helix-loop-helix family of transcriptional regulatory proteins [136]. Upregulation of Twist expression has been shown to promote EMT in breast cancer, while downregulation suppresses metastasis [137]. In HNSCC, Twist has been correlated to lymph node metastasis through tissue microarray screening [101]. Higher Twist expression was also observed in metastatic samples when compared to primary tumors, significantly correlating with reduced survival [138]. Another study correlated high tumor grade to Twist1 expression in HNSCC cases and while not statistically significant, Twist1 expression was associated with poor prognosis [139]. Several in vitro studies have implicated Twist expression in the acquisition of chemotherapeutic resistance for various cancer types [90,129,140]. One of these was nasopharyngeal carcinoma, where decreased Twist expression by RNAi enhances sensitivity to chemotherapeutic compounds such as taxol and cisplatin [90,129,140].
The transcription factor Snail is a zinc-finger transcriptional repressor that induces EMT by suppressing expression of E-cadherin, a component of adherens junctions that maintains epithelial cell-cell adhesion [136]. Expression of Snail was also found at higher levels in metastatic HNSCC samples [138] and was positively associated with higher-grade tumors, lower survival rates [141,142], increased invasion depth and development of metastases [142]. Additionally, another earlier retrospective study found higher Snail expression correlated with cervical lymph node and distant metastasis. When co-expressed with NBS1, Snail expression resulted in higher probability of metastasis and shorter survival periods [45]. Like Twist, Snail has also been associated with chemoresistance [143]. Snail promoted cisplatin resistance in HNSCC cell lines via upregulation of excision repair cross-complementation group 1 (ERCC1), a protein important in nucleotide excision. IHC analysis of HNSCC patients who had undergone cisplatin treatment revealed a higher risk of resistance with Snail expression. Twist1 was also evaluated in this study and was correlated with greater resistance [143]. Given their function in HNSCC and other tumor types, Twist1 and Snail expression levels are likely good candidates for monitoring the invasive and metastatic potential of primary HNSCC, and may be useful in predicting patient response to chemotherapy.
5.3. Hif-1α
In response to the low oxygen (hypoxic) environment present in primary tumors as they proliferate and increase in size, tumor cells activate hypoxia-inducible factor 1 (HIF-1) to upregulate proteins necessary for preventing cell death [144,145]. HIF-1 is a heterodimeric protein consisting of an alpha and a beta subunit [144]. The HIF-1 alpha subunit is needed for HIF-1 to function as a transcription factor, and responds to cellular oxygen levels by activating transcription of genes such as VEGF, platelet-derived growth factor (PDGF) and transforming growth factor-α (TGF-α) to survive under hypoxic conditions [144]. In OSCC, high HIF-1 alpha expression was correlated to worse outcome in metastatic OSCC samples [138,146]. Overexpression of HIF-1 alpha was frequently observed in HNSCC patients with lymph node metastasis, and was significantly higher when compared with node-negative cases [146]. Hypoxic tumor cells are known to influence other factors required for HNSCC cell survival and growth including EMT, ECM invasion and angiogenesis [147]. HIF-1 alpha performs these functions through regulation of expression and/or activity of multiple proteins, including Twist, MMP-2, MMP-9, VEGF and CXCR4/SDF-1 [147,148]. Hypoxia and HIF-1 alpha have been cited as one of the major causes of drug resistance to anti-angiogenic therapies in many human cancers [149]. These studies suggest that HIF-1 alpha expression may be a potential candidate to serve as a pro-metastatic biomarker in HNSCC and also predict treatment response.
6. HPV and HNSCC
Increased risk of HNSCC has largely been attributed to tobacco exposure and alcohol use; however in recent years, studies have also implicated human papillomavirus (HPV) infection as an additional risk [4,150,151]. Approximately 20-25% of HNSCC are HPV-positive, with the majority of these cases arising in the oropharynx [4,150,151]. The HPV proteins E6 and E7 are key players in carcinogenesis, causing the destabilization and degradation of cell cycle regulators p53 and pRb [150,151]. As a result, deregulation of cell cycle checkpoints and downregulation of other cell cycle regulatory proteins occurs, leading to genomic instability and uncontrolled proliferation [150,151]. There are multiple differences between HPV-positive and HPV-negative HNSCC, making them clinically distinct and requiring different management strategies. In HPV-positive HNSCC cases, genome-wide alterations in DNA copy number that are frequent in HPV-negative tumors are fewer in number; there are also fewer TP53 mutations, and epigenetic changes such as p16INK4A gene silencing due to downstream Rb degradation [4,151]. Another major difference is in cell cycle regulatory pathways, with upregulation of cyclins D and E being a common occurrence in HPV-positive tumors. An important distinction to note is the expression of p21WAF1/Cip1, a cell cycle regulator that normally functions to promote cell cycle arrest through binding and interfering with cyclin-dependent kinases (CDKs) 1 and 2, as well as proliferating cell nuclear antigen (PCNA) [152] to produce cell senescence. p21WAF1/Cip1 also promotes resistance to apoptosis [153] and is either up or down regulated depending on cancer type, giving it dual oncogenic and tumor suppressor properties [152]. In a retrospective study involving 117 patient samples, p21WAF1/Cip1 was overexpressed in HNSCC samples from pharynx and larynx, and its increased expression was correlated with lymph node metastasis, locoregional relapse and decreased survival rate [154]. HPV status was not determined in this report. In a more definitive study of HPV-positive HNSCC, p21WAF1/Cip1 overexpression was associated with favorable outcome [155]. In general, HPV-positive cancers have better prognosis and response to radio- and chemotherapies, with fewer distant metastases when there has been no tobacco exposure [4,151,156]. For these reasons, HPV-positive HNSCC should be treated as a separate subtype with specific biomarkers and warrants determination of HPV as standard practice in order to accurately format appropriate treatment strategies.
7. Conclusions
Development of lymph node metastases remains a major prognostic factor in HNSCC. Many patients present with clinically advanced disease, where surgery, radiation and chemotherapy are the standard of care. Given the highly disfigurative nature of HNSCC surgical treatment and typically repeated exposure to high-dose radiation, identification of primary HNSCC tumors with enhanced metastatic potential by molecular means can aid clinicians in tailoring appropriate treatment strategies, especially in cases that have no apparent nodal involvement. Current histological procedures can be limited in their ability to detect nodal metastasis, highlighting a need for detailed, accurate molecular analysis of individual HNSCC tumors to determine specific deleterious protein expression patterns. Such a molecular analysis would theoretically result in an increased ability to identify patients with a greater risk for metastasis formation, allowing for rational treatment design to be tailored for the best possible patient outcome [157].
Early detection of oral premalignant lesions is one route to improving patient prognosis. Markers predicting progression of these lesions to cancer have not been extensively studied, however a recent gene expression profiling study revealed a signature useful in predicting OSCC development [158]. The biomarkers of focus here have been more extensively characterized in tumor spread beyond the primary site, however it would be interesting to find out if they could also be useful in identifying precancerous lesions at risk for progression to carcinoma. Some of these proteins are more useful in later stages of HNSCC. Expression of cortactin, CD44, NBS1, CXCR4, Snail and VEGF in patients with metastatic disease has been correlated to the development of distant metastases. These patients may benefit from induction and maintenance therapy in order to prevent spread below the clavicles. Monitoring serum levels of HGF, MMPs and VEGF has also been shown to be beneficial in predicting patient outcome, and further studies involving prospective analyses could provide an easier route to identification of patients at higher risk for metastasis.
