Catalytic Activities of Ribozymes and DNAzymes in Water and Mixed Aqueous Media


1
Department of Nanobiochemistry, Faculty of Frontiers of Innovative Research in Science and Technology (FIRST), Konan University, 7-1-20, Minatojima-Minamimachi, Chuo-ku, Kobe 650-0047, Japan
2
Frontier Institute for Biomolecular Engineering Research (FIBER), Konan University, 7-1-20, Minatojima-Minamimachi, Chuo-ku, Kobe 650-0047, Japan
*
Author to whom correspondence should be addressed.
Catalysts 2017, 7(12), 355; https://doi.org/10.3390/catal7120355
Submission received: 1 November 2017
/
Revised: 13 November 2017
/
Accepted: 20 November 2017
/
Published: 23 November 2017
(This article belongs to the Special Issue Homogeneous Catalysis and Mechanisms in Water and Biphasic Media)
Abstract
:Catalytic nucleic acids are regarded as potential therapeutic agents and biosensors. The catalytic activities of nucleic acid enzymes are usually investigated in dilute aqueous solutions, although the physical properties of the reaction environment inside living cells and that in the area proximal to the surface of biosensors in which they operate are quite different from those of pure water. The effect of the molecular environment is also an important focus of research aimed at improving and expanding nucleic acid function by addition of organic solvents to aqueous solutions. In this study, the catalytic activities of RNA and DNA enzymes (hammerhead ribozyme, 17E DNAzyme, R3C ribozyme, and 9DB1 DNAzyme) were investigated using 21 different mixed aqueous solutions comprising organic compounds. Kinetic measurements indicated that these enzymes can display enhanced catalytic activity in mixed solutions with respect to the solution containing no organic additives. Correlation analyses revealed that the turnover rate of the reaction catalyzed by hammerhead ribozyme increased in a medium with a lower dielectric constant than water, and the turnover rate of the reaction catalyzed by 17E DNAzyme increased in conditions that increased the strength of DNA interactions. On the other hand, R3C ribozyme and 9DB1 DNAzyme displayed no significant turnover activity, but their single-turnover rates increased in many mixed solutions. Our data provide insight into the activity of catalytic nucleic acids under various conditions that are applicable to the medical and technology fields, such as in living cells and in biosensors.
1. Introduction
The catalytic activities of RNA enzymes (ribozymes) and DNA enzymes (DNAzymes) are useful in the development of medical and biotechnology tools. Several types of nucleic acid enzymes have been found in living cells or were artificially made through in vitro selection using libraries of random sequence [1]. Specifically, the hammerhead ribozyme and the 8–17 DNAzyme, which effectively catalyze the site-specific cleavage of the phosphodiester bond of an RNA substrate, have been used as inhibitors of intracellular gene expression [2,3] and as biosensors for detecting target metal ions, molecules, or specific DNA and RNA sequences [4,5,6,7,8,9]. The flexibility in nucleic acid sequence design enables to convert these enzymes into ligand-responsive aptazyme systems regulating gene expression levels and into sensitive electrochemical sensors [10,11,12,13].
Nucleic acid enzymes are highly negatively charged, so they exhibit catalytic activity in the presence of metal ions, like Mg2+. In general, cation binding is required for the hybridization of the enzyme with its substrate strand, for the formation of a catalytically active complex, and for the catalytic chemical process to take place [14,15,16,17,18,19]. Studies using simple aqueous solutions have provided detailed information regarding the catalytic activity of many types of ribozymes and DNAzymes and the metal ion conditions necessary for catalysis. However, the environment inside living cells and that in the area proximal to the surface of biosensors, where nucleic acid enzymes exert their therapeutic and diagnostic activity, are quite different from the aqueous solutions commonly used for in vitro evaluation of catalytic activity. In these particular cases, nucleic acid interactions occur under sterically-restricted conditions that alter the diffusion, dynamics, and effective concentration of molecules. Moreover, intracellular water and that in the vicinity of the surface of a biosensor have different physical properties (e.g., dielectric constant and water activity) from pure water. The characteristics of these environments are known to affect the thermodynamics and kinetics of nucleic acid interactions [20]. The molecular environment of an aqueous solution is also substantially affected by addition of organic solvents. Solvents like ethanol, methanol, dimethyl sulfoxide (DMSO), and N,N-dimethylformamide (DMF) have been employed to improve oligonucleotide function in the polymerase chain reaction [21,22], molecular beacon assays [23], DNA strand exchanges [24], and to assist DNAzyme functionalization [25,26,27,28]. The presence of organic solvents changes the physical properties of a solution, and the organic molecules themselves may interact with nucleic acids. Understanding the effects of the molecular environment on the activity of nucleic acid enzymes is helpful for the use of the enzymes in practical applications.
Aqueous solutions containing water-soluble organic compounds at a concentration of a few to several tenths of a percent have been used to investigate how environmental factors influence nucleic acid behavior. These types of studies have shown that the solution composition influences base-pair interaction energy, RNA folding, hydration status, and nucleic acid–metal ion interactions [20,29]. Polyethylene glycol (PEG) is a commonly used reagent in these studies. This polymer is water-soluble, and highly purified PEGs of different molecular weights are commercially available. Notably, low-molecular-weight PEGs exist as liquids and high-molecular-weight PEGs as solids at room temperature. Mixed solutions, PEG-containing solutions in many cases, have been reported to increase the rate of RNA cleavage by ribozymes, such as the group I intron ribozyme, human delta virus-like ribozyme, hairpin ribozyme, leadzyme, and hammerhead ribozyme (see the references in [30]). Several reports also exist on the catalytic activity of DNAzymes; in these cases, the reaction rates in mixed solutions containing ethanol or other organic solvents were higher than those in the solution containing no organic solvents [26,27,28]. These studies suggest the possibility of using mixed solutions to enhance the activity of nucleic acid enzymes. However, the effects of solution composition on nucleic acid behavior have been explained by a variety of different environmental factors: excluded volume interactions (molecular crowding environment), reduced water activity (dehydrating environment), and reduced dielectric constant (stronger electrostatic interaction environment) [31]. This uncertainty is partly the result of the limited use of mixed solutions to discuss the major environmental factor in these studies. In addition, most of the reported studies evaluated the catalytic activity under single-turnover conditions. In contrast to the case of protein enzymes, the turnover of nucleic acid enzymes is intrinsically difficult to achieve, because of the stable base-pairing interactions that are established with substrate strands and because of the slow kinetics of Watson-Crick base-pair opening. Methods for enhancing the turnover number would be useful to increase the effectiveness of technologies based on the catalytic activity of nucleic acid enzymes.
