Mixed Enzyme Systems for Delignification of Lignocellulosic Biomass

Department of Chemistry, Biochemistry, & Physics, Marist College, Poughkeepsie, NY 12601, USA
Catalysts 2014, 4(1), 1-35; https://doi.org/10.3390/catal4010001
Submission received: 9 October 2013
/
Revised: 13 December 2013
/
Accepted: 18 December 2013
/
Published: 6 January 2014
Abstract
:The application of enzymes such as laccase and xylanase for the preparation of cellulose from lignocellulosic material is an option for those industries seeking to reduce the use of chlorine-containing bleach agents, thus minimizing the environmental impact of their processes. Mixed hydrolytic and oxidative enzyme systems have been well described in the context of biopulping, and thus provide good precedent regarding effectiveness, despite the susceptibility of xylanase to inactivation by laccase-generated oxidants. This paper examines the progress towards development of sequential and simultaneous mixed enzyme systems to accomplish delignification.
Keywords:
biomass; delignification; hemicellulose; laccase; lignin; lignocellulose; manganese peroxidase; mediator; xylanase1. Introduction
The processing of lignocellulosic biomass is essential for the production of cellulosic pulp as a raw material for paper manufacture and also as a feedstock for chemical synthesis, including biofuel. In nature, lignocellulose degradation is a multienzyme process involving both hydrolytic and oxidative transformations, due to the complexity of lignified plant material [1,2,3]. The enzyme systems must be robust in order to recycle the vast amount of carbon in terrestrial ecosystems [1,4]. The discovery of peroxide-dependent lignin degrading enzymes in cultures of the white rot fungus Phanerochaete chrysosporium [5,6] marked the beginning of the development of isolated enzyme systems for applied biomass delignification.
1.1. Precedent
One of the earliest studies of a mixed enzyme treatment applied to cellulosic pulp was by Poppius-Levlin et al. [7] who enhanced the laccase-mediator treatment of pulp by a pretreatment with xylanase. Further studies documented the benefit of xylanase application to pulp followed by delignification using laccase in combination with a redox mediator such as HBT or NHA [8,9,10]. Successive application of the two enzyme systems, notably xylanase and the LMS, resulted in improved delignification, to the extent removal of 60% of the residual lignin in softwood kraft pulp and approaching 70% in wheat straw chemical pulp [9,11,12,13,14].
1.2. Impact
The US consumes 140 billion gallons per year of gasoline. According to the Energy Independence and Security Act of 2007, the Renewable Fuel Standard calls for 36 billion gallons per year of renewable biofuel by 2022, including 22 billion gallons of cellulosic biofuel and other advanced biofuels. The US EPA had targeted 250 million gallons of cellulosic biofuel in 2010, but achieved only 10% of that goal (25.5 million gallons). This can be attributed to the difficulty in saccharifying cellulose isolated from lignocellulosic substrates despite its relative abundance in a variety of materials, as detailed in Table 1. The International Energy Agency estimates that a targeted 50% reduction of greenhouse gases by 2050 will require a four-fold increase of biomass-derived energy to 150 EJ/year (where 1 EJ = 1018 J), representing over 20% of the world energy supply [15].
1.3. Objective
The objective is ultimately to develop a simultaneous delignification system to maximize the green production of cellulose for subsequent use in papermaking or as a chemical synthesis feedstock. The hypothesis considered in this review paper is that the simultaneous action of hydrolytic and oxidative enzymes may be a promising strategy for pulp delignification and more generally for biomass processing. Xylanase and laccase are readily commercially available and thus are the primary focus of this discussion. Xylanase exposes lignin which can be degraded by laccase with a mediator [16]. The simultaneous application of the two classes of enzymes, in the context of pulp delignification, could be incorporated into most bleach plants without significant capital investment. In general, biomass processing using a mixed enzyme system for cellulose isolation would be economically beneficial given opportunity to use agricultural lignocellulosic waste as substrate [17]. This paper first introduces each of the two classes of enzymes, and then shows how the enzymes work in tandem in nature, from the perspective of biopulping. A review of the results of mixed enzyme systems applied to pulp, both sequential and simultaneous, is provided.

{kind=link}
{kind=link}
{kind=link}
| Lignocellulosic material | Cellulose (%) | Hemicellulose (%) | Lignin (%) |
|---|---|---|---|
| Hardwood Stems | 40–55 | 24–40 | 18–25 |
| Softwood Stems | 45–50 | 25–35 | 25–35 |
| Corn Cobs | 45 | 35 | 15 |
| Grasses | 25–40 | 35–50 | 10–30 |
| Paper | 85–99 | 0 | 0–15 |
| Wheat Straw | 30 | 50 | 15 |
| Leaves | 15–20 | 80–85 | 0 |
| Newspaper | 40–55 | 25–40 | 18–30 |
| Switchgrass | 45 | 31 | 12 |
2. Oxidative Enzymes for Lignin Removal
In nature, three key enzymes are broadly recognized for their ability to oxidize lignin: LiP, MnP, and laccase. Of these three enzymes, laccase exhibits the greatest potential for commercial development for several reasons. First, unlike LiP and MnP, laccase does not require the presence of a heme cofactor. Second, laccase does not require a low, steady supply of hydrogen peroxide for activity; oxygen serves as the terminal electron acceptor for this enzyme. This section introduces both laccase and MnP, motivated by accounts of synergy in lignin oxidation derived from the combination of xylanase with these oxidative enzymes [20,21,22].
2.1. Laccase and the Mediator System
Laccase (benzenediol:oxygen oxidoreductase, EC 1.10.3.2) is a blue copper enzyme with broad substrate specificity that catalyzes the oxidation of phenols, anilines, and aromatic thiols [23], with the concomitant four electron reduction of O2 to H2O. The enzyme occurs in fungi, plants, trees, insects, and bacteria [24]. Laccase is recognized as an industrially significant enzyme in pulp and textile manufacture, beverage and food processing, and pollutant detoxification [25].
Laccase contains four active site copper atoms, one at the T1 site and three at the T2/T3 site, arranged as a trinuclear copper cluster; the three different types of copper, T1, T2, and T3, are classified according to the EPR spectrum of each copper atom [26]. Oxygen reduction occurs at the T2/T3 site, while the electrochemical potential, typically ranging from 0.4 to 0.8 V, is determined at the T1 site [27,28]. Laccases have relatively low redox potentials (≤ 0.8 V) relative to ligninolytic peroxidases (>1 V) [25].
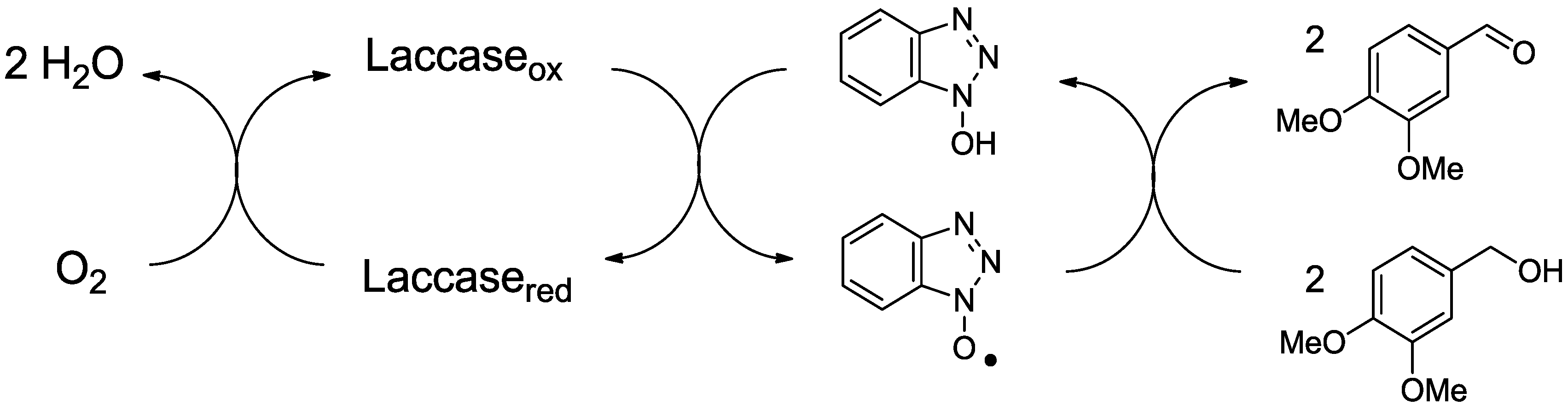
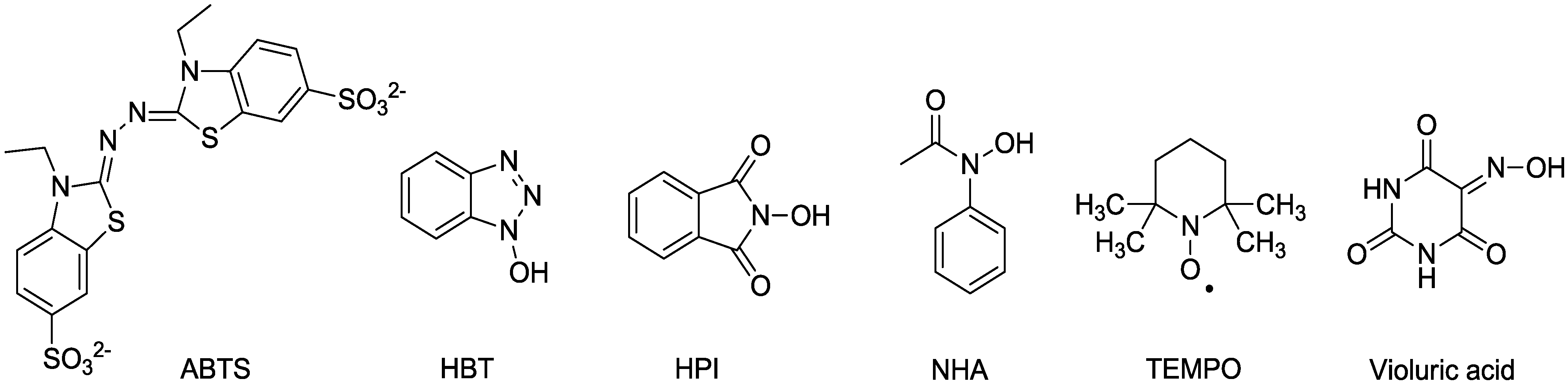
Laccase was long considered to effect the degradation of only phenolic lignin and phenolic lignin model compounds [29], since nonphenolic substrates with redox potentials greater than 1.3 V could not be oxidized by laccase directly. Given that phenolic lignin structures account for 10–20% of the lignin substructures in the plant cell wall, laccase was therefore not expected to play a role as a delignifying agent [30,31]. In 1989, however, the combination of laccase and syringaldehyde was shown to oxidize the nonphenolic lignin model compound veratryl alcohol [32]. This account was followed soon after by a report that laccase could degrade nonphenolic lignin model compounds when used in conjunction with the redox mediator ABTS [33]. Laccase alone cannot oxidize veratryl alcohol to veratraldehyde, but laccase with a mediator can do so (Figure 1). The system cleaves or oxidizes nonphenolic lignin model dimers at the alpha carbon [34]. Subsequently, laccase plus ABTS was reported to delignify kraft pulp [35]. The combination of laccase plus a low molecular weight redox mediator is known as the laccase mediator system (LMS). Mediators include ABTS, HBT, HPI, NHA, TEMPO, and violuric acid (Figure 2), with HBT and NHA being particularly effective [36,37,38,39].
Figure 1.
HBT-mediated oxidation of veratryl alcohol to veratraldehyde by laccase. Adapted with permission from Bendl et al. [40]. Copyright 2008 Elsevier.
Figure 1.
HBT-mediated oxidation of veratryl alcohol to veratraldehyde by laccase. Adapted with permission from Bendl et al. [40]. Copyright 2008 Elsevier.

Figure 2.
Laccase redox mediators.

Oxidation of nonphenolic lignin may occur via either electron transfer, as with ABTS, or hydrogen atom transfer, as with the hydroxamic acids [41]. The best mediators for removal of recalcitrant nonphenolic lignin are of the N-OH variety, including HBT, HPI, NHA, and violuric acid, since the oxidation gives nitroxyl radicals. Nitroxyl radicals oxidize the target substrate by hydrogen atom transfer mechanism, and the driving force for the oxidation of substrate is the energy difference between the dissociated bond (C-H) in the target substrate and the forming bond (NO-H) in the mediator [25,42,43]. Oxidation of the stable nitroxyl radical TEMPO by high redox potential laccases occurs via a non-radical, ionic mechanism to the oxoammonium ion (>N=O+) [25,41]. Consistent with the difference in oxidation mechanism as a function of mediator type, analysis of residual lignin from laccase-HBT treatment versus laccase-NHA treatment via NMR showed greater enrichment of carboxylic acid functionality in laccase-HBT lignin than in laccase-NHA lignin [44].
The LMS is effective for both wood and non-wood (sisal) pulp bleaching and delignification, yielding pulp with properties equal or better than can be achieved using only chemical reagents [25,38,45,46,47]. The optimum oxygen pressure required for the LMS is relatively low and can be generated by the hydraulic pressure in a conventional bleaching tower [34]. The LMS can replace either an ozone stage or an oxygen delignification stage in kraft pulp bleaching [7,48]. The LMS using HBT as mediator followed by an alkaline extraction can reach as much as 70% delignification, and if the extraction is followed by a second LMS treatment, the delignification can be extended to 80%, indicating that the LMS with HBT as mediator could reach even very unreactive lignin [36]. The kappa number is best measured following an alkaline extraction, otherwise a modified, alkaline extractable lignin is left behind after the laccase stage that interferes with kappa [49,50].
Limitations of the LMS are primarily a consequence of the mediator. While the LMS delignification scheme predicts that the mediator should be continuously recycled (Figure 1), HBT, a particularly potent mediator, undergoes ineffective conversion to benzotriazole. Concern has also been expressed regarding the cost of HBT and the toxicity of HBT and its by-products [25,51,52,53], although the toxicity of laccase-HBT treatments has been shown to fall in non-hazardous limits [51]. Another drawback to the LMS if the objective is to isolate cellulose for papermaking is that the system produces hydroxyl radicals, which can lead to cellulose oxidation and consequent decrease in the degree of polymerization, manifested as viscosity decrease upon alkaline hydrolysis. Cellulose oxidation can be minimized by following LMS with a reducing stage [45,46,54].
Disadvantageously, the laccase-generated HBT oxidant is reactive enough to lead to laccase oxidation and inactivation. For example, in a treatment where HBT was applied at a dose of 1% on pulp, only 2% residual laccase was detected at the end of the laccase stage [34]. This is difficult to moderate since the minimum effective HBT dose is 0.5% [55]. Further, a slower rate of O2 uptake was observed with HBT than with a mediator like ABTS, another indication of loss of laccase activity. However, some of the activity loss may be attributed to laccase adsorption to fiber. For example, in a study using Pycnoporus cinnabarinus laccase to bleach flax pulp, laccase adsorption to the pulp was estimated to be around 20%, based upon residual enzyme activity in the absence of mediator [1].
Given the high cost of HBT and the tendency of the HBT oxidant to inactivate laccase, other mediators, both synthetic and natural, have been sought [39,52,53,56,57,58,59,60,61,62]. For example, the synthetic mediator NHA has been described as both cost effective and biodegradable, with another particular advantage that laccase retains about 80% of its initial activity in an LMS stage with this mediator [63]. Other synthetic hydroxamic acids such as N-(4-cyanophenyl)-acetohydroxamic acid and 7-cyano-4-hydroxy-2H-1,4-benzoxazine-3-one have been reported to be as effective as NHA for bleaching of softwood kraft pulp [64]. Naturally occurring phenolic mediators such as syringaldehyde and acetosyrignone can provide up to 25% lignin removal [52]. Phenolic p-hydroxycinnamic acid derivatives have also been considered for nonwood pulp treatment, and while less laccase inactivation was observed, pulp bleaching was less efficient than with HBT. Phenolic compounds tend to bind to pulp fibers [59], according to results previously reported for p-coumaric acid [52]. Overall, the ideal mediator would be a small molecule that is oxidizable to a stable radical that would not inactivate the enzyme. In addition, laccase mediators should be environmentally benign, available at low cost, and would be needed in catalytic rather than stoichiometric quantities due to recycling without degeneration [25].