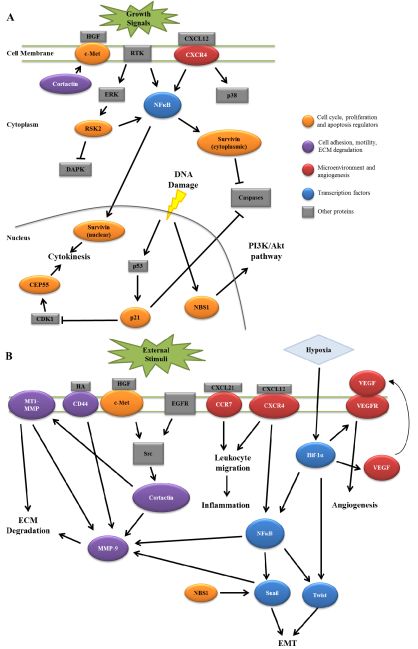
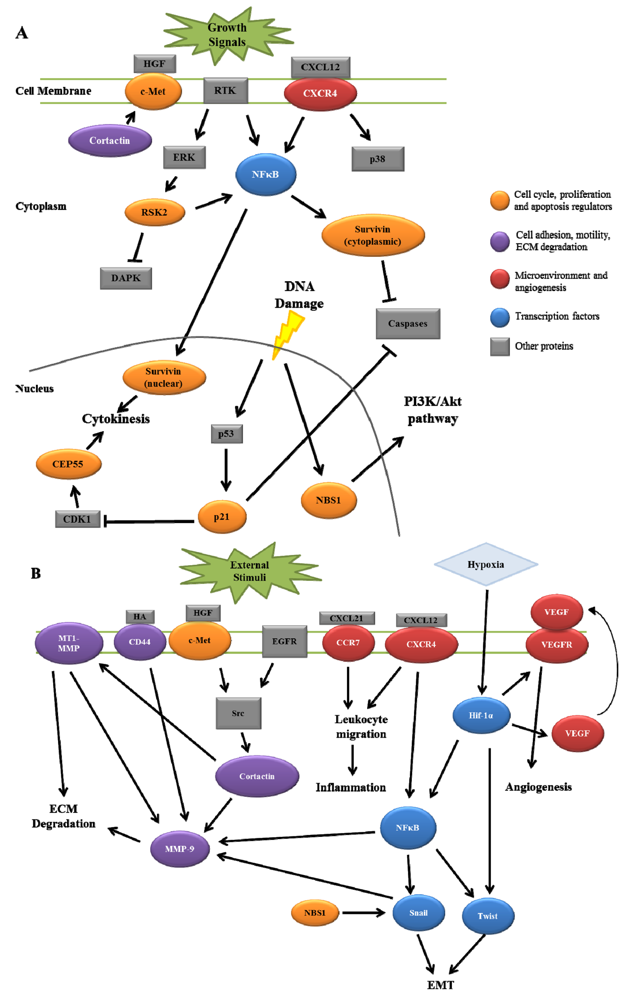
HNSCC invasion and nodal metastasis is a complex process involving multiple signaling pathways and protein components. As reviewed here, several proteins have recently been characterized that give insight into HNSCC progression that have been documented to interact in signaling pathways ultimately resulting in lymph node metastasis. Figure 1 summarizes the potential interactions between these signaling pathways. Combinations of multiple protein expression patterns may potentially produce an accurate lymph node metastasis signature that could serve as a predictive tool for analyzing patient tumors. While the implementation of such a signature would require further validation in both experimental and clinical settings, the outcome of such work would provide an improved understanding of HNSCC as a disease, with the combined goal of enhancing overall patient quality of life.


{kind=link}
{kind=link}
{kind=link}
| Biomarker | Cellular Function | Relevant References |
|---|---|---|
| c-Met | Proliferation | [23] |
| CEP55 | Cell cycle regulation, cytokinesis | [39,40] |
| NBS1 | Cell cycle regulation, DNA double-strand break repair | [43,45] |
| Survivin | Inhibitor of apoptosis | [51,55,61] |
| RSK2 | Cell cycle regulation, proliferation, apoptosis | [65] |
| Cortactin | Cell motility and invasion | [74-76,79] |
| CD44 | Cell-cell and cell-matrix adhesions | [81-83] |
| MMP-9 | ECM degradation | [81,88] |
| MT1-MMP | ECM degradation | [90] |
| CXCR4 | SDF-1 chemokine receptor, chemoattraction | [98-101] |
| CCR7 | Chemokine receptor, chemoattraction | [100,101,103] |
| VEGF/R | Angiogenesis | [110,115,117,119] |
| NFκB | Proinflammatory TF | [126,127] |
| Twist | TF, regulator of EMT | [101,138] |
| Snail | TF, regulator of EMT | [138,141,142] |
| Hif-1α | TF, hypoxia | [138,146] |
| p21WAF1/Cip1 | Cell cycle regulation, proliferation, apoptosis | [154,155] |
MMP: matrix metalloproteinase; ECM: extracellular matrix; TF: transcription factor;EMT: epithelial-mesenchymal transition.
Acknowledgements
This work was supported by a subproject of National Institute of Health grant RR16440 to SAW and by the West Virginia University Office of Research and Graduate Education (ELW).
References
- Leemans, C.R.; Tiwari, R.; Nauta, J.J.; van der Waal, I.; Snow, G.B. Regional Lymph Node Involvement and its Significance in the Development of Distant Metastases in Head and Neck Carcinoma. Cancer 1993, 71, 452–456. [Google Scholar]
- Jemal, A.; Siegel, R.; Xu, J.; Ward, E. Cancer Statistics, 2010. CA Cancer J. Clin. 2010, 60, 277–300. [Google Scholar]
- Altekruse, S.F.; Kosary, C.L.; Krapcho, M.; Neyman, N.; Aminou, R.; Waldron, W.; Ruhl, J.; Howlader, N.; Tatalovich, Z.; Cho, H.; et al. SEER Cancer Statistics Review, 1975-2007; National Cancer Institute: Bethesda, MD, USA, 2010. Available online: http://seer.Cancer.gov/csr/1975_2007/ (Based on November 2009 SEER Data Submission, Posted to the SEER Web Site, 2010).
- Pai, S.I.; Westra, W.H. Molecular Pathology of Head and Neck Cancer: Implications for Diagnosis, Prognosis, and Treatment. Annu. Rev. Pathol. 2009, 4, 49–70. [Google Scholar]
- Le Tourneau, C.; Velten, M.; Jung, G.M.; Bronner, G.; Flesch, H.; Borel, C. Prognostic Indicators for Survival in Head and Neck Squamous Cell Carcinomas: Analysis of a Series of 621 Cases. Head Neck 2005, 27, 801–808. [Google Scholar]
- Dunne, A.A.; Muller, H.H.; Eisele, D.W.; Kessel, K.; Moll, R.; Werner, J.A. Meta-Analysis of the Prognostic Significance of Perinodal Spread in Head and Neck Squamous Cell Carcinomas (HNSCC) Patients. Eur. J. Cancer 2006, 42, 1863–1868. [Google Scholar]
- Liu, M.; Weynand, B.; Delos, M.; Marbaix, E. Prognostic Factors in Squamous Cell Carcinomas of the Head and Neck. Acta Otorhinolaryngol. Belg. 1999, 53, 155–160. [Google Scholar]
- Hanahan, D.; Weinberg, R. A. The Hallmarks of Cancer. Cell 2000, 100, 57–70. [Google Scholar]
- Cortesina, G.; Martone, T. Molecular Metastases Markers in Head and Neck Squamous Cell Carcinoma: Review of the Literature. Acta Otorhinolaryngol. Ital. 2006, 26, 317–325. [Google Scholar]
- El-Naggar, A.K. Pathobiology of Head and Neck Squamous Tumorigenesis. Curr. Cancer Drug Targets 2007, 7, 606–612. [Google Scholar]
- Takes, R.P.; Rinaldo, A.; Rodrigo, J.P.; Devaney, K.O.; Fagan, J.J.; Ferlito, A. Can Biomarkers Play a Role in the Decision about Treatment of the Clinically Negative Neck in Patients with Head and Neck Cancer? Head Neck 2008, 30, 525–538. [Google Scholar]
- Molinolo, A.A.; Amornphimoltham, P.; Squarize, C.H.; Castilho, R.M.; Patel, V.; Gutkind, J.S. Dysregulated Molecular Networks in Head and Neck Carcinogenesis. Oral Oncol. 2009, 45, 324–334. [Google Scholar]
- Thomas, G.R.; Nadiminti, H.; Regalado, J. Molecular Predictors of Clinical Outcome in Patients with Head and Neck Squamous Cell Carcinoma. Int. J. Exp. Pathol. 2005, 86, 347–363. [Google Scholar]
- Hensen, E.F.; De Herdt, M.J.; Goeman, J.J.; Oosting, J.; Smit, V.T.; Cornelisse, C.J.; Baatenburg de Jong, R.J. Gene-Expression of Metastasized Versus Non-Metastasized Primary Head and Neck Squamous Cell Carcinomas: A Pathway-Based Analysis. BMC Cancer 2008, 8, 168. [Google Scholar]
- Rickman, D.S.; Millon, R.; De Reynies, A.; Thomas, E.; Wasylyk, C.; Muller, D.; Abecassis, J.; Wasylyk, B. Prediction of Future Metastasis and Molecular Characterization of Head and Neck Squamous-Cell Carcinoma Based on Transcriptome and Genome Analysis by Microarrays. Oncogene 2008, 27, 6607–6622. [Google Scholar]