The hypothesis can be made that organic solvents that reduce the strength of base-pairing interactions facilitate the release of reaction products and thus increase the catalytic turnover of the reactions catalyzed by ribozymes and DNAzymes. Here, we investigated the activities of several types of RNA-cleaving and RNA-ligating nucleic acid enzymes under single- and multiple-turnover conditions, using 21 different mixed aqueous solutions containing PEGs with an average molecular weight ranging from 2 × 104 to 2 × 102, ethylene glycol derivatives (ethylene glycol (EG), glycerol (Glyc), 1,3-propanediol (PDO), 2-methoxyethanol (MME), and 1,2-dimethoxyethane (DME)), small primary alcohols (methanol (MeOH), ethanol (EtOH), and 1-propanol (PrOH)), amide compounds (urea, formamide (FA), DMF, and acetamide (AcAm)), aprotic compounds (acetonitrile (AcCN), DMSO, and 1,4-dioxane (DOX)), or dextran with an average molecular weight of 1 × 104 (Dex). These solutions have different dielectric constant, water activity, and viscosity. Furthermore, the additives may directly interact with nucleic acids, and additives of large size occupy large volumes from which other molecules are excluded. These factors potentially influence the thermodynamics and kinetics of nucleic acid interactions [20]. Results show that the ribozymes and DNAzymes used in this study remain active in many mixed solutions, and they may even display enhanced catalytic activity. Although destabilization of base pairing between the enzyme and substrate was expected to beneficial for the catalytic turnover, the mixed solutions did not increase the turnover rate of the enzymes having extended base pairing. Our data provide insight into the catalytic activity of nucleic acid enzymes in the presence of organic compounds. This insight can improve the quality of medical and technological applications of nucleic acid enzymes, such as their use in living cells or as biosensors.
2. Results
2.1. Substrate Cleavage by Ribozymes
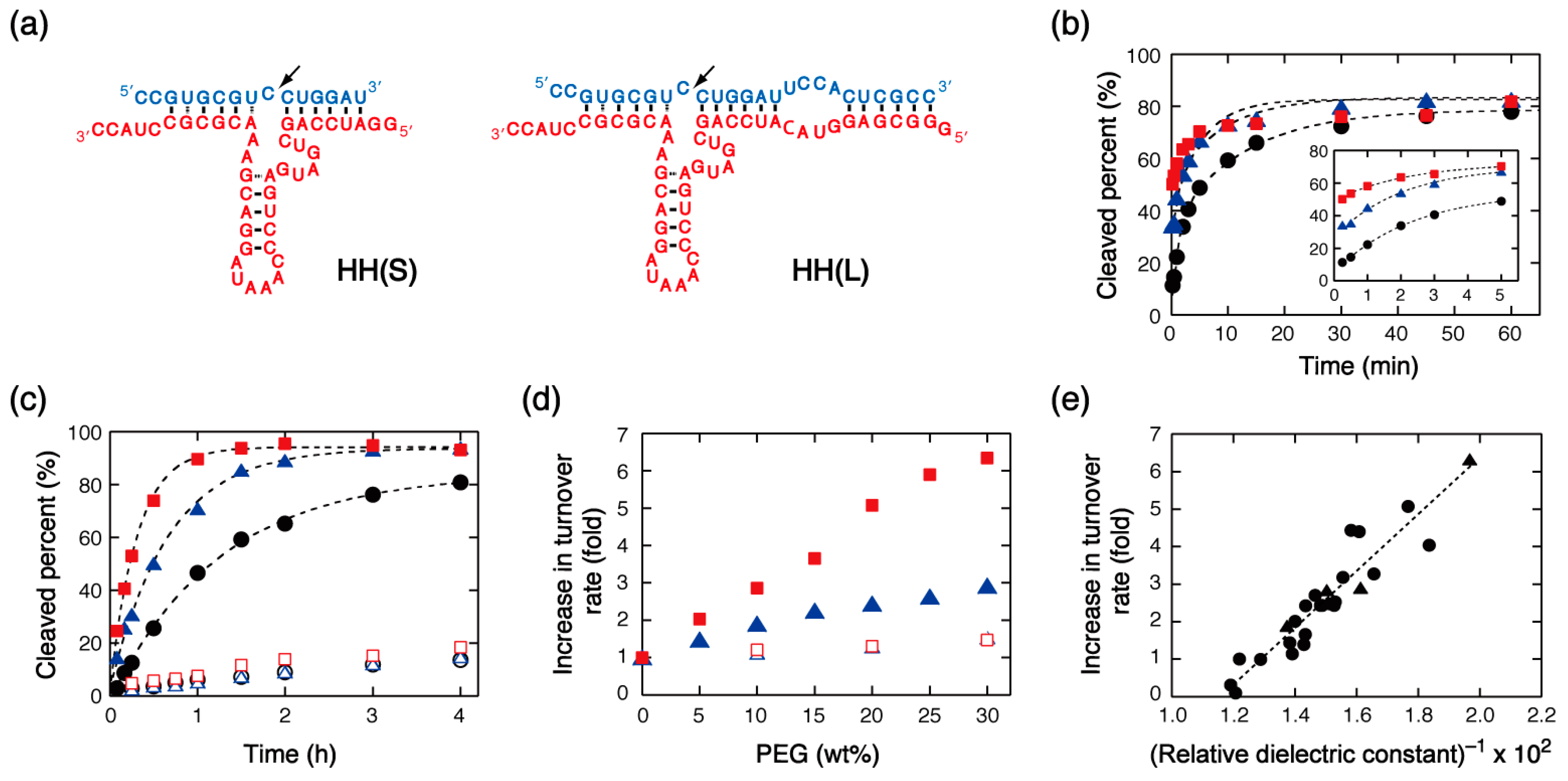
The hammerhead ribozyme is a small RNA-cleaving ribozyme found in nature. Because of its small size and high nuclease activity, this ribozyme is an attractive candidate for therapeutic and diagnostic uses [2]. HH(S) and HH(L) investigated in this study are hammerhead ribozymes having different number of base pairs formed with the substrate RNA (Figure 1a). HH(S) is a small sequence motif that can form the catalytic core. HH(L) has the extended base pairing that allows the ribozyme to form loop-loop tertiary interactions required for fast cleavage of the substrate [32,33]. Both ribozymes exhibited high RNA cleavage activity under single-turnover condition in the presence of 10 mM MgCl2, but the kinetic traces associated with substrate cleavage differed for the two systems. The time course of substrate cleavage by HH(S) was closely approximated by a single-exponential function [34], whereas HH(L) showed a biphasic kinetic behavior, whereby ~10% of the substrate was cleaved at a fast rate (within less than a few seconds) and ~70% of it was cleaved at a slow rate (taking a few tens of minutes), so that 80% substrate conversion was achieved after completion of the slow phase of the reaction (Figure 1b). This observation is consistent with a previous report that an extended ribozyme having a similar sequence to HH(L) showed a biphasic kinetic behavior [32]. It is regarded that substrate cleavage by the extended ribozyme proceeds through multiple reaction pathways: the fast phase of the reaction reflects the cleavage of the catalytically active complex, whereas its slow phase reflects the contribution of the slow conversion of kinetically trapped conformations, such as those due to misfolding or interfering secondary structures, to the active complex.