In a study of the mechanism of oxidation of the three N-OH compounds HBT, NHA, and violuric acid with seven different laccases, a bell-shaped pH activity profile was observed with optimum activity over the range of 4–7. The oxidation rate was found to be dependent on the redox potential difference between laccase and the N-OH substrate [65]. A correlation between enzyme redox potential and activity towards substrate had previously been reported [66]. The use of a laccase with a higher redox potential or an N-OH compound with a lower redox potential produced a higher rate of oxidation.
When laccase from Trametes versicolor was used as a component of the LMS, delignification was accomplished over a broad range of acidic pH [67]. Ultimately the optimum pH for delignification may depend on the mediator, since the pH optimum for substrate oxidation in the absence of pulp varies. For example, the pH optimum was determined to be 5.0 with o-dianisidine as substrate and 3.0 with ABTS as substrate. Laccases generally have a low pH optimum with ABTS as substrate [68].
Organisms such as Ceriporiopsis subvermispora produce laccase isozymes of variable pH optimum activity and stability; the C. subvermispora laccases are like those of T. versicolor in that they oxidize only the methoxybenzene congener with the lowest redox potential, TMB. While oxidation of ABTS and TMB was found to be optimal at pH 2-3 for both the L1 and L2 isozymes, L2 exhibited a broader range of activity as a function of pH. L1 was barely active with respect to ABTS oxidation at pH 5, whereas L2 still exhibited 60% of activity for ABTS oxidation relative to that at pH 3. For guaiacol oxidation, the maximum L2 activity was at pH 5, versus maximum L1 activity at pH 3. The pH stability followed the same trend, with L1 and L2 maximally stable at pH 5 and pH 3-4, respectively. The key point is that since the pH during the degradation of lignin in wood by C. subvermispora gradually decreases from 5.0 to near 3.5, the different pH optima of L1 and L2 ensures that oxidation can occur over a wider range of pH [69].
2.2. MnP
MnP (Mn(II):hydrogen-peroxide oxidoreductase, EC 1.11.1.13), first characterized in 1984, is another key oxidative enzyme for lignin degradation [70]. MnP oxidizes Mn2+ to Mn3+ in a reaction that is dependent on H2O2; the manganese ion is chelated to an organic acid. Mn3+ is a strong oxidant (1.54 V) and can act on a variety of phenolic compounds as a diffusible oxidant through the lignin matrix [17]. The MnP system can also oxidize nonphenolics, including the model compounds veratryl alcohol and benzyl alcohol. The redox potential of chelated Mn3+ depends on the chelator [71].
MnP effectively reduces the kappa number of chemical pulp produced via the kraft process, but the treated pulp also exhibits increased bleachability, especially in subsequent peroxide stages [72]. This indicates that MnP not only removes lignin, but the enzyme also oxidizes residual lignin to a form that is more easily bleached by alkaline peroxide [34]. This is perhaps due to mobilization and removal of manganese ions in wood. In a study of the biobleaching of kraft pulp by white rot fungi during solid-state fermentation, a direct relationship was observed between cumulative MnP activity and brightness [73].
A disadvantage with MnP use is that residual Mn2+ ion can impact brightness development in subsequent stages, specifically, high manganese concentrations can reduce the effectiveness of the peroxide stage [74], and it may also produce darkening of pulps at high temperature and pH values [75]. Another concern is the susceptibility of inactivation of MnP by peroxide. Generally, fungal peroxidases are inactivated by peroxide concentrations greater than 0.10 mM [72,76]. H2O2 is best added in pulses, for example, as aliquots of 0.042 mM H2O2 every 5 min to a total charge of 0.50 mM over 1 h [17].
Manganese ion chelators include organic acids such as oxalic and malonic. In systems where glucose oxidase and glucose are used to generate H2O2 for MnP, the resulting gluconic acid can chelate manganese ion [17]. A Mn2+ concentration of 33 μM was found to be optimal in a study of the bleaching of a eucalyptus kraft pulp with oxalic acid as chelator [17], although in a previous study using malonic acid as a chelator, 100–500 μM Mn2+ was found to be optimal [77]. Unsaturated fatty acids such of linoleic acid, linolenic acid, and Tween 80 can assist the MnP system by enhancing brightness [17,34]; linoleic and linolenic acids are more effective than Tween 80. Mechanistically, Tween 80 was proposed to facilitate oxidation of a nonphenolic β-O-4 lignin model compound via peroxyl radical [78]. Since white rot fungi contain lipids within their extracellular membranes, these lipids could play important an role in the fungal degradation of lignin [78].
3. Hydrolytic Enzymes for Lignin Removal
3.1. Xylanase
Xylan is the most abundant hemicellulose in the biosphere [79], accounting for one-third of the polysaccharide component of renewable plant biomass [80,81]. Xylan occurs as a heteropolysaccharide with a backbone of 1,4-linked xylose monomers that may be branched with arabinose, glucuronic acid, and acetate substituents at the 2- and 3-position of xylose [79]. This polysaccharide can be used by bacteria and fungi as a primary carbon source [82].
Hemicellulase systems are complex and variable from organism to organism [83] and potentially include endo-1,4-β-d-xylanase (1,4-β-d-xylan xylanohydrolase, EC 3.2.1.8), cleaving internal bonds in xylan chains, β‑xylosidase (EC 3.2.1.37), cleaving xylose units from xylooligosaccharides, endo-1,4-β-d-mannanase (EC 3.2.1.78), cleaving internal bonds in mannan chains, and β-mannosidase (EC 3.2.1.25), cleaving mannose units from mannooligosaccharides. Of these hemicellulases, xylanase has industrial significance as a bleaching agent in the pulp, paper, and textile industries, as an aid for improving the digestibility of animal feed, as an aid to improve dough processing for breadmaking, and also has application in bioethanol fuel production [84].
Xylanase has long been recognized as an effective bleaching pretreatment, with the first report of xylanase aided bleaching dating back to 1986 [24,85]; pretreatment with xylanase was shown to increase bleachability in subsequent steps, giving up to 25% bleach savings to reach target brightness [86,87]. As of 2007, according to Viikari et al. [88], about 20 mills in North America and Scandinavia used enzymes in kraft pulp bleaching, and the approximate price of xylanase treatment in that year was less than $2 per ton of pulp. Enzyme products are available that function at both elevated pH and temperature, e.g., approaching pH 10 and 90–100 °C. There is also continued interest in the development of new xylanases and product formulations to enhance their compatibility for industrial applications [84].
Xylanases are widely used in the pulp and paper industry since they increase lignin extractability from kraft pulp by depolymerizing the hemicellulosic material closely associated with lignin in the plant cell wall [31,89]; this includes hydrolysis of precipitated xylan or hydrolysis of the xylan component of LCC in fibers. Consequently, a key reason for the improvement in the bleach response of pulps treated with xylanase is an increase in fiber permeability [87]. Xylanase acts as a bleaching aid rather than as a true delignification agent [24], since the enzyme does not directly degrade lignin. Another proposed mechanism of xylanase action involves removal of hexenuronic acid residues, which consume bleach chemicals and contribute to kappa number, the latter by consuming permanganate [8].
3.2. Aspects of Xylanase Structure Relative to Catalysis and Substrate Binding
Xylanases are distributed among two families of glycosyl hydrolases, family 10 and family 11 (formerly family F and family G); classification is determined by hydrophobic cluster analysis and sequence alignment [79,90,91]. Family 10 xylanases generally have a high molecular mass (>30 kDa) and low pI value (pH < 6.5) while family 11 xylanases generally have a low molecular mass (<30 kDa) and high pI value (8.0–10.0), although there are some low pI value family 11 xylanases of fungal origin [92,93,94]. There are also occurrences of family 5 and family 8 bacterial xylanases [94,95]. The broad range of xylanase activities and properties has been proposed to reflect the diversity of xylan structure [94]. Table 2 lists representative xylanases as a function of family; entries include those enzymes for which the structure has been reported or the enzyme has been characterized with respect to modification by NBS. A comprehensive review of xylanase and xylanase families has been prepared by Collins et al. [95].
All xylanases are known to catalyze hydrolysis via a double displacement mechanism, resulting in retention of configuration at the anomeric carbon [82,91,96]. The residues that are directly active in xylan hydrolysis are highly conserved in all family 11 xylanases, and include Glu78 (referring to the Bacillus subtilis sequence, equivalent to Thermomyces lanuginosus Glu86) as the active site nucleophile and Glu172 (T. lanuginosus Glu178) as the acid-base catalyst [79,97]. A very highly conserved arginine residue, Arg112 (T. lanuginosus Arg122), helps stabilize the negative charge on Glu78 [79,84,98]. Other conserved xylanase residues include six clustered tyrosine residues (Bacillus Tyr65, Tyr69, Tyr79, Tyr80, Tyr166, and Tyr174) and several tryptophan residues (Bacillus Trp9, Trp30, Trp71, Trp 85, Trp129, and Trp153) [84,98]. The two catalytic Glu residues are situated 6–7 Å apart in the Bacillus circulans [99] and Bacillus agaradhaerens [100] xylanases. T. lanuginosus Glu86 and Glu178 are solvent inaccessible, but Glu86 is hydrogen bonded to Gln136, Tyr77, and Tyr88, while Glu178 is hydrogen bonded only to Asn44. T. lanuginosus Glu86 has a typical pKa value of 3.9‑4.3, appropriate since this residue needs to be ionized to function as a nucleophile, while Glu178 has a pKa of 9.4.
Commercial xylanases for pulp bleaching are most often from family 11. This may attributed to the exclusive specificity of this family of xylanases for substrates containing d-xylose; in contrast, the family 5 xylanases possess carboxymethylcellulase activity while the family 10 xylanases are active on cellulose of low molecular mass [95]. Family 11 xylanases are comprised of a single domain with two antiparallel β sheets and one α helix [79,101]. Overall, the protein resembles a right hand, where the two β sheets form the fingers, and the combination of the twisted part of one of these sheets plus the sole α helix comprise the palm. Two loop regions form the thumb and the cord. A long substrate-binding cleft spans one entire side of the enzyme. Subsites for substrate docking are represented as ‑2, ‑1, +1, and +2, where cleavage occurs between subsites −1 and +1, and the negative numbers refer to residues in the nonreducing end direction, while positive numbers refer to residues in the reducing end direction [102].
| Xylanase † | Family | Mass (Da) ‡ | pI ‡ | Entry | Reference |
|---|---|---|---|---|---|
| Aspergillus niger Xylanase C | 10 | 35,486 | 6.17 | XYNC_ASPNC | [96] |
| Cellulomonas fimi CEX/Xylanase B | 10 | 51,219 | 6.19 | GUX_CELFI | [79,90] |
| Clostridium thermocellum Xylanase Z | 10 | 92,263 | 5.56 | XYNZ_CLOTH | [79,90] |
| Pseudomonas fluorescens Xylanase A | 10 | 64,804 | 6.08 | XYNA_CELJU | [79,90] |
| Pseudomonas fluorescens Xylanase B | 10 | 64,363 | 6.37 | XYNB_CELJU | [79,90] |
| Streptomyces lividans Xylanase A | 10 | 51,163 | 6.18 | XYNA_STRLI | [79,91] |
| Thermoascus aurantiacus Xylanase | 10 | 35,686 | 5.82 | XYNA_THEAU | [90] |
| Aspergillus kawachii Xylanase 3 | 11 | 22,627 | 4.07 | XYN3_ASPKW | [96] |
| Bacillus agaradhaerens Xylanase | 11 | 23,152 | 8.58 | Q7SIE3_BACAG | [96,103] |
| Bacillus circulans Xylanase | 11 | 23,359 | 9.44 | XYNA_BACCI | [79,90,96] |
| Bacillus pumilus Xylanase A | 11 | 25,491 | 9.33 | XYNA_BACPU | [90,96] |
| Bacillus subtilis Xylanase A | 11 | 23,345 | 9.44 | XYNA_BACSU | [90] |
| Clostridium saccharobutylicum Xylanase | 11 | 29,033 | 9.24 | XYNA_CLOSA | [90] |
| Fibrobacter succinogenes Xylanase C | 11 | 66,403 | 7.96 | XYNC_FIBSS | [96] |
| Penicillium citrinium Xylanase | 11 | 23,480 | 4.69 | Q2PGY1 | [104] |
| Thermomyces lanuginosus Xylanase | 11 | 24,356 | 4.77 | XYNA_THELA | [79,96] |
| Trichoderma harzianum Xylanase | 11 | 20,703 | 8.71 | XYN_TRIHA | [79] |
| Trichoderma reesei Xylanase I | 11 | 24,583 | 5.00 | XYN1_HYPJE | [79,96] |
| Trichoderma reesei Xylanase II | 11 | 24,172 | 8.89 | XYN2_HYPJE | [79,96] |
†Entries include those enzymes for which the structure has been reported or the enzyme has been characterized with respect to modification by NBS; ‡values are calculated for the complete sequence and do not consider protein processing.
With respect to binding of substrate by xylanase, arginine, tryptophan, and tyrosine residues may be involved via either stacking interactions or hydrogen bonding with the xylose residues [82,97,99,100,103,105,106,107,108]. Stacking interactions between carbohydrate and aromatic residues are common in glycosidases [109,110]. The interaction between substrate and a tryptophan residue, for example, is proposed to stabilize the transition state for glycosidic bond hydrolysis [111]; a tryptophan is essential for binding of the enzyme to substrate in order to carry out hydrolysis [80,112]. Modeling studies with xyloheptose in the T. lanuginosus active site show that Trp18 is involved in a stacking interaction with the sugar at subsite −2. X‑ray crystallographic analysis of inactivated B. circulans Xylanase A (XynA) co-crystallized with xylotetraose revealed a large number of interactions of carbohydrate with aromatic residues [99], including a stacking interaction between Trp9 and one of the xylose residues. The analogous Trp18 of Trichoderma reesei Xylanase II (Xyn II) is found in the substrate binding cleft, along with Trp96 and Tyr179 [101]. Inactivation of T. reesei Xyn II with the mechanism-based inhibitor 2',3'-epoxypropanyl-β-d-xyloside and crystallographic analysis of the modified protein revealed a covalently modified Glu86, with the xylosyl residue occupying subsite −2 packed against Trp18 [102]. The binding of substrate was proposed to induce a conformational change that closes the thumb, necessary for optimal catalytic activity. In the T. lanuginosus xylanase, Ile128 of the thumb is in position to interact with the indole ring of Trp79; this interaction perhaps stabilizes the loop [79].
Within the group of family 11 xylanases, there is peak activity at 40–80 °C and pH 4.0–6.5 [113]. Xylanases isolated from B. circulans and B. subtilis have an optimum pH of about 6 and an optimum temperature of 55 °C, while xylanase from T. lanuginosus has an optimum pH of 6.5 and optimum temperature of 70 °C [79]. Generally, family 11 xylanases with pH optima <5.0 have an aspartic acid adjacent to their general acid-base catalytic residue, and those with optima >5.0 have an asparagine (B. subtilis N44) in the same place [82]. The xylanase obtained from C. subvermispora, a white rot fungus commonly used for biopulping, possesses an optimum pH of 5.0 and an optimum temperature of 60 °C [83]. The xylanase exhibits good thermal stability at 40 °C and 50 °C, but some activity loss occurs within a few hours at 60 °C. Surprisingly, no sequence or structure has been published for a C. subvermispora xylanase, so therefore the family to which this enzyme belongs is not known.
Figure 3.
Clustal W 2.1 xylanase multiple sequence alignment [114]. The catalytic Glu residues are indicated with arrows. Identical residues for all sequences are given by “*”, while conserved and semi-conserved substitutions are given by “:”and “.” respectively. Accession numbers for these sequences are Bacillus agaradhaerens PDB 1QH7 [103], Bacillus circulans 1BCX_A [99], Bacillus subtilis 2Z79_B [115], Fibrobacter succinogenes P35811 [116,117], Penicillium citrinium BAE71133 [118], Thermomyces lanuginosus SwissProt O43097 and PDB 1YNA [119], and Trichoderma reesei ACB38137 [120]. The F. succinogenes XynC-B sequence does not show a linker region of four serine residues at the N terminus.