- Roepman, P.; de Jager, A.; Groot Koerkamp, M.J.; Kummer, J.A.; Slootweg, P.J.; Holstege, F.C. Maintenance of Head and Neck Tumor Gene Expression Profiles upon Lymph Node Metastasis. Cancer Res. 2006, 66, 11110–11114. [Google Scholar]
- Gentile, A.; Trusolino, L.; Comoglio, P.M. The Met Tyrosine Kinase Receptor in Development and Cancer. Cancer Metastasis Rev. 2008, 27, 85–94. [Google Scholar]
- Comoglio, P.M.; Trusolino, L. Invasive Growth: From Development to Metastasis. J. Clin. Invest. 2002, 109, 857–862. [Google Scholar]
- Benvenuti, S.; Comoglio, P.M. The MET Receptor Tyrosine Kinase in Invasion and Metastasis. J. Cell. Physiol. 2007, 213, 316–325. [Google Scholar]
- Mueller, K.L.; Hunter, L.A.; Ethier, S.P.; Boerner, J.L. Met and c-Src Cooperate to Compensate for Loss of Epidermal Growth Factor Receptor Kinase Activity in Breast Cancer Cells. Cancer Res. 2008, 68, 3314–3322. [Google Scholar]
- Sen, B.; Peng, S.H.; Saigal, B.; Williams, M.D.; Johnson, F.M. Distinct Interactions between c-Src and c-Met in Mediating Resistance to c-Src Inhibition in Head and Neck Cancer. Clin. Cancer Res. 2010. [Google Scholar] [CrossRef]
- Egloff, A.M.; Grandis, J.R. Targeting Epidermal Growth Factor Receptor and SRC Pathways in Head and Neck Cancer. Semin. Oncol. 2008, 35, 286–297. [Google Scholar]
- Kim, C.H.; Koh, Y.W.; Han, J.H.; Kim, J.W.; Lee, J.S.; Baek, S.J.; Hwang, H.S.; Choi, E.C. C-Met Expression as an Indicator of Survival Outcome in Patients with Oral Tongue Carcinoma. Head Neck 2010, 32, 1655–1664. [Google Scholar]
- Kim, C.H.; Lee, J.S.; Kang, S.O.; Bae, J.H.; Hong, S.P.; Kahng, H. Serum Hepatocyte Growth Factor as a Marker of Tumor Activity in Head and Neck Squamous Cell Carcinoma. Oral Oncol. 2007, 43, 1021–1025. [Google Scholar]
- Druzgal, C.H.; Chen, Z.; Yeh, N.T.; Thomas, G.R.; Ondrey, F.G.; Duffey, D.C.; Vilela, R.J.; Ende, K.; McCullagh, L.; Rudy, S.F.; et al. A Pilot Study of Longitudinal Serum Cytokine and Angiogenesis Factor Levels as Markers of Therapeutic Response and Survival in Patients with Head and Neck Squamous Cell Carcinoma. Head Neck 2005, 27, 771–784. [Google Scholar]
- Allen, C.; Duffy, S.; Teknos, T.; Islam, M.; Chen, Z.; Albert, P.S.; Wolf, G.; Van Waes, C. Nuclear Factor-kappaB-Related Serum Factors as Longitudinal Biomarkers of Response and Survival in Advanced Oropharyngeal Carcinoma. Clin. Cancer Res. 2007, 13, 3182–3190. [Google Scholar]
- Kim, C.H.; Moon, S.K.; Bae, J.H.; Lee, J.H.; Han, J.H.; Kim, K.; Choi, E.C. Expression of Hepatocyte Growth Factor and c-Met in Hypopharyngeal Squamous Cell Carcinoma. Acta Otolaryngol. 2006, 126, 88–94. [Google Scholar]
- Uchida, D.; Kawamata, H.; Omotehara, F.; Nakashiro, K.; Kimura-Yanagawa, T.; Hino, S.; Begum, N.M.; Hoque, M.O.; Yoshida, H.; Sato, M.; et al. Role of HGF/c-Met System in Invasion and Metastasis of Oral Squamous Cell Carcinoma Cells in vitro and its Clinical Significance. Int. J. Cancer 2001, 93, 489–496. [Google Scholar]
- Marshall, D.D.; Kornberg, L.J. Overexpression of Scatter Factor and its Receptor (c-Met) in Oral Squamous Cell Carcinoma. Laryngoscope 1998, 108, 1413–1417. [Google Scholar]
- Galeazzi, E.; Olivero, M.; Gervasio, F.C.; De Stefani, A.; Valente, G.; Comoglio, P.M.; Di Renzo, M.F.; Cortesina, G. Detection of MET oncogene/hepatocyte Growth Factor Receptor in Lymph Node Metastases from Head and Neck Squamous Cell Carcinomas. Eur. Arch. Otorhinolaryngol. 1997, 254 (Suppl. 1), S138–S143. [Google Scholar]
- Bishop, E.A.; Lengyel, E.R.; Yamada, S.D.; Montag, A.; Temkin, S.M. The Expression of Hepatocyte Growth Factor (HGF) and c-Met in Uterine Serous Carcinoma. Gynecol. Oncol. 2010. [Google Scholar] [CrossRef]
- Zucali, P.A.; Ruiz, M.G.; Giovannetti, E.; Destro, A.; Varella-Garcia, M.; Floor, K.; Ceresoli, G.L.; Rodriguez, J.A.; Garassino, I.; Comoglio, P.; et al. Role of cMET Expression in Non-Small-Cell Lung Cancer Patients Treated with EGFR Tyrosine Kinase Inhibitors. Ann. Oncol. 2008, 19, 1605–1612. [Google Scholar]
- Mueller, K.L.; Yang, Z.Q.; Haddad, R.; Ethier, S.P.; Boerner, J.L. EGFR/Met Association Regulates EGFR TKI Resistance in Breast Cancer. J. Mol. Signal 2010, 5, 8. [Google Scholar]
- Liska, D.; Chen, C.T.; Bachleitner-Hofmann, T.; Christensen, J.G.; Weiser, M.R. HGF Rescues Colorectal Cancer Cells from EGFR Inhibition Via MET Activation. Clin. Cancer Res. 2010, 17, 472–482. [Google Scholar]
- Kasahara, K.; Arao, T.; Sakai, K.; Matsumoto, K.; Sakai, A.; Kimura, H.; Sone, T.; Horiike, A.; Nishio, M.; Ohira, T.; et al. Impact of Serum Hepatocyte Growth Factor on Treatment Response to Epidermal Growth Factor Receptor Tyrosine Kinase Inhibitors in Patients with Non-Small Cell Lung Adenocarcinoma. Clin. Cancer Res. 2010, 16, 4616–4624. [Google Scholar]
- Eder, J.P.; Vande Woude, G.F.; Boerner, S.A.; LoRusso, P.M. Novel Therapeutic Inhibitors of the c-Met Signaling Pathway in Cancer. Clin. Cancer Res. 2009, 15, 2207–2214. [Google Scholar]
- Fabbro, M.; Zhou, B.B.; Takahashi, M.; Sarcevic, B.; Lal, P.; Graham, M.E.; Gabrielli, B.G.; Robinson, P.J.; Nigg, E.A.; Ono, Y.; et al. Cdk1/Erk2- and Plk1-Dependent Phosphorylation of a Centrosome Protein, Cep55, is Required for its Recruitment to Midbody and Cytokinesis. Dev. Cell 2005, 9, 477–488. [Google Scholar]
- Gemenetzidis, E.; Bose, A.; Riaz, A.M.; Chaplin, T.; Young, B.D.; Ali, M.; Sugden, D.; Thurlow, J.K.; Cheong, S.C.; Teo, S.H.; et al. FOXM1 Upregulation is an Early Event in Human Squamous Cell Carcinoma and it is Enhanced by Nicotine during Malignant Transformation. PLoS One 2009, 4, e4849. [Google Scholar]
- Chen, C.H.; Chien, C.Y.; Huang, C.C.; Hwang, C.F.; Chuang, H.C.; Fang, F.M.; Huang, H.Y.; Chen, C.M.; Liu, H.L.; Huang, C.Y. Expression of FLJ10540 is Correlated with Aggressiveness of Oral Cavity Squamous Cell Carcinoma by Stimulating Cell Migration and Invasion through Increased FOXM1 and MMP-2 Activity. Oncogene 2009, 28, 2723–2737. [Google Scholar]