The presence of 20% PEG8000 (with an average molecular weight of 8 × 103) or PEG200 (2 × 102) influenced the single-turnover reaction rate. We have previously reported the enhancement of the rate of cleavage catalyzed by HH(S) in mixed solutions containing PEG8000 or PEG200 [34,35]. When HH(L) was investigated, the effects of PEG on the fast and slow phases of the reaction were different: the amplitude of the fast phase conversion increased 9.8-fold with PEG8000 and 5.3-fold with PEG200, although the rates were too fast for the rate constants to be determined. The rate of the slow phase was not affected by the addition of PEGs, and the amplitude after completion of the slow phase reached ~80% conversion, the same level observed in the absence of PEG (Figure 1b). These results suggest that the molecular environment made up of PEG-containing aqueous solutions increases the amount of the catalytically active complex with respect to the case of the solution containing no organic additives; however, it does not significantly affect the rate of the conformational change necessary for the formation of the active complex.
The cleavage rates of HH(S) and HH(L) under multiple-turnover conditions were investigated using a 20-fold excess of substrate RNA to ribozyme. In principle, at least two kinetic stages exist for the reaction: the first round of substrate cleavage (accounting at most for 5% of the reaction yield) and the subsequent rounds of cleavage. The kinetic data showed that HH(S) had high turnover activity in the presence of 10 mM MgCl2 (the turnover number was greater than 10 after 1.5 h), and the turnover rate increased 5.1-fold after addition of PEG8000 and 2.4-fold after addition of PEG200 (Figure 1c). By contrast, the extended ribozyme HH(L) displayed lower turnover activity than HH(S), both in the presence and absence of PEGs (the turnover number was less than three after 6 h). The turnover rate of the HH(S)-catalyzed reaction increased linearly with the amount of PEG (5–30%), but the turnover rate of the HH(L)-catalyzed reaction did not change significantly even when using 30% PEG solutions (Figure 1d).
To identify the environmental factor causing the observed effect of PEGs on HH(S), we investigated the effects of other mixed solutions having various physical properties. The kinetic data showed that the turnover rate increased in many mixed solutions with respect to the solution containing no organic additives (Table 1). Analysis of the correlation with the values of solution properties revealed that the turnover rate was highly correlated with the dielectric constant of the medium (Figure 1e), but the rate was not correlated with water activity, viscosity, or the additives’ molecular weight (Figure S1). These results suggest that the rate enhancement in mixed solutions was predominantly caused by electrostatic effects. These same correlations were previously reported for the single-turnover rates [35], suggesting that reaction rate enhancements resulted from the same mechanism in both reaction conditions. The decrease in dielectric constant observed as a result of the formation of mixed solutions may strengthen the binding interaction between nucleic acids and Mg2+ ions, which is a required process for the formation of the active complex and for catalysis. Consistent with the strengthening of Mg2+ binding to the ribozyme, the increase in turnover rate was more pronounced when MgCl2 concentration decreased to 1 mM. In particular, the turnover rate increased more than 10-fold in mixed solutions containing PEG20000, PEG8000, and PEG2000 (13, 17, and 10-fold, respectively) with respect to the solution containing no organic additives (Table 1).
2.2. Substrate Cleavage by DNAzymes
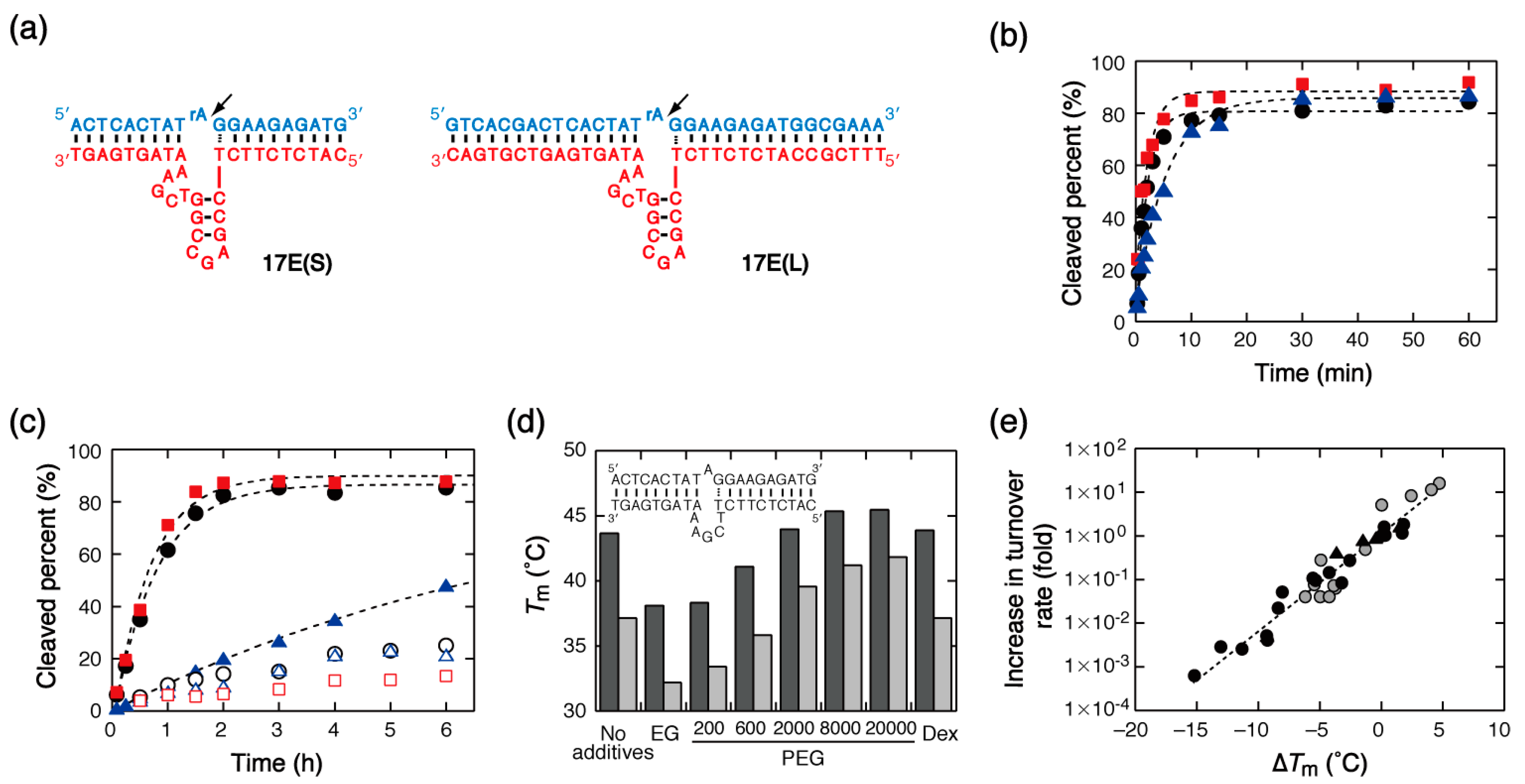
The DNA enzyme named 17E DNAzyme is a variant of the 8–17 DNAzyme obtained through in vitro selection [36,37]. This enzyme is relatively small in size and can cleave a substrate sequence in the presence of Mg2+ or other metal ions. Notably, it can be engineered to have nuclease activity in various situations, including in vivo, in blood serum, and when anchored on the surface of a biosensor [7,38,39,40]. Notably, this enzyme is being tested in clinical trials [3]. 