Figure 3.
Clustal W 2.1 xylanase multiple sequence alignment [114]. The catalytic Glu residues are indicated with arrows. Identical residues for all sequences are given by “*”, while conserved and semi-conserved substitutions are given by “:”and “.” respectively. Accession numbers for these sequences are Bacillus agaradhaerens PDB 1QH7 [103], Bacillus circulans 1BCX_A [99], Bacillus subtilis 2Z79_B [115], Fibrobacter succinogenes P35811 [116,117], Penicillium citrinium BAE71133 [118], Thermomyces lanuginosus SwissProt O43097 and PDB 1YNA [119], and Trichoderma reesei ACB38137 [120]. The F. succinogenes XynC-B sequence does not show a linker region of four serine residues at the N terminus.

3.3. Susceptibility of Xylanase to Oxidative Inactivation
Incubation of T. lanuginosus xylanase with the LMS using T. versicolor laccase and HBT as mediator resulted in rapid xylanase inactivation. This was demonstrated to be an active site directed process by observing saturation of the inactivation and protection by the xylanase substrate xylan [40]. Inactivation occurred as a result of tryptophan oxidation, evidenced as a decrease in protein fluorescence and identification via mass spectrometry of conversion of at least Trp17 and Trp18 to oxindolealanine residues. The LMS was also demonstrated to inactivate xylanase using NHA as mediator [8].
In an effort to identify key catalytic residues, many investigators have treated xylanase with active-site directed agents, including, most relevant to this discussion, the oxidizing agent NBS. NBS is specific for tryptophan, effecting the conversion of tryptophan residues to oxindolealanine residues. There are many accounts of xylanase modification by NBS, providing evidence for an active site tryptophan, including the Bacillus sp. [82], Chainia sp. [121], Fibrobacter succinogenes [122], Schizophyllum commune [123], and Streptomyces sp. T7 [124] enzymes. As an early example of this method applied to another type of glycosidase, lysozyme, X‑ray crystallography revealed that oxidation and concomitant rotation of the indole moiety of Trp62 was sufficient to completely inhibit binding of substrate and therefore inactivate the enzyme [96,125]. Generally, inactivation studies of xylanase by NBS follow the absorbance change at 250–280 nm, from which the number of tryptophan residues oxidized per mole of enzyme can be calculated [126,127,128]. This topic is pertinent due to the susceptibility xylanase to oxidative inactivation by the LMS.
Titration of the alkalophilic Bacillus sp. Xylanase C with NBS resulted in loss of 85% activity concomitant with modification of just one Trp residue [82]. The Km of the partially inactivated xylanase increased, implicating a tryptophan in substrate binding. Xylan protected the enzyme against inactivation, a further indication of tryptophan in the active site. Bacillus sp. Xylanase C shows extreme similarity to the sequence of B. agaradhaerens xylanase, although the former has a pH optimum at pH 8.0 while the latter has a pH optimum of 5.6 [129]. In the alignment of seven family 11 xylanases shown in Figure 3, Trp19 of B. agaradhaerens, equivalent to Trp9 of B. subtilis, is conserved in all, and is also highly conserved among family 10 xylanases. Trp19 has been implicated in xylanase activity through chemical modification, leading many investigators to site the key role of tryptophan at the active site [40,99,124,130,131,132].
F. succinogenes S85 Xylanase C (XynC), with a molecular mass 63,850 Da, is composed of the three distinct domains A, B, and C, each separated by serine-rich linker peptides [96]. XynC-A and XynC-B are family 11 xylanases. Despite 42.9% identity and 71.3% similarity in amino acid sequence homology, XynC-A and XynC-B exhibit different substrate specificity. XynC-A contains four tryptophan residues, all oxidizable by NBS, and XynC-B contains seven tryptophan residues, including three found in XynC-A, but only two oxidizable by NBS [122]. The addition of NBS to F. succinogenes XynC-B demonstrated that at 18:1 molar ratio of NBS to enzyme, six of the seven tryptophan residues in XynC-B were oxidized. A molar ratio of about two NBS molecules per molar equivalent of protein was sufficient to inhibit 50% of enzyme activity; the further addition of five molar equivalents of NBS resulted in less than 10% activity. On average, oxidation of three tryptophan residues resulted in inactivation. Xylose prevented the oxidation of two tryptophan residues, and the enzyme retained 60% activity even after addition of near 10-fold molar excess of NBS. With xylohexaose, about 80% activity was preserved even at a molar ratio of 10:1 NBS:xylanase, with only one of the three most susceptible tryptophan residues oxidized at that same NBS:xylanase ratio as in the absence of xylohexaose. These three most labile tryptophan residues were found to be Trp135, Trp161, and Trp202 (equivalent to B. subtilis Trp85, Tyr108, and Trp153). In contrast to Trp135 and Trp161, which are often replaced by phenylalanine or tyrosine in other family 11 xylanases [133], Trp202 is one of two totally conserved tryptophan residues in the family (the other is F. succinogenes Trp125, equivalent to B. subtilis Trp71) and occurs in the α helix, at the interface between the helix and the back of the β sheet which forms the active site cleft. Trp202 is not anticipated to directly interact with substrate, but is solvent accessible so could be subject to oxidation by NBS.
In a departure from this pattern, the addition of NBS to Penicillium citrinium xylanase resulted in no loss of xylanase activity, even though fluorescence decreased up to a molar ratio of 10:1 NBS:xylanase. The authors suggested that tryptophan residues may be present in the microenvironment but not involved in catalysis [104].
3.4. Protection of Xylanase via Adsorption to Pulp
Xylanase that is applied for biobleaching is often adsorbed to pulp, which prevents recovery and recycling of the enzyme. Additionally, xylanase may be inhibited by unidentified components of pulp leachate [134]. However, xylanase that is adsorbed to fiber is more likely to be resistant to oxidative damage. To further understand these phenomena, a variety of hardwood and softwood pulps, from both kraft and sulfite processes, were provided as substrates for xylanases from Aureobasidium pullulans, B. circulans, S. commune, Thermoascus aurantiacus, and Trichoderma harzianum. Various degrees of recovery were observed for the T. harzianum xylanase in filtrate, and the differences were based upon the pulp type, where more xylanase was recovered from pulps of higher kappa. This may be an indication of either the accessibility of xylanase to fiber based on lignin content or there may be differences attributable to lignin type resulting in variable enzyme binding properties [135]. The A. pullulans and S. commune enzymes were well recovered from a variety of pulps, and this was especially true for S. commune xylanase, with even greater than 100% activity in some cases, perhaps due to activation of the enzyme by some pulp constituent. For example, while p-coumaric acid is known to inhibit F. succinogenes xylanase, both ferulic acid and vanillin activate this enzyme [136].
In another study aimed to evaluate the adsorption of hydrolytic enzymes to pulp, a mix of three T. reesei cellulases, a T. reesei xylanase (Xyn11), and an Aspergillus niger β-glucosidase was applied to three different types of pulp [137]. Previous studies had indicated that adsorption equilibrium for cellulases is achieved within 10–20 min at pH 4.8–5.6, irrespective of temperature, enzyme dose, enzyme purity, and substrate lignin content, although increasing pH leads to decreased adsorption due to changes in the charge distribution of the enzyme [138]. In the analysis, the adsorption of a mixture of five different enzymes with total protein load of 25 mg/g cellulose was assessed in a reaction at 2% cellulose consistency, pH 5.0, and 35 °C with constant stirring. The time zero point of adsorption on the substrates was obtained by incubating the enzyme mixture on the substrates at 4 °C for 30 min. The pulp substrates included steam pretreated softwood, catalytically delignified spruce, and a commercial microcrystalline cellulose, Avicel. The delignified spruce and Avicel samples represent almost pure (at least 94%) cellulose, while the steam pretreated softwood contained 51.5% carbohydrate and 32.5% lignin on a dry weight basis.
The xylanase evaluated in this mixed enzyme study did not possess a xylan- or cellulose-binding module. Therefore, as expected, xylanase was initially adsorbed to the substrates to a much lower extent than the cellulases. With respect to the xylanase, at time zero only a little over 25% of xylanase activity was detectable in supernatant of steam pretreated softwood application, versus about 60% for catalytically delignified spruce and about 80% for Avicel. Confirmation that the proteins were adsorbed to the substrate rather than in an inactive form in the supernatant was accomplished by quantifying the amount of protein in the hydrolysis supernatants via gel separation and imaging. Xylanase activity correlated with the amount of protein in the hydrolysis supernatant. The enzyme was demonstrated to bind most well to steam pretreated softwood, in that the enzyme was adsorbed rather rapidly after the hydrolysis of steam pretreated softwood started, and after 6 h less than 1% of the original enzyme remained in the supernatant. Within 24 h xylanase was completely bound to Avicel as well, with no enzyme activity or protein found in the hydrolysis supernatant.
The fate of S. lividans xylanase was considered to ensure that no residual enzyme remained on fiber that might come into contact with food or personal care items [139]. Radiolabeled xylanase was applied to softwood kraft pulp at 50 units/g od pulp (about 10 times higher than the commercial rate of 5 units/g od pulp), and then the xylanase stage was followed with (C+D)EDED bleaching. After the xylanase stage, 91% of the original activity of xylanase was associated with the pulp, while 3.8% was measured in the filtrate, amounting to recovery of 95% of the activity. The pulp, without washing, was carried onto the bleaching sequence. After the first stage, the C+D stage, 2% of the original xylanase activity could still be detected on pulp and 1.25% was found in the filtrate. The loss of enzyme through the bleach sequence was attributed to the extreme temperature and pH (2 for the C+D stage), although enzyme inactivation via oxidation must be considered too.
4. Mixed Enzyme Systems on Wood, Straw, and Grass (Biopulping)
The effectiveness of a complement of hydrolytic and oxidative enzymes to simultaneously digest a fibrous substrate is well documented in the biopulping literature. Biopulping is based upon the ability of some fungi to adsorb to wood chips and to produce extracellular enzymes and selectively degrade lignin in wood while leaving cellulose substantially intact [140,141]. For example, bagasse and sawdust were both effective substrates for production of xylanase and phenol oxidase by the thermophilic fungus T. aurantiacus [142]; the activities were measured at day 10 of culture growth. Similarly, maize bran, wheat bran, and sugar beet pulp could each induce the simultaneous production of both laccase and xylanase by P. cinnabarinus, measured at days 6–12 [31].
Generally, in the case of white rot fungi, there are two possible routes of wood decay: (1) simultaneous degradation of all wood components, where several polymer-degrading enzymes act on the accessible wood cell wall surfaces simultaneously, with progressive degradation from the lumen to the middle lamella resulting in a thinning of the wood cell wall [143], and (2) a more selective degradation of lignin and polyoses without progressive thinning of wood cell walls [144]. In the latter case, the wood cell wall is inaccessible to enzymes with molar mass greater than about 40 kDa [145]. The white rot fungus C. subvermispora, with demonstrated selectiveness and effectiveness on Eucalyptus grandis and Pinus taeda wood chips [146], exemplifies this mode of wood degradation. Since C. subvermispora enzymes cannot penetrate the wood cells, low molecular weight agents such as chelated Mn3+ must accomplish the decomposition. These low molecular weight oxidants are generated and activated by enzymes in the cell lumina [147], resulting in extensive degradation of lignin in the middle lamellae.
C. subvermispora does not express lignin peroxidase, although LiP-like genes have been detected. The main oxidative enzyme utilized by this fungus during growth on wood is MnP. Laccase production during the initial stages of decay can be stimulated by the addition of readily available carbon and nitrogen sources [146,148]. C. subvermispora produces hemicellulases and an incomplete set of cellulases, the latter described as such due to the absence of cellobiohydrolase activity [69,149]. The organism is also devoid of β‑xylosidase, given no detectable xylose in wood extracts [141].
C. subvermispora produces low molecular mass compounds such as oxalic acid, glyoxylic acid, and unsaturated fatty acids [146]. Degradation of nonphenolic lignin proceeds by one electron oxidation of the aromatic ring [150] or by hydrogen atom abstraction at the benzyl position [151]. Mn3+-oxalate, produced in the MnP cycle, is proposed to oxidize unsaturated lipids to peroxyl radicals [152]. Unlike chelated Mn3+, which cannot oxidize nonphenolics [148], the MnP-lipid system is strong enough to oxidize nonphenolic β-O-4 lignin model compounds [153].
C. subvermispora grows aggressively and can be used on both hardwoods and softwoods [146]. Ferraz et al. considered the biopulping of E. grandis by C. subvermispora; the authors were interested in the connection between the pattern of wood decay and enzyme production [146]. Solid state growth of the organism on wood chips was performed using malt extract as a co-substrate, and the activities of xylanase, cellulase, β-glucosidase, laccase, and peroxidase were followed, while also measuring the changes in polysaccharide and lignin contents. When assessed over time, up to 90 days, there was little glucan loss until about 60 days, with the maximum loss of just 7.3% at 90 days. Coincident with this, there was virtually no measurable cellulase activity; this was confirmed in a subsequent report, also of C. subvermispora grown on either E. grandis or P. taeda wood chips, where hemicellulases predominated over cellulases, with no detectable cellobiohydrolase [83]. Total cellulose hydrolysis activities were low but there was some cellulose depolymerization, evidenced as a viscosity decrease. Fenton chemistry, based upon Fe2+ and H2O2, was proposed to be responsible for this. In the same time frame (90 days) the lignin loss was 27% and the extractives loss was 42%. This confirms the selectiveness of C. subvermispora towards lignin and extractives removal [146]. Xylanase activity peaked at days 15 (960 units/culture) and 30 (942 units/culture), while laccase activity was greatest at day 30 (12.6 units/culture); most significantly, the peak of these hydrolytic and oxidative enzymes occurred at the same culture age. Of special note is that laccase was detected when C. subvermispora was grown on E. grandis, but not when grown on P. taeda. This might be attributable to differences in the copper content of the wood [141]; copper is a known inducer of laccase. Only peroxidases were detected in the case of C. subvermispora growth on P. taeda, and the lignin was extensively depolymerized at the initial wood decay stages [146].
To determine the impact of various culture supplements such as glucose, corn steep liquor, and Mn2+ on enzyme production, C. subvermispora was grown on E. grandis for two weeks, and then the extracted enzymes were evaluated, including MnP, laccase, and xylanase [154]. MnP was detected under all culture conditions, as was xylanase. For both MnP and xylanase, culture conditions that promoted fungal growth also raised the level of these enzymes. Concomitant with fungal growth, the pH of the wood chips decreased from 5.0 to 3.5, attributable in part to oxalic acid production. Laccase was observed in cultures supplemented with corn steep liquor or glucose, with a peak in activity on day 4, and then a drop to zero by about day 6. Laccase activity and xylanase activity were concurrently present. Under all culture conditions assayed, carbohydrate losses were lower than 3%, while lignin loss varied from only about 3% to 9%. This is a typical initial stage decay pattern for wood degradation by C. subvermispora. Significantly, high levels of xylanases were not sufficient to degrade large amounts of xylan. Cultures with large amounts of secreted xylanases had previously been reported to be inefficient for xylan degradation due to the limited lignin removal [155]. Lignin degradation or transformation did not correlate with MnP levels; even low amounts of MnP could be sufficient to result in significant lignin removal. The lack of correlation between lignin degradation and MnP activity was also observed in biobleaching experiments with T. versicolor [156]. The low water solubility of unsaturated fatty acids was proposed to limit lignin degradation by MnP-generated peroxyl radicals in cultures where high levels of MnP were observed.