- Waseem, A.; Ali, M.; Odell, E.W.; Fortune, F.; Teh, M.T. Downstream Targets of FOXM1: CEP55 and HELLS are Cancer Progression Markers of Head and Neck Squamous Cell Carcinoma. Oral Oncol. 2010, 46, 536–542. [Google Scholar]
- Antoccia, A.; Kobayashi, J.; Tauchi, H.; Matsuura, S.; Komatsu, K. Nijmegen Breakage Syndrome and Functions of the Responsible Protein, NBS1. Genome Dyn. 2006, 1, 191–205. [Google Scholar]
- Lamarche, B.J.; Orazio, N.I.; Weitzman, M.D. The MRN Complex in Double-Strand Break Repair and Telomere Maintenance. FEBS Lett. 2010, 584, 3682–3695. [Google Scholar]
- Hsu, D.S.; Chang, S.Y.; Liu, C.J.; Tzeng, C.H.; Wu, K.J.; Kao, J.Y.; Yang, M.H. Identification of Increased NBS1 Expression as a Prognostic Marker of Squamous Cell Carcinoma of the Oral Cavity. Cancer Sci. 2010, 101, 1029–1037. [Google Scholar]
- Yang, M.H.; Chiang, W.C.; Chou, T.Y.; Chang, S.Y.; Chen, P.M.; Teng, S.C.; Wu, K.J. Increased NBS1 Expression is a Marker of Aggressive Head and Neck Cancer and Overexpression of NBS1 Contributes to Transformation. Clin. Cancer Res. 2006, 12, 507–515. [Google Scholar]
- Yang, M.H.; Chang, S.Y.; Chiou, S.H.; Liu, C.J.; Chi, C.W.; Chen, P.M.; Teng, S.C.; Wu, K.J. Overexpression of NBS1 Induces Epithelial-Mesenchymal Transition and Co-Expression of NBS1 and Snail Predicts Metastasis of Head and Neck Cancer. Oncogene 2007, 26, 1459–1467. [Google Scholar]
- Park, S.L.; Bastani, D.; Goldstein, B.Y.; Chang, S.C.; Cozen, W.; Cai, L.; Cordon-Cardo, C.; Ding, B.; Greenland, S.; He, N.; et al. Associations between NBS1 Polymorphisms, Haplotypes and Smoking-Related Cancers. Carcinogenesis 2010, 31, 1264–1271. [Google Scholar]
- Schuetz, J.M.; MaCarthur, A.C.; Leach, S.; Lai, A.S.; Gallagher, R.P.; Connors, J.M.; Gascoyne, R.D.; Spinelli, J.J.; Brooks-Wilson, A.R. Genetic Variation in the NBS1, MRE11, RAD50 and BLM Genes and Susceptibility to Non-Hodgkin Lymphoma. BMC Med. Genet. 2009, 10, 117. [Google Scholar]
- di Masi, A.; Antoccia, A. NBS1 Heterozygosity and Cancer Risk. Curr. Genomics 2008, 9, 275–281. [Google Scholar]
- Duffy, M.J.; O'Donovan, N.; Brennan, D.J.; Gallagher, W.M.; Ryan, B.M. Survivin: A Promising Tumor Biomarker. Cancer Lett. 2007, 249, 49–60. [Google Scholar]
- Guha, M.; Altieri, D.C. Survivin as a Global Target of Intrinsic Tumor Suppression Networks. Cell Cycle 2009, 8, 2708–2710. [Google Scholar]
- Kim, Y.H.; Kim, S.M.; Kim, Y.K.; Hong, S.P.; Kim, M.J.; Myoung, H. Evaluation of Survivin as a Prognostic Marker in Oral Squamous Cell Carcinoma. J. Oral Pathol. Med. 2010, 39, 368–375. [Google Scholar]
- Khan, Z.; Tiwari, R.P.; Mulherkar, R.; Sah, N.K.; Prasad, G.B.; Shrivastava, B.R.; Bisen, P.S. Detection of Survivin and p53 in Human Oral Cancer: Correlation with Clinicopathologic Findings. Head Neck 2009, 31, 1039–1048. [Google Scholar]
- Lippert, B.M.; Knauer, S.K.; Fetz, V.; Mann, W.; Stauber, R.H. Dynamic Survivin in Head and Neck Cancer: Molecular Mechanism and Therapeutic Potential. Int. J. Cancer 2007, 121, 1169–1174. [Google Scholar]
- Qi, G.; Kudo, Y.; Ando, T.; Tsunematsu, T.; Shimizu, N.; Siriwardena, S.B.; Yoshida, M.; Keikhaee, M.R.; Ogawa, I.; Takata, T. Nuclear Survivin Expression is Correlated with Malignant Behaviors of Head and Neck Cancer Together with Aurora-B. Oral Oncol. 2010, 46, 263–270. [Google Scholar]
- Lin, J.; Guan, Z.; Wang, C.; Feng, L.; Zheng, Y.; Caicedo, E.; Bearth, E.; Peng, J.R.; Gaffney, P.; Ondrey, F.G. Inhibitor of Differentiation 1 Contributes to Head and Neck Squamous Cell Carcinoma Survival Via the NF-kappaB/survivin and Phosphoinositide 3-kinase/Akt Signaling Pathways. Clin. Cancer Res. 2010, 16, 77–87. [Google Scholar]
- Preuss, S.F.; Weinell, A.; Molitor, M.; Semrau, R.; Stenner, M.; Drebber, U.; Wedemeyer, I.; Hoffmann, T.K.; Guntinas-Lichius, O.; Klussmann, J.P. Survivin and Epidermal Growth Factor Receptor Expression in Surgically Treated Oropharyngeal Squamous Cell Carcinoma. Head Neck 2008b, 30, 1318–1324. [Google Scholar]
- Preuss, S.F.; Weinell, A.; Molitor, M.; Stenner, M.; Semrau, R.; Drebber, U.; Weissenborn, S.J.; Speel, E.J.; Wittekindt, C.; Guntinas-Lichius, O.; et al. Nuclear Survivin Expression is Associated with HPV-Independent Carcinogenesis and is an Indicator of Poor Prognosis in Oropharyngeal Cancer. Br. J. Cancer 2008a, 98, 627–632. [Google Scholar]
- Fetz, V.; Bier, C.; Habtemichael, N.; Schuon, R.; Schweitzer, A.; Kunkel, M.; Engels, K.; Kovacs, A.F.; Schneider, S.; Mann, W.; et al. Inducible NO Synthase Confers Chemoresistance in Head and Neck Cancer by Modulating Survivin. Int. J. Cancer 2009, 124, 2033–2041. [Google Scholar]
- Wang, S.; Huang, X.; Lee, C.K.; Liu, B. Elevated Expression of erbB3 Confers Paclitaxel Resistance in erbB2-Overexpressing Breast Cancer Cells Via Upregulation of Survivin. Oncogene 2010, 29, 4225–4236. [Google Scholar]
- Promkan, M.; Liu, G.; Patmasiriwat, P.; Chakrabarty, S. BRCA1 Modulates Malignant Cell Behavior, the Expression of Survivin and Chemosensitivity in Human Breast Cancer Cells. Int. J. Cancer 2009, 125, 2820–2828. [Google Scholar]
- Wang, Q.; Wu, P.C.; Roberson, R.S.; Luk, B.V.; Ivanova, I.; Chu, E.; Wu, D.Y. Survivin and Escaping in Therapy-Induced Cellular Senescence. Int. J. Cancer 2010. [Google Scholar] [CrossRef]
- Su, L.; Wang, Y.; Xiao, M.; Lin, Y.; Yu, L. Up-Regulation of Survivin in Oral Squamous Cell Carcinoma Correlates with Poor Prognosis and Chemoresistance. Oral Surg. Oral Med. Oral Pathol. Oral Radiol. Endod. 2010, 110, 484–491. [Google Scholar]