17E(S) and 17E(L) are DNAzymes having different number of base pairs formed with the substrate DNA–RNA chimeric strand, and substrate hydrolysis occurs at a single RNA nucleotide (rA) site (Figure 2a). With these DNAzymes, rapid substrate cleavage was observed, reaching ~90% efficiency under single-turnover conditions in the presence of 10 mM MgCl2. The reaction rates of both enzymes were approximated by single exponential functions characterized by similar rate constants (0.39 min−1 and 0.50 min−1, for 17E(S) and 17E(L), respectively) in the solution containing no organic additives. This observation indicates the absence of kinetically trapped alternate conformations of the extended DNAzyme 17E(L). In contrast to the hammerhead ribozyme, 17E(S) did not show an increase in the single-turnover rate in the presence of PEG8000 (0.20 min−1) and showed a decreased rate in the presence of PEG200 (0.097 min−1), as shown in Figure 2b. The same effects of the presence of PEGs in solution were observed for 17E(L) (0.39 min−1 with PEG8000 and 0.11 min−1 with PEG200).
The rate of the substrate cleavage catalyzed by 17E(S) under multiple-turnover conditions did not show any marked change in the presence of PEG8000, and it showed a reduction in the presence of PEG200 (Figure 2c). When other mixed solutions were investigated, all mixed solutions, except those containing PEG20000 and Dex, substantially decreased the cleavage rate (Table 2). Specifically, addition of amide compounds almost completely eliminated the catalytic activity and addition of primary alcohols or aprotic compounds greatly decreased the turnover rate. The decreases in turnover rate observed in mixed solutions were characterized by the decreases in single-turnover rate (Table S1). This result suggests that the turnover rate is primarily determined by the single-turnover activity and that the effect of mixed solutions on product release is unlikely to be important. Correlation analyses revealed that the turnover rate (and also the single-turnover rate) had no significant correlation with the values of solution properties (Figure S2a–c). In addition, although the catalytic activity was observed to increase with the size of the additives, only a weak correlation was determined to exist with the additives’ molecular weight (Figure S2d). It was also found that the effects of large PEGs and Dex were particularly pronounced in the presence of 1 mM MgCl2, with 16-, 11-, 8.2-, and 5.1-fold rate enhancements in the presence of PEG20000, PEG8000, PEG2000, and Dex, respectively (Table 2).
To understand the effects of the mixed solutions on the turnover rate of the 17E(S)-catalyzed reaction, the thermal stability of the complex between the DNAzyme and its substrate was studied using a model DNA sequence of base pairs that gave the thermal melting temperature (Tm) of a two-state transition. Analysis of the melting curve indicated that the DNA structure was destabilized in the presence of small PEGs or EG, but it was slightly stabilized in the presence of large PEGs or Dex (Figure 2d). When MgCl2 concentration decreased to 1 mM, the stabilization effects of large PEGs increased. These results are in agreement with previous reports according to which small PEGs reduce the stability of DNA base pairing [41,42,43] and large PEGs are able to increase the stability at low salt concentrations [44]. Figure 2e shows the turnover rate increments plotted against the Tm changes (ΔTm) obtained in the mixed solutions, excluding the data for reactions characterized by a very slow rate (with a rate constant smaller than 1 × 10−3 h−1) as a degree of uncertainty exists for them. The turnover rate decreased as the Tm decreased, and the correlation plots for the solutions containing 10 mM MgCl2 and those containing 1 mM MgCl2 overlapped with each other. Furthermore, the data obtained with different PEG8000, PEG200, or EtOH concentrations (10% and 30%) fell on the correlation line. The high level of correlation between the increments of the turnover rate and the ΔTm of a model DNA sequence of base pairs indicates the importance of the stability of DNA interactions for the catalytic activity. In addition, the analysis using the free energy change (ΔG°) associated with complex formation inferred from the melting curve showed that the turnover rate constant is also correlated with the value of ΔG° (Figure S2e). These correlation plots may be useful for predicting the turnover rate in other mixed solutions or in solutions containing Mg2+ at concentrations not employed in this study. They may also be used to find the conditions that can enhance the turnover activity of the DNAzyme. Conversely, the presence of organic additives did not increase the turnover rate of the reaction catalyzed by the extended DNAzyme 17E(L), as shown in Figure 2c, regardless of the ability of the mixed solutions to cause changes in the single-turnover rate comparable to those of 17E(S) (Figure S2f).