In a study examining the hydrolytic and oxidative enzymes produced by white rot fungi and brown rot fungi during decay of E. grandis over 150 days, the fungi providing the highest values of lignin loss were also responsible for the highest values of polyose removal [155]. While only white rot fungi produce oxidative enzymes, both brown rot and white rot fungi produce hydrolytic enzymes throughout the biodegradation process, with brown rot organisms producing higher amounts. For example, Wolfiporia cocos, a brown rot fungus, produced a peak near 1250 xylanase units at day 15, and then again a smaller peak of 127 units at day 150. In contrast, T. versicolor, a white rot fungus, produced only 335 xylanase units at day 15 and then just near 40 units of laccase on day 60. T. versicolor and Pycnoporus coccineus, which produced the highest final lignin loss values, were also the best producers of oxidative enzymes, particularly laccase. Overall, however, the production of a large amount of enzymes did not necessarily correlate with loss of the particular wood component, and polysaccharide degradation was closely dependent on the extent of lignin removal. Lignin removal caused wood cell wall permeability, facilitating xylanase diffusion into and action on the polyose of the wood cell walls [145,155].
5. Mixed Enzyme Systems on Pulp
Xylanase functions to remove xylan, consequently breaking the link between cellulose and lignin and facilitating removal of the latter in subsequent bleaching stages [9,10,157,158]. Xylanase also induces lignin removal from the fiber surface and thus improves reagent penetration in subsequent bleach stages [157,159]. By either mechanism, xylanase makes fibers more accessible for penetration of the LMS. This has been demonstrated by a number of investigators using a variety of pulp types and enzyme preparations. A combined xylanase plus LMS treatment is more effective than a xylanase treatment alone due to the direct action of LMS on lignin [33,42,45,47,62]. Specific examples of mixed enzyme systems follow. Summary results are provided in Table 3 and Table 4.
5.1. Sequential Xylanase and Laccase Application
As early as 1993, Kantelinen et al. considered application of lignin-modifying Phlebia radiata enzymes for bleaching of pine kraft pulp [160] in combination with Streptomyces olivochromogenes xylanase, prior to a peroxide stage. When laccase was used alone, there was an insignificant change in kappa, and there was a decrease in brightness, the latter likely due to oxidation of phenols to quinones. In a sequential xylanase then laccase treatment, the hemicellulase was demonstrated to improve the accessibility of lignin to laccase, since after a peroxide stage the brightness increased (from 55.8 to 56.5 ISO) and the kappa decreased (from 16.5 to 15.8) in comparison to a xylanase only treatment. No mediator was used in this study.
A sequential xylanase-laccase treatment has also been applied to wheat straw pulp. Non-wood pulps such as wheat and rice straws are particularly important in China [9]. The study used a commercial xylanase, Pulpzyme HC (Novo Nordisk). The laccase treatment employed the laccase mediator system with HBT as mediator; the laccase was isolated from P. cinnabarinus, induced by ferulic acid. While the XE and LE sequences effected 35.4% and 47.0% delignification, respectively, the XLE sequence accomplished 64.6% delignification. Further, the molecular mass of the alkaline extracted lignin decreased, but only after the LE or XLE treatments, not after the XE treatment. This indicates that the laccase mediator system accomplishes lignin depolymerization. The enzyme treatment reduced the chlorine consumption and resulted in higher final brightness [36]. Due to the differences in the pH optima of the two enzymes, a combined treatment at the optimum conditions of either of the enzymes did not improve the results obtained with the sequential enzyme treatment.
Valls and Roncero [49], in an examination of LMS process variables, used oxygen-delignified Eucalyptus globulus kraft pulp as a substrate for the sequential enzyme treatment with Bacillus sp. xylanase, Trametes villosa laccase (Novozyme NS-51002), and HBT as mediator. HBT was demonstrated to be effective at lower dose when the laccase stage was preceded by a xylanase stage. If only a laccase stage was performed, a kappa decrease and brightness increase was only significant with an HBT dose of 1.4–2.5% on pulp. In contrast, the XL sequence resulted in a kappa decrease and brightness increase over the entire range (0.5–2.5%) of HBT on pulp. All XL pulps exhibited a smaller kappa number. When a xylanase stage preceded the LMS, a similar brightness and smaller kappa number could be obtained with 30% less laccase (14.3 units/g od pulp), 80% less HBT (0.5%) and a 45% shorter reaction time (4 h), compared to the LMS alone. Overall, the xylanase treatment was demonstrated to facilitate access to cellulose fibers, thereby boosting the effect of the LMS in reducing the residual lignin content and releasing more hexenuronic acids.
In a continuation study, using the same pulp and enzyme sources, Valls et al. [161] proceeded to vary laccase dose (1–20 units/g od pulp), mediator charge (0.5–2.5% od pulp), and time (1‑7 h); the xylanase dose remained constant at 3 units/g od pulp. The pulp treated with the sequential XL treatment was determined to have a kappa number 1–2 units smaller than pulp treated only with an L stage, but brightness results were similar. Based on modeling of the XL results, the point at 20 laccase units/g od pulp, 2.5% HBT on pulp, reacting for 5.3 h would be expected to provide the smallest kappa number (4.6) and that at 8.3 laccase units/g od pulp, 2.5% HBT on pulp, reacting for 7 h would give the highest brightness (62.1% ISO). With respect to pulp viscosity, low process levels of laccase dose, HBT charge, and time, had no effect on viscosity while medium and high levels reduced viscosity slightly, by about 100 mL/g. The LMS reduced the HexA content regardless of whether a xylanase stage was performed, but HexA content was lower in XL pulp than in L pulp. There was a lack of brightness development at high laccase dose attributed to quinone formation [55,162]. Further, while low process levels led to only modest laccase activity loss (26%), at medium and high levels laccase activity loss was substantial, at 90% and 97%, respectively, consistent with previous observations of laccase inactivation by oxidized mediators [24,34,51,163].
Valls et al. [161] further determined that a shorter reaction time could effectively remove lignin, measured as kappa decrease, but there was not much change in brightness. In contrast, a longer reaction time did not result in much more lignin removal, since there was no further kappa change, but there was a brightness increase. As a consequence the authors proposed that during the first phase of the reaction all accessible lignin is oxidized and in second phase of the reaction brightness increases due to modification of previously oxidized lignin or due to removal of colored compounds containing carbonyl or carboxyl groups.
The combined application of xylanase and laccase can be useful for removal of HexA, including from sisal (non-wood) fibers [164,165]. HexA is produced during kraft pulping via conversion of 4-O-methylglucuronic acid residues in xylan to the unsaturated hexenuronic acid [165]. HexA units contribute to kappa number and retain metal ions via chelation, and so therefore increase bleach chemical consumption. HexA units also cause brightness reversion and contribute to process scaling via calcium oxalate formation [49,166]. Xylanases can reduce HexA by releasing xylan from the fiber surface [49,158,161].
The change in the HexA content of sisal pulp was followed during a bleaching sequence performed with and without a xylanase stage and including an LMS treatment, where either the natural compound sinapyl aldehyde or the synthetic compound violuric acid was used as laccase mediator. Sinapyl aldehyde is reported to be the best of the p-hydroxycinnamic compounds as a natural laccase mediator [59]. When Agave sisalana soda-anthraquinone pulp was treated with xylanase (Novozyme Pulpzyme HC) and with T. villosa laccase (Novozyme NS-51002), there was an unexpected increase in kappa by about one unit when laccase was used in combination with sinapyl alcohol, regardless of prior xylanase treatment relative to the control pulp. There was also loss of brightness with the laccase-sinapyl alcohol system. The authors proposed a different mode of action for sinapyl alcohol as mediator versus violuric acid, attributable to phenoxyl radical formation upon laccase oxidation of sinapyl alcohol; this is not possible for violuric acid. Overall, the pulp samples treated with the laccase-sinapyl alcohol system showed no reduction in HexA, in contrast to the laccase-violuric acid system, an indication that mediators follow distinct mechanistic paths.
In a further study of the impact of the xylanase and laccase on HexA removal, E. globulus kraft pulp was bleached via the LE and XLE sequences using xylanase isolated from Bacillus sp. and commercial T. villosa laccase (Novozyme) [165]. The laccase HBT system was determined to remove HexA only after all accessible lignin was removed, and the extent of HexA removal was only dependent on laccase and mediator doses, not on reaction time. While more HexA was removed by the laccase HBT treatment relative to laccase only, HexA was even more effectively removed by XLE sequence rather than just the LE sequence. Xylanase was able to increase the accessibility of the laccase HBT system to HexA on the fiber surfaces, resulting in a reduction in the kappa number. When high levels of process variables were applied, the XLE sequence accomplished 55% reduction of kappa number, versus a 44% reduction for the LE sequence.
The combination of cellulase or hemicellulase with the laccase-violuric acid system has also been considered for deinking to produce pulps with improved physical and optical properties; the enzyme combination is anticipated to be highly efficient and have low environmental impact [9,160,167]. The work was based on the proposal that the combination of a xylanase stage followed by a laccase-gallic acid treatment may have a synergistic delignification effect [168]. Shredded ONP was pulped with either cellulase (Novozyme 476) or hemicellulase (Pulpzyme HC) for 25 min at 55–60 °C, and then with laccase (Novozyme NS-51003) and violuric acid for 30 min at the same temperature. The greatest brightness increase was observed after combination of the hemicellulase then laccase-violuric acid system; this pulp also showed a good increase in specific surface area and specific volume, although the greatest specific surface area was observed for pulp treated only with laccase-violuric. ESEM images showed changes in the fiber surface, in that the surface was rougher and more fibrils were discernible after either the cellulase or hemicellulase treatment. Generally, changes in fiber morphology such as cracks, pores, and peeling were observed in enzyme-combining deinked pulps. These cracks and pores were proposed to facilitate the diffusion of lignin out through the fiber cell wall.
5.2. Simultaneous Xylanase and Laccase Application
In an early study of the simultaneous use of T. versicolor laccase and T. reesei xylanase, application of the LMS using HBT as mediator with xylanase in a single stage was found to be relatively ineffective, and this was attributed to the inactivation of xylanase by the oxidized mediator [8,88]. Laccase was also susceptible to inactivation [34,169].
Since NHA is known to cause less damage than HBT to laccase, and so then perhaps xylanase, the combination of NHA with xylanase and laccase was evaluated [8]. Both the simultaneous and sequential combination of T. reesei xylanase with T. hirsuta laccase-mediator bleaching systems resulted in an enhancement of pulp bleachability. The greatest delignification occurred in the sequential treatment (70.6% delignification), followed next by the simultaneous enzyme application (67.2% delignification) versus 63.8% for the LMS alone. Significantly, the simultaneous enzyme treatment with NHA as mediator produced greater delignification than the sequential enzyme treatment with HBT as mediator (67.2% versus 65.5%). The beneficial effects of combined treatments extended also to pulp brightness values, with the highest brightness value observed in pulps treated sequentially with xylanase and the LMS using NHA as mediator (62.2% ISO versus 61.3% for the simultaneous treatment).
The combination of T. hirsuta laccase and Ecopulp X-200 (Primalco Ltd Biotec, Rajamäki, Finland), a commercial xylanase, whether sequential or simultaneous, was determined to be better than either enzyme stage alone [7]. A xylanase stage, before or together with the LMS, using HBT as mediator, slightly improved the effect of LMS and gave a higher final brightness after peroxide bleaching than without the xylanase treatment. For example, the brightness after the sequential XEnQP sequence was 88.5% versus 88.1% after the simultaneous (X+En)QP sequence. Further, the pulp bleached with the simultaneous enzyme combination consumed less H2O2 (1.2% versus 1.4%) and exhibited a higher final viscosity (750 mL/g versus 720 mL/g) than the pulp bleached with a sequential enzyme combination.
Given that the successive application of xylanase followed by laccase had been reported to increase delignification [9,11], then the simultaneous application of laccase, cultivated from T. versicolor on wood, and a commercial xylanase (Novozyme Pulpzyme HC) was expected to be at least as effective [170]. A eucalyptus pulp, kappa 18.2, was subjected to sequential and simultaneous xylanase and laccase stages, using HBT as mediator for the laccase treatment. While the LEpDoEpD1 sequence resulted in a 45.6% reduction of ClO2, both the simultaneous ([X+L]EpDoEpD1) and sequential (XLEpDoEpD1) sequences provided a 55.0% reduction of ClO2 to achieve final pulp properties comparable to the reference pulp. The simultaneous and the sequential combinations of xylanase with the LMS showed the same reduction in bleach chemical consumption.
One further mixed enzyme system based on xylanase and laccase was demonstrated to be effective despite no mediator addition. Penicillium oxalicum and Pleurotus ostreatus were co-cultured for xylanase and laccase, respectively, via solid-state fermentation in an intermittent rotating bioreactor [16]. The obtained laccase-xylanase mixture was applied at 8 units/g od pulp in a ratio of 22:1 xylanase:laccase to a 60:40 eucalyptus:poplar pulp mix. No exogenous mediator was applied, although a native mediator might have been present in the enzyme extract. The simultaneous enzyme application resulted in a 21% reduction in kappa (from 14.0 to 11.1) and an 8% increase in brightness (from 35.5 ISO to 38.3 ISO); there was no subsequent extraction. In contrast, a xylanase only treatment provided just a 4% reduction in kappa (to 13.4) and a 4% increase in brightness (to 37.0 ISO). The investigators proposed that the synergistic action of xylanase and laccase caused simultaneous dissociation and degradation of lignin into smaller phenolic compounds, resulting in more effective delignification of pulp [16]. The action of xylanase and laccase also resulted in improved fiber quality: the fiber morphology after enzyme treatment exhibited greater porosity, swelling, separation, and peeling of pulp fibers in comparison to the smooth surface of untreated pulp [16].
5.3. Xylanase and MnP Application
There have been several accounts of the effectiveness of xylanase in combination with an oxidative enzyme other than laccase, notably, xylanase in a sequential process with either LiP or MnP [21,22]. For example, Niku-Paavola et al. [22] evaluated the sequential treatment of oxygen delignified pine kraft pulp with xylanase and MnP. There was a significant improvement in brightness when the sequential enzyme treatment was followed by two peroxide bleach stages, demonstrating that a lignin-modifying enzyme such as MnP can increase the bleachability of lignin when combined with xylanase. Xylanase was noted to increase the permeability of pulp which will therefore improve bleachability.
One of the most convincing accounts of synergistic delignification by a mixed enzyme system was demonstrated via application of MnP and xylanase. As noted by Bajpai et al. [36], the results of Bermek et al. [20] using the combination of MnP and xylanase clearly show that single enzymes are not able to mimic the complete biological system. The study was accomplished using hardwood kraft pulp at pH 4.5 treated with T. versicolor MnP at 50 units/g od pulp and T. reesei xylanase at 4.5 units/g od pulp [20], with key variations of temperature and peroxide application. The enzyme bleached pulp was then treated with EDTA prior to an alkaline peroxide stage.
There was a fairly dramatic impact of temperature on the delignification, with an optimum temperature of 40 °C. Further, the slow addition of H2O2, at 1.2 mL/h, to a maximum of 0.0612 mmole/g od pulp, in a simultaneous enzyme treatment provided the greatest brightness increase and kappa number reduction. Overall, when MnP and xylanase were used together, the pulp bleaching effect was much better than their individual effects. As shown in Table 4, pulp bleaching with xylanase and MnP simultaneously gave significantly higher pulp brightness than that obtained after the treatment of pulp with xylanase and MnP sequentially, indicating that MnP and xylanase exhibit a strong synergistic effect on pulp bleaching. Xylanase was proposed to increase MnP access to lignin, which is more effectively accomplished in the simultaneous application.
Enzyme stability was also considered, with and without pulp, at 40 °C over 24 h. MnP was stable without pulp in both the presence and absence of xylanase; however, MnP activity gradually decreased by 60% over 24 h in the presence of pulp, regardless of the presence or absence of xylanase. The investigators proposed the activity loss might be due to MnP adsorption to the pulp fibers. While xylanase was stable with and without pulp in absence of MnP, xylanase gradually lost activity in the presence of MnP, by 65% in 24 h without pulp. MnP also induced loss of xylanase activity in the presence of pulp, although much more gradually and to a lesser extent: the xylanase activity decreased by 30% in the first 3 hr and then remained stable over the next 21 h. In the case of xylanase in the presence of pulp the authors note that even at time zero there was much less xylanase activity than in the xylanase sample without pulp, a certain indication of xylanase adsorption to the fiber [20].
