
Involvement of the Integrin α1β1 in the Progression of Colorectal Cancer


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
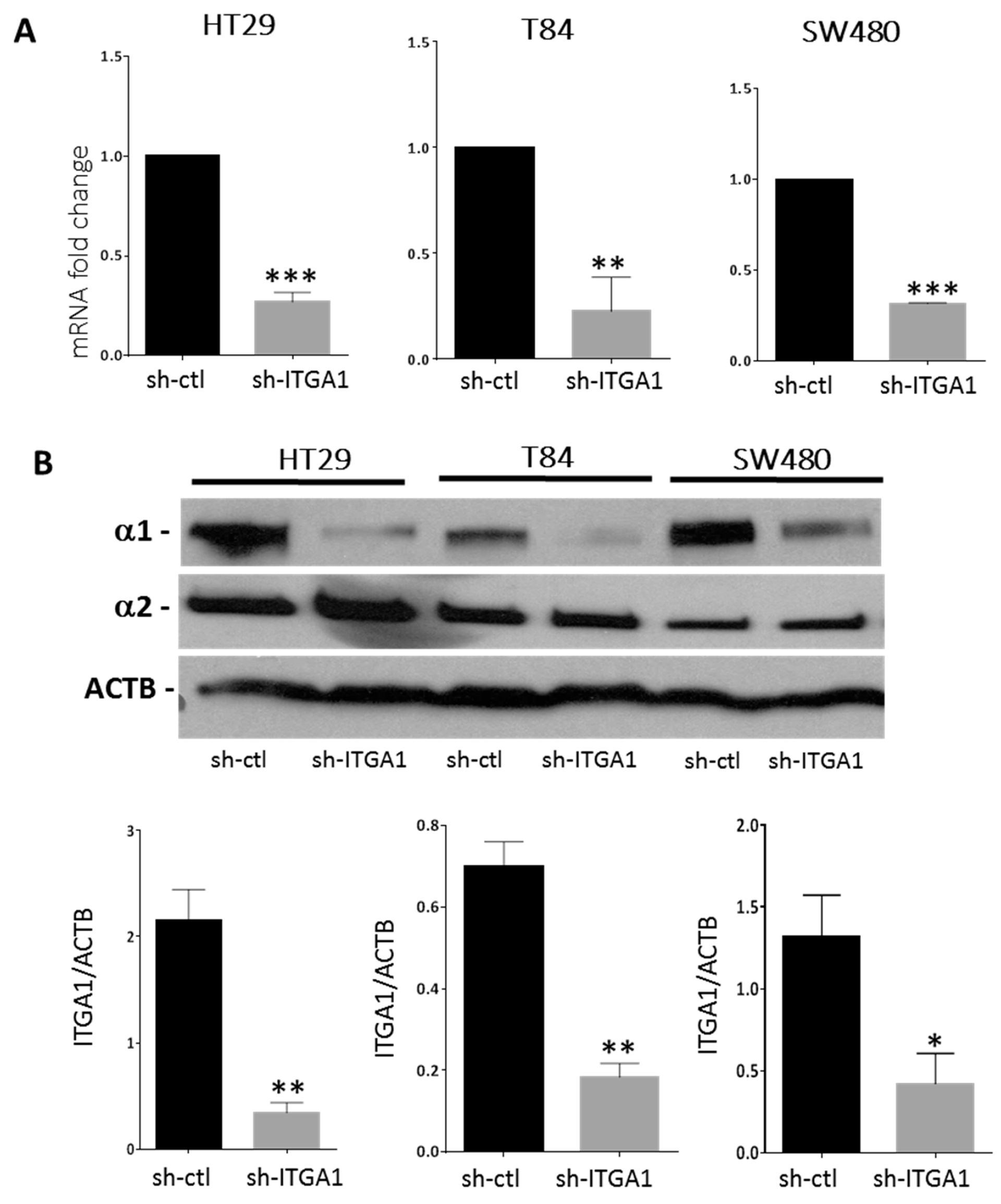
2.1. Integrin α1 Subunit/ITGA1 Knockdown in CRC Cells
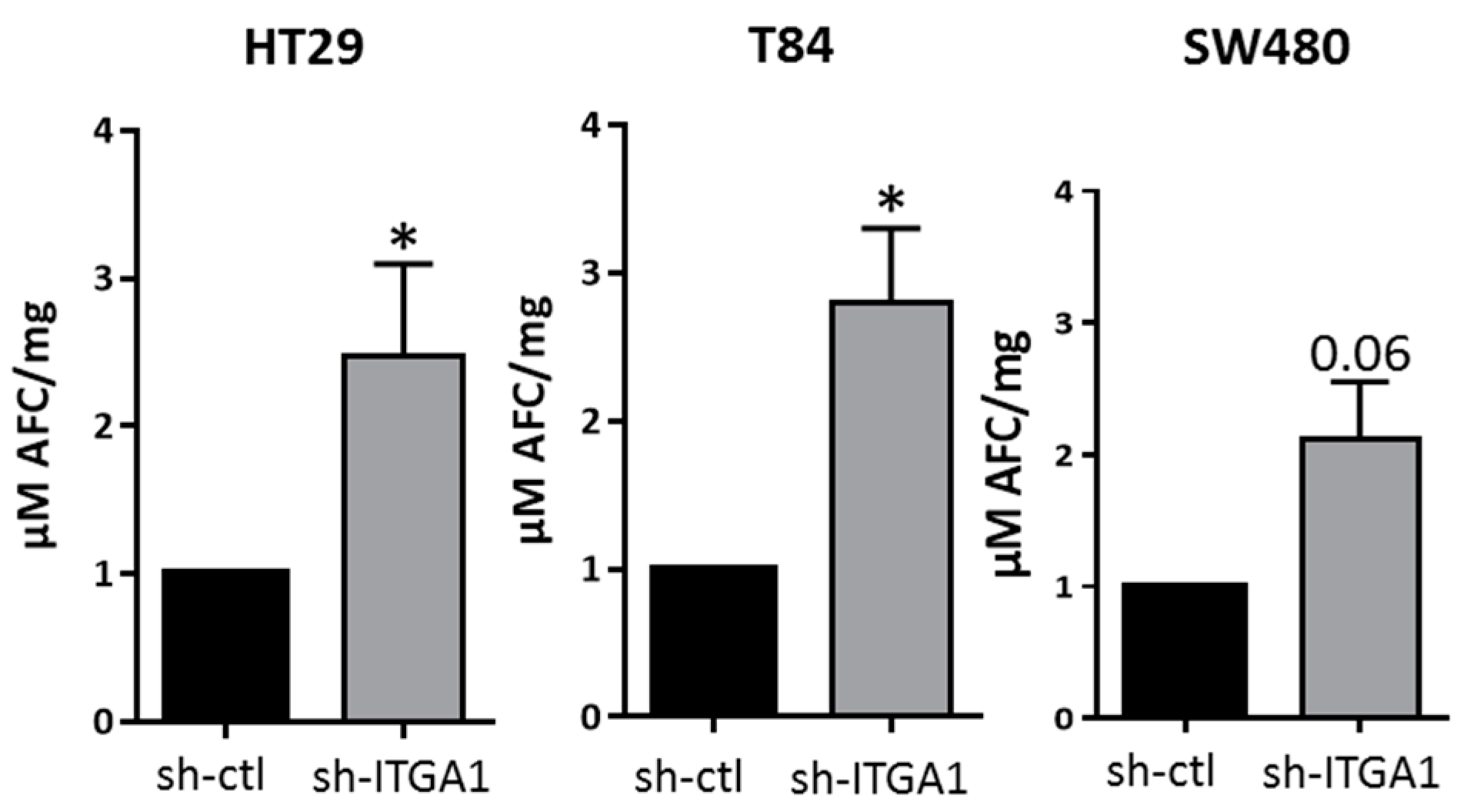
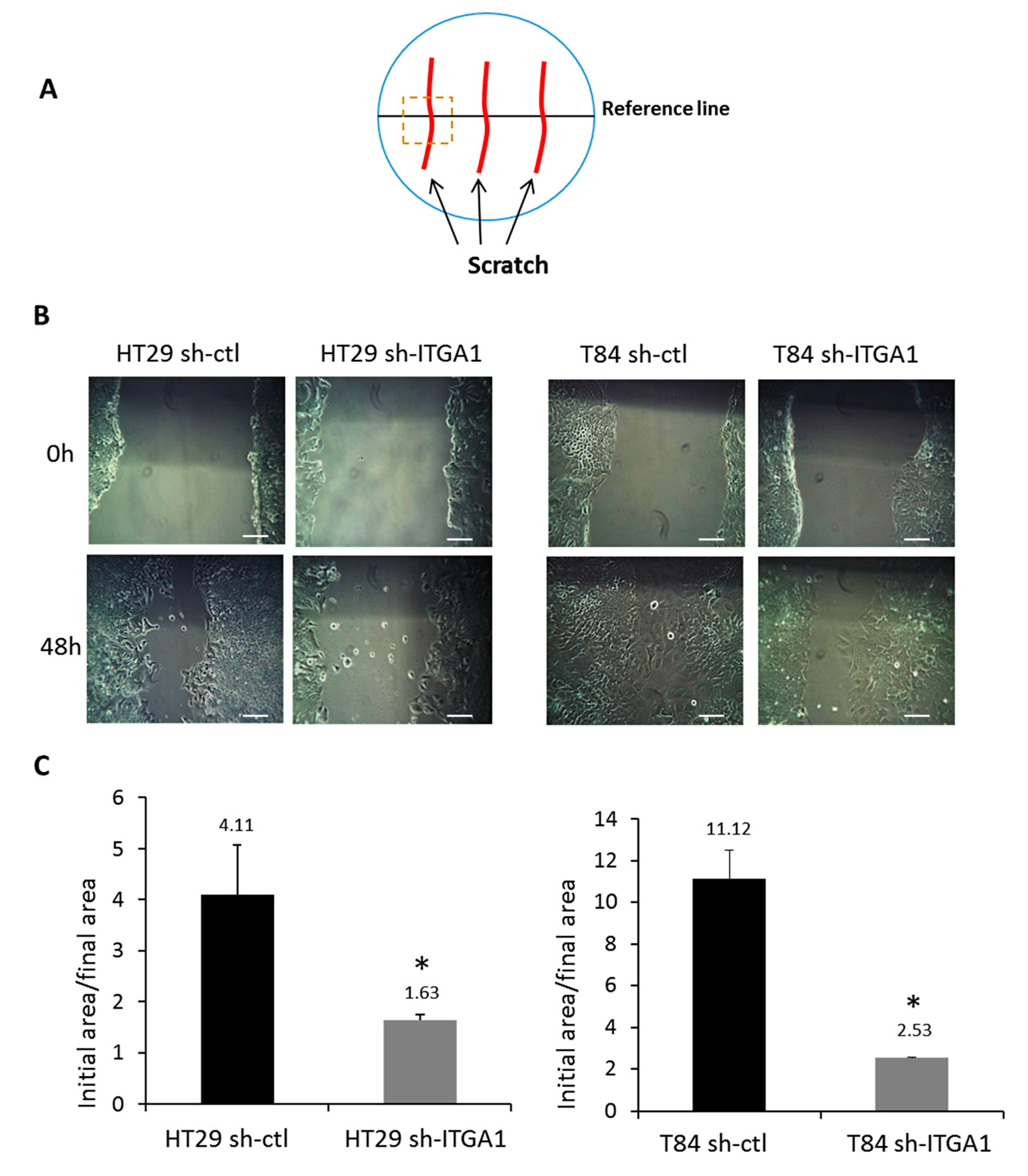
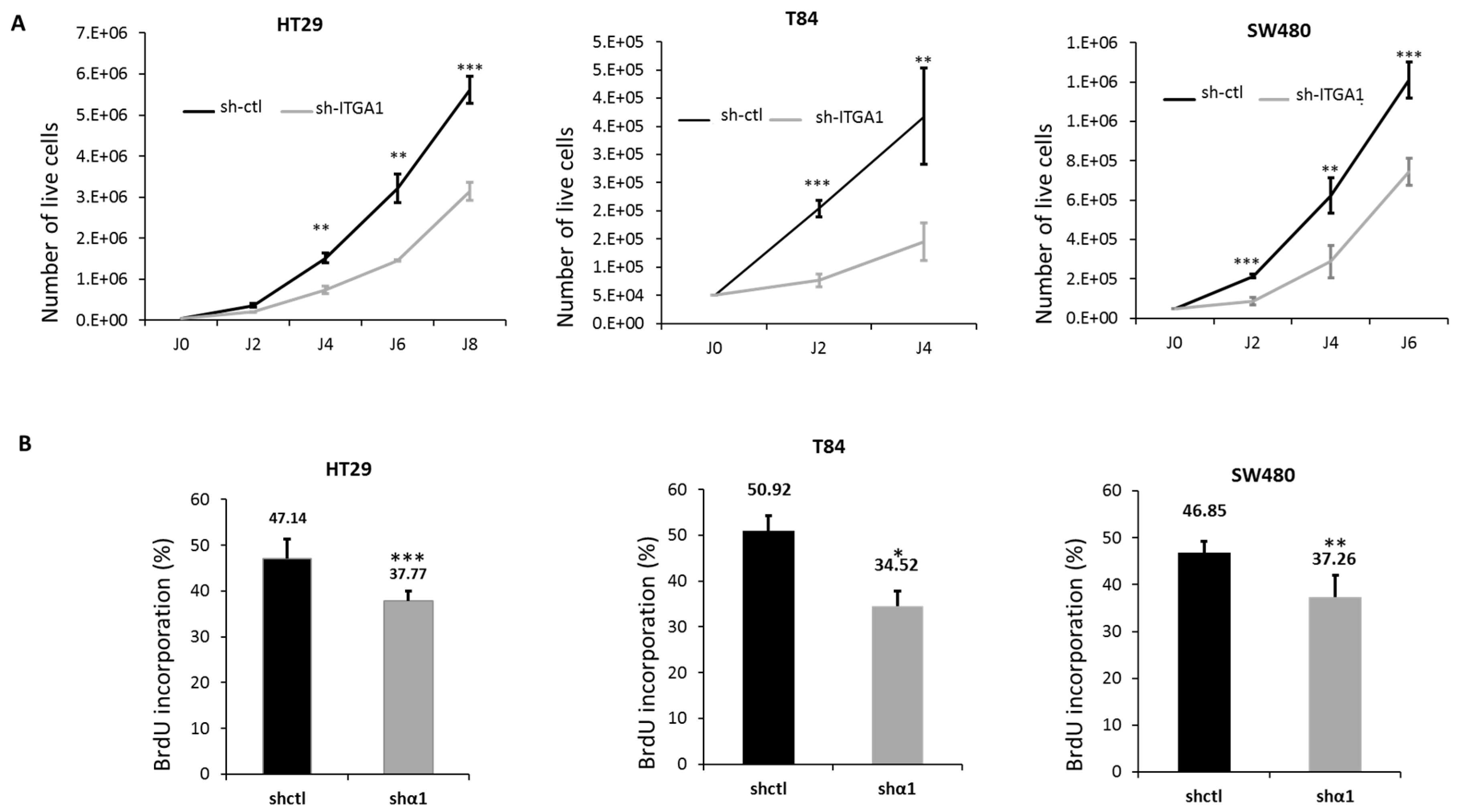
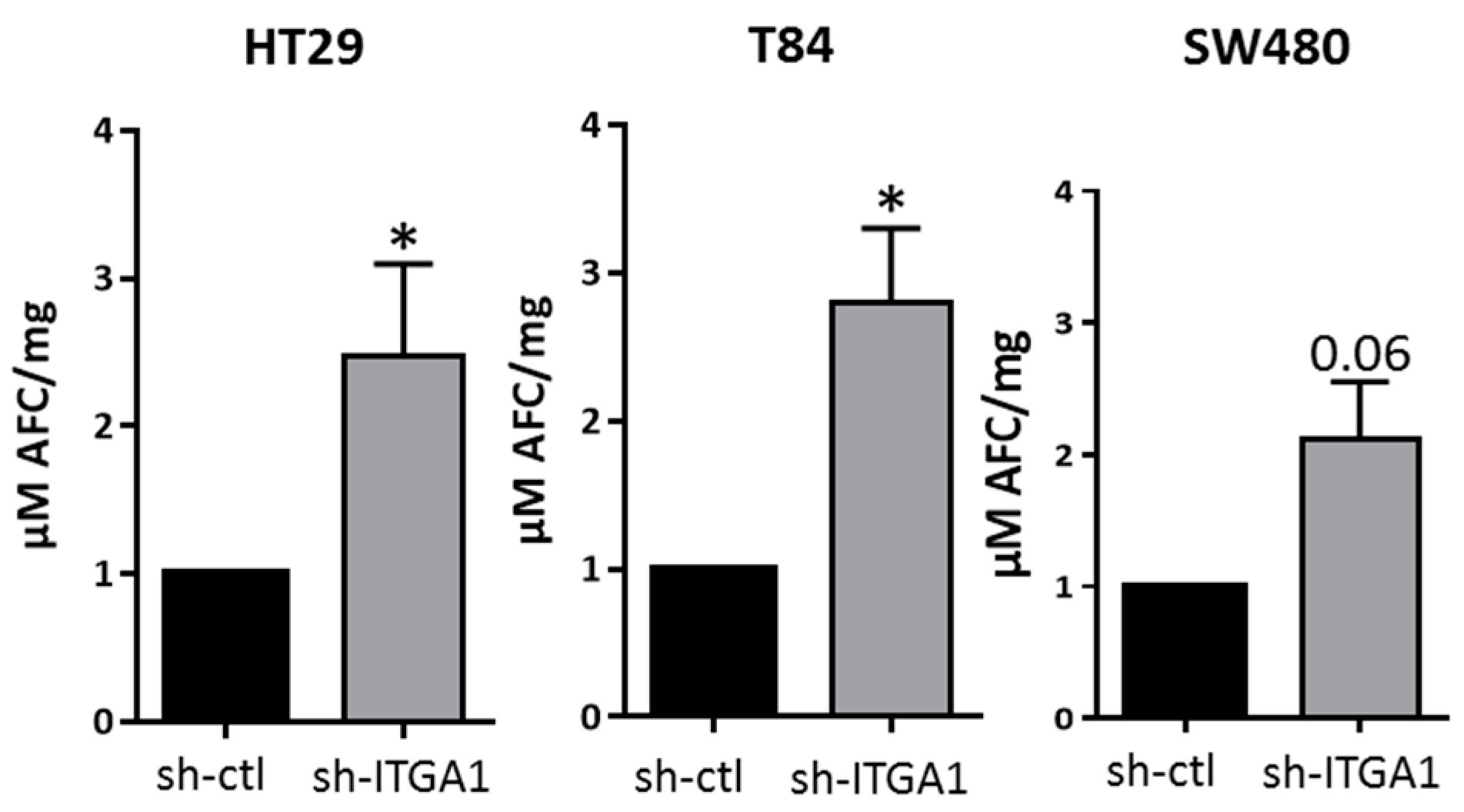
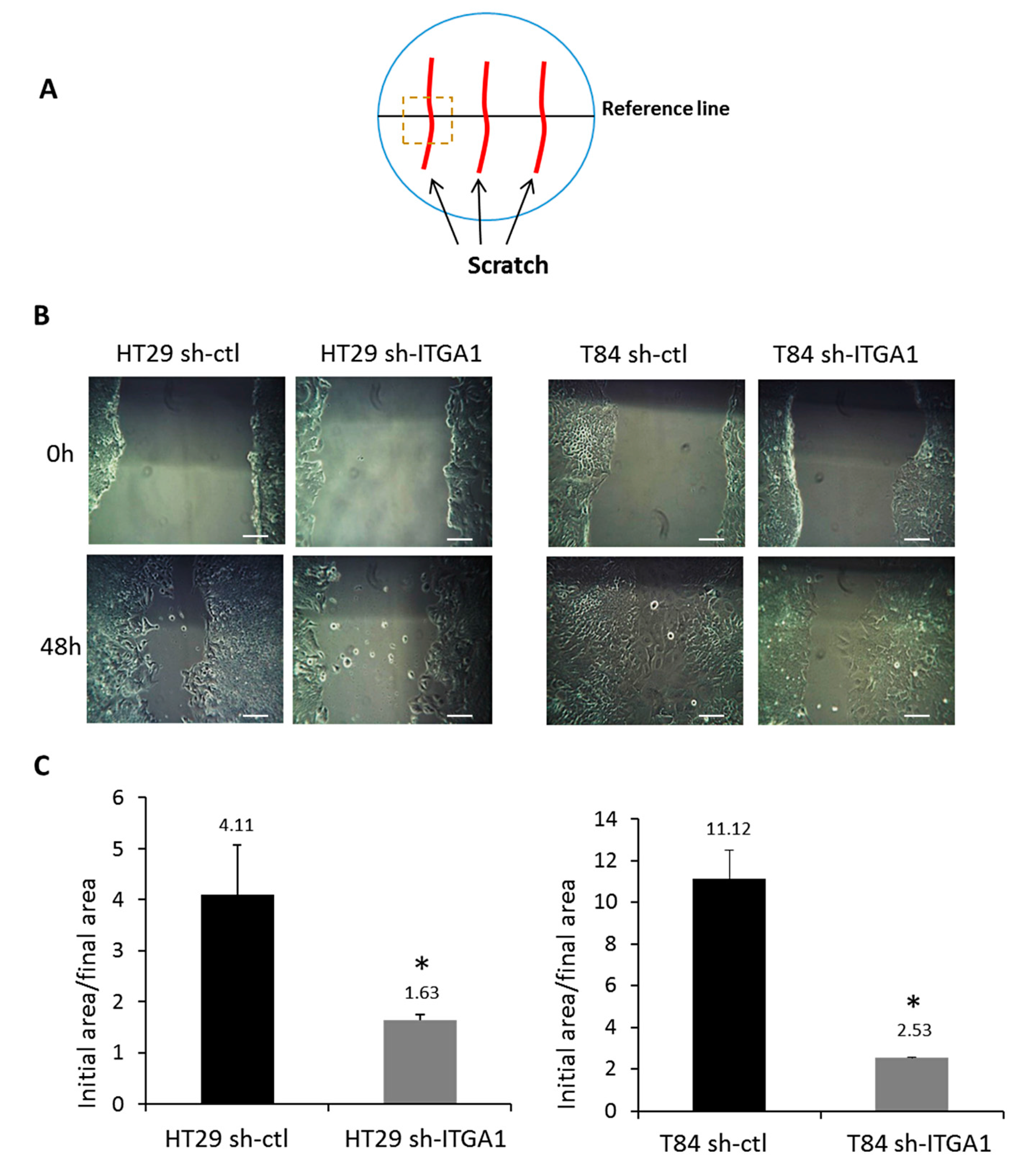
2.2. Integrin α1β1 Regulates Proliferation, Anoikis and Migration in CRC Cells
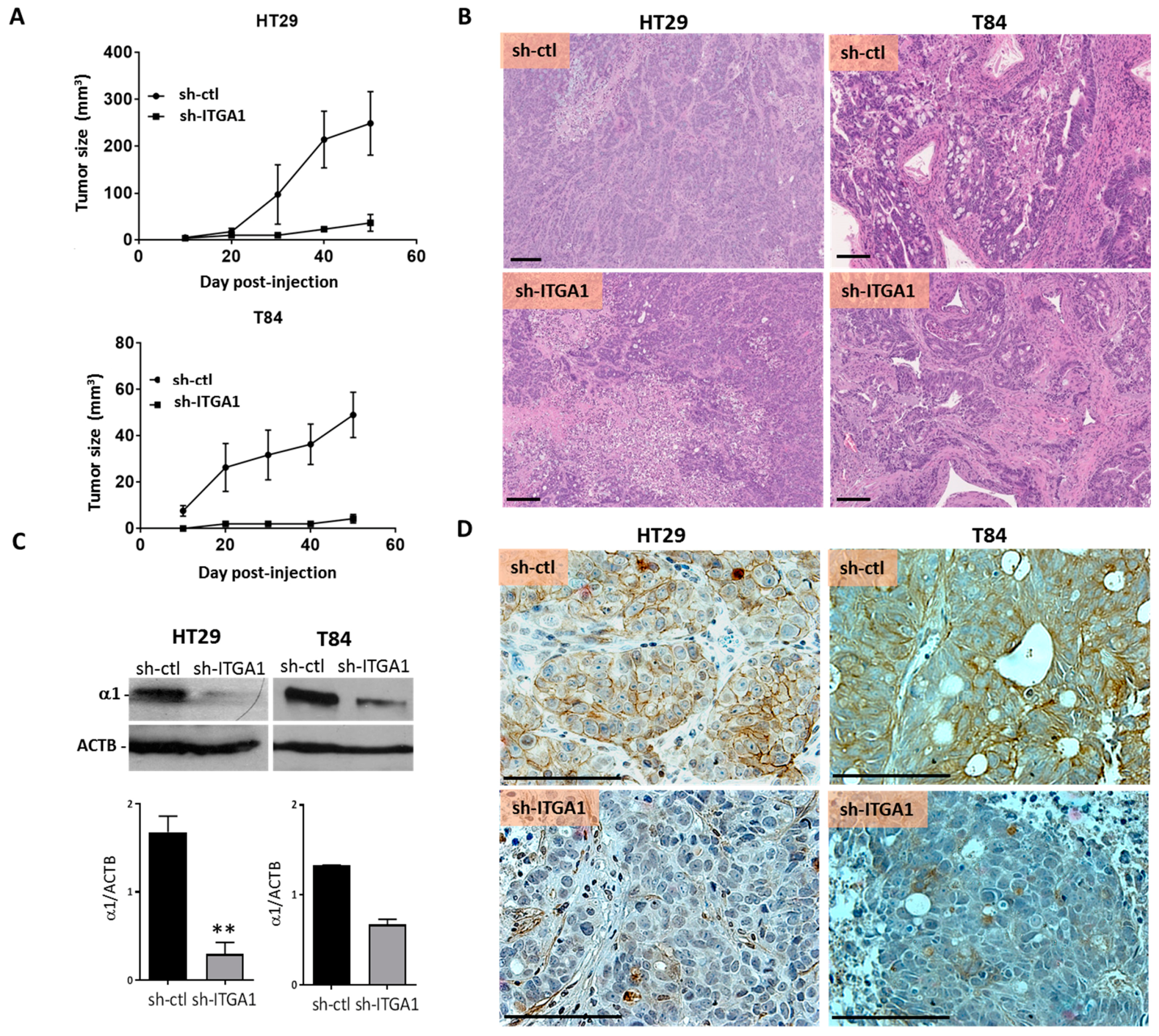
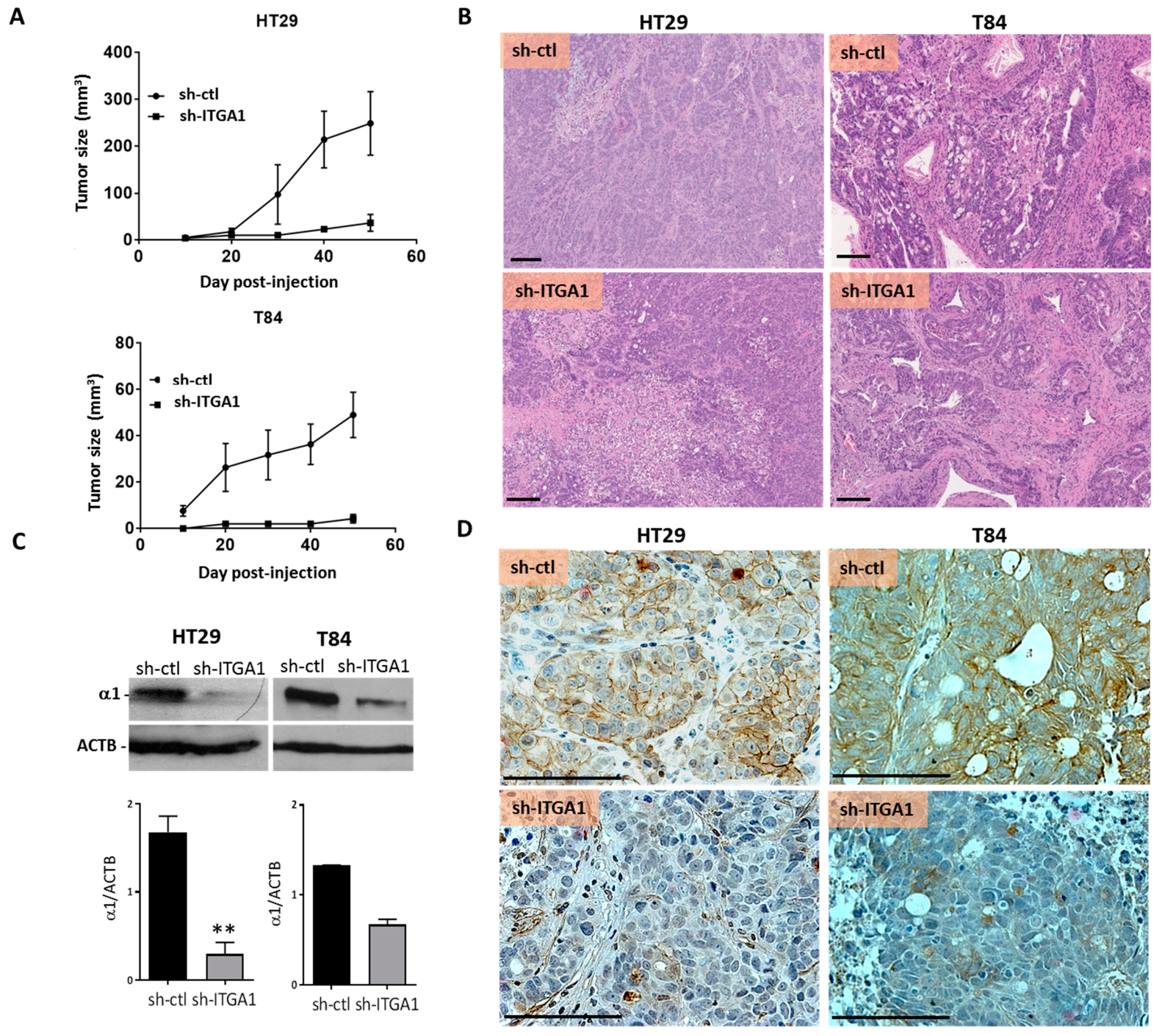
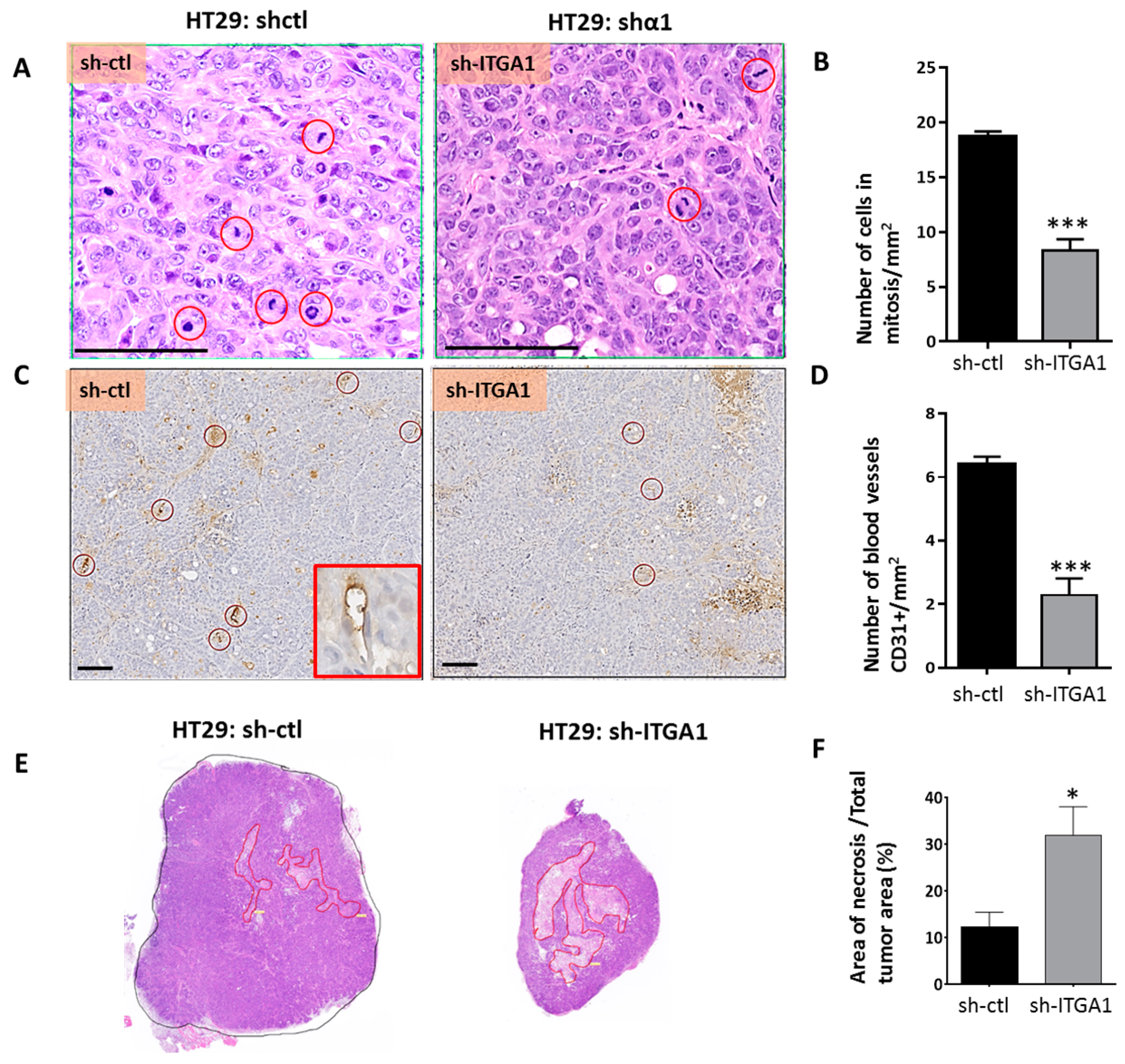
2.3. Integrin α1β1 Promotes Tumor Growth
3. Discussion
4. Materials and Methods
4.1. Cell Culture and Lentivirus-Mediated RNA Interference
4.2. Western Blot Analysis
4.3. Growth Curve Assay
4.4. Cell Proliferation Assay
4.5. Migration Assay
4.6. Anoikis Assays
4.7. Caspase 3 Activity Assays
4.8. Xenografts and Tumor Immunohistochemical Staining
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Giancotti, F.G. Integrin signaling: Specificity and control of cell survival and cell cycle progression. Curr. Opin. Cell Biol. 1997, 9, 691–700. [Google Scholar]
- Luo, B.H.; Carman, C.V.; Springer, T.A. Structural basis of integrin regulation and signaling. Annu. Rev. Immunol. 2007, 25, 619–647. [Google Scholar]
- Vachon, P.H. Integrin signaling, cell survival, and anoikis: Distinctions, differences, and differentiation. J. Signal Transduct. 2011, 2011, 738137. [Google Scholar] [CrossRef]
- Pozzi, A.; Wary, K.K.; Giancotti, F.G.; Gardner, H.A. Integrin alpha1beta1 mediates a unique collagen-dependent proliferation pathway in vivo. J. Cell Biol. 1998, 142, 587–594. [Google Scholar]