2.3. Substrate Ligation by Ribozyme and DNAzyme
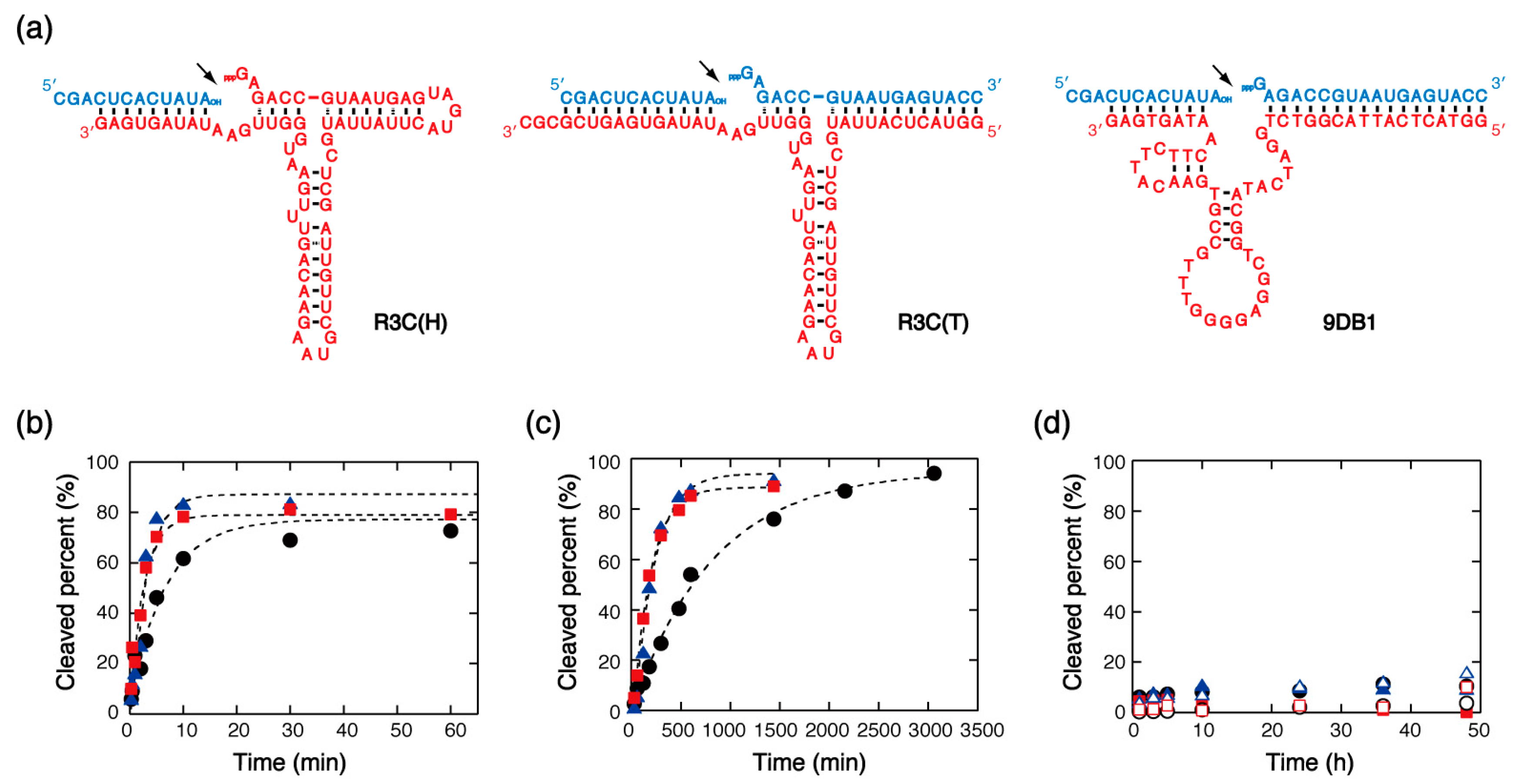
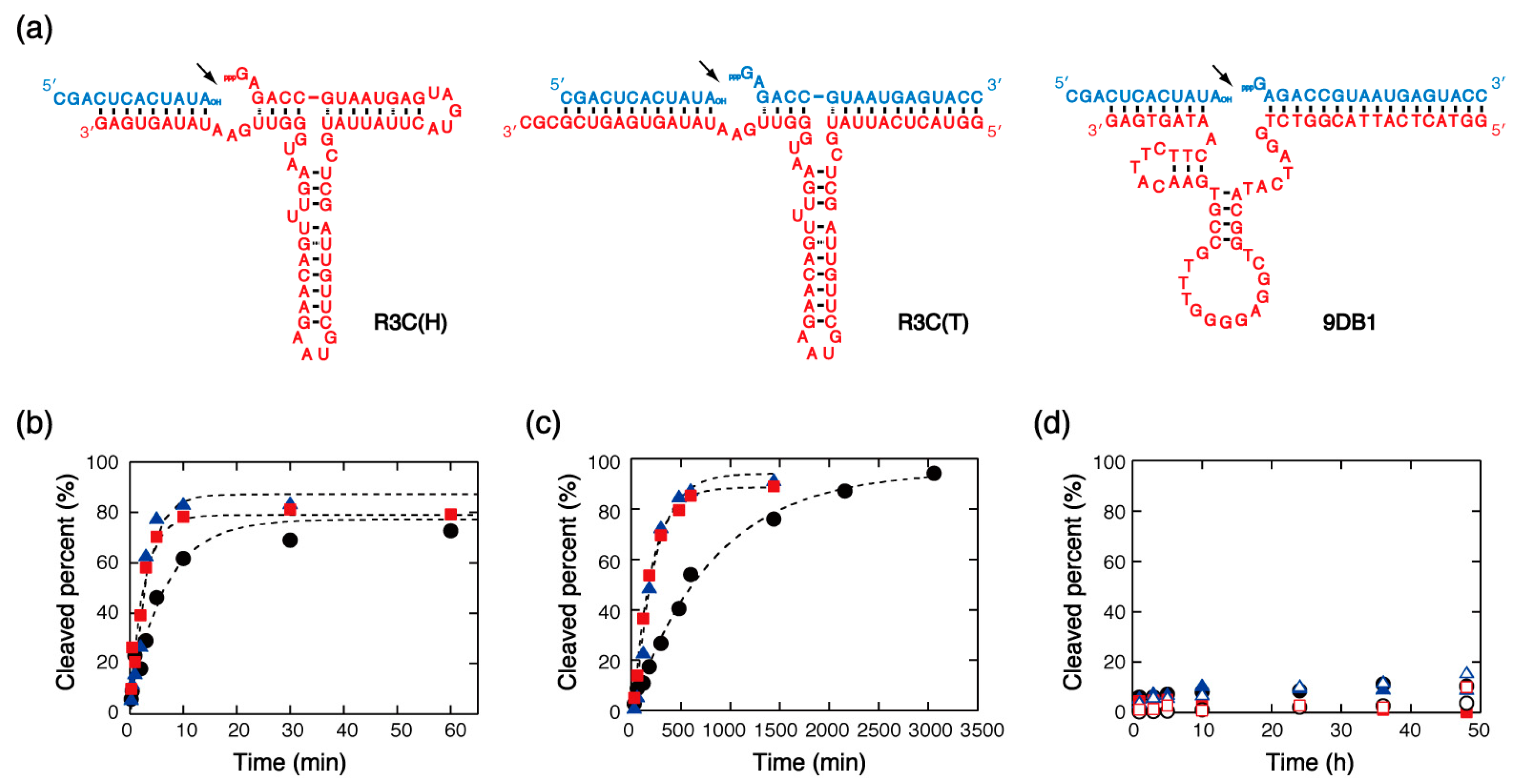
Several types of RNA-ligating ribozymes exist that catalyze the formation of covalent bonds between two RNA strands. This study investigated the ligase ribozyme derived from the R3C ribozyme obtained through in vitro selection [45,46]. This ribozyme catalyzes the formation of a phosphodiester bond between the 5′-triphosphate and 3′-hydroxyl termini of substrate RNAs in the presence of Mg2+. R3C(H) catalyzes the ligation of the ribozyme with a substrate RNA, and R3C(T) catalyzes the ligation of two substrate RNAs (Figure 3a). R3C(H) was used for the analysis of single-turnover rate, and R3C(T) was used to analyze the turnover rate. We also investigated 9DB1 shown in Figure 3a, derived from the 9DB1 DNAzyme obtained through in vitro selection [47]. This DNAzyme catalyzes the formation of a 3′,5′-phosphodiester bond of two substrate RNAs in the presence of Mg2+, and it was designed to catalyze the ligation of the same substrates as the ligase ribozymes.