6. Obstacles
There are two obvious considerations regarding simultaneous application of mixed enzyme systems such as xylanase and laccase to a lignocellulosic substrate. First, the enzymes often possess different pH optima, with xylanases and laccases generally showing maximum activity from pH of 4–6.5 and 2‑5, respectively. However, as noted by Sigoillot et al. in their study of P. cinnabarinus grown on substrates such as sugar beet pulp and maize for the production of an enzyme cocktail for bleached pulp production, the temperature and pH profiles of xylanase and laccase were found to be very similar, with an optimal pH of about 5.0 and a thermal stability of up to 55 °C [31]. Ultimately, the extent of enzyme compatibility may depend on both the enzyme source and the substrate [171]. Since both enzymes are readily commercially available, enzyme dose could be increased to compensate for decreased enzyme activity at a given pH, provided the process economics could accommodate the increased enzyme charge.
The second challenge to a mixed enzyme system application is that the LMS is known to inactivate xylanase and in some accounts even laccase itself [8,34,40,88,169]. However, xylanase inactivation can be slowed via protection by its substrate xylan; in fact, loss of xylanase activity from supernatant in heterogeneous pulp systems may be attributable to enzyme binding to fiber, not to inactivation. As precedent for the dual function of mixed enzyme systems, organisms such as C. subvermispora and T. versicolor produce oxidative and hydrolytic enzymes simultaneously.
Overall, despite great commercial potential for a mixed enzyme system application, boundaries for compatibility of these two enzyme systems have not been clearly defined. In particular, there has been no systematic study of the susceptibility of xylanase to oxidative inactivation as a function of the oxidizing environment. In contrast to xylanase inactivation by the laccase-HBT system, there are no reports of loss of hemicellulolytic activity when xylanase is incubated with laccase-ABTS. Given the commercial availability of xylanase and laccase, delineation of conditions that would allow for simultaneous application of such a mixed enzyme system would be extremely advantageous.
| Year/Reference | Xylanase | Laccase | Mediator | Sequential or simultaneous? | Reaction conditions | Pulp type | Initial kappa | Bleach sequence following enzyme(s) | End bleach sequence | Delignification (%) | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Dose (per g od pulp) | Source organism or commercial product | Dose (per g od pulp) | Source organism or commercial product | Dose | ID | pH | Buffer | Temperature (°C) | Pulp consistency (%) | Time (h) | Brightness | Kappa | ||||||
| 1993/[160] | - | - | - | - | - | - | - | - | - | - | - | - | Pine Kraft | 33.7 | P | 35.9 | 21.8 | 35.3 |
| 2 units | Streptomyces olivochromogenes | - | - | - | - | - | 5.0 | citrate | 45 | 2.5 | 24 | Pine Kraft | 31.8 | P | 55.8 | 16.5 | 48.1 | |
| - | - | 6 units | Phlebia radita | none | none | - | 4.5 | dimethyl- succinate | room temperature | 2.5 | 24 | Pine Kraft | 33.7 | P | 38.8 | 22.1 | 34.4 | |
| 2 units | Streptomyces olivochromogenes | 6 units | Phlebia radita | none | none | Sequential XL | 5.0 then 4.5 | citrate then dimethyl- succinate | 45 then room temperature | 2.5 | 24 then 24 | Pine Kraft | 31.8 | P | 56.5 | 15.8 | 50.3 | |
| 1997/[7] | - | - | - | - | - | - | - | - | - | - | - | - | O2 Pine Kraft | 8.6 | EQP | 80.3 | 4.8 | 44.2 |
| 6 units | Ecopulp X-200 | - | - | - | - | - | 4.5 | none | 45 | 10 | 2 | O2 Pine Kraft | 8.6 | QP | 83.2 | 3.4 | 60.5 | |
| - | - | 40 units | Trametes hirsuta | 1% | HBT | - | 4.5 | none | 45 | 10 | 2 | O2 Pine Kraft | 8.6 | QP | 87.5 | 2.8 | 67.4 | |
| 6 units | Ecopulp X-200 | 40 units | Trametes hirsuta | 1% | HBT | Sequential XL | 4.5 | none | 45 | 10 | 2 | O2 Pine Kraft | 8.6 | QP | 88.5 | 2.3 | 73.3 | |
| 6 units | Ecopulp X-200 | 40 units | Trametes hirsuta | 1% | HBT | Sequential LX | 4.5 | none | 45 | 10 | 2 | O2 Pine Kraft | 8.6 | QP | 87.6 | 2.3 | 73.3 | |
| 6 units | Ecopulp X-200 | 40 units | Trametes hirsuta | 1% | HBT | Simultaneous | 4.5 | none | 45 | 10 | 2 | O2 Pine Kraft | 8.6 | QP | 88.1 | 2.5 | 70.9 | |
| 1997/[13] | - | - | - | - | - | - | - | - | - | - | - | - | Pine Kraft | 22.6 | EP | 40.1 | 19.3 | 14.6 |
| 12 units | Trichoderma reesei | - | - | - | - | - | 5.0 | none | 45 | 5 | 2 | Pine Kraft | 22.6 | EP | 43.9 | 15.7 | 30.5 | |
| - | - | 30 units | Trametes versicolor | 1% | HBT | - | 4.5 | none | 45 | 10 | 2 | Pine Kraft | 22.6 | EP | 46.4 | 12.8 | 43.4 | |
| 12 units | Trichoderma reesei | 30 units | Trametes versicolor | 1% | HBT | Sequential XL | 5.0 then 4.5 | none | 45 | 5 then 10 | 2 then 2 | Pine Kraft | 22.6 | EP | 50.8 | 10.4 | 54.0 | |
| 2002/[8] | - | - | - | - | - | - | - | 4.5 | none | 45 | 4 | 2 | Pine Kraft | 23.5 | EP | 52.1 | 13.5 | 42.5 |
| 6 units | Trichoderma reesei | - | - | - | - | - | 4.5 | none | 45 | 4 | 2 | Pine Kraft | 23.5 | EP | 55.4 | 11.4 | 51.5 | |
| - | - | 30 units | Trametes hirsuta | 1% | HBT | - | 4.5 | none | 45 | 4 | 2 | Pine Kraft | 23.5 | EP | 58.4 | 9.7 | 58.7 | |
| - | - | 30 units | Trametes hirsuta | 1% | NHA | - | 4.5 | none | 45 | 4 | 2 | Pine Kraft | 23.5 | EP | 59.5 | 8.5 | 63.8 | |
| 6 units | Trichoderma reesei | 30 units | Trametes hirsuta | 1% | HBT | Sequential XL | 4.5 | none | 45 | 4 | 2 | Pine Kraft | 23.5 | EP | 61.1 | 8.1 | 65.5 | |
| 6 units | Trichoderma reesei | 30 units | Trametes hirsuta | 1% | NHA | Sequential XL | 4.5 | none | 45 | 4 | 2 | Pine Kraft | 23.5 | EP | 62.2 | 6.9 | 70.6 | |
| 6 units | Trichoderma reesei | 30 units | Trametes hirsuta | 1% | HBT | Simultaneous | 4.5 | none | 45 | 4 | 2 | Pine Kraft | 23.5 | EP | 58.3 | 9.4 | 60.0 | |
| 6 units | Trichoderma reesei | 30 units | Trametes hirsuta | 1% | NHA | Simultaneous | 4.5 | none | 45 | 4 | 2 | Pine Kraft | 23.5 | EP | 61.3 | 7.7 | 67.2 | |
| 2002/[9] | - | - | - | - | - | - | - | - | - | - | - | - | Wheat Straw | 24.0 | E | nr | 17.3 | 27.9 |
| 1 unit | Pulpzyme HC | - | - | - | - | - | 7.0 | phosphate | 50 | 5 | 2 | Wheat Straw | 24.0 | E | nr | 15.5 | 35.4 | |
| - | - | 25 units | Pycnoporus cinnabarinus | 3% | HBT | - | 5.0 | acetate | 50 | 5 | 4 | Wheat Straw | 24.0 | E | nr | 12.7 | 47.0 | |
| 1 unit | Pulpzyme HC | 25 units | Pycnoporus cinnabarinus | 3% | HBT | Sequential XL | 7.0 then 5.0 | phosphate then acetate | 50 | 5 | 2 then 4 | Wheat Straw | 24.0 | E | nr | 8.5 | 64.6 | |
| 2006/[170] | - | - | - | Trametes versicolor | 3% | HBT | - | 4.0 | acetate | 45 | 15 | 5 | Eucalyptus Kraft | 18.2 | EpDoEpD1 (0.15 kappa factor) | 88.0 | nr | nr |
| 0.75 mg | Pulpzyme-HC | 60 units | Trametes versicolor | 3% | HBT | Sequential XL | 8.0 then 4.0 | none then acetate | 50 then 45 | 10 then 15 | 2 then 5 | Eucalyptus Kraft | 18.2 | EpDoEpD1 (0.12 kappa factor) | 88.0 | nr | nr | |
| 0.75 mg | Pulpzyme-HC | 60 units | Trametes versicolor | 3% | HBT | Simultaneous | 4.0 | acetate | 45 | 15 | 5 | Eucalyptus Kraft | 18.2 | EpDoEpD1 (0.12 kappa factor) | 88.3 | nr | nr | |
| 2009/[49] | 3 units | Bacillus sp. | - | - | - | - | - | 7.0 | Tris | 50 | 10 | 2 | O2 Eucalyptus Kraft | 8.4 | none | 53.7 | 8.4 | 0 |
| - | - | 10.5 units | NS-51002
(T. villosa) | 1.5% | HBT | - | 4.0 | tartrate | 30 | 5 | 4 | O2 Eucalyptus Kraft | 8.4 | E | 67.0 | 5.0 | 40.5 | |
| 3 units | Bacillus sp. | 10.5 units | NS-51002
(T. villosa) | 1.5% | HBT | Sequential XL | 7.0 then 4.0 | Tris then tartrate | 50 then 30 | 10 then 5 | 2 then 4 | O2 Eucalyptus Kraft | 8.4 | E | 70.2 | 4.0 | 52.4 | |
| 2010/[161] | 3 units | Bacillus sp. | - | - | - | - | - | 7.0 | Tris | 50 | 10 | 2 | O2 Eucalyptus Kraft | 8.4 | none | 53.7 | 8.4 | 0 |
| - | - | 10.5 units | NS-51002
(T. villosa) | 1.5% | HBT | - | 4.0 | tartrate | 30 | 5 | 4 | O2 Eucalyptus Kraft | 8.4 | none | 59.8 | 7.0 | 16.7 | |
| 3 units | Bacillus sp. | 10.5 units | NS-51002
(T. villosa) | 1.5% | HBT | Sequential XL | 7.0 then 4.0 | Tris then tartrate | 50 then 30 | 10 then 5 | 2 then 4 | O2 Eucalyptus Kraft | 8.4 | none | 58.8 | 5.2 | 38.1 | |
| 2010/[16] | 10 units | Penicillium oxalicum | - | - | - | - | - | 9.0 | none | 55 | 10 | 2 | 60:40 Eucalyptus: Poplar | 14.0 | none | 37.0 | 13.4 | 4.3 |
| 9.6 units | Penicillium oxalicum | 0.4 units | Pleurotus ostreatus | none | none | Simultaneous | 9.0 | none | 55 | 10 | 2 | 60:40 Eucalyptus: Poplar | 14.0 | none | 38.3 | 12.7 | 9.3 | |
| 7.7 units | Penicillium oxalicum | 0.3 units | Pleurotus ostreatus | none | none | Simultaneous | 9.0 | none | 55 | 10 | 3 | 60:40 Eucalyptus: Poplar | 14.0 | none | 38.3 | 11.1 | 20.7 | |
nr = not reported.
Table 4.
Trichoderma reesei Xylanase and Trametes versicolor MnP Treatment of Hardwood Kraft Pulp. Adapted with permission of authors from Bermek et al. [20].
| Treatment † prior to Ep stage | Kappa number | Delignification (%) | ISO brightness (%) |
|---|---|---|---|
| None | 8.0 | - | 59.3 |
| Xylanase (no H2O2) | 6.2 | 23 | 65.6 |
| MnP (0.0612 mmole H2O2/g pulp) * | 6.6 ‡ | 18 | 62.5‡ |
| Xylanase + MnP (simultaneous) (0.0612 mmole H2O2/g pulp) * | 5.2 | 35 | 70.5 |
| Xylanase + MnP (simultaneous) ** | 4.3 | 46 | 78.1 |
| Xylanase→MnP (sequential) ** | 6.6 | 18 | 64.4 |
| MnP→Xylanase (sequential) ** | 6.4 | 20 | 65.9 |
†Treatment of 1 g pulp HWKP (starting kappa 13.7 and brightness 35.3%) for 24 h, at 2% csc, pH 4.5, with 1 mM MnSO4, and 100 μL 10% Tween 80; when applied, MnP was used at 50 units/g pulp and xylanase at 4.5 units/g pulp; *H2O2 added in one portion at reaction start; **H2O2 added at a rate of 1.2 mL/h to the enzyme stages for a total of 0.1224 mmole for the sequential treatment and 0.0612 mmole for the simultaneous treatment; ‡value estimated from figure in original text.
7. Conclusions
Biomass processing with mixed hydrolytic and oxidative enzyme systems has been well documented in the biopulping and pulp biobleaching literature. The removal of xylan by hemicellulase improves the accessibility of lignin substrates to laccase and thus enhances the extractability of lignin from fibers. Moreover, lignin removal increases wood cell wall permeability, thus facilitating xylanase diffusion into and action on the hemicellulose component of the wood cell walls [145,155]. The application of a large dose of a single type of enzyme to wood does not correlate with loss of the particular wood component, but polysaccharide degradation is closely dependent on the extent of lignin removal. Biopulping cultures that produce large amounts of xylanases but no oxidases are ineffective for xylan degradation due to the limited lignin removal [155].
Even though the effectiveness of mixed hydrolytic and oxidative systems might be moderated when used simultaneously due to the inactivating effect of the redox mediator on the enzymes, in practice it would be beneficial to combine the treatments [8,88]. Fiber is expected to have a protective effect, and lignin will provide an alternate target for the oxidized mediators [8]. The identification of hemicellulases more robust to the oxidative environment generated by the LMS is an opportunity not yet exploited. For example, while most xylanases are readily inactivated by the tryptophan modifying agent NBS, the P. citrinium xylanase was not inactivated by even a ten-fold molar excess of this oxidant [104]. The xylanase from C. subvermispora may also exhibit stability in an oxidative environment, since the enzyme is produced along with laccase and MnP during biopulping. Inactivation studies have not been reported for the C. subvermispora xylanase, nor has sequence and structural information been published.
In consideration of simultaneous enzyme application, enzymes of similar pH optima and thermal stability would be required [31]; alternatively, a complement of enzymes of varying pH optimum may be useful. In wood degradation by C. subvermispora, the pH gradually decreases from 5.0 to near 3.5, so the production of laccase isozymes of different pH optima ensures that oxidation occurs over this pH range [69]. Different substrates are also known to exhibit different pH optima.
Overall, the combination of xylanase with an oxidative enzyme system provides an opportunity to prepare cellulose for papermaking or as a feedstock for chemical synthesis, including biofuel production. The enzyme systems may even act synergistically, evidenced as the markedly lower kappa and higher brightness pulp from simultaneous application of T. reesei xylanase and T. versicolor MnP to hardwood kraft pulp in comparison to treatment with either enzyme alone or in sequence.
Acknowledgements
The author gratefully acknowledges the support of BASF Corporation and Marist College.
Abbreviations
| ABTS | 2,2'-azinobis(3-ethylbenzothiazoline-6-sulfonic acid) |
| Ep | peroxide fortified extraction |
| EPR | electron paramagnetic resonance |
| ESEM | environmental scanning electron microscopy |
| HBT | 1‑hydroxybenzotriazole |
| HexA | hexenuronic acid |
| HPI | N-hydroxyphthalimide |
| kDa | kilodalton |
| HWKP | hardwood kraft pulp |
| L | laccase treatment stage |
| LCC | lignin carbohydrate complex |
| LiP | lignin peroxidase |
| LMS | laccase mediator system |
| MnP | manganese peroxidase |
| NHA | N-hydroxyacetanilide |
| NMR | nuclear magnetic resonance |
| od | oven dried |
| ONP | old newspaper pulp |
| P | peroxide stage |
| TEMPO | 2,2,6,6-tetramethyl-1-piperidinyloxy, free radical |
| TMB | 1,2,3,4-tetramethoxybenzene |
| X | xylanase treatment stage |
Conflicts of Interest
The author declares no conflict of interest.
