Effect of the Premalignant and Tumor Microenvironment on Immune Cell Cytokine Production in Head and Neck Cancer

{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results and Discussion
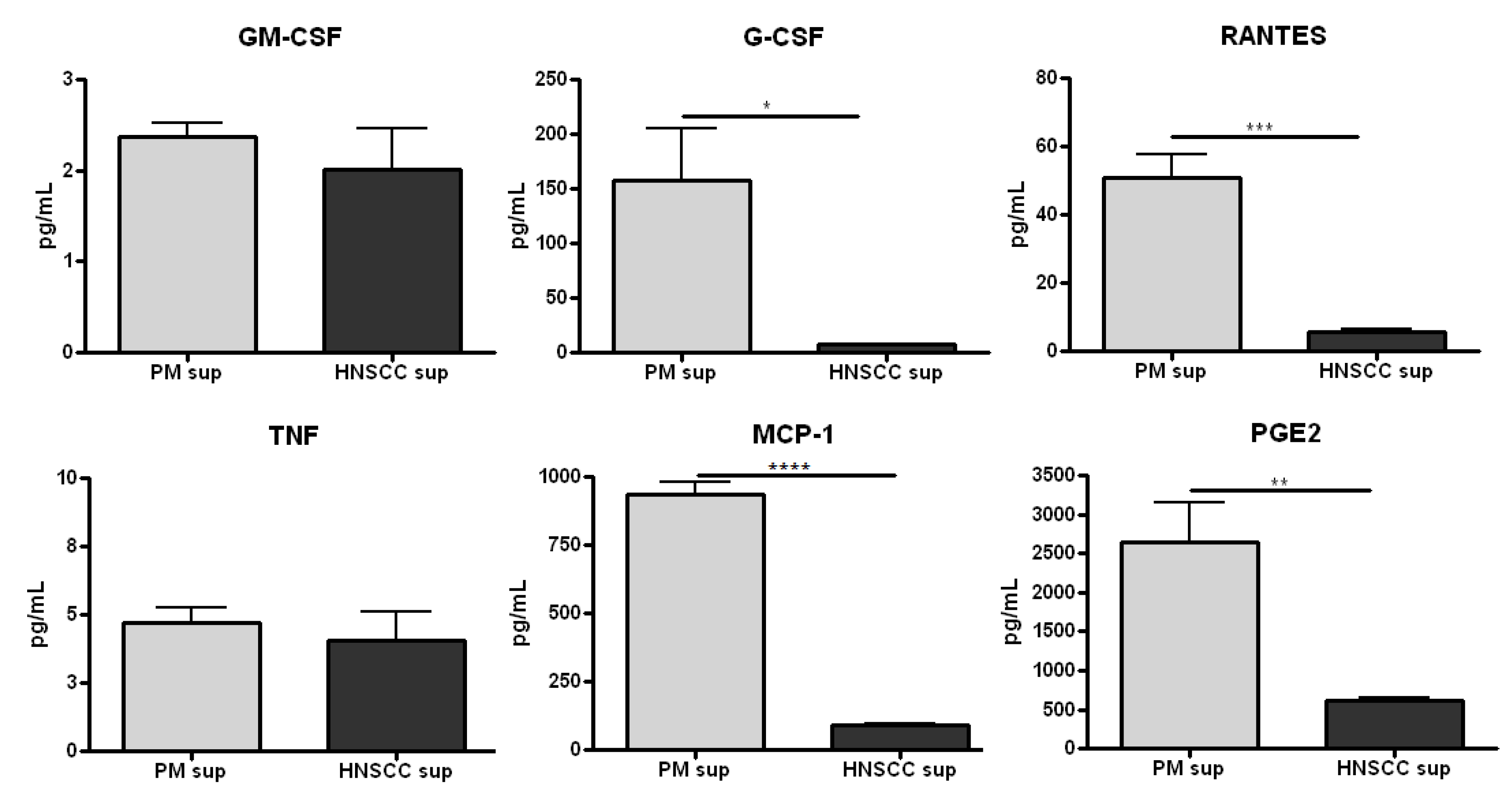
2.1. Premalignant Lesion Cells Release Significantly Increased Levels of Pro-Inflammatory Mediators Compared to HNSCC Cells in Vitro

2.2. Splenocytes Cultured with Premalignant Supernatant Secrete Significantly Increased Levels of Innate Proinflammatory Mediators Compared to Splenocytes Cultured with HNSCC Supernatant

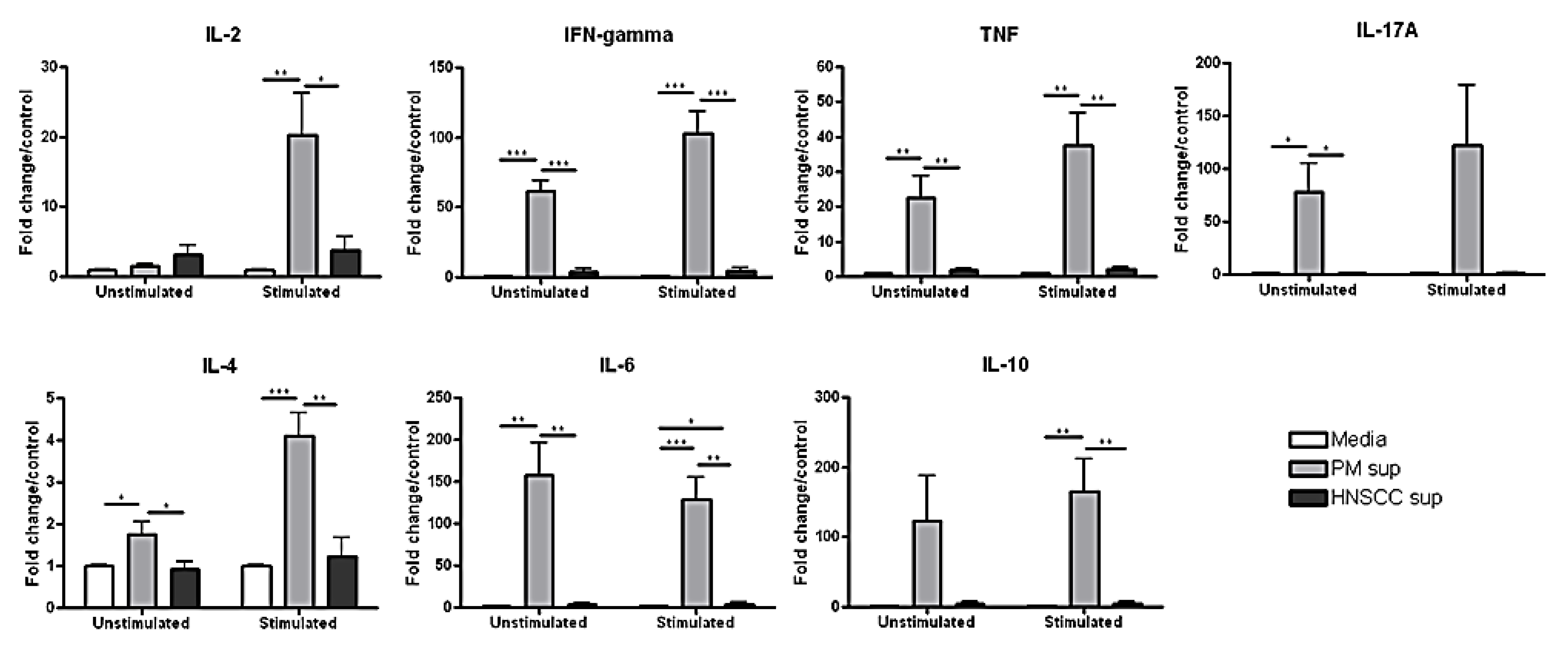
2.3. Splenocytes Cultured with Premalignant Supernatant Secrete Significantly Increased Levels of Th1-, Th2-, and Th17-Associated Cytokines Compared to Splenocytes Cultured with HNSCC Supernatant

3. Experimental
3.1. Oral HNSCC Carcinogenesis
3.2. Premalignant Lesion and HNSCC Cell Lines
3.3. Spleen Processing
3.4. Splenocyte Cultures
3.5. Cytokine Bead Array
3.6. Prostaglandin E2 ELISA
3.7. Statistical Analysis [33]
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Duray, A.; Demoulin, S.; Hubert, P.; Delvenne, P.; Saussez, S. Immune suppression in head and neck cancers: A review. Clin. Dev. Immunol. 2010. [Google Scholar] [CrossRef]
- Grandis, J.R.; Pietenpol, J.A.; Greenberger, J.S.; Pelroy, R.A.; Mohla, S. Head and neck cancer: Meeting summary and research opportunities. Cancer Res. 2004, 64, 8126–8129. [Google Scholar] [CrossRef]
- Bergmann, C.; Strauss, L.; Wang, Y.; Szczepanski, M.J.; Lang, S.; Johnson, J.T.; Whiteside, T.L. T regulatory type I cells in squamous cell carcinoma of the head and neck: Mechanisms of suppression and expansion in advanced disease. Clin. Cancer Res. 2008, 14, 3706–3715. [Google Scholar] [CrossRef]
- Badoual, C.; Hans, S.; Rodriguez, J.; Peyrard, S.; Klein, C.; Agueznay Nel, H.; Mosseri, V.; Laccourreye, O.; Bruneval, P.; Fridman, W.H.; et al. Prognostic value of tumor-infiltrating CD4+ T-cell subpopulations in head and neck cancers. Clin. Cancer Res. 2006, 12, 465–472. [Google Scholar] [CrossRef]