- Carriere, A.; Ray, H.; Blenis, J.; Roux, P.P. The RSK Factors of Activating the Ras/MAPK Signaling Cascade. Front. Biosci. 2008, 13, 4258–4275. [Google Scholar]
- Anjum, R.; Blenis, J. The RSK Family of Kinases: Emerging Roles in Cellular Signalling. Nat. Rev. Mol. Cell Biol. 2008, 9, 747–758. [Google Scholar]
- Kang, S.; Elf, S.; Lythgoe, K.; Hitosugi, T.; Taunton, J.; Zhou, W.; Xiong, L.; Wang, D.; Muller, S.; Fan, S.; et al. P90 Ribosomal S6 Kinase 2 Promotes Invasion and Metastasis of Human Head and Neck Squamous Cell Carcinoma Cells. J. Clin. Invest. 2010, 120, 1165–1177. [Google Scholar]
- Cho, Y.Y.; Yao, K.; Kim, H.G.; Kang, B.S.; Zheng, D.; Bode, A.M.; Dong, Z. Ribosomal S6 Kinase 2 is a Key Regulator in Tumor Promoter Induced Cell Transformation. Cancer Res. 2007, 67, 8104–8112. [Google Scholar]
- Nguyen, T.L. Targeting RSK: An Overview of Small Molecule Inhibitors. Anticancer Agents Med. Chem. 2008, 8, 710–716. [Google Scholar]
- Ammer, A.G.; Weed, S.A. Cortactin Branches Out: Roles in Regulating Protrusive Actin Dynamics. Cell Motil. Cytoskeleton 2008, 65, 687–707. [Google Scholar]
- Rodrigo, J.P.; Garcia, L.A.; Ramos, S.; Lazo, P.S.; Suarez, C. EMS1 Gene Amplification Correlates with Poor Prognosis in Squamous Cell Carcinomas of the Head and Neck. Clin. Cancer Res. 2000, 6, 3177–3182. [Google Scholar]
- Rothschild, B.L.; Shim, A.H.; Ammer, A.G.; Kelley, L.C.; Irby, K.B.; Head, J.A.; Chen, L.; Varella-Garcia, M.; Sacks, P.G.; Frederick, B.; et al. Cortactin Overexpression Regulates Actin-Related Protein 2/3 Complex Activity, Motility, and Invasion in Carcinomas with Chromosome 11q13 Amplification. Cancer Res. 2006, 66, 8017–8025. [Google Scholar]
- Meredith, S.D.; Levine, P.A.; Burns, J.A.; Gaffey, M.J.; Boyd, J.C.; Weiss, L.M.; Erickson, N.L.; Williams, M.E. Chromosome 11q13 Amplification in Head and Neck Squamous Cell Carcinoma. Association with Poor Prognosis. Arch. Otolaryngol. Head. Neck. Surg. 1995, 121, 790–794. [Google Scholar]
- Williams, M.E.; Gaffey, M.J.; Weiss, L.M.; Wilczynski, S.P.; Schuuring, E.; Levine, P.A. Chromosome 11Q13 Amplification in Head and Neck Squamous Cell Carcinoma. Arch. Otolaryngol. Head. Neck. Surg. 1993, 119, 1238–1243. [Google Scholar]
- Luo, M.L.; Shen, X.M.; Zhang, Y.; Wei, F.; Xu, X.; Cai, Y.; Zhang, X.; Sun, Y.T.; Zhan, Q.M.; Wu, M.; et al. Amplification and Overexpression of CTTN (EMS1) Contribute to the Metastasis of Esophageal Squamous Cell Carcinoma by Promoting Cell Migration and Anoikis Resistance. Cancer Res. 2006, 66, 11690–11699. [Google Scholar]
- Yamada, S.I.; Yanamoto, S.; Kawasaki, G.; Mizuno, A.; Nemoto, T.K. Overexpression of Cortactin Increases Invasion Potential in Oral Squamous Cell Carcinoma. Pathol. Oncol. Res. 2010, 4, 523–531. [Google Scholar]
- Rodrigo, J.P.; Garcia-Carracedo, D.; Garcia, L.A.; Menendez, S.; Allonca, E.; Gonzalez, M.V.; Fresno, M.F.; Suarez, C.; Garcia-Pedrero, J.M. Distinctive Clinicopathological Associations of Amplification of the Cortactin Gene at 11q13 in Head and Neck Squamous Cell Carcinomas. J. Pathol. 2009, 217, 516–523. [Google Scholar]
- Hofman, P.; Butori, C.; Havet, K.; Hofman, V.; Selva, E.; Guevara, N.; Santini, J.; Van Obberghen-Schilling, E. Prognostic Significance of Cortactin Levels in Head and Neck Squamous Cell Carcinoma: Comparison with Epidermal Growth Factor Receptor Status. Br. J. Cancer 2008, 98, 956–964. [Google Scholar]
- Timpson, P.; Wilson, A.S.; Lehrbach, G.M.; Sutherland, R.L.; Musgrove, E.A.; Daly, R.J. Aberrant Expression of Cortactin in Head and Neck Squamous Cell Carcinoma Cells is Associated with Enhanced Cell Proliferation and Resistance to the Epidermal Growth Factor Receptor Inhibitor Gefitinib. Cancer Res. 2007, 67, 9304–9314. [Google Scholar]
- Fantozzi, I.; Grall, D.; Cagnol, S.; Stanchi, F.; Sudaka, A.; Brunstein, M.C.; Bozec, A.; Fischel, J.L.; Milano, G.; Van Obberghen-Schilling, E. Overexpression of Cortactin in Head and Neck Squamous Cell Carcinomas can be Uncoupled from Augmented EGF Receptor Expression. Acta Oncol. 2008, 47, 1502–1512. [Google Scholar]
- Gibcus, J.H.; Mastik, M.F.; Menkema, L.; de Bock, G.H.; Kluin, P.M.; Schuuring, E.; van der Wal, J.E. Cortactin Expression Predicts Poor Survival in Laryngeal Carcinoma. Br. J. Cancer 2008, 98, 950–955. [Google Scholar]
- Orian-Rousseau, V. CD44, a Therapeutic Target for Metastasising Tumours. Eur. J. Cancer 2010, 46, 1271–1277. [Google Scholar]
- Wang, S.J.; Wong, G.; de Heer, A.M.; Xia, W.; Bourguignon, L.Y. CD44 Variant Isoforms in Head and Neck Squamous Cell Carcinoma Progression. Laryngoscope 2009, 119, 1518–1530. [Google Scholar]
- Yuce, I.; Bayram, A.; Cagli, S.; Canoz, O.; Bayram, S.; Guney, E. The Role of CD44 and Matrix Metalloproteinase-9 Expression in Predicting Neck Metastasis of Supraglottic Laryngeal Carcinoma. Am. J. Otolaryngol. 2010. [Google Scholar] [CrossRef]
- Lin, J.T.; Chang, T.H.; Chang, C.S.; Wang, W.H.; Su, B.W.; Lee, K.D.; Chang, P.J. Prognostic Value of Pretreatment CD44 mRNA in Peripheral Blood of Patients with Locally Advanced Head and Neck Cancer. Oral Oncol. 2010, 46, e29–e33. [Google Scholar]
- Kessenbrock, K.; Plaks, V.; Werb, Z. Matrix Metalloproteinases: Regulators of the Tumor Microenvironment. Cell 2010, 141, 52–67. [Google Scholar]
- Yu, Q.; Stamenkovic, I. Cell Surface-Localized Matrix Metalloproteinase-9 Proteolytically Activates TGF-Beta and Promotes Tumor Invasion and Angiogenesis. Genes Dev. 2000, 14, 163–176. [Google Scholar]
- Lee, J.I.; Jin, B.H.; Kim, M.A.; Yoon, H.J.; Hong, S.P.; Hong, S.D. Prognostic Significance of CXCR-4 Expression in Oral Squamous Cell Carcinoma. Oral Surg. Oral Med. Oral Pathol. Oral Radiol. Endod. 2009, 107, 678–684. [Google Scholar]