- Wary, K.K.; Mariotti, A.; Zurzolo, C.; Giancotti, F.G. A requirement for caveolin-1 and associated kinase fyn in integrin signaling and anchorage-dependent cell growth. Cell 1998, 94, 625–634. [Google Scholar]
- Abair, T.D.; Bulus, N.; Borza, C.; Sundaramoorthy, M.; Zent, R.; Pozzi, A. Functional analysis of the cytoplasmic domain of the integrin 1 subunit in endothelial cells. Blood 2008, 112, 3242–3254. [Google Scholar]
- Senger, D.R.; Perruzzi, C.A.; Streit, M.; Koteliansky, V.E.; de Fougerolles, A.R.; Detmar, M. The alpha(1)beta(1) and alpha(2)beta(1) integrins provide critical support for vascular endothelial growth factor signaling, endothelial cell migration, and tumor angiogenesis. Am. J. Pathol. 2002, 160, 195–204. [Google Scholar]
- Gardner, H. Integrin alpha1beta1. Adv. Exp. Med. Biol. 2014, 819, 21–39. [Google Scholar]
- Van Slambrouck, S.; Grijelmo, C.; De Wever, O.; Bruyneel, E.; Emami, S.; Gespach, C.; Steelant, W.F. Activation of the fak-src molecular scaffolds and p130cas-jnk signaling cascades by alpha1-integrins during colon cancer cell invasion. Int. J. Oncol. 2007, 31, 1501–1508. [Google Scholar]
- Ferlay, J.; Soerjomataram, I.; Dikshit, R.; Eser, S.; Mathers, C.; Rebelo, M.; Parkin, D.M.; Forman, D.; Bray, F. Cancer incidence and mortality worldwide: Sources, methods and major patterns in globocan 2012. Int. J. Cancer 2015, 136, E359–E386. [Google Scholar]
- Boudjadi, S.; Carrier, J.C.; Beaulieu, J.F. Integrin alpha1 subunit is up-regulated in colorectal cancer. Biomark. Res. 2013, 1, 16. [Google Scholar]
- Boudjadi, S.; Carrier, J.C.; Groulx, J.F.; Beaulieu, J.F. Integrin alpha1beta1 expression is controlled by c-myc in colorectal cancer cells. Oncogene 2016, 35, 1671–1678. [Google Scholar]