Kinetic data showed that the single-turnover rate of the reaction catalyzed by R3C(H) in the presence of 10 mM MgCl2 was comparable to the rates obtained for the nuclease enzymes. Addition of PEGs to the solution increased the ligation rate (Figure 3b); and the same effect was observed for many organic additives, but addition of amide compounds greatly decreased the rate (Table 3). When the MgCl2 concentration decreased to 1 mM, almost no reaction was observed either with or without organic additives. Similar results were observed for 9DB1, although the increments of the ligation rate with respect to the solution containing no organic additives were, in most cases, greater than those observed for R3C(H) (Figure 3c and Table 3). Correlation analyses showed a less significant correlation between the single-turnover rates and the values of solution properties, the additives’ molecular weight, or base-pair stability (data not shown), suggesting that the activity of these enzymes is not simply determined by a single environmental factor. On the other hand, experiments with R3C(T) and 9DB1 showed no turnover activity whether or not organic additives were present (Figure 3d). In the case of these ligase enzymes, therefore, mixed solutions are not useful to enhance turnover activity, but they are when it comes to the single-turnover activity.
3. Discussion
3.1. Classification of the Mixed Solutions Based on the Effects on Nucleic Acids
In this study, we investigated the single- and multiple-turnover kinetics of nucleic acid enzymes in various mixed aqueous solutions that contain water-soluble organic cosolvents or cosolutes. These organic additives may alter the physical properties of solution, interact with nucleic acids, and introduce areas of excluded volume [20,31]. Most mixed solutions have lower dielectric constant values than pure water, and the additives, such as large PEGs, MME, DME, EtOH PrOH, and DOX, significantly lowers the dielectric constant of solution. The dielectric constant affects the strength of electrostatic interactions, including metal ion binding to nucleic acid phosphate groups. Hydrophilic additives, such as EG, small primary alcohols, and AcCN significantly reduce the water activity of solution. Water activity affects base pairing and tertiary folding accompanied by water association or release (hydration changes). In addition, preferential binding to or exclusion of additives from nucleic acid surface also affects the stability of nucleic acid interactions. The additives, such as MME, PrOH, amide compounds, AcCN, and DOX, significantly reduce the stability of base-pairing interactions. On the other hand, large PEGs and Dex occupy large volumes and increase the viscosity of solution. Viscosity affects the rate of substrate hybridization and the excluded-volume effect facilitates nucleic acid hybridization and folding into a compact structure by increasing effective concentration and restricting conformational dynamics. Accordingly, the molecular environment of mixed solutions is assumed to influence the activity of the nucleic acid enzymes through a combination of mechanisms.
The data shown in Table 1, Table 2 and Table 3 suggest the possibility to employ mixed solutions as reaction media of these enzymes. The solutions containing large PEGs increased the catalytic activity of all tested ribozymes and DNAzymes, even although these enzymes have different catalytic core sequences, tertiary interactions, roles of metal ions, and mechanism of catalysis. The solutions containing small PEGs, ethylene glycol derivatives, small primary alcohols, and aprotic compounds increased the turnover rate of the reactions catalyzed by HH(S) and the single-turnover rates catalyzed by R3C(H) and 9DB1, but they reduced the turnover rate catalyzed by 17E(S). The solutions containing amide compounds greatly reduced the catalytic activity of these enzymes, particularly at the low MgCl2 concentration of 1 mM. This classification based on the chemical structure and size of additives can provide some explanation for the effects on the activity of the nucleic acid enzymes, and the correlation analyses have further determined the environmental factors causing the observed effect of the additives on HH(S) and 17E(S). Conversely, the mixed solutions did not increase the turnover rate of the extended nuclease enzymes HH(L) and 17E(L) and the ligase enzymes R3C(T) and 9DB1. These observations were attributed to the reduced tendency to allow the dissociation of the long reaction products, due to the high stability of base-pairing interactions.
3.2. Comparison of the Effects of Mixed Solutions on the Nucleic Acid Enzymes
3.2.1. Effects on the Hammerhead Ribozyme
The turnover rate of the HH(S)-catalyzed reaction increased in many mixed solutions with respect to the solution containing no organic additives. This result indicates that many kinds of organic additives are useful for enhancing the turnover activity of the hammerhead ribozyme. It has been reported that the stability of base pairing and tertiary folding is reduced in the presence of organic solvents, particularly amide compounds that are known to substantially destabilize nucleic acid structures [31,44]. However, this destabilization effect seemed not to cause a disruption of interactions necessary for the formation of the active conformation, such as a network of hydrogen bonds [48], in the presence of 10 mM MgCl2. The correlation plot of Figure 1e suggests that the dielectric constant influences the turnover rate. A simple interpretation of this correlation is that the transition state is electrostatically stabilized in less-polar media. However, our previous study identified an apparent increase in Mg2+-binding affinity to HH(S) in mixed solutions with a low dielectric constant [34,35]. The substrate cleavage catalyzed by the hammerhead ribozyme requires the involvement of several Mg2+ ions, and at least one Mg2+ ion binds to the site with highly negative electrostatic potential located near the scissile phosphate of the substrate [49]. This mechanism is consistent with the dielectric constant effect that facilitates electrostatic Mg2+ binding and allows greater enhancement of the turnover activity observed at the low Mg2+ concentration of 1 mM. In the cellular environment, the dielectric constant is relatively low [50,51] and the concentration of Mg2+ is as low as 1 mM or less [52]. Thus, it is speculated that the hammerhead ribozyme is suitable for use in living cells, for example to achieve intracellular mRNA cleavage and in vivo sensing.