- Sterz, C.M.; Kulle, C.; Dakic, B.; Makarova, G.; Bottcher, M.C.; Bette, M.; Werner, J.A.; Mandic, R. A Basal-Cell-Like Compartment in Head and Neck Squamous Cell Carcinomas Represents the Invasive Front of the Tumor and is Expressing MMP-9. Oral Oncol. 2010, 46, 116–122. [Google Scholar]
- Liu, C.J.; Chang, K.W.; Lin, S.C.; Cheng, H.W. Presurgical Serum Levels of Matrix Metalloproteinase-9 and Vascular Endothelial Growth Factor in Oral Squamous Cell Carcinoma. Oral Oncol. 2009, 45, 920–925. [Google Scholar]
- Ruokolainen, H.; Paakko, P.; Turpeenniemi-Hujanen, T. Serum Matrix Metalloproteinase-9 in Head and Neck Squamous Cell Carcinoma is a Prognostic Marker. Int. J. Cancer 2005, 116, 422–427. [Google Scholar]
- Zhang, H.; Liu, M.; Sun, Y.; Lu, J. MMP-14 can Serve as a Prognostic Marker in Patients with Supraglottic Cancer. Eur. Arch. Otorhinolaryngol. 2009, 266, 1427–1434. [Google Scholar]
- Clark, E.S.; Whigham, A.S.; Yarbrough, W.G.; Weaver, A.M. Cortactin is an Essential Regulator of Matrix Metalloproteinase Secretion and Extracellular Matrix Degradation in Invadopodia. Cancer Res. 2007, 67, 4227–4235. [Google Scholar]
- Clark, E.S.; Weaver, A.M. A New Role for Cortactin in Invadopodia: Regulation of Protease Secretion. Eur. J. Cell Biol. 2008, 87, 581–590. [Google Scholar]
- Coussens, L.M.; Fingleton, B.; Matrisian, L.M. Matrix Metalloproteinase Inhibitors and Cancer: Trials and Tribulations. Science 2002, 295, 2387–2392. [Google Scholar]
- Dorman, G.; Cseh, S.; Hajdu, I.; Barna, L.; Konya, D.; Kupai, K.; Kovacs, L.; Ferdinandy, P. Matrix Metalloproteinase Inhibitors: A Critical Appraisal of Design Principles and Proposed Therapeutic Utility. Drugs 2010, 70, 949–964. [Google Scholar]
- Kakinuma, T.; Hwang, S.T. Chemokines, Chemokine Receptors, and Cancer Metastasis. J. Leukoc. Biol. 2006, 79, 639–651. [Google Scholar]
- Ben-Baruch, A. Site-Specific Metastasis Formation: Chemokines as Regulators of Tumor Cell Adhesion, Motility and Invasion. Cell. Adh Migr. 2009, 3, 328–333. [Google Scholar]
- Teicher, B.A.; Fricker, S.P. CXCL12 (SDF-1)/CXCR4 Pathway in Cancer. Clin. Cancer Res. 2010, 16, 2927–2931. [Google Scholar]
- Tan, C.T.; Chu, C.Y.; Lu, Y.C.; Chang, C.C.; Lin, B.R.; Wu, H.H.; Liu, H.L.; Cha, S.T.; Prakash, E.; Ko, J.Y.; et al. CXCL12/CXCR4 Promotes Laryngeal and Hypopharyngeal Squamous Cell Carcinoma Metastasis through MMP-13-Dependent Invasion Via the ERK1/2/AP-1 Pathway. Carcinogenesis 2008, 29, 1519–1527. [Google Scholar]
- Meng, X.; Wuyi, L.; Yuhong, X.; Xinming, C. Expression of CXCR4 in Oral Squamous Cell Carcinoma: Correlations with Clinicopathology and Pivotal Role of Proliferation. J. Oral Pathol. Med. 2010, 39, 63–68. [Google Scholar]
- Ueda, M.; Shimada, T.; Goto, Y.; Tei, K.; Nakai, S.; Hisa, Y.; Kannagi, R. Expression of CC-Chemokine Receptor 7 (CCR7) and CXC-Chemokine Receptor 4 (CXCR4) in Head and Neck Squamous Cell Carcinoma. Auris Nasus Larynx 2010, 37, 488–495. [Google Scholar]
- Ou, D.L.; Chien, H.F.; Chen, C.L.; Lin, T.C.; Lin, L.I. Role of Twist in Head and Neck Carcinoma with Lymph Node Metastasis. Anticancer Res. 2008, 28, 1355–1359. [Google Scholar]
- Pitkin, L.; Luangdilok, S.; Corbishley, C.; Wilson, P.O.; Dalton, P.; Bray, D.; Mady, S.; Williamson, P.; Odutoye, T.; Rhys Evans, P.; et al. Expression of CC Chemokine Receptor 7 in Tonsillar Cancer Predicts Cervical Nodal Metastasis, Systemic Relapse and Survival. Br. J. Cancer 2007, 97, 670–677. [Google Scholar]
- Shang, Z.J.; Liu, K.; Shao, Z. Expression of Chemokine Receptor CCR7 is Associated with Cervical Lymph Node Metastasis of Oral Squamous Cell Carcinoma. Oral Oncol. 2009, 45, 480–485. [Google Scholar]
- Wang, J.; Seethala, R.R.; Zhang, Q.; Gooding, W.; van Waes, C.; Hasegawa, H.; Ferris, R.L. Autocrine and Paracrine Chemokine Receptor 7 Activation in Head and Neck Cancer: Implications for Therapy. J. Natl. Cancer Inst. 2008, 100, 502–512. [Google Scholar]
- Wolf, M.; Albrecht, S.; Marki, C. Proteolytic Processing of Chemokines: Implications in Physiological and Pathological Conditions. Int. J. Biochem. Cell Biol. 2008, 40, 1185–1198. [Google Scholar]
- Yu, T.; Wu, Y.; Helman, J.I.; Wen, Y.; Wang, C.; Li, L. CXCR4 Promotes Oral Squamous Cell Carcinoma Migration and Invasion through Inducing Expression of MMP-9, 13 Via the ERK Signaling Pathway. Mol. Cancer. Res. 2011. [Google Scholar] [CrossRef]
- Raman, D.; Baugher, P.J.; Thu, Y.M.; Richmond, A. Role of Chemokines in Tumor Growth. Cancer Lett. 2007, 256, 137–165. [Google Scholar]
- Wang, J.; Xi, L.; Hunt, J.L.; Gooding, W.; Whiteside, T.L.; Chen, Z.; Godfrey, T.E.; Ferris, R.L. Expression Pattern of Chemokine Receptor 6 (CCR6) and CCR7 in Squamous Cell Carcinoma of the Head and Neck Identifies a Novel Metastatic Phenotype. Cancer Res. 2004, 64, 1861–1866. [Google Scholar]
- Margaritescu, C.; Pirici, D.; Simionescu, C.; Mogoanta, L.; Raica, M.; Stinga, A.; Ciurea, R.; Stepan, A.; Stinga, A.; Ribatti, D. VEGF and VEGFRs Expression in Oral Squamous Cell Carcinoma. Rom. J. Morphol. Embryol. 2009, 50, 527–548. [Google Scholar]
- Joo, Y.H.; Jung, C.K.; Kim, M.S.; Sun, D.I. Relationship between Vascular Endothelial Growth Factor and Notch1 Expression and Lymphatic Metastasis in Tongue Cancer. Otolaryngol. Head. Neck. Surg. 2009, 140, 512–518. [Google Scholar]
- Hicklin, D.J.; Ellis, L.M. Role of the Vascular Endothelial Growth Factor Pathway in Tumor Growth and Angiogenesis. J. Clin. Oncol. 2005, 23, 1011–1027. [Google Scholar]
- Winder, T.; Lenz, H.J. Vascular Endothelial Growth Factor and Epidermal Growth Factor Signaling Pathways as Therapeutic Targets for Colorectal Cancer. Gastroenterology 2010, 138, 2163–2176. [Google Scholar]