- Bartolome, R.A.; Barderas, R.; Torres, S.; Fernandez-Acenero, M.J.; Mendes, M.; Garcia-Foncillas, J.; Lopez-Lucendo, M.; Casal, J.I. Cadherin-17 interacts with alpha2beta1 integrin to regulate cell proliferation and adhesion in colorectal cancer cells causing liver metastasis. Oncogene 2014, 33, 1658–1669. [Google Scholar]
- Abair, T.D.; Sundaramoorthy, M.; Chen, D.; Heino, J.; Ivaska, J.; Hudson, B.G.; Sanders, C.R.; Pozzi, A.; Zent, R. Cross-talk between integrins alpha1beta1 and alpha2beta1 in renal epithelial cells. Exp. Cell Res. 2008, 314, 3593–3604. [Google Scholar]
- Giancotti, F.G. Complexity and specificity of integrin signalling. Nat. Cell Biol. 2000, 2, E13–E14. [Google Scholar]
- Macias-Perez, I.; Borza, C.; Chen, X.; Yan, X.; Ibanez, R.; Mernaugh, G.; Matrisian, L.M.; Zent, R.; Pozzi, A. Loss of integrin alpha1beta1 ameliorates kras-induced lung cancer. Cancer Res. 2008, 68, 6127–6135. [Google Scholar]
- Sterry, W.; Mielke, V.; Konter, U.; Kellner, I.; Boehncke, W.H. Role of beta 1-integrins in epidermotropism of malignant t cells. Am. J. Pathol. 1992, 141, 855–860. [Google Scholar]
- Gogali, A.; Charalabopoulos, K.; Constantopoulos, S. Integrin receptors in primary lung cancer. Exp. Oncol. 2004, 26, 106–110. [Google Scholar]
- Desgrosellier, J.S.; Cheresh, D.A. Integrins in cancer: Biological implications and therapeutic opportunities. Nat. Rev. Cancer 2010, 10, 9–22. [Google Scholar]
- Beausejour, M.; Thibodeau, S.; Demers, M.J.; Bouchard, V.; Gauthier, R.; Beaulieu, J.F.; Vachon, P.H. Suppression of anoikis in human intestinal epithelial cells: Differentiation state-selective roles of alpha2beta1, alpha3beta1, alpha5beta1, and alpha6beta4 integrins. BMC Cell Biol. 2013, 14, 53. [Google Scholar]