- Young, M.R.; Wright, M.A.; Lozano, Y.; Prechel, M.M.; Benefield, J.; Leonetti, J.P.; Collins, S.L.; Petruzzelli, G.J. Increased recurrence and metastasis in patients whose primary head and neck squamous cell carcinomas secreted granulocyte-macrophage colony-stimulating factor and contained CD34+ natural suppressor cells. Int. J. Cancer 1997, 74, 69–74. [Google Scholar] [CrossRef]
- Chikamatsu, K.; Sakakura, K.; Whiteside, T.L.; Furuya, N. Relationships between regulatory T cells and CD8+ effector populations in patients with squamous cell carcinoma of the head and neck. Head Neck 2007, 29, 120–127. [Google Scholar]
- Drennan, S.; Stafford, N.D.; Greenman, J.; Green, V.L. Increased frequency and suppressive activity of CD127(low/−) regulatory T cells in the peripheral circulation of patients with head and neck squamous cell carcinoma are associated with advanced stage and nodal involvement. Immunology 2013, 140, 335–343. [Google Scholar]
- Koontongkaew, S. The tumor microenvironment contribution to development, growth, invasion and metastasis of head and neck squamous cell carcinomas. J. Cancer 2013, 4, 66–83. [Google Scholar] [CrossRef]
- Badoual, C.; Sandoval, F.; Pere, H.; Hans, S.; Gey, A.; Merillon, N.; van Ryswick, C.; Quintin-Colonna, F.; Bruneval, P.; Brasnu, D.; et al. Better understanding tumor-host interaction in head and neck cancer to improve the design and development of immunotherapeutic strategies. Head Neck 2010, 32, 946–958. [Google Scholar]
- Young, M.R. Protective mechanisms of head and neck squamous cell carcinomas from immune assault. Head Neck 2006, 28, 462–470. [Google Scholar] [CrossRef]
- Ferris, R.L.; Whiteside, T.L.; Ferrone, S. Immune escape associated with functional defects in antigen-processing machinery in head and neck cancer. Clin. Cancer Res. 2006, 12, 3890–3895. [Google Scholar] [CrossRef]
- Ogino, T.; Shigyo, H.; Ishii, H.; Katayama, A.; Miyokawa, N.; Harabuchi, Y.; Ferrone, S. Hla class I antigen down-regulation in primary laryngeal squamous cell carcinoma lesions as a poor prognostic marker. Cancer Res. 2006, 66, 9281–9289. [Google Scholar] [CrossRef]
- Grandis, J.R.; Falkner, D.M.; Melhem, M.F.; Gooding, W.E.; Drenning, S.D.; Morel, P.A. Human leukocyte antigen class I allelic and haplotype loss in squamous cell carcinoma of the head and neck: Clinical and immunogenetic consequences. Clin. Cancer Res. 2000, 6, 2794–2802. [Google Scholar]