- Guo, S.; Colbert, L.S.; Fuller, M.; Zhang, Y.; Gonzalez-Perez, R.R. Vascular Endothelial Growth Factor Receptor-2 in Breast Cancer. Biochim. Biophys. Acta. 2010, 1806, 108–121. [Google Scholar]
- Delli Carpini, J.; Karam, A.K.; Montgomery, L. Vascular Endothelial Growth Factor and its Relationship to the Prognosis and Treatment of Breast, Ovarian, and Cervical Cancer. Angiogenesis 2010, 13, 43–58. [Google Scholar]
- Hong, D.Y.; Lee, B.J.; Lee, J.C.; Choi, J.S.; Wang, S.G.; Ro, J.H. Expression of VEGF, HGF, IL-6, IL-8, MMP-9, Telomerase in Peripheral Blood of Patients with Head and Neck Squamous Cell Carcinoma. Clin. Exp. Otorhinolaryngol. 2009, 2, 186–192. [Google Scholar]
- Lv, X.; Xiang, Y.Q.; Cao, S.M.; Qian, C.N.; Li, N.W.; Guo, L.; Mai, H.Q.; Chen, Q.Y.; Huang, P.Y.; Luo, D.; et al. Prospective Validation of the Prognostic Value of Elevated Serum Vascular Endothelial Growth Factor in Patients with Nasopharyngeal Carcinoma: More Distant Metastases and Shorter overall Survival After Treatment. Head Neck 2010. [Google Scholar] [CrossRef]
- Sugiura, T.; Inoue, Y.; Matsuki, R.; Ishii, K.; Takahashi, M.; Abe, M.; Shirasuna, K. VEGF-C and VEGF-D Expression is Correlated with Lymphatic Vessel Density and Lymph Node Metastasis in Oral Squamous Cell Carcinoma: Implications for use as a Prognostic Marker. Int. J. Oncol. 2009, 34, 673–680. [Google Scholar]
- Borggrefe, T.; Oswald, F. The Notch Signaling Pathway: Transcriptional Regulation at Notch Target Genes. Cell Mol. Life Sci. 2009, 66, 1631–1646. [Google Scholar]
- Shao, Z.; Zhang, W.F.; Chen, X.M.; Shang, Z.J. Expression of EphA2 and VEGF in Squamous Cell Carcinoma of the Tongue: Correlation with the Angiogenesis and Clinical Outcome. Oral Oncol. 2008, 44, 1110–1117. [Google Scholar]
- Pentheroudakis, G.; Angouridakis, N.; Wirtz, R.; Nikolaou, A.; Kalogeras, K.T.; Pavlidis, N.; Fountzilas, G. Transcriptional Activity of Human Epidermal Growth Factor Receptor Family and Angiogenesis Effectors in Locoregionally Recurrent Head and Neck Squamous Cell Carcinoma and Correlation with Patient Outcome. J. Oncol. 2009, 2009, 854127. [Google Scholar]
- Rueda, A.; Cazorla, O.; Perez, L.; Alvarez, M.; Redondo, M.; Gallego, E.; Saez, M.; Medina, J.A.; Solano, J.; Matilla, A. Vascular Endothelial Growth Factor and Vascular Endothelial Growth Factor Receptor-2 Tumor Expression in Patients with Advanced Laryngeal Cancer After Induction Chemotherapy for Organ Preservation. Head Neck 2010. [Google Scholar] [CrossRef]
- Faustino, S.E.; Oliveira, D.T.; Nonogaki, S.; Landman, G.; Carvalho, A.L.; Kowalski, L.P. Expression of Vascular Endothelial Growth Factor-C does Not Predict Occult Lymph-Node Metastasis in Early Oral Squamous Cell Carcinoma. Int. J. Oral Maxillofac. Surg. 2008, 37, 372–378. [Google Scholar]
- Luqman, S.; Pezzuto, J.M. NFkappaB: A Promising Target for Natural Products in Cancer Chemoprevention. Phytother. Res. 2010, 24, 949–963. [Google Scholar]
- Aggarwal, B.B. Nuclear Factor-kappaB: The Enemy within Cancer. Cell 2004, 6, 203–208. [Google Scholar]
- Mantovani, A.; Allavena, P.; Sica, A.; Balkwill, F. Cancer-Related Inflammation. Nature 2008, 454, 436–444. [Google Scholar]
- Yan, M.; Xu, Q.; Zhang, P.; Zhou, X.J.; Zhang, Z.Y.; Chen, W.T. Correlation of NF-kappaB Signal Pathway with Tumor Metastasis of Human Head and Neck Squamous Cell Carcinoma. BMC Cancer 2010, 10, 437. [Google Scholar]
- Huang, C.; Huang, K.; Wang, C.; Jiang, Z.D.; Li, X.X.; Wang, H.P.; Chen, H.Y. Overexpression of Mitogen-Activated Protein Kinase Kinase 4 and Nuclear Factor-kappaB in Laryngeal Squamous Cell Carcinoma: A Potential Indicator for Poor Prognosis. Oncol. Rep. 2009, 22, 89–95. [Google Scholar]
- Yoshida, K.; Sasaki, R.; Nishimura, H.; Okamoto, Y.; Suzuki, Y.; Kawabe, T.; Saito, M.; Otsuki, N.; Hayashi, Y.; Soejima, T.; et al. Nuclear Factor-kappaB Expression as a Novel Marker of Radioresistance in Early-Stage Laryngeal Cancer. Head Neck 2010, 32, 646–655. [Google Scholar]
- Pham, C.G.; Bubici, C.; Zazzeroni, F.; Knabb, J.R.; Papa, S.; Kuntzen, C.; Franzoso, G. Upregulation of Twist-1 by NF-kappaB Blocks Cytotoxicity Induced by Chemotherapeutic Drugs. Mol. Cell. Biol. 2007, 27, 3920–3935. [Google Scholar]
- julien, S.; Puig, I.; Caretti, E.; Bonaventure, J.; Nelles, L.; van Roy, F.; Dargemont, C.; de Herreros, A.G.; Bellacosa, A.; Larue, L. Activation of NF-kappaB by Akt Upregulates Snail Expression and Induces Epithelium Mesenchyme Transition. Oncogene 2007, 26, 7445–7456. [Google Scholar]
- Rehman, A.O.; Wang, C.Y. SDF-1alpha Promotes Invasion of Head and Neck Squamous Cell Carcinoma by Activating NF-kappaB. J. Biol. Chem. 2008, 283, 19888–19894. [Google Scholar]
- Chaturvedi, M.M.; Sung, B.; Yadav, V.R.; Kannappan, R.; Aggarwal, B.B. NF-kappaB Addiction and its Role in Cancer: 'One Size does Not Fit all'. Oncogene 2010. [Google Scholar] [CrossRef]
- Liu, F.Y.; Zhao, Z.J.; Li, P.; Ding, X.; Guo, N.; Yang, L.L.; Zong, Z.H.; Sun, C.F. NF-kappaB Participates in Chemokine Receptor 7-Mediated Cell Survival in Metastatic Squamous Cell Carcinoma of the Head and Neck. Oncol. Rep. 2011, 25, 383–391. [Google Scholar]
- Iwatsuki, M.; Mimori, K.; Yokobori, T.; Ishi, H.; Beppu, T.; Nakamori, S.; Baba, H.; Mori, M. Epithelial-Mesenchymal Transition in Cancer Development and its Clinical Significance. Cancer Sci. 2010, 101, 293–299. [Google Scholar]
- Yilmaz, M.; Christofori, G. EMT, the Cytoskeleton, and Cancer Cell Invasion. Cancer Metastasis Rev. 2009, 28, 15–33. [Google Scholar]
- Ouyang, G.; Wang, Z.; Fang, X.; Liu, J.; Yang, C.J. Molecular Signaling of the Epithelial to Mesenchymal Transition in Generating and Maintaining Cancer Stem Cells. Cell Mol. Life Sci. 2010, 67, 2605–2618. [Google Scholar]