- Bian, B.; Mongrain, S.; Cagnol, S.; Langlois, M.J.; Boulanger, J.; Bernatchez, G.; Carrier, J.C.; Boudreau, F.; Rivard, N. Cathepsin b promotes colorectal tumorigenesis, cell invasion, and metastasis. Mol. Carcinog. 2016, 55, 671–687. [Google Scholar]
- Shattil, S.J.; Kim, C.; Ginsberg, M.H. The final steps of integrin activation: The end game. Nat. Rev. Mol. Cell Biol. 2010, 11, 288–300. [Google Scholar]
- Barczyk, M.; Carracedo, S.; Gullberg, D. Integrins. Cell Tissue Res. 2010, 339, 269–280. [Google Scholar]
- Beausejour, M.; Noel, D.; Thibodeau, S.; Bouchard, V.; Harnois, C.; Beaulieu, J.F.; Demers, M.J.; Vachon, P.H. Integrin/fak/src-mediated regulation of cell survival and anoikis in human intestinal epithelial crypt cells: Selective engagement and roles of pi3-k isoform complexes. Apoptosis 2012, 17, 566–578. [Google Scholar]
- Soung, Y.H.; Clifford, J.L.; Chung, J. Crosstalk between integrin and receptor tyrosine kinase signaling in breast carcinoma progression. BMB Rep. 2010, 43, 311–318. [Google Scholar]
- Uhlen, M.; Fagerberg, L.; Hallstrom, B.M.; Lindskog, C.; Oksvold, P.; Mardinoglu, A.; Sivertsson, A.; Kampf, C.; Sjostedt, E.; Asplund, A.; et al. Tissue-based map of the human proteome. Science 2015. [Google Scholar] [CrossRef]
- Pomerleau, V.; Landry, M.; Bernier, J.; Vachon, P.H.; Saucier, C. MET receptor-induced GRB2 or shc signals both promote transformation of intestinal epithelial cells, albeit they are required for distinct oncogenic functions. BMC Cancer 2014, 14, 1–15. [Google Scholar]
- Basu Roy, U.K.; Henkhaus, R.S.; Loupakis, F.; Cremolini, C.; Gerner, E.W.; Ignatenko, N.A. Caveolin-1 is a novel regulator of k-ras-dependent migration in colon carcinogenesis. Int. J. Cancer 2013, 133, 43–57. [Google Scholar]
