Simultaneous Expression of Cancer Stem Cell-Like Properties and Cancer-Associated Fibroblast-Like Properties in a Primary Culture of Breast Cancer Cells

Abstract
:1. Introduction
2. Results and Discussion
2.1. Gene Expression Profile of MCKH Cells

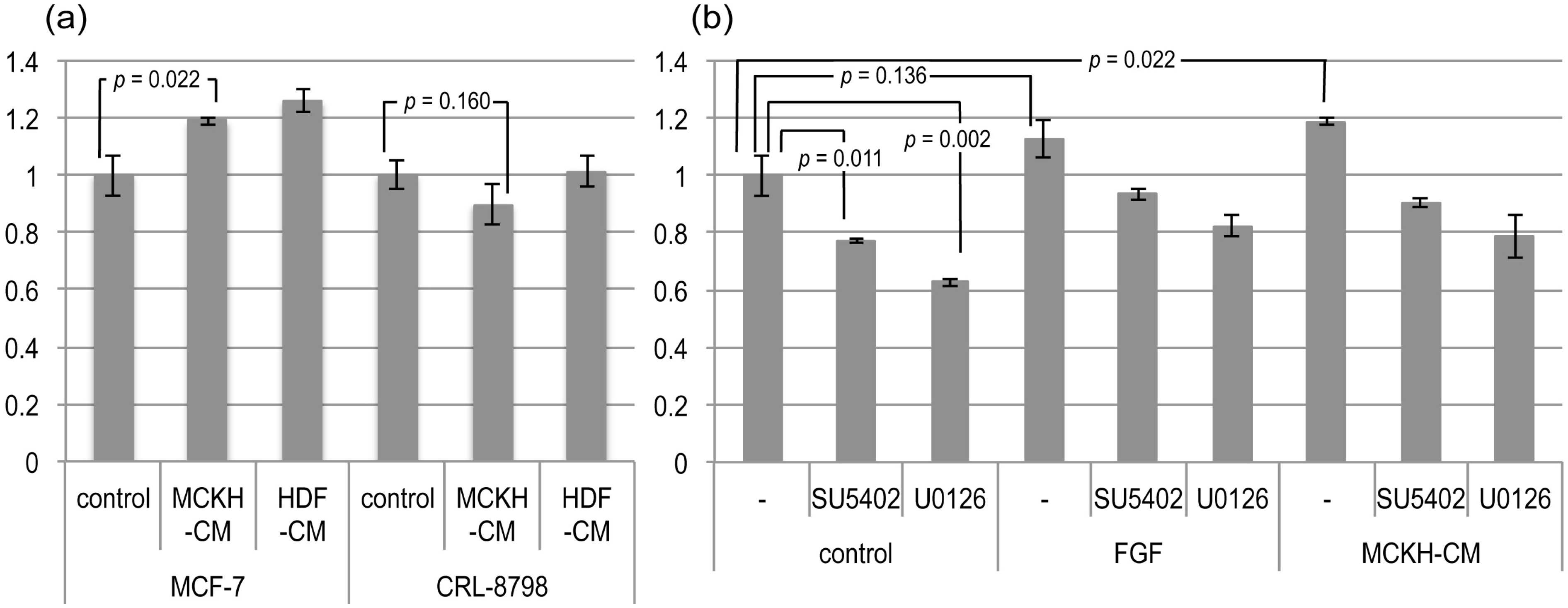{kind=link}
| Gene name | Gene | malignant tissue * | CRL8798 * | MCKH * | |||
|---|---|---|---|---|---|---|---|
| transcription factors | |||||||
| HIF1A | hypoxia inducible factor 1, α subunit | 1.28 ± 0.16 | 3.93 ± 0.16 | 9.40 ± 1.30 | |||
| NFKB2 | NF-κB2 | 1.04 ± 0.12 | 4.25 ± 0.50 | 2.07 ± 0.30 | |||
| BNIP3 | BCL2/adenovirus E1B interacting protein 3 | 0.77 ± 0.02 | 11.72 ± 0.23 | 20.85 ± 0.72 | |||
| glycolytic enzymes | |||||||
| GAPDH | glyceraldehyde-3-phosphate dehydrogenase | 1.08 ± 0.02 | 8.11 ± 0.23 | 16.78 ± 0.47 | |||
| LDHA | lactate dehydrogenase A | 1.05 ± 0.05 | 21.22 ± 3.08 | 32.17 ± 5.40 | |||
| PGAM1 | phosphoglycerate mutase 1 | 1.3 ± 0.50 | 10.84 ± 0.49 | 17.22 ± 2.79 | |||
| PGK1 | phosphoglycerate kinase 1 | 1.35 ± 0.04 | 10.83 ± 0.19 | 15.67 ± 0.62 | |||
| ALDOA | aldolase A, fructose-bisphosphate | 1.81 | 5.95 | 8.39 | |||
| ENO1 | enolase 1, α | 0.58 | 12.00 | 10.15 | |||
| PKM2 | pyruvate kinase, muscle | 1.31 | 11.59 | 18.56 | |||
| TPI1 | triosephosphate isomerase 1 | 1.06 | 9.04 | 13.87 | |||