3.2.2. Effects on the 17E DNAzyme
The activity of 17E(S) was not enhanced in the presence of many organic additives under both single- and multiple-turnover conditions, in contrast to the case of HH(S). The result shows that increasing the activity of the 17E DNAzyme is difficult to achieve using mixed aqueous solutions. Correlation analysis revealed no significant contribution of the dielectric constant to the catalytic activity of 17E(S). Conversely, the correlation plot shown in Figure 2e suggests that the stability of DNA interactions strongly influences the turnover rate. However, the effects of mixed solutions on the single-turnover rate of the reaction catalyzed by the extended DNAzyme 17E(L) were similar to those measured for 17E(S). Thus, the destabilization effect on base pairing between the enzyme and substrate is unlikely to be an important factor. It is possible that the mixed solutions reduce the stability of DNA interactions important for formation of the catalytically active complex. Although the detailed structure and enzymatic mechanism of the 17E DNAzyme have not been elucidated, biophysical studies have shown the importance of Mg2+ binding and non-canonical hydrogen bonding in the catalytic core for catalysis [53,54,55]. It is noted that an RNA-cleaving DNAzyme exhibiting high activity in the presence of high ethanol concentration have been reported [27,28]. The active complex of this enzyme might be stable enough to overcome the destabilization effect of ethanol, unlike the case of 17E(S). The turnover activity of 17E(S) was enhanced in the presence of large PEGs and Dex, which increased the stability of DNA interactions. As the large-sized additives are characterized by an excluded-volume effect, this DNAzyme might be suitable for use as a probe anchored on the surface of a sensing material, where the excluded volume of immobilized molecules is large.
3.2.3. Effects on the R3C Ribozyme and 9DB1 DNAzyme
The turnover of the reactions catalyzed by ligases is assumed to be more difficult to achieve than it is for nucleases, because ligation leads to an increase in the number of base pairs, thus strengthening the bonding between the enzyme and reaction products. As expected, no turnovers were observed in the reactions catalyzed by R3C(T) and 9DB1 even in mixed solutions that greatly reduced the stability of base-pairing interactions. By contrast, many mixed solutions increased the single-turnover rates. The result indicates that the active complexes of these enzymes are stable enough to overcome the destabilization effect of the organic solvents. However, addition of amide compounds greatly decreased the ligation activity, suggesting the importance of the stability of catalytic active complex in the mixed solutions for catalysis. Enhancements of the rate of the ligation catalyzed by 9DB1 DNAzyme in mixed solutions containing ethanol, methanol, isoamyl alcohol, acetone, DMSO, or DMF have been previously reported [26]. Our results exhibited that many other organic compounds also have an ability to enhance the activity of this DNAzyme, although the activity was completely abolished at the low MgCl2 concentration of 1 mM. Although the detailed mechanism and structure of the R3C ribozyme have not been elucidated, the crystal structure of 9DB1 DNAzyme has been reported, and it shows the presence of tertiary interactions in the catalytic domain; however, the role of metal ions in the reaction remains unclear [56]. The mixed solutions enhanced the catalytic activities of both R3C(T) and 9DB1, implying a similar mechanism by which the environmental factors influence the activity of these enzymes, whereas correlation analyses suggested that changes in the activity of these enzymes was not determined by a single environmental factor. It is mentioned that the cleavage and ligation of RNA strands are related to RNA evolution and the RNA world, where the prebiotic environment is likely to contain organic compounds. It is fascinating to speculate on the contribution of environmental factors to the evolution of functional RNAs.
4. Materials and Methods
4.1. Materials
The hammerhead ribozyme, ligase ribozyme, and substrate RNAs having a 5′-triphosphate group were prepared by in vitro transcription using T7 RNA polymerase and purified by polyacrylamide gel electrophoresis, as described previously [34]. Other RNAs, DNAs, and a DNA–RNA chimeric strand, purified by high performance liquid chromatography, were purchased from Hokkaido System Science (Sapporo, Japan) or Fasmac (Atsugi, Japan). All reagents used for preparing buffer solutions were purchased from Wako Chemicals (Osaka, Japan) with the following exceptions: 4-(2-hydroxyethyl)-1-piperazineethanesulfonic acid (HEPES), 3-[4-(2-hydroxyethyl)-1-piperazinyl]propanesulfonic acid (HEPPS), and the disodium salt of ethylenediaminetetraacetic acid (Na2EDTA) were purchased from Dojindo (Kumamoto, Japan); PEG with an average molecular weight of 8 × 103 was purchased from MP Biomedicals (Tokyo, Japan); 2-methoxyethanol and 1,2-dimethoxyethane were purchased from TCI (Tokyo, Japan); and dextran with an average molecular weight of 1 × 104 from Sigma-Aldrich (St. Louis, MO, USA). Values of solution properties (relative dielectric constant, water activity, and viscosity) were determined, as described previously [44].
4.2. Kinetic Studies on the Ribozymes and DNAzymes
Kinetics of the reactions catalyzed by hammerhead ribozymes and 17E DNAzymes were measured in buffer solutions comprising 50 mM HEPES, 50 mM NaCl, and 0.1 mM Na2EDTA. The pH of this solution was adjusted to 7.0 at 37 °C. Kinetics of the reactions catalyzed by R3C ligase ribozymes and 9DB1 DNAzymes were measured in buffer solutions comprising 50 mM HEPPS, 50 mM NaCl, and 0.1 mM Na2EDTA. The pH of this solution was adjusted to 8.0 at 37 °C. Mixed aqueous solutions were prepared by adding a cosolvent or cosolute at a concentration of 20% by weight, unless otherwise stated.