- Salari, K.; Spulak, M.E.; Cuff, J.; Forster, A.D.; Giacomini, C.P.; Huang, S.; Ko, M.E.; Lin, A.Y.; van de Rijn, M.; Pollack, J.R. CDX2 is an amplified lineage-survival oncogene in colorectal cancer. Proc. Natl. Acad. Sci. USA 2012, 109, E3196–E3205. [Google Scholar]
- Roberts, P.J.; Der, C.J. Targeting the raf-mek-erk mitogen-activated protein kinase cascade for the treatment of cancer. Oncogene 2007, 26, 3291–3310. [Google Scholar]
- Montagut, C.; Settleman, J. Targeting the RAF-MEK-ERK pathway in cancer therapy. Cancer Lett. 2009, 283, 125–134. [Google Scholar]
- Di Magliano, M.P.; Logsdon, C.D. Roles for kRAS in pancreatic tumor development and progression. Gastroenterology 2013, 144, 1220–1229. [Google Scholar]
- Jun, J.E.; Rubio, I.; Roose, J.P. Regulation of RAS exchange factors and cellular localization of ras activation by lipid messengers in T cells. Front. Immunol. 2013, 4, 239. [Google Scholar] [CrossRef]
- Aplin, A.E.; Juliano, R.L. Integrin and cytoskeletal regulation of growth factor signaling to the map kinase pathway. J. Cell Sci. 1999, 112, 695–706. [Google Scholar]
- Aplin, A.E.; Stewart, S.A.; Assoian, R.K.; Juliano, R.L. Integrin-mediated adhesion regulates ERK nuclear translocation and phosphorylation of ALK-1. J. Cell Biol. 2001, 153, 273–282. [Google Scholar]
- Bouchard, V.; Demers, M.J.; Thibodeau, S.; Laquerre, V.; Fujita, N.; Tsuruo, T.; Beaulieu, J.F.; Gauthier, R.; Vezina, A.; Villeneuve, L.; et al. Fak/src signaling in human intestinal epithelial cell survival and anoikis: Differentiation state-specific uncoupling with the PI3-k/Akt-1 and MEK/ERK pathways. J. Cell Physiol. 2007, 212, 717–728. [Google Scholar]