- Zeng, X.; Chen, Q.; Nie, M. The relationship of Fas and FasL protein expression in oral carcinogenesis. West China J. Stomatol. 2003, 21, 259–262. [Google Scholar]
- Cho, Y.A.; Yoon, H.J.; Lee, J.I.; Hong, S.P.; Hong, S.D. Relationship between the expressions of PD-L1 and tumor-infiltrating lymphocytes in oral squamous cell carcinoma. Oral Oncol. 2011, 47, 1148–1153. [Google Scholar] [CrossRef]
- Jewett, A.; Head, C.; Cacalano, N.A. Emerging mechanisms of immunosuppression in oral cancers. J. Dent. Res. 2006, 85, 1061–1073. [Google Scholar] [CrossRef]
- Bose, A.; Chakraborty, T.; Chakraborty, K.; Pal, S.; Baral, R. Dysregulation in immune functions is reflected in tumor cell cytotoxicity by peripheral blood mononuclear cells from head and neck squamous cell carcinoma patients. Cancer Immun. 2008, 8, 1–10. [Google Scholar]
- Lathers, D.M.; Achille, N.J.; Young, M.R. Incomplete Th2 skewing of cytokines in plasma of patients with squamous cell carcinoma of the head and neck. Hum. Immunol. 2003, 64, 1160–1166. [Google Scholar] [CrossRef]
- Lu, S.L.; Reh, D.; Li, A.G.; Woods, J.; Corless, C.L.; Kulesz-Martin, M.; Wang, X.J. Overexpression of transforming growth factor beta1 in head and neck epithelia results in inflammation, angiogenesis, and epithelial hyperproliferation. Cancer Res. 2004, 64, 4405–4410. [Google Scholar] [CrossRef]
- Wahl, S.M.; Wen, J.; Moutsopoulos, N. TGF-beta: A mobile purveyor of immune privilege. Immunol. Rev. 2006, 213, 213–227. [Google Scholar] [CrossRef]
- Nitsch, S.M.; Pries, R.; Wollenberg, B. Head and neck cancer triggers increased IL-6 production of CD34+ stem cells from human cord blood. In Vivo 2007, 21, 493–498. [Google Scholar]
- Chen, Z.; Malhotra, P.S.; Thomas, G.R.; Ondrey, F.G.; Duffey, D.C.; Smith, C.W.; Enamorado, I.; Yeh, N.T.; Kroog, G.S.; Rudy, S.; et al. Expression of proinflammatory and proangiogenic cytokines in patients with head and neck cancer. Clin. Cancer Res. 1999, 5, 1369–1379. [Google Scholar]