References
- Sigoillot, C.; Camarero, S.; Vidal, T.; Record, E.; Asther, M.; Pérez-Boada, M.; Martínez, M.J.; Sigoillot, J.-C.; Asther, M.; Colom, J.F.; et al. Comparison of different fungal enzymes for bleaching high-quality paper pulps. J. Biotechnol. 2005, 115, 333–343. [Google Scholar] [CrossRef]
- Eriksson, K.-E.L.; Blanchette, R.A.; Ander, P. Microbial and Enzymatic Degradation of Wood and Wood Components; Springer-Verlag: New York, NY, USA, 1990; pp. 1–87. [Google Scholar]
- Sweeney, M.D.; Xu, F. Biomass converting enzymes as industrial biocatalysts for fuels and chemicals: Recent developments. Catalysts 2012, 2, 244–263. [Google Scholar] [CrossRef]
- Kirk, T.K.; Farrell, R.L. Enzymatic “combustion”: The microbial degradation of lignin. Annu. Rev. Microbiol. 1987, 41, 465–505. [Google Scholar] [CrossRef]
- Tien, M.; Kirk, T.K. Lignin-degrading enzyme from the Hymenomycete Phanerochaete chrysosporium burds. Science 1983, 221, 661–663. [Google Scholar]
- Glenn, J.K.; Morgan, M.A.; Mayfield, M.B.; Kuwahara, M.; Gold, M.H. An extracellular H2O2-requiring enzyme preparation involved in lignin biodegradation by the white rot basidiomycete Phanerochaete chrysosporium. Biochem. Biophys. Res. Commun. 1983, 114, 1077–1083. [Google Scholar] [CrossRef]
- Poppius-Levlin, K.; Wang, W.; Ranua, M.; Niku-Paavola, M.L.; Viikari, L. Biobleaching of chemical pulps by laccase/mediator systems. In Proceedings of TAPPI Biological Science Symposium, San Francisco, CA, USA, 20–23 October 1997; TAPPI: San Francisco, CA, USA, 1997; pp. 329–333. [Google Scholar]
- Oksanen, T.; Buchert, J.; Amann, M.; Candussio, A.; Viikari, L. Boosting of LMS-bleaching with hemicellulases. In Progress in Biotechnology; Viikari, L., Lantto, R., Eds.; Elsevier: Amsterdam, The Netherlands, 2002; Volume 21, pp. 255–262. [Google Scholar]
- Herpoël, I.; Jeller, H.; Fang, G.; Petit-Conil, M.; Bourbonnais, R.; Robert, J.L.; Asther, M.; Sigoillot, J.C. Efficient enzymatic delignification of wheat straw pulp by a sequential xylanase-laccase mediator treatment. J. Pulp Pap. Sci. 2002, 28, 67–71. [Google Scholar]
- Surma-Ślusarska, B.; Leks-Stępień, J. TCF bleaching of kraft pulps with laccase and xylanase. J. Wood Chem. Technol. 2001, 21, 361–370. [Google Scholar] [CrossRef]
- Viikari, L.; Oksanen, T.; Buchert, J.; Amann, M.; Candussio, A. Combined action of hemicellulases and oxidases in bleaching. In Proceedings of 10th International Symposium on Wood and Pulping Chemistry–10th Biennial ISWPC, Yokohama, Japan, 7–10 June 1999; Japan TAPPI: Yokohama, Japan, 1999; pp. 504–508. [Google Scholar]
- Freudenreich, J.; Amann, M.; Fritz-Langhals, E.; Stohrer, J. Understanding the Lignozym-process. In Proceedings of International Pulp Bleaching Conference, Helsinki, Finland, 1998; IBBC: Helsinki, Finland, 1998; pp. 71–76. [Google Scholar]
- Oksanen, T.; Amann, M.; Candussio, A.; Buchert, J.; Viikari, L. Role of xylanase in laccase-mediator delignification of kraft pulps. In Proceedings of 9th International Symposium on Wood and Pulping Chemistry, Montreal, Canada, 9–12 June 1997; CPPA: Montreal, Canada, 1997; pp. 83-81–83-84. [Google Scholar]
- Nelson, P.J.; Chin, C.W.J.; Viikari, L.; Tenkanen, M. The use of a laccase-mediator stage in bleaching of eucalypt kraft pulps. Appita J. 1998, 51, 451–455. [Google Scholar]
- Richard, T.L. Challenges in Scaling Up Biofuels Infrastructure. Science 2010, 329, 793–796. [Google Scholar] [CrossRef]
- Dwivedi, P.; Vivekanand, V.; Pareek, N.; Sharma, A.; Singh, R.P. Bleach enhancement of mixed wood pulp by xylanase-laccase concoction derived through co-culture strategy. Appl. Biochem. Biotechnol. 2010, 160, 255–268. [Google Scholar] [CrossRef]
- Feijoo, G.; Moreira, M.T.; Álvarez, P.; Lú-Chau, T.A.; Lema, J.M. Evaluation of the enzyme manganese peroxidase in an industrial sequence for the lignin oxidation and bleaching of eucalyptus kraft pulp. J. Appl. Polym. Sci. 2008, 109, 1319–1327. [Google Scholar] [CrossRef]
- Jørgensen, H.; Kristensen, J.B.; Felby, C. Enzymatic conversion of lignocellulose into fermentable sugars: Challenges and opportunities. Biofuel Bioprod. Bior. 2007, 1, 119–134. [Google Scholar] [CrossRef]
- Kumar, P.; Barrett, D.M.; Delwiche, M.J.; Stroeve, P. Methods for pretreatment of lignocellulosic biomass for efficient hydrolysis and biofuel production. Ind. Eng. Chem. Res. 2009, 48, 3713–3729. [Google Scholar] [CrossRef]
- Bermek, H.; Li, K.; Eriksson, K.-E.L. Pulp bleaching with manganese peroxidase and xylanase: A synergistic effect. Tappi J. 2000, 83, 69. [Google Scholar]
- Olsen, W.L.; Gallagher, H.P.; Burris, K.A.; Bhattacharjee, S.S.; Slocomb, J.P.; DeWitt, D.M. Enzymatic delignification of lignocellulosic material. EP0406617, 9 January 1991. [Google Scholar]
- Niku-Paavola, M.L.; Ranua, M.; Suurnäkki, A.; Kantelinen, A. Effects of lignin-modifying enzymes on pine kraft pulp. Bioresour. Technol. 1994, 50, 73–77. [Google Scholar] [CrossRef]
- Thurston, C.F. The structure and function of fungal laccases. Microbiology 1994, 140, 19–26. [Google Scholar] [CrossRef]
- Rochefort, D.; Leech, D.; Bourbonnais, R. Electron transfer mediator systems for bleaching of paper pulp. Green Chem. 2004, 6, 14–24. [Google Scholar] [CrossRef]
- Cañas, A.I.; Camarero, S. Laccases and their natural mediators: Biotechnological tools for sustainable eco-friendly processes. Biotechnol. Adv. 2010, 28, 694–705. [Google Scholar] [CrossRef]
- Solomon, E.I.; Sundaram, U.M.; Machonkin, T.E. Multicopper oxidases and oxygenases. Chem. Rev. 1996, 96, 2563–2606. [Google Scholar] [CrossRef]
- Morozova, O.V.; Shumakovich, G.P.; Gorbacheva, M.A.; Shleev, S.V.; Yaropolov, A.I. “Blue” laccases. Biochemistry 2007, 72, 1136–1150. [Google Scholar]
- Yaropolov, A.I.; Skorobogat’ko, O.V.; Vartanov, S.S.; Varfolomeyev, S.D. Laccase. Appl. Biochem. Biotechnol. 1994, 49, 257–280. [Google Scholar]
- Higuchi, T. Lignin biochemistry: Biosynthesis and biodegradation. Wood Sci. Technol. 1990, 24, 23–63. [Google Scholar] [CrossRef]
- Kawai, S.; Umezawa, T.; Shimada, M.; Higuchi, T.; Koide, K.; Nishida, T.; Morohoshi, N.; Haraguchi, T. Cα-Cβ cleavage of phenolic β-1 lignin substructure model compound by laccase of Coriolus versicolor. Mokuzai Gakkaishi 1987, 33, 792–797. [Google Scholar]
- Sigoillot, C.; Lomascolo, A.; Record, E.; Robert, J.L.; Asther, M.; Sigoillot, J.-C. Lignocellulolytic and hemicellulolytic system of Pycnoporus cinnabarinus: Isolation and characterization of a cellobiose dehydrogenase and a new xylanase. Enzyme Microb. Technol. 2002, 31, 876–883. [Google Scholar] [CrossRef]
- Kawai, S.; Umezawa, T.; Higuchi, T. Oxidation of methoxylated benzyl alcohols by laccase of Coriolus versicolor in the presence of syringaldehyde. Wood Res. 1989, 76, 10–16. [Google Scholar]
- Bourbonnais, R.; Paice, M.G. Oxidation of non-phenolic substrates: An expanded role for laccase in lignin biodegradation. FEBS Lett. 1990, 267, 99–102. [Google Scholar] [CrossRef]
- Paice, M.G.; Bourbonnais, R.; Reid, I.; Archibald, F.S. Kraft pulp bleaching by redox enzymes. In Proceedings of International Symposium on Wood and Pulping Chemistry, Montreal, Canada, 9–12 June 1997; CPPA: Montreal, Canada, 1997; pp. PL1–1–PL1–4. [Google Scholar]
- Bourbonnais, R.; Paice, M.G. Demethylation and delignification of kraft pulp by Trametes versicolor laccase in the presence of 2,2′-azinobis-(3-ethylbenzthiazoline-6-sulphonate). Appl. Microbiol. Biotechnol. 1992, 36, 823–827. [Google Scholar]
- Bajpai, P.; Anand, A.; Bajpai, P.K. Bleaching with lignin-oxidizing enzymes. Biotechnol. Annu. Rev. 2006, 12, 349–378. [Google Scholar] [CrossRef]
- Call, H.P. Process for modifying, breaking down or bleaching lignin, materials containing lignin or like substances. WO/1994/029510, 22 December 1994. [Google Scholar]
- Call, H.P.; Mücke, I. History, overview and applications of mediated lignolytic systems, especially laccase-mediator-systems (Lignozym®-process). J. Biotechnol. 1997, 53, 163–202. [Google Scholar] [CrossRef]
- Bourbonnais, R.; Paice, M.; Freiermuth, B.; Bodie, E.; Borneman, S. Reactivities of various mediators and laccases with kraft pulp and lignin model compounds. Appl. Environ. Microbiol. 1997, 63, 4627–4632. [Google Scholar]
- Bendl, R.F.; Kandel, J.M.; Amodeo, K.D.; Ryder, A.M.; Woolridge, E.M. Characterization of the oxidative inactivation of xylanase by laccase and a redox mediator. Enzyme Microb. Technol. 2008, 43, 149–156. [Google Scholar] [CrossRef]
- Fabbrini, M.; Galli, C.; Gentili, P. Comparing the catalytic efficiency of some mediators of laccase. J. Mol. Catal. 2002, 16, 231–240. [Google Scholar]
- Barreca, A.M.; Fabbrini, M.; Galli, C.; Gentili, P.; Ljunggren, S. Laccase/mediated oxidation of a lignin model for improved delignification procedures. J. Mol. Catal. 2003, 26, 105–110. [Google Scholar]
- Cantarella, G.; Galli, C.; Gentili, P. Free radical versus electron-transfer routes of oxidation of hydrocarbons by laccase/mediator systems: Catalytic or stoichiometric procedures. J. Mol. Catal. 2003, 22, 135–144. [Google Scholar]
- Chakar, F.S.; Ragauskas, A.J. The effects of oxidative alkaline extraction stages after laccase HBT and laccase NHAA treatments—An NMR study of residual lignins. J. Wood Chem. Technol. 2000, 20, 169–184. [Google Scholar] [CrossRef]
- Camarero, S.; García, O.; Vidal, T.; Colom, J.; del Río, J.C.; Gutiérrez, A.; Gras, J.M.; Monje, R.; Martínez, M.J.; Martínez, Á.T. Efficient bleaching of non-wood high-quality paper pulp using laccase-mediator system. Enzyme Microb. Technol. 2004, 35, 113–120. [Google Scholar] [CrossRef] [Green Version]
- Garcia, O.; Camarero, S.; Colom, J.F.; Martínez, A.T.; Martínez, M.J.; Monje, R.; Vidal, T. Optimization of a laccase-mediator stage for TCF bleaching of flax pulp. Holzforschung 2003, 57, 513–519. [Google Scholar]
- Ibarra, D.; Camarero, S.; Romero, J.; Martínez, M.J.; Martínez, A.T. Integrating laccase–mediator treatment into an industrial-type sequence for totally chlorine-free bleaching of eucalypt kraft pulp. J. Chem. Technol. Biotechnol. 2006, 81, 1159–1165. [Google Scholar] [CrossRef]
- Poppius-Levlin, K.; Wang, W.; Tamminen, T.; Hortling, B.; Viikari, L.; Niku-Paavola, M.L. Effects of laccase/HBT treatment on pulp and lignin structures. J. Pulp Pap. Sci. 1999, 25, 90–94. [Google Scholar]
- Valls, C.; Roncero, M.B. Using both xylanase and laccase enzymes for pulp bleaching. Bioresour. Technol. 2009, 100, 2032–2039. [Google Scholar] [CrossRef]
- Moldes, D.; Vidal, T. Laccase-HBT bleaching of eucalyptus kraft pulp: Influence of the operating conditions. Bioresour. Technol. 2008, 99, 8565–8570. [Google Scholar] [CrossRef]
- Fillat, U.; Roncero, M.B. Effect of process parameters in laccase mediator system delignification of flax pulp. Part II. Impact on effluents properties. Chem. Eng. J. 2009, 152, 330–338. [Google Scholar] [CrossRef] [Green Version]
- Camarero, S.; Ibarra, D.; Martínez, Á.T.; Romero, J.; Gutiérrez, A.; del Río, J.C. Paper pulp delignification using laccase and natural mediators. Enzyme Microb. Technol. 2007, 40, 1264–1271. [Google Scholar] [CrossRef] [Green Version]
- Johannes, C.; Majcherczyk, A. Natural mediators in the oxidation of polycyclic aromatic hydrocarbons by laccase mediator systems. Appl. Environ. Microbiol. 2000, 66, 524–528. [Google Scholar] [CrossRef]
- Roncero, M.B.; Colom, J.F.; Vidal, T. Cellulose protection during ozone treatments of oxygen delignified Eucalyptus kraft pulp. Carbohydr. Polym. 2003, 51, 243–254. [Google Scholar] [CrossRef]
- Fillat, U.; Blanca Roncero, M. Effect of process parameters in laccase-mediator system delignification of flax pulp. Part I. Pulp properties. Chem. Eng. J. 2009, 152, 322–329. [Google Scholar] [CrossRef]
- Moldes, D.; Díaz, M.; Tzanov, T.; Vidal, T. Comparative study of the efficiency of synthetic and natural mediators in laccase-assisted bleaching of eucalyptus kraft pulp. Bioresour. Technol. 2008, 99, 7959–7965. [Google Scholar] [CrossRef]
- Shleev, S.; Persson, P.; Shumakovich, G.; Mazhugo, Y.; Yaropolov, A.; Ruzgas, T.; Gorton, L. Interaction of fungal laccases and laccase-mediator systems with lignin. Enzyme Microb. Technol. 2006, 39, 841–847. [Google Scholar] [CrossRef]
- Valls, C.; Colom, J.F.; Baffert, C.; Gimbert, I.; Roncero, M.B.; Sigoillot, J.-C. Comparing the efficiency of the laccase-NHA and laccase-HBT systems in eucalyptus pulp bleaching. Biochem. Eng. J. 2010, 49, 401–407. [Google Scholar] [CrossRef]
- Aracri, E.; Colom, J.F.; Vidal, T. Application of laccase-natural mediator systems to sisal pulp: An effective approach to biobleaching or functionalizing pulp fibres? Bioresour. Technol. 2009, 100, 5911–5916. [Google Scholar] [CrossRef]