- Demers, M.J.; Thibodeau, S.; Noel, D.; Fujita, N.; Tsuruo, T.; Gauthier, R.; Arguin, M.; Vachon, P.H. Intestinal epithelial cancer cell anoikis resistance: EGFR-mediated sustained activation of Src overrides fak-dependent signaling to mek/erk and/or PI3-k/Akt-1. J. Cell Biochem. 2009, 107. [Google Scholar] [CrossRef]
- Pignatelli, M.; Smith, M.E.; Bodmer, W.F. Low expression of collagen receptors in moderate and poorly differentiated colorectal adenocarcinomas. Br. J. Cancer 1990, 61, 636–638. [Google Scholar]
- Stallmach, A.; von Lampe, B.; Matthes, H.; Bornhoft, G.; Riecken, E.O. Diminished expression of integrin adhesion molecules on human colonic epithelial cells during the benign to malign tumour transformation. Gut 1992, 33, 342–346. [Google Scholar]
- Benoit, Y.D.; Larrivee, J.F.; Groulx, J.F.; Stankova, J.; Vachon, P.H.; Beaulieu, J.F. Integrin alpha8beta1 confers anoikis susceptibility to human intestinal epithelial crypt cells. Biochem. Biophys. Res. Commun. 2010, 399, 434–439. [Google Scholar]
- Klekotka, P.A.; Santoro, S.A.; Zutter, M.M. Alpha 2 integrin subunit cytoplasmic domain-dependent cellular migration requires p38 mapk. J. Biol. Chem. 2001, 276, 9503–9511. [Google Scholar]
- Vicente-Manzanares, M.; Choi, C.K.; Horwitz, A.R. Integrins in cell migration--the actin connection. J. Cell Sci. 2009, 122, 199–206. [Google Scholar]
- Becker, H.M.; Rullo, J.; Chen, M.; Ghazarian, M.; Bak, S.; Xiao, H.; Hay, J.B.; Cybulsky, M.I. Alpha1beta1 integrin-mediated adhesion inhibits macrophage exit from a peripheral inflammatory lesion. J. Immunol. 2013, 190, 4305–4314. [Google Scholar]