- Bronte, V.; Chappell, D.B.; Apolloni, E.; Cabrelle, A.; Wang, M.; Hwu, P.; Restifo, N.P. Unopposed production of granulocyte-macrophage colony-stimulating factor by tumors inhibits CD8+ T cell responses by dysregulating antigen-presenting cell maturation. J. Immunol. 1999, 162, 5728–5737. [Google Scholar]
- Hambek, M.; Baghi, M.; Wagenblast, J.; Schmitt, J.; Baumann, H.; Knecht, R. Inverse correlation between serum PGE2 and T classification in head and neck cancer. Head Neck 2007, 29, 244–248. [Google Scholar] [CrossRef]
- Liss, C.; Fekete, M.J.; Hasina, R.; Lam, C.D.; Lingen, M.W. Paracrine angiogenic loop between head-and-neck squamous-cell carcinomas and macrophages. Int. J. Cancer 2001, 93, 781–785. [Google Scholar] [CrossRef]
- Forastiere, A.; Koch, W.; Trotti, A.; Sidransky, D. Head and neck cancer. N. Engl. J. Med. 2001, 345, 1890–1900. [Google Scholar] [CrossRef]
- Schoop, R.A.; Noteborn, M.H.; Baatenburg de Jong, R.J. A mouse model for oral squamous cell carcinoma. J. Mol. Histol. 2009, 40, 177–181. [Google Scholar] [CrossRef]
- De Costa, A.M.; Schuyler, C.A.; Walker, D.D.; Young, M.R. Characterization of the evolution of immune phenotype during the development and progression of squamous cell carcinoma of the head and neck. Cancer Immunol. Immunother. 2012, 61, 927–939. [Google Scholar] [CrossRef]
- Smith, C.W.; Chen, Z.; Dong, G.; Loukinova, E.; Pegram, M.Y.; Nicholas-Figueroa, L.; van Waes, C. The host environment promotes the development of primary and metastatic squamous cell carcinomas that constitutively express proinflammatory cytokines IL-1alpha, IL-6, GM-CSF, and KC. Clin. Exp. Metastasis 1998, 16, 655–664. [Google Scholar] [CrossRef]
- Mann, E.A.; Spiro, J.D.; Chen, L.L.; Kreutzer, D.L. Cytokine expression by head and neck squamous cell carcinomas. Am. J. Surg. 1992, 164, 567–573. [Google Scholar] [CrossRef]
- Whiteside, T.L. Immunobiology of head and neck cancer. Cancer Metastasis Rev. 2005, 24, 95–105. [Google Scholar] [CrossRef]
- Reichert, T.E.; Rabinowich, H.; Johnson, J.T.; Whiteside, T.L. Mechanisms responsible for signaling and functional defects. J. Immunother. 1998, 21, 295–306. [Google Scholar] [CrossRef]
- GraphPad Prism, verion 6.0.; GraphPad-Prism Software Inc.: Sandiego, CA, USA, 2013.