| transporters | |||||||
| SLC16A1 | monocarboxylic acid transporter 1 | 0.97 | 14.34 | 12.03 | |||
| SLC16A3 | monocarboxylic acid transporter 4 | 5.44 | 116.02 | 136.50 | |||
| SLC1A5 | neutral amino acid transporter | 0.50 | 4.16 | 25.87 | |||
| others | |||||||
| COLA2 | collagen, type I, α2 | 26.20 ± 1.70 | 0.0006 ± 0.0002 | 35.32 ± 2.13 | |||
| MMP1** | matrix metallopeptidase 1 | 1.05 ± 0.55 | 52.62 ± 19.16 | 2942.30 ± 1107.47 | |||
| CAV1 | caveolin 1, caveolae protein | 0.28 ± 0.02 | 4.17 ± 0.36 | 0.95 ± 0.05 | |||
| CAV2 | caveolin 2 | 0.34 ± 0.03 | 4.64 ± 1.40 | 2.31 ± 0.67 | |||
| AKT1 | v-akt murine thymoma viral oncogene homolog 1 | 1.33 | 2.22 | 3.22 | |||
| ANGPT1 | angiopoietin 1 (ANGPT1) | 0.63 | 0.07 | 6.59 | |||
| FGF1 | fibroblast growth factor 1 (acidic) | 0.66 ± 0.41 | 0.53 ± 0.68 | 2.49 ± 0.97 | |||
| FGF2 | fibroblast growth factor 2 (basic) | 0.22 ± 0.01 | 0.98 ± 0.05 | 9.38 ± 0.67 | |||
| TGFB1** | transforming growth factor, β1 | 3.59 ± 0.20 | 6.31 ± 0.28 | 10.04 ± 0.54 | |||
| VEGFA | vascular endothelial growth factor A | 0.86 ± 0.01 | 1.96 ± 0.33 | 8.88 ± 2.73 | |||
| CXCL12 | chemokine (C-X-C motif) ligand 12 | 0.20 ± 0.005 | 0.0003 ± 0.0001 | 0.03 ± 0.002 | |||
| CXCR4 | chemokine (C-X-C motif) receptor 4 | 2.87 ± 0.07 | 0.003 ± 0.0003 | 0.008 ± 0.0008 | |||