- Lochter, A.; Navre, M.; Werb, Z.; Bissell, M.J. Alpha1 and alpha2 integrins mediate invasive activity of mouse mammary carcinoma cells through regulation of stromelysin-1 expression. Mol. Biol. Cell 1999, 10, 271–282. [Google Scholar]
- Yang, C.; Zeisberg, M.; Lively, J.C.; Nyberg, P.; Afdhal, N.; Kalluri, R. Integrin alpha1beta1 and alpha2beta1 are the key regulators of hepatocarcinoma cell invasion across the fibrotic matrix microenvironment. Cancer Res. 2003, 63, 8312–8317. [Google Scholar]
- Bolte, G.; Knauss, M.; Metzdorf, I.; Stern, M. Dot blot chemiluminescence assay for studying food protein binding to small intestinal brush border membranes in vitro. J. Biochem. Biophys. Methods 1997, 34, 189–203. [Google Scholar]
- Marvin-Guy, L.F.; Duncan, P.; Wagniere, S.; Antille, N.; Porta, N.; Affolter, M.; Kussmann, M. Rapid identification of differentiation markers from whole epithelial cells by matrix-assisted laser desorption/ionisation time-of-flight mass spectrometry and statistical analysis. Rapid Commun. Mass Spectrom. 2008, 22, 1099–1108. [Google Scholar]
- Beaulieu, J.F. Differential expression of the vla family of integrins along the crypt-villus axis in the human small intestine. J. Cell Sci. 1992, 102, 427–436. [Google Scholar]
- Boudjadi, S.; Beaulieu, J.F. Integrin a1b1 regulates intestinal cell junctions. 2017; manuscript in preparation. [Google Scholar]
- Chen, N.; Zhou, Q. Constructing tissue microarrays without prefabricating recipient blocks: A novel approach. Am. J. Clin. Pathol. 2005, 124, 103–107. [Google Scholar]
- McGuire, T.F.; Sajithlal, G.B.; Lu, J.; Nicholls, R.D.; Prochownik, E.V. In vivo evolution of tumor-derived endothelial cells. PLoS ONE 2012, 7, e37138. [Google Scholar]