- Almand, B.; Clark, J.I.; Nikitina, E.; van Beynen, J.; English, N.R.; Knight, S.C.; Carbone, D.P.; Gabrilovich, D.I. Increased production of immature myeloid cells in cancer patients: A mechanism of immunosuppression in cancer. J. Immunol. 2001, 166, 678–689. [Google Scholar]
- Pak, A.S.; Wright, M.A.; Matthews, J.P.; Collins, S.L.; Petruzzelli, G.J.; Young, M.R. Mechanisms of immune suppression in patients with head and neck cancer: Presence of CD34+ cells which suppress immune functions within cancers that secrete granulocyte-macrophage colony-stimulating factor. Clin. Cancer Res. 1995, 1, 95–103. [Google Scholar]
- Ninck, S.; Reisser, C.; Dyckhoff, G.; Helmke, B.; Bauer, H.; Herold-Mende, C. Expression profiles of angiogenic growth factors in squamous cell carcinomas of the head and neck. Int. J. Cancer 2003, 106, 34–44. [Google Scholar] [CrossRef]
- Sugimoto, C.; Fujieda, S.; Sunaga, H.; Noda, I.; Tanaka, N.; Kimura, Y.; Saito, H.; Matsukawa, S. Granulocyte colony-stimulating factor (G-CSF)-mediated signaling regulates type IV collagenase activity in head and neck cancer cells. Int. J. Cancer 2001, 93, 42–46. [Google Scholar] [CrossRef]
- Tsuzuki, H.; Fujieda, S.; Sunaga, H.; Noda, I.; Saito, H. Expression of granulocyte colony-stimulating factor receptor correlates with prognosis in oral and mesopharyngeal carcinoma. Cancer Res. 1998, 58, 794–800. [Google Scholar]
- Linkov, F.; Lisovich, A.; Yurkovetsky, Z.; Marrangoni, A.; Velikokhatnaya, L.; Nolen, B.; Winans, M.; Bigbee, W.; Siegfried, J.; Lokshin, A.; et al. Early detection of head and neck cancer: Development of a novel screening tool using multiplexed immunobead-based biomarker profiling. Cancer Epidemiol. Biomark. Prev. 2007, 16, 102–107. [Google Scholar] [CrossRef]
- Young, M.R.; Wright, M.A.; Lozano, Y.; Matthews, J.P.; Benefield, J.; Prechel, M.M. Mechanisms of immune suppression in patients with head and neck cancer: Influence on the immune infiltrate of the cancer. Int. J. Cancer 1996, 67, 333–338. [Google Scholar] [CrossRef]
- Abrahao, A.C.; Castilho, R.M.; Squarize, C.H.; Molinolo, A.A.; dos Santos-Pinto, D., Jr.; Gutkind, J.S. A role for Cox2-derived PGE2 and PGE2-receptor subtypes in head and neck squamous carcinoma cell proliferation. Oral Oncol. 2010, 46, 880–887. [Google Scholar] [CrossRef]
- Bao, Y.S.; Zhang, P.; Xie, R.J.; Wang, M.; Wang, Z.Y.; Zhou, Z.; Zhai, W.J.; Feng, S.Z.; Han, M.Z. The regulation of CD4+ T cell immune responses toward Th2 cell development by prostaglandin E2. Int. Immunopharmacol. 2011, 11, 1599–1605. [Google Scholar] [CrossRef]
- Kalinski, P. Regulation of immune responses by prostaglandin E2. J. Immunol. 2012, 188, 21–28. [Google Scholar] [CrossRef]
- MacKenzie, K.F.; Clark, K.; Naqvi, S.; McGuire, V.A.; Noehren, G.; Kristariyanto, Y.; van den Bosch, M.; Mudaliar, M.; McCarthy, P.C.; Pattison, M.J.; et al. PGE(2) induces macrophage IL-10 production and a regulatory-like phenotype via a protein kinase A-SIK-CRTC3 pathway. J. Immunol. 2013, 190, 565–577. [Google Scholar] [CrossRef]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Johnson, S.D.; De Costa, A.-M.A.; Young, M.R.I. Effect of the Premalignant and Tumor Microenvironment on Immune Cell Cytokine Production in Head and Neck Cancer. Cancers 2014, 6, 756-770. https://doi.org/10.3390/cancers6020756
Johnson SD, De Costa A-MA, Young MRI. Effect of the Premalignant and Tumor Microenvironment on Immune Cell Cytokine Production in Head and Neck Cancer. Cancers. 2014; 6(2):756-770. https://doi.org/10.3390/cancers6020756
Chicago/Turabian StyleJohnson, Sara D., Anna-Maria A. De Costa, and M. Rita I. Young. 2014. "Effect of the Premalignant and Tumor Microenvironment on Immune Cell Cytokine Production in Head and Neck Cancer" Cancers 6, no. 2: 756-770. https://doi.org/10.3390/cancers6020756