| SACTA2 | actin, α2, smooth muscle | 0.43 | 0.04 | 2.10 | |||
2.2. CAF Properties of MCKH Cells
| Gene name | Gene | malignant tissue * | CRL8798 * | MCKH * |
|---|---|---|---|---|
| telomerase reverse transcriptase | ||||
| TERT | TERT | 1.05 ± 0.22 | 1.13 ± 0.25 | 0.82 ± 0.33 |
| cyclins | ||||
| CCNA2 | cyclin A2 | 1.97 ± 0.83 | 10.27 ± 4.05 | 7.92 ± 3.06 |
| CCNB1 | cyclin B1 | 0.86 ± 0.15 | 6.83 ± 1.37 | 5.72 ± 1.43 |
| CCND1 | cyclin D1 | 3.23 ± 0.11 | 5.79 ± 0.30 | 7.46 ± 0.54 |
| CCNE1 | cyclin E1 | 1.70 | 10.10 | 4.67 |
| cyclin-dependent kinases | ||||
| CDK1 | CDK1 | 1.23 ± 0.30 | 20.13 ± 3.86 | 5.36 ± 1.18 |
| CDK3 | CDK3 | 1.50 | 1.74 | 2.88 |
| CDK5 | CDK5 | 2.06 | 5.98 | 6.08 |
| CDK7 | CDK7 | 1.07 | 3.34 | 2.72 |
| cyclin-dependent kinase inhibitors | ||||
| CDKN1A ** | p21, Cip1 | 3.46 | 7.81 | 19.86 |
| CDKN2A | p16, inhibits CDK4 | 5.36 ± 1.79 | 5.10 ± 1.80 | 23.37 ± 10.36 |
| CDKN2B | p15, inhibits CDK4 | 4.69 | 7.66 | 34.42 |
| CDKN3 | CDKN3 | 2.90 ± 0.75 | 9.10 ± 1.87 | 14.23 ± 7.23 |
| others | ||||
| TP53 ** | p53 | 0.85 ± 0.24 | 2.32 ± 0.71 | 1.54 ± 0.52 |
| MDM2 | MDM2 | 2.44 ± 0.67 | 4.21 ± 0.74 | 2.61 ± 0.57 |
| RB1 ** | retinoblastoma 1 | 1.15 ± 0.06 | 1.00 ± 0.04 | 0.84 ± 0.05 |
2.3. What Are MCKH Cells?

3. Experimental
3.1. Cells and Cell Culture
3.2. Gene Expression Profiles
3.3. CAF Assays
3.4. Inhibition Assays
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Fedele, C.; Tothill, R.W.; McArthur, G.A. Navigating the challenge of tumor heterogeneity in cancer therapy. Cancer Discov. 2014, 4, 146–148. [Google Scholar] [CrossRef] [PubMed]
- Hanahan, D.; Weinberg, R.A. Hallmarks of cancer: The next generation. Cell 2011, 144, 646–674. [Google Scholar] [CrossRef] [PubMed]
- Orimo, A.; Gupta, P.B.; Sgroi, D.C.; Arenzana-Seisdedos, F.; Delaunay, T.; Naeem, R.; Carey, V.J.; Richardson, A.L.; Weinberg, R.A. Stromal fibroblasts present in invasive human breast carcinomas promote tumor growth and angiogenesis through elevated SDF-1/CXCL12 secretion. Cell 2005, 121, 335–348. [Google Scholar] [CrossRef] [PubMed]
- Pietras, K.; Ostman, A. Hallmarks of cancer: Interactions with the tumor stroma. Exp. Cell Res. 2010, 316, 1324–1331. [Google Scholar] [CrossRef] [PubMed]
- Bhowmick, N.A.; Neilson, E.G.; Moses, H.L. Stromal fibroblasts in cancer initiation and progression. Nature 2004, 18, 332–337. [Google Scholar] [CrossRef]
- Tripathi, M.; Billet, S.; Bhowmick, N.A. Understanding the role of stromal fibroblasts in cancer progression. Cell Adhes. Migr. 2012, 6, 231–235. [Google Scholar] [CrossRef]
- Nishikata, T.; Ishikawa, M.; Matsuyama, T.; Takamatsu, K.; Fukuhara, T.; Konishi, Y. Primary culture of breast cancer: A model system for epithelial-mesenchymal transition and cancer stem cells. Anticancer Res. 2013, 33, 2867–2874. [Google Scholar] [PubMed]
- Sappino, A.P.; Skalli, O.; Jackson, B.; Schürch, W.; Gabbiani, G. Smooth-muscle differentiation in stromal cells of malignant and non-malignant breast tissues. Int. J. Cancer 1988, 41, 707–712. [Google Scholar] [CrossRef] [PubMed]
- Xouri, G.; Christian, S. Origin and function of tumor stroma fibroblasts. Semin. Cell Dev. Biol. 2010, 21, 40–46. [Google Scholar] [CrossRef] [PubMed]
- Eck, S.M.; Côté, A.L.; Winkelman, W.D.; Brinckerhoff, C.E. CXCR4 and matrix metalloproteinase-1 are elevated in breast carcinoma-associated fibroblasts and in normal mammary fibroblasts exposed to factors secreted by breast cancer cells. Mol. Cancer Res. 2009, 7, 1033–1044. [Google Scholar] [CrossRef] [PubMed]