- Camarero, S.; Ibarra, D.; Martinez, M.J.; Martinez, A.T. Lignin-derived compounds as efficient laccase mediators for decolorization of different types of recalcitrant dyes. Appl. Environ. Microbiol. 2005, 71, 1775–1784. [Google Scholar] [CrossRef]
- Eggert, C.; Temp, U.; Dean, J.F.D.; Eriksson, K.-E.L. A fungal metabolite mediates degradation of non-phenolic lignin structures and synthetic lignin by laccase. FEBS Lett. 1996, 391, 144–148. [Google Scholar] [CrossRef]
- Fillat, A.; Colom, J.F.; Vidal, T. A new approach to the biobleaching of flax pulp with laccase using natural mediators. Bioresour. Technol. 2010, 101, 4104–4110. [Google Scholar] [CrossRef]
- Amann, M. The Lignozym process coming closer to the mill. In Proceedings of International Symposium of Wood and Pulping Chemistry, Montreal, Canada, 9–12 June 1997; CPPA: Montreal, Canada, 1997; pp. F4–1–F4–5. [Google Scholar]
- Geng, X.; Li, K.; Xu, F. Investigation of hydroxamic acids as laccase-mediators for pulp bleaching. Appl. Microbiol. Biotechnol. 2004, 64, 493–496. [Google Scholar] [CrossRef]
- Xu, F.; Kulys, J.J.; Duke, K.; Li, K.; Krikstopaitis, K.; Deussen, H.-J.W.; Abbate, E.; Galinyte, V.; Schneider, P. Redox chemistry in laccase-catalyzed oxidation of N-hydroxy compounds. Appl. Environ. Microbiol. 2000, 66, 2052–2056. [Google Scholar] [CrossRef]
- Xu, F.; Shin, W.; Brown, S.H.; Wahleithner, J.A.; Sundaram, U.M.; Solomon, E.I. A study of a series of recombinant fungal laccases and bilirubin oxidase that exhibit significant differences in redox potential, substrate specificity, and stability. Biochim. Biophys. Acta 1996, 1292, 303–311. [Google Scholar] [CrossRef]
- Bourbonnais, R.; Paice, M.G. Enzymatic delignification of kraft pulp using laccase and a mediator. Tappi J. 1996, 79, 199–204. [Google Scholar]
- Baldrian, P. Fungal laccases—Occurrence and properties. FEMS Microbiol. Rev. 2006, 30, 215–242. [Google Scholar] [CrossRef]
- Fukushima, Y.; Kirk, T. Laccase component of the Ceriporiopsis subvermispora lignin-degrading system. Appl. Environ. Microbiol. 1995, 61, 872–876. [Google Scholar]
- Gold, M.H.; Glenn, J.K. Manganese peroxidase from Phanerochaete chrysosporium. Methods Enzymol. 1988, 161, 258–264. [Google Scholar]
- Cui, F.; Dolphin, D. The role of manganese in model systems related to lignin biodegradation. Holzforschung 1990, 44, 279–283. [Google Scholar] [CrossRef]
- Paice, M.G.; Bourbonnais, R.; Reid, I.D. Bleaching kraft pulps with oxidative enzymes and alkaline hydrogen peroxide. Tappi J. 1995, 78, 161–169. [Google Scholar]
- Katagiri, N.; Tsutsumi, Y.; Nishida, T. Correlation of brightening with cumulative enzyme activity related to lignin biodegradation during biobleaching of kraft pulp by white rot fungi in the solid-state fermentation system. Appl. Environ. Microbiol. 1995, 61, 617–622. [Google Scholar]
- Ni, Y.; Ju, Y.; Ohi, H. Further understanding of the manganese-induced decomposition of hydrogen peroxide. J. Pulp Pap. Sci. 2000, 26, 90–94. [Google Scholar]
- Leary, G.; Giampaolo, D. The darkening reactions of TMP and BTMP during alkaline peroxide bleaching. J. Pulp Pap. Sci. 1999, 25, 141–147. [Google Scholar]
- Glenn, J.K.; Gold, M.H. Purification and characterization of an extracellular Mn(II)-dependent peroxidase from the lignin-degrading basidiomycete, Phanerochaete chrysosporium. Arch. Biochem. Biophys. 1985, 242, 329–341. [Google Scholar] [CrossRef]
- Moreira, M.T.; Sierra-Alvarez, R.; Lema, J.M.; Feijoo, G.; Field, J.A. Oxidation of lignin in eucalyptus kraft pulp by manganese peroxidase from Bjerkandera sp. strain BOS55. Bioresour. Technol. 2001, 78, 71–79. [Google Scholar] [CrossRef]
- Bermek, H.; Li, K.; Eriksson, K.-E.L. Studies on mediators of manganese peroxidase for bleaching of wood pulps. Bioresour. Technol. 2002, 85, 249–252. [Google Scholar] [CrossRef]
- Gruber, K.; Klintschar, G.; Hayn, M.; Schlacher, A.; Steiner, W.; Kratky, C. Thermophilic xylanase from Thermomyces lanuginosus: High-resolution X-ray structure and modeling studies. Biochemistry 1998, 37, 13475–13485. [Google Scholar] [CrossRef]
- Bandivadekar, K.R.; Deshpande, V.V. Structure-function relationship of xylanase: Fluorimetric analysis of the tryptophan environment. Biochem. J. 1996, 315, 583–587. [Google Scholar]
- Jeffries, T.W. Conversion of xylose to ethanol under aerobic conditions by Candida tropicalis. Biotechnol. Lett. 1981, 3, 213–218. [Google Scholar] [CrossRef]
- Balakrishnan, H.; Satyanarayana, L.; Gaikwad, S.M.; Suresh, C.G. Structural and active site modification studies implicate Glu, Trp and Arg in the activity of xylanase from alkalophilic Bacillus sp. (NCL 87–6-10). Enzyme Microb. Technol. 2006, 39, 67–73. [Google Scholar] [CrossRef]
- Heidorne, F.O.; Magalhães, P.O.; Ferraz, A.L.; Milagres, A.M.F. Characterization of hemicellulases and cellulases produced by Ceriporiopsis subvermispora grown on wood under biopulping conditions. Enzyme Microb. Technol. 2006, 38, 436–442. [Google Scholar] [CrossRef]
- Vieira, D.S.; Degrève, L.; Ward, R.J. Characterization of temperature dependent and substrate-binding cleft movements in Bacillus circulans family 11 xylanase: A molecular dynamics investigation. Biochim. Biophys. Acta 2009, 1790, 1301–1306. [Google Scholar] [CrossRef]
- Viikari, L.; Ranua, M.; Kantelinen, A.; Sundquist, J.; Linko, M. Bleaching with enzymes. In Proceedings of the Third International Conference on Biotechnology in the Pulp and Paper Industry, Stockholm, 16–19 June 1986; STFI: Stockholm, Sweden, 1986; p. 67. [Google Scholar]
- Scott, B.P.; Young, F.; Paice, M.G. Mill-scale enzyme treatment of a softwood kraft pulp prior to bleaching: Brightness was maintained while reducing the active chlorine multiple. Pulp Pap. Can. 1993, 94, 57–61. [Google Scholar]
- Kantelinen, A.; Hortling, B.; Sundquist, J.; Linko, M.; Viikari, L. Proposed mechanism of the enzymatic bleaching of kraft pulp with xylanases. Holzforschung 1993, 47, 318–324. [Google Scholar] [CrossRef]
- Viikari, L.; Suurnäkki, A.; Grönqvist, S.; Raaska, L.; Ragauskas, A. Forest Products: Biotechnology in Pulp and Paper Processing. In Encyclopedia of Microbiology; Moselio, S., Ed.; Academic Press: Oxford, UK, 2009; pp. 80–94. [Google Scholar]
- Viikari, L.; Kantelinen, A.; Sundquist, J.; Linko, M. Xylanases in bleaching: From an idea to the industry. FEMS Microbiol. Rev. 1994, 13, 335–350. [Google Scholar] [CrossRef]
- Henrissat, B. A classification of glycosyl hydrolases based on amino acid sequence similarities. Biochem. J. 1991, 280, 309–316. [Google Scholar]
- Davies, G.; Henrissat, B. Structures and mechanisms of glycosyl hydrolases. Structure 1995, 3, 853–859. [Google Scholar] [CrossRef]
- Wong, K.K.; Tan, L.U.; Saddler, J.N. Multiplicity of beta-1,4-xylanase in microorganisms: Functions and applications. Microbiol. Rev. 1988, 52, 305–317. [Google Scholar]
- Georis, J.; Esteves, F.D.L.; Lamotte-Brasseur, J.; Bougnet, V.; Giannotta, F.; Frère, J.-M.; Devreese, B.; Granier, B. An additional aromatic interaction improves the thermostability and thermophilicity of a mesophilic family 11 xylanase: Structural basis and molecular study. Protein Sci. 2000, 9, 466–475. [Google Scholar]
- Collins, T.; Meuwis, M.-A.; Stals, I.; Claeyssens, M.; Feller, G.; Gerday, C. A novel family 8 xylanase, functional and physicochemical characterization. J. Biol. Chem. 2002, 277, 35133–35139. [Google Scholar] [CrossRef]
- Collins, T.; Gerday, C.; Feller, G. Xylanases, xylanase families and extremophilic xylanases. FEMS Microbiol. Rev. 2005, 29, 3–23. [Google Scholar] [CrossRef]
- McAllister, K.A.; Marrone, L.; Clarke, A.J. The role of tryptophan residues in substrate binding to catalytic domains A and B of xylanase C from Fibrobacter succinogenes S85. Biochim. Biophys. Acta 2000, 1480, 342–352. [Google Scholar] [CrossRef]
- Maio, S.; Ziser, L.; Aebersold, R.; Withers, S.G. Identification of glutamic acid 78 as the active site nucleophile in Bacillus subtilis xylanase using electrospray tandem mass spectrometry. Biochemistry 1994, 33, 7027–7032. [Google Scholar] [CrossRef]
- Wakarchuk, W.W.; Sung, W.L.; Campbell, R.L.; Cunningham, A.; Watson, D.C.; Yaguchi, M. Thermostabilization of the Bacillus circulans xylanase by the introduction of disulfide bonds. Protein Eng. 1994, 7, 1379–1386. [Google Scholar] [CrossRef]
- Wakarchuk, W.W.; Campbell, R.L.; Sung, W.L.; Davoodi, J.; Yaguchi, M. Mutational and crystallographic analysis of the active site residues of the Bacillus circulans xylanase. Protein Sci. 1994, 3, 467–475. [Google Scholar]
- Sabini, E.; Wilson, K.S.; Danielson, S.; Schülein, M.; Davies, G.J. Oligosaccharide binding to family 11 xylanases: Both covalent intermediate and mutant product complexes display 2,5B conformations at the active centre. Acta Crystallogr. D 2001, 57, 1344–1347. [Google Scholar] [CrossRef]
- Törrönen, A.; Harkki, A.; Rouvinen, J. Three-dimensional structure of endo-1,4-β-xylanase II from Trichoderma reesei: Two conformational states in the active site. EMBO J. 1994, 13, 2493–2501. [Google Scholar]
- Havukainen, R.; Torronen, A.; Laitinen, T.; Rouvinen, J. Covalent binding of three epoxyalkyl xylosides to the active site of endo-1,4-xylanase II from Trichoderma reesei. Biochemistry 1996, 35, 9617–9624. [Google Scholar] [CrossRef]
- Sabini, E.; Sulzenbacher, G.; Dauter, M.; Dauter, Z.; Jørgensen, P.L.; Schülein, M.; Dupont, C.; Davies, G.J.; Wilson, K.S. Catalysis and specificity in enzymatic glycoside hydrolysis: A 2,5B conformation for the glycosyl-enzyme intermediate revealed by the structure of the Bacillus agaradhaerens family 11 xylanase. Chem. Biol. 1999, 6, 483–492. [Google Scholar] [CrossRef]
- Dutta, T.; Sahoo, R.; Sinha Ray, S.; Bhattacharjee, A.; Sengupta, R.; Ghosh, S. Probing the active site environment of alkaliphilic family 11 xylanase from Penicillium citrinum: Evidence of essential histidine residue at the active site. Enzyme Microb. Technol. 2007, 41, 440–446. [Google Scholar] [CrossRef]
- Wouters, J.; Gerois, J.; Engher, D.; Vandenhaute, J.; Dusart, J.; Frere, J.M.; Depiereux, E.; Charlier, P. Crystallographic analysis of family 11 endo-β-1,4-xylanase Xyl1 from Streptomyces sp. S38. Acta Crystallogr. D 2001, 57, 1813–1819. [Google Scholar] [CrossRef]
- Krengel, U.B.; Dijkstra, W. Three dimensional analysis of endo-1,4-β-xylanase I from Aspergillus niger: Molecular basis for its low pH optimum. J. Mol. Biol. 1996, 263, 70–78. [Google Scholar] [CrossRef]
- Törrönen, A.; Rovinen, J. Structural comparison of two major endo-1,4-β-xylanases from Trichoderma reesei. Biochemistry 1995, 34, 847–856. [Google Scholar] [CrossRef]
- Withers, S.G.; Aebersold, R.R. Approaches to labeling and identification of active site residues in glycosidases. Protein Sci. 1995, 4, 361–372. [Google Scholar] [CrossRef]
- Juy, M.; Amrt, A.G.; Alzari, P.M.; Poljak, R.J.; Claeyssens, M.; Béguin, P.; Aubert, J.-P. Three-dimensional structure of a thermostable bacterial cellulase. Nature 1992, 357, 89–91. [Google Scholar] [CrossRef]
- Vyas, N.K. Atomic features of protein-carbohydrate interactions. Curr. Opin. Struct. Biol. 1991, 1, 732–740. [Google Scholar] [CrossRef]
- Sierks, M.R.; Ford, C.; Reilly, P.J.; Sevensson, B. Catalytic mechanism of fungal glucoamylase as defined by mutagenesis of Asp176, Glu179 and Glu180 in the enzyme from Aspergillus awamori. Protein Eng. 1990, 3, 193–198. [Google Scholar] [CrossRef]
- Clarke, A.J. Essential tryptophan residues in the function of cellulase from Schizophyllum commune. Biochim. Biophys. Acta 1987, 912, 424–431. [Google Scholar] [CrossRef]
- Polizeli, M.L.; Rizzatti, A.C.; Monti, R.; Terenzi, H.F.; Jorge, J.A.; Amorim, D.S. Xylanases from fungi: Properties and industrial applications. Appl. Microbiol. Biotechnol. 2005, 67, 577–591. [Google Scholar] [CrossRef]
- Larkin, M.A.; Blackshields, G.; Brown, N.P.; Chenna, R.; McGettigan, P.A.; McWilliam, H.; Valentin, F.; Wallace, I.M.; Wilm, A.; Lopez, R.; et al. Clustal W and Clustal X version 2.0. Bioinformatics 2007, 23, 2947–2948. [Google Scholar] [CrossRef]
- Vandermarliere, E.; Bourgois, T.M.; Rombouts, S.; van Campenhout, S.; Volckaert, G.; Strelkov, S.V.; Delcour, J.A.; Rabijns, A.; Courtin, C.M. Crystallographic analysis shows substrate binding at the −3 to +1 active-site subsites and at the surface of glycoside hydrolase family 11 endo-1,4-β-xylanases. Biochem. J. 2008, 410, 71–79. [Google Scholar] [CrossRef]
- Marrone, L.; McAllister, K.A.; Clarke, A.J. Characterization of function and activity of domains A, B and C of xylanase C from Fibrobacter succinogenes S85. Protein Eng. 2000, 13, 593–601. [Google Scholar] [CrossRef]
- Paradis, F.W.; Zhu, H.; Krell, P.J.; Phillips, J.P.; Forsberg, C.W. The xynC gene from Fibrobacter succinogenes S85 codes for a xylanase with two similar catalytic domains. J. Bacteriol. 1993, 175, 7666–7672. [Google Scholar]