Substrate strands were fluorescently-labeled either with carboxyfluorescein (FAM) or carboxytetramethylrhodamine (TAMRA) at the 5′-end. For single-turnover reactions, substrate cleavage was measured using a 2 μM concentration of ribozyme (or DNAzyme) and a 0.1 μM concentration of substrate, and substrate ligation was measured using a 1 μM concentration of ribozyme (or DNAzyme) and a 0.1 μM concentration of substrate. For multiple-turnover reactions, substrate cleavage was measured using a 5 nM concentration of ribozyme (or DNAzyme) and a 0.1 μM concentration of substrate, and substrate ligation was measured using a 10 nM concentration of ribozyme (or DNAzyme) and a 0.1 μM concentration of substrate or a 0.1 μM concentration of fluorescently-labeled 5′-fragment substrate and a 1 μM concentration of non-labeled 3′-fragment substrate to be used for the ligation of two substrate RNAs. Before initiating the reaction by adding MgCl2 at 37 °C, the enzyme and substrate(s) were annealed in the buffer solution at 60 °C, in the case of the nuclease enzymes, or 80 °C, in the case of the ligase enzymes. The reactions were quenched by mixing the reaction mixture with a 90% formamide solution containing 100 mM Na2EDTA. The quenched solutions were then loaded onto a 20% polyacrylamide gel containing 7 M urea. After gel electrophoresis, the fluorescence emission from fluorescently-labeled strands was quantified using a fluorescence scanner (FLA-7000, Fujifilm, Tokyo, Japan).
The rate constant (k) for substrate cleavage or ligation was determined from a plot of the fraction of the reaction product against time, fitted to a single-exponential equation f = f0 + (fmax − f0){1 − exp(−kt)}, where f0 is the fraction at time zero and fmax is the fraction at the end of the reaction. A linear approximation was used to calculate the k value of very slow reactions.
4.3. Thermal Stability Measurements
Thermal melting curve of a DNA structure was obtained by monitoring absorption at 260 nm using a spectrophotometer (UV1800, Shimadzu, Kyoto, Japan) equipped with a temperature controller. Oligonucleotides at a strand concentration of 2 μM were prepared using a buffer solution comprising 50 mM HEPES, 50 mM NaCl, 10 mM MgCl2, and 0.1 mM Na2EDTA. The pH of this solution was adjusted to 7.0. Mixed solutions were prepared at a concentration of organic additives of 20% by weight, unless otherwise stated. The solutions were placed in a cuvette sealed with an adhesive sheet. After annealing at a cooling rate of 2 °C min−1, the melting curve was measured at a heating rate of 0.5 °C min−1. The value of Tm, defined as the temperature at which half of the structure was denatured, and ΔG° at 37 °C were determined from the shape of the melting curve, as described previously [43].
5. Conclusions
The molecular environment of mixed aqueous solutions that contain organic compounds can enhance the catalytic activity of the hammerhead ribozyme, 17E DNAzyme, R3C ribozyme, and 9DB1 DNAzyme. In particular, the addition of large PEGs to aqueous reaction media effectively increased the rate of the reactions catalyzed by these enzymes, but systematic investigation using different kinds of mixed solutions and correlation analyses revealed that the effect of PEGs on the catalytic reaction rates had different origins with different enzymes. The activity of the hammerhead ribozyme was enhanced by a decrease in dielectric constant of the medium, because such decrease led to stronger Mg2+ binding interactions with the ribozyme. The activity of the 17E DNAzyme increased in the presence of large-sized additives, as these compounds caused an increase in the stability of DNA interactions. Although destabilization of base pairing between the enzyme and substrate was expected to beneficial for the catalytic turnover, the mixed solutions did not increase the turnover rate of the hammerhead ribozyme and 17E DNAzyme having extended base pairing. The ribozyme and DNAzyme ligases evaluated in this study also showed no significant turnover activity, but their single-turnover rates increased in many mixed solutions with respect to the solution containing no organic additives. In this case, however, the effects did not seem to be determined by a single environmental factor. These results suggest that the mixed solutions can be widely used to enhance the activity of nucleic acid enzymes, such as by modulating the physical properties of a solution and the stability of nucleic acid interactions. The correlation analysis data appears to be a useful tool for predicting the effect of mixed solutions, including those not evaluated in this study, on the activity of these enzymes. These findings provide insight into the catalytic activity of nucleic acid enzymes under various conditions that are applicable to the medical and technology fields, for example in living cells and in biosensors.
Supplementary Materials
The following are available online at www.mdpi.com/2073-4344/7/12/355/s1, Table S1: Increments of the single-turnover rates of the 17E(S)-catalyzed reaction, Figure S1: Correlation plots of the turnover rates of the HH(S)-catalyzed reaction, Figure S2: Correlation plots of the turnover rates of the 17E(S)-catalyzed reaction, and the increments of the single-turnover rates of the reactions catalyzed by 17E(S) and 17E(L).
Acknowledgments
We thank Junpei Ueno for technical assistance. This work was supported in part by Grants-in-Aid for Scientific Research from Japan Society for the Promotion of Science (JSPS) (24550200 and 15K05575) and MEXT-Supported Program for the Strategic Research Foundation at Private Universities (2009–2014).
Author Contributions
S.N. conceived and designed the experiments; S.N., M.H., and M.K. performed the experiments; S.N., M.H., and M.K. analyzed the data; S.N. and N.S. contributed reagents and materials tools; S.N. wrote the paper.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Jäschke, A.; Seelig, B. Evolution of DNA and RNA as catalysts for chemical reactions. Curr. Opin. Chem. Biol. 2000, 4, 257–262. [Google Scholar]
- Citti, L.; Rainaldi, G. Synthetic hammerhead ribozymes as therapeutic tools to control disease genes. Curr. Gene Ther. 2005, 5, 11–24. [Google Scholar] [CrossRef] [PubMed]
- Fokina, A.A.; Stetsenko, D.A.; Francois, J.C. DNA enzymes as potential therapeutics: Towards clinical application of 10–23 DNAzymes. Expert Opin. Biol. Ther. 2015, 15, 689–711. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Lu, Y. A highly sensitive and selective catalytic DNA biosensor for lead ions. J. Am. Chem. Soc. 2001, 122, 10466–10467. [Google Scholar] [CrossRef]
- Rueda, D.; Walter, N.G. Fluorescent energy transfer readout of an aptazyme-based biosensor. Methods Mol. Biol. 2006, 335, 289–310. [Google Scholar] [PubMed]
- Li, S.; Nosrati, M.; Kashani-Sabet, M. Knockdown of telomerase RNA using hammerhead ribozymes and RNA interference. Methods Mol. Biol. 2008, 405, 113–131. [Google Scholar]
- Liu, J.; Cao, Z.; Lu, Y. Functional nucleic acid sensors. Chem. Rev. 2009, 109, 1948–1998. [Google Scholar] [CrossRef] [PubMed]
- Lan, T.; Lu, Y. Metal ion-dependent DNAzymes and their applications as biosensors. Met. Ions Life Sci. 2012, 10, 217–248. [Google Scholar] [PubMed]
- Gong, L.; Zhao, Z.; Lv, Y.F