- Franco, O.E.; Shaw, A.K.; Strand, D.W.; Hayward, S.W. Cancer associated fibroblasts in cancer pathogenesis. Semin. Cell Dev. Biol. 2010, 21, 33–39. [Google Scholar] [CrossRef] [PubMed]
- Zhong, H.; de Marzo, A.M.; Laughner, E.; Lim, M.; Hilton, D.A.; Zagzag, D.; Buechler, P.; Issacs, W.B.; Semenza, G.L.; Simons, J.W. Overexpression of hypoxia-inducible factor 1α in common human cancers and their metastases. Cancer Res. 1999, 59, 5830–5835. [Google Scholar] [PubMed]
- Martinez-Outschoorn, U.E.; Trimmer, C.; Lin, Z.; Whitaker-Menezes, D.; Chiavarina, B.; Zhou, J.; Wang, C.; Pavlides, S.; Martinez-Cantarin, M.P.; Capozza, F.; et al. Autophagy in cancer associated fibroblasts promotes tumor cell survival: Role of hypoxia, HIF1 induction and NF-κB activation in the tumor stromal microenvironment. Cell Cycle 2010, 9, 3515–3533. [Google Scholar] [CrossRef] [PubMed]
- Bonuccelli, G.; Whitaker-Menezes, D.; Castello-Cros, R.; Pavlides, S.; Pestell, R.G.; Fatatis, A.; Witkiewicz, A.K.; vander Heiden, M.G.; Migneco, G.; Chiavarina, B.; et al. The reverse Warburg effect: Glycolysis inhibitors prevent the tumor promoting effects of caveolin-1 deficient cancer associated fibroblasts. Cell Cycle 2010, 9, 1960–1971. [Google Scholar]
- Carreira, S.; Goodall, J.; Aksan, I.; la Rocca, S.A.; Galibert, M.D.; Denat, L.; Larue, L.; Goding, C.R. Mitf cooperates with Rb1 and activates p21(Cip1) expression to regulate cell cycle progression. Nature 2005, 433, 764–769. [Google Scholar] [CrossRef] [PubMed]
- Shintani, Y.; Abulaiti, A.; Kimura, T.; Funaki, S.; Nakagiri, T.; Inoue, M.; Sawabata, N.; Minami, M.; Morii, E.; Okumura, M. Pulmonary fibroblasts induce epithelial mesenchymal transition and some characteristics of stem cells in non-small cell lung cancer. Ann. Thorac. Surg. 2013, 96, 425–433. [Google Scholar] [CrossRef] [PubMed]
- Kiyonari, H.; Kaneko, M.; Abe, S.; Aizawa, S. Three inhibitors of FGF receptor, ERK, and GSK3 establishes germline-competent embryonic stem cells of C57BL/6N mouse strain with high efficiency and stability. Genesis 2010, 48, 317–327. [Google Scholar] [PubMed]
- Favata, M.F.; Horiuchi, K.Y.; Manos, E.J.; Daulerio, A.J.; Stradley, D.A.; Feeser, W.S.; van Dyk, D.E.; Pitts, W.J.; Earl, R.A.; Hobbs, F.; et al. Identification of a novel inhibitor of mitogen-activated protein kinase kinase. J. Biol. Chem. 1998, 273, 18623–18632. [Google Scholar] [CrossRef] [PubMed]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Ishikawa, M.; Inoue, T.; Shirai, T.; Takamatsu, K.; Kunihiro, S.; Ishii, H.; Nishikata, T. Simultaneous Expression of Cancer Stem Cell-Like Properties and Cancer-Associated Fibroblast-Like Properties in a Primary Culture of Breast Cancer Cells. Cancers 2014, 6, 1570-1578. https://doi.org/10.3390/cancers6031570
Ishikawa M, Inoue T, Shirai T, Takamatsu K, Kunihiro S, Ishii H, Nishikata T. Simultaneous Expression of Cancer Stem Cell-Like Properties and Cancer-Associated Fibroblast-Like Properties in a Primary Culture of Breast Cancer Cells. Cancers. 2014; 6(3):1570-1578. https://doi.org/10.3390/cancers6031570
Chicago/Turabian StyleIshikawa, Mami, Takahiro Inoue, Takuma Shirai, Kazuhiko Takamatsu, Shiori Kunihiro, Hirokazu Ishii, and Takahito Nishikata. 2014. "Simultaneous Expression of Cancer Stem Cell-Like Properties and Cancer-Associated Fibroblast-Like Properties in a Primary Culture of Breast Cancer Cells" Cancers 6, no. 3: 1570-1578. https://doi.org/10.3390/cancers6031570