- Sajithlal, G.B.; McGuire, T.F.; Lu, J.; Beer-Stolz, D.; Prochownik, E.V. Endothelial-like cells derived directly from human tumor xenografts. Int. J. Cancer 2010, 127, 2268–2278. [Google Scholar]
- Boudjadi, S.; Beaulieu, J.F. Myc and integrins interplay in colorectal cancer. Oncoscience 2016, 3, 50–51. [Google Scholar]
- Dydensborg, A.B.; Teller, I.C.; Basora, N.; Groulx, J.F.; Auclair, J.; Francoeur, C.; Escaffit, F.; Pare, F.; Herring, E.; Menard, D.; et al. Differential expression of the integrins alpha6abeta4 and alpha6bbeta4 along the crypt-villus axis in the human small intestine. Histochem. Cell Biol. 2009, 131, 531–536. [Google Scholar]
- Dufour, G.; Demers, M.J.; Gagne, D.; Dydensborg, A.B.; Teller, I.C.; Bouchard, V.; Degongre, I.; Beaulieu, J.F.; Cheng, J.Q.; Fujita, N.; et al. Human intestinal epithelial cell survival and anoikis. Differentiation state-distinct regulation and roles of protein kinase b/akt isoforms. J. Biol. Chem. 2004, 279, 44113–44122. [Google Scholar]
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Boudjadi, S.; Bernatchez, G.; Sénicourt, B.; Beauséjour, M.; Vachon, P.H.; Carrier, J.C.; Beaulieu, J.-F. Involvement of the Integrin α1β1 in the Progression of Colorectal Cancer. Cancers 2017, 9, 96. https://doi.org/10.3390/cancers9080096
Boudjadi S, Bernatchez G, Sénicourt B, Beauséjour M, Vachon PH, Carrier JC, Beaulieu J-F. Involvement of the Integrin α1β1 in the Progression of Colorectal Cancer. Cancers. 2017; 9(8):96. https://doi.org/10.3390/cancers9080096
Chicago/Turabian StyleBoudjadi, Salah, Gérald Bernatchez, Blanche Sénicourt, Marco Beauséjour, Pierre H. Vachon, Julie C. Carrier, and Jean-François Beaulieu. 2017. "Involvement of the Integrin α1β1 in the Progression of Colorectal Cancer" Cancers 9, no. 8: 96. https://doi.org/10.3390/cancers9080096