- Tanaka, H.; Nakamura, T.; Hayashi, S.; Ohta, K. Purification and properties of an extracellular endo-1,4-beta-xylanase from Penicillium citrinum and characterization of the encoding gene. J. Biosci. Bioeng. 2005, 100, 623–630. [Google Scholar] [CrossRef]
- Schlacher, A.; Holzmann, K.; Hayn, M.; Steiner, W.; Schwab, H. Cloning and characterization of the gene for the thermostable xylanase XynA from Thermomyces lanuginosus. J. Biotechnol. 1996, 49, 211–218. [Google Scholar] [CrossRef]
- He, J.; Yu, B.; Zhang, K.; Ding, X.; Chen, D. Expression of a Trichoderma reesei β-xylanase gene in Escherichia coli and activity of the enzyme on fiber-bound substrates. Protein Expression Purif. 2009, 67, 1–6. [Google Scholar] [CrossRef]
- Deshpande, V.; Hinge, J.; Rao, M. Chemical modification of xylanases: Evidence for essential tryptophan and cysteine residues at the active site. Biochim. Biophys. Acta 1990, 1041, 172–177. [Google Scholar] [CrossRef]
- Zhu, H.; Paradis, F.W.; Krell, P.J.; Phillips, J.P.; Forsberg, C.W. Enzymatic specificities and modes of action of the two catalytic domains of the XynC xylanase from Fibrobacter succinogenes S85. J. Bacteriol. 1994, 176, 3885–3894. [Google Scholar]
- Bray, M.R.; Clarke, A.J. Identification of an essential tyrosyl residue in the binding site of Schizophyllum commune xylanase A. Biochemistry 1995, 34, 2006–2014. [Google Scholar] [CrossRef]
- Keskar, S.S.; Srinivasan, M.C.; Deshpande, V.V. Chemical modification of a xylanase from thermotolerant Streptomyces. Biochem. J. 1989, 261, 49–55. [Google Scholar]
- Blake, C.C.F.; Cassels, R.; Dobson, C.M.; Poulsen, F.M.; Williams, R.J.P.; Wilson, K.S. Structure and binding properties of hen lysozyme modified at tryptophan 62. J. Mol. Biol. 1981, 147, 73–95. [Google Scholar] [CrossRef]
- Spande, T.F.; Witkop, B. Determination of the tryptophan content of proteins with N-bromosuccinimide. Methods Enzymol. 1967, 11, 498–506. [Google Scholar]
- Witkop, B. Nonenzymatic methods for the preferential and selective cleavage and modification of proteins. Adv. Protein Chem. 1962, 16, 221–321. [Google Scholar] [CrossRef]
- Bray, M.R.; Carriere, A.D.; Clarke, A.J. Quantitation of tryptophan and tyrosine residues in proteins by fourth-derivative spectroscopy. Anal. Biochem. 1994, 221, 278–284. [Google Scholar] [CrossRef]
- Poon, D.K.Y.; Webster, P.; Withers, S.G.; McIntosh, L.P. Characterizing the pH-dependent stability and catalytic mechanism of the family 11 xylanase from the alkalophilic Bacillus agaradhaerens. Carbohydr. Res. 2003, 338, 415–421. [Google Scholar] [CrossRef]
- Biswas, S.R.; Jana, S.C.; Mishra, A.K.; Nanda, G. Production, purification and characterization of xylanase from hyperxylanolytic mutant of Aspergillus ochraceus. Biotechnol. Bioeng. 1990, 35, 244–251. [Google Scholar] [CrossRef]
- Kubacková, M.; Karácsonyi, S.; Bilisics, L.; Toman, R. Some properties of an endo-1,4-beta-d-xylanase from the ligniperdous fungus Trametes hirsuta. Folia Microbiol. 1978, 23, 202–209. [Google Scholar] [CrossRef]
- Nath, D.; Rao, M. Structural and functional role of tryptophan in xylanase from an extremophilic Bacillus: Assessment of the active site. Biochem. Biophys Res. Commun. 1998, 249, 207–212. [Google Scholar] [CrossRef]
- Hakulinen, N.; Turunen, O.; Jänis, J.; Leisola, M.; Rouvinen, J. Three-dimensional structures of thermophilic β-1,4-xylanases from Chaetomium thermophilum and Nonomuraea flexuosa. Eur. J. Biochem. 2003, 270, 1399–1412. [Google Scholar] [CrossRef]
- Senior, D.J.; Mayers, P.R.; Saddler, J.N. The interaction of xylanases with commercial pulps. Biotechnol. Bioeng. 1991, 37, 274–279. [Google Scholar] [CrossRef]
- Tatsumoto, K.; Baker, J.O.; Tucker, M.P.; Oh, K.K.; Mohagheghi, A.; Grohmann, K.; Himmel, M.E. Digestion of pretreated aspen substrates: Hydrolysis rates and adsorptive loss of cellulase enzymes. Appl. Biochem. Biotechnol. 1988, 18, 159–174. [Google Scholar] [CrossRef]
- Montgomery, L.; Flesher, B.; Stahl, D. Transfer of Bacteroides succinogenes (Hungate) to Fibrobacter gen. nov. as Fibrobacter succinogenes comb. nov. and description of Fibrobacter intestinalis sp. nov. Int. J. Syst Bacteriol 1988, 38, 430–435. [Google Scholar] [CrossRef]
- Várnai, A.; Viikari, L.; Marjamaa, K.; Siika-aho, M. Adsorption of monocomponent enzymes in enzyme mixture analyzed quantitatively during hydrolysis of lignocellulose substrates. Bioresour. Technol. 2011, 102, 1220–1227. [Google Scholar] [CrossRef]
- Kyriacou, A.; Neufeld, R.J.; MacKenzie, C.R. Effect of physical parameters on the adsorption characteristics of fractionated Trichoderma reesei cellulase components. Enzyme Microb. Technol. 1988, 10, 675–681. [Google Scholar] [CrossRef]
- Bernier, R.L., Jr.; Gray, A.L.; Moser, G.P.; Hamilton, J.; Roberge, M.; Senior, D.J. Fate of residual xylanase after treatment and bleaching of softwood kraft pulp. Bioresour. Technol. 1994, 50, 79–83. [Google Scholar] [CrossRef]
- Ferraz, A.; Guerra, A.; Mendonça, R.; Masarin, F.; Vicentim, M.P.; Aguiar, A.; Pavan, P.C. Technological advances and mechanistic basis for fungal biopulping. Enzyme Microb. Technol. 2008, 43, 178–185. [Google Scholar] [CrossRef]
- de Souza-Cruz, P.B.; Freer, J.; Siika-Aho, M.; Ferraz, A. Extraction and determination of enzymes produced by Ceriporiopsis subvermispora during biopulping of Pinus taeda wood chips. Enzyme Microb. Technol. 2004, 34, 228–234. [Google Scholar] [CrossRef]
- Machuca, A.; Durán, N. Phenol oxidases production and wood degradation by a thermophilic fungus Thermoascus aurantiacus. Appl. Biochem. Biotechnol. 1993, 43, 37–44. [Google Scholar] [CrossRef]
- Guerra, A.; Mendonça, R.; Ferraz, A. Molecular weight distribution of wood components extracted from Pinus taeda biotreated by Ceriporiopsis subvermispora. Enzyme Microb. Technol 2003, 33, 12–18. [Google Scholar] [CrossRef]
- Aguiar, A.; Souza-Cruz, P.B.D.; Ferraz, A. Oxalic acid, Fe3+-reduction activity and oxidative enzymes detected in culture extracts recovered from Pinus taeda wood chips biotreated by Ceriporiopsis subvermispora. Enzyme Microb. Technol. 2006, 38, 873–878. [Google Scholar] [CrossRef]
- Blanchette, R.A.; Krueger, E.W.; Haight, J.E.; Masood, A.; Akin, D.E. Cell wall alterations in loblolly pine wood decayed by the white-rot fungus, Ceriporiopsis subvermispora. J. Biotechnol. 1997, 53, 203–213. [Google Scholar] [CrossRef]
- Ferraz, A.; Córdova, A.M.; Machuca, A. Wood biodegradation and enzyme production by Ceriporiopsis subvermispora during solid-state fermentation of Eucalyptus grandis. Enzyme Microb. Technol. 2003, 32, 59–65. [Google Scholar] [CrossRef]
- Watanabe, T.; Koller, K.; Messner, K. Copper-dependent depolymerization of lignin in the presence of fungal metabolite, pyridine. J. Biotechnol. 1998, 62, 221–230. [Google Scholar] [CrossRef]
- Enoki, M.; Watanabe, T.; Nakagame, S.; Koller, K.; Messner, K.; Honda, Y.; Kuwahara, M. Extracellular lipid peroxidation of selective white-rot fungus, Ceriporiopsis subvermispora. FEMS Microbiol. Lett. 1999, 180, 205–211. [Google Scholar] [CrossRef]
- Sethuraman, A.; Akin, D.E.; Eriksson, K.-E.L. Plant-cell-wall-degrading enzymes produced by the white-rot fungus Ceriporiopsis subvermispora. Biotechnol. Appl. Biochem. 1998, 27, 37–47. [Google Scholar]
- Srebotnik, E.; Jensen, K.A.; Kawai, S.; Hammel, K. Evidence that Ceriporiopsis subvermispora degrades nonphenolic lignin structures by a one-electron-oxidation mechanism. Appl. Environ. Microbiol. 1997, 63, 4435–4440. [Google Scholar]
- Bao, W.; Fukushima, Y.; Jensen, K.A., Jr.; Moen, M.A.; Hammel, K.E. Oxidative degradation of non-phenolic lignin during lipid peroxidation by fungal manganese peroxidase. FEBS Lett. 1994, 354, 297–300. [Google Scholar] [CrossRef]
- Kapich, A.N.; Jensen, K.A.; Hammel, K.E. Peroxyl radicals are potential agents of lignin biodegradation. FEBS Lett. 1999, 461, 115–119. [Google Scholar] [CrossRef]
- Jensen, K.A.; Bao, W.; Kawai, S.; Srebotnik, E.; Hammel, K. Manganese-dependent cleavage of nonphenolic lignin structures by Ceriporiopsis subvermispora in the absence of lignin peroxidase. Appl. Environ. Microbiol. 1996, 62, 3679–3686. [Google Scholar]
- Vicentim, M.P.; Ferraz, A. Enzyme production and chemical alterations of Eucalyptus grandis wood during biodegradation by Ceriporiopsis subvermispora in cultures supplemented with Mn2+, corn steep liquor and glucose. Enzyme Microb. Technol. 2007, 40, 645–652. [Google Scholar] [CrossRef]
- Machuca, A.; Ferraz, A. Hydrolytic and oxidative enzymes produced by white- and brown-rot fungi during Eucalyptus grandis decay in solid medium. Enzyme Microb. Technol. 2001, 29, 386–391. [Google Scholar] [CrossRef]
- Addleman, K.; Dumonceaux, T.; Paice, M.G.; Bourbonnais, R.; Archibald, F.S. Production and characterization of Trametes versicolor mutants unable to bleach hardwood kraft pulp. Appl. Environ. Microbiol. 1995, 61, 3687–3694. [Google Scholar]
- Torres, A.L.; Roncero, M.B.; Colom, J.F.; Pastor, F.I.J.; Blanco, A.; Vidal, T. Effect of a novel enzyme on fibre morphology during ECF bleaching of oxygen delignified Eucalyptus kraft pulps. Bioresour. Technol. 2000, 74, 135–140. [Google Scholar] [CrossRef]
- Roncero, M.B.; Torres, A.L.; Colom, J.F.; Vidal, T. The effect of xylanase on lignocellulosic components during the bleaching of wood pulps. Bioresour. Technol. 2005, 96, 21–30. [Google Scholar]
- Roncero, M.B.; Torres, A.L.; Colom, J.F.; Vidal, T. Effect of xylanase on ozone bleaching kinetics and properties of Eucalyptus kraft pulp. J. Chem. Technol. Biotechnol. 2003, 78, 1023–1031. [Google Scholar] [CrossRef]
- Kantelinen, A.; Hortling, B.; Ranua, M.; Viikari, L. Effects of fungal and enzymatic treatments on isolated lignins and on pulp bleachability. Holzforschung 1993, 47, 29–35. [Google Scholar] [CrossRef]
- Valls, C.; Vidal, T.; Roncero, M.B. Boosting the effect of a laccase-mediator system by using a xylanase stage in pulp bleaching. J. Hazard. Mater. 2010, 177, 586–592. [Google Scholar] [CrossRef]
- Chakar, F.S.; Ragauskas, A.J. Formation of quinonoid structures in laccase-mediator reactions. In Oxidative Delignification Chemistry; American Chemical Society: Washington, DC, USA, 2001; Volume 785, pp. 444–455. [Google Scholar]
- Ibarra, D.; Romero, J.; Martínez, M.J.; Martínez, A.T.; Camarero, S. Exploring the enzymatic parameters for optimal delignification of eucalypt pulp by laccase-mediator. Enzyme Microb. Technol. 2006, 39, 1319–1327. [Google Scholar]
- Aracri, E.; Vidal, T. Xylanase- and laccase-aided hexenuronic acids and lignin removal from specialty sisal fibres. Carbohydr. Polym. 2011, 83, 1355–1362. [Google Scholar] [CrossRef]
- Valls, C.; Vidal, T.; Roncero, M.B. The role of xylanases and laccases on hexenuronic acid and lignin removal. Process. Biochem. 2010, 45, 425–430. [Google Scholar] [CrossRef]
- Cadena, E.M.; Vidal, T.; Torres, A.L. Can the laccase mediator system affect the chemical and refining properties of the eucalyptus pulp? Bioresour. Technol. 2010, 101, 8199–8204. [Google Scholar] [CrossRef]
- Xu, Q.; Fu, Y.; Gao, Y.; Qin, M. Performance and efficiency of old newspaper deinking by combining cellulase/hemicellulase with laccase-violuric acid system. Waste Manag. 2009, 29, 1486–1490. [Google Scholar] [CrossRef]
- Chandra, R.P.; Ragauskas, A.J. Modification of high-lignin kraft pulps with laccase. Part 2. Xylanase-enhanced strength benefits. Biotechnol. Prog. 2005, 21, 1302–1306. [Google Scholar] [CrossRef]
- Pfaller, R.; Amann, M.; Freudenreich, J. Analysis of laccase and mediator interactions in the laccase mediator system. In Proceedings of 7th International Conference on Biotechnology in the Pulp and Paper Industry, Vancouver, Canada, 16–19 June 1998; CPPA: Vancouver, Canada, 1998; pp. A99–A102. [Google Scholar]
- Bajpai, P.; Anand, A.; Sharma, N.; Mishra, S.P.; Bajpai, P.K.; Lachenal, D. Enzymes improve ECF bleaching of pulp. BioResources 2006, 1, 34–44. [Google Scholar]
- Freixo, M.D.R.; Karmali, A.; Frazão, C.; Arteiro, J.M. Production of laccase and xylanase from Coriolus versicolor grown on tomato pomace and their chromatographic behaviour on immobilized metal chelates. Process. Biochem. 2008, 43, 1265–1274. [Google Scholar] [CrossRef]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
MDPI and ACS Style
Woolridge, E.M. Mixed Enzyme Systems for Delignification of Lignocellulosic Biomass. Catalysts 2014, 4, 1-35. https://doi.org/10.3390/catal4010001
AMA Style
Woolridge EM. Mixed Enzyme Systems for Delignification of Lignocellulosic Biomass. Catalysts. 2014; 4(1):1-35. https://doi.org/10.3390/catal4010001
Chicago/Turabian StyleWoolridge, Elisa M. 2014. "Mixed Enzyme Systems for Delignification of Lignocellulosic Biomass" Catalysts 4, no. 1: 1-35. https://doi.org/10.3390/catal4010001