- Yang, J.; Mani, S.A.; Donaher, J.L.; Ramaswamy, S.; Itzykson, R.A.; Come, C.; Savagner, P.; Gitelman, I.; Richardson, A.; Weinberg, R.A. Twist, a Master Regulator of Morphogenesis, Plays an Essential Role in Tumor Metastasis. Cell 2004, 117, 927–939. [Google Scholar]
- Yang, M.H.; Wu, M.Z.; Chiou, S.H.; Chen, P.M.; Chang, S.Y.; Liu, C.J.; Teng, S.C.; Wu, K.J. Direct Regulation of TWIST by HIF-1alpha Promotes Metastasis. Nat. Cell Biol. 2008, 10, 295–305. [Google Scholar]
- Yang, M.H.; Hsu, D.S.; Wang, H.W.; Wang, H.J.; Lan, H.Y.; Yang, W.H.; Huang, C.H.; Kao, S.Y.; Tzeng, C.H.; Tai, S.K.; et al. Bmi1 is Essential in Twist1-Induced Epithelial-Mesenchymal Transition. Nat. Cell Biol. 2010, 12, 982–992. [Google Scholar]
- Shiota, M.; Izumi, H.; Onitsuka, T.; Miyamoto, N.; Kashiwagi, E.; Kidani, A.; Yokomizo, A.; Naito, S.; Kohno, K. Twist Promotes Tumor Cell Growth through YB-1 Expression. Cancer Res. 2008, 68, 98–105. [Google Scholar]
- Yu, C.C.; Lo, W.L.; Chen, Y.W.; Huang, P.I.; Hsu, H.S.; Tseng, L.M.; Hung, S.C.; Kao, S.Y.; Chang, C.J.; Chiou, S.H. Bmi-1 Regulates Snail Expression and Promotes Metastasis Ability in Head and Neck Squamous Cancer-Derived ALDH1 Positive Cells. J. Oncol. 2011, 2011. [Google Scholar] [CrossRef]
- Hayry, V.; Makinen, L.K.; Atula, T.; Sariola, H.; Makitie, A.; Leivo, I.; Keski-Santti, H.; Lundin, J.; Haglund, C.; Hagstrom, J. Bmi-1 Expression Predicts Prognosis in Squamous Cell Carcinoma of the Tongue. Br. J. Cancer 2010, 102, 892–897. [Google Scholar]
- Hsu, D.S.; Lan, H.Y.; Huang, C.H.; Tai, S.K.; Chang, S.Y.; Tsai, T.L.; Chang, C.C.; Tzeng, C.H.; Wu, K.J.; Kao, J.Y.; et al. Regulation of Excision Repair Cross-Complementation Group 1 by Snail Contributes to Cisplatin Resistance in Head and Neck Cancer. Clin. Cancer Res. 2010, 16, 4561–4571. [Google Scholar]
- Semenza, G.L. Defining the Role of Hypoxia-Inducible Factor 1 in Cancer Biology and Therapeutics. Oncogene 2010, 29, 625–634. [Google Scholar]
- Mizukami, Y.; Kohgo, Y.; Chung, D.C. Hypoxia Inducible Factor-1 Independent Pathways in Tumor Angiogenesis. Clin. Cancer Res. 2007, 13, 5670–5674. [Google Scholar]
- Uehara, M.; Sano, K.; Ikeda, H.; Nonaka, M.; Asahina, I. Hypoxia-Inducible Factor 1 Alpha in Oral Squamous Cell Carcinoma and its Relation to Prognosis. Oral Oncol. 2009, 45, 241–246. [Google Scholar]
- Gort, E.H.; Groot, A.J.; van der Wall, E.; van Diest, P.J.; Vooijs, M.A. Hypoxic Regulation of Metastasis Via Hypoxia-Inducible Factors. Curr. Mol. Med. 2008, 8, 60–67. [Google Scholar]
- Yang, M.H.; Wu, K.J. TWIST Activation by Hypoxia Inducible Factor-1 (HIF-1): Implications in Metastasis and Development. Cell Cycle 2008, 7, 2090–2096. [Google Scholar]
- Rapisarda, A.; Melillo, G. Role of the Hypoxic Tumor Microenvironment in the Resistance to Anti-Angiogenic Therapies. Drug Resist. Updat. 2009, 12, 74–80. [Google Scholar]
- Goon, P.K.; Stanley, M.A.; Ebmeyer, J.; Steinstrasser, L.; Upile, T.; Jerjes, W.; Bernal-Sprekelsen, M.; Gorner, M.; Sudhoff, H.H. HPV & Head and Neck Cancer: A Descriptive Update. Head Neck Oncol. 2009, 1, 36. [Google Scholar]
- Chung, C.H.; Gillison, M.L. Human Papillomavirus in Head and Neck Cancer: Its Role in Pathogenesis and Clinical Implications. Clin. Cancer Res. 2009, 15, 6758–6762. [Google Scholar]
- Abbas, T.; Dutta, A. P21 in Cancer: Intricate Networks and Multiple Activities. Nat. Rev. Cancer 2009, 9, 400–414. [Google Scholar]
- Roninson, I.B. Oncogenic Functions of Tumour Suppressor p21(Waf1/Cip1/Sdi1): Association with Cell Senescence and Tumour-Promoting Activities of Stromal Fibroblasts. Cancer Lett. 2002, 179, 1–14. [Google Scholar]
- Fischer, C.A.; Jung, M.; Zlobec, I.; Green, E.; Storck, C.; Tornillo, L.; Lugli, A.; Wolfensberger, M.; Terracciano, L.M. Co-Overexpression of p21 and Ki-67 in Head and Neck Squamous Cell Carcinoma Relative to a significantly Poor Prognosis. Head Neck 2010, 33, 267–273. [Google Scholar]
- Hafkamp, H.C.; Mooren, J.J.; Claessen, S.M.; Klingenberg, B.; Voogd, A.C.; Bot, F.J.; Klussmann, J.P.; Hopman, A.H.; Manni, J.J.; Kremer, B.; et al. P21 Cip1/WAF1 Expression is Strongly Associated with HPV-Positive Tonsillar Carcinoma and a Favorable Prognosis. Mod. Pathol. 2009, 22, 686–698. [Google Scholar]
- Maxwell, J.H.; Kumar, B.; Feng, F.Y.; Worden, F.P.; Lee, J.S.; Eisbruch, A.; Wolf, G.T.; Prince, M.E.; Moyer, J.S.; Teknos, T.N.; et al. Tobacco use in Human Papillomavirus-Positive Advanced Oropharynx Cancer Patients Related to Increased Risk of Distant Metastases and Tumor Recurrence. Clin. Cancer Res. 2010, 16, 1226–1235. [Google Scholar]
- McDermott, U.; Downing, J.R.; Stratton, M.R. Genomics and the Continuum of Cancer Care. N Engl. J. Med. 2011, 364, 340–350. [Google Scholar]
- Saintigny, P.; Zhang, L.; Fan, Y.H.; El-Naggar, A.K.; Papadimitrakopoulou, V.A.; Feng, L.; Lee, J.J.; Kim, E.S.; Hong, W.K.; Mao, L. Gene Expression Profiling Predicts the Development of Oral Cancer. Cancer Prev. Res. 2011, 4, 218. [Google Scholar]
© 2011 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Walk, E.L.; Weed, S.A. Recently Identified Biomarkers That Promote Lymph Node Metastasis in Head and Neck Squamous Cell Carcinoma. Cancers 2011, 3, 747-772. https://doi.org/10.3390/cancers3010747
Walk EL, Weed SA. Recently Identified Biomarkers That Promote Lymph Node Metastasis in Head and Neck Squamous Cell Carcinoma. Cancers. 2011; 3(1):747-772. https://doi.org/10.3390/cancers3010747
Chicago/Turabian StyleWalk, Elyse L., and Scott A. Weed. 2011. "Recently Identified Biomarkers That Promote Lymph Node Metastasis in Head and Neck Squamous Cell Carcinoma" Cancers 3, no. 1: 747-772. https://doi.org/10.3390/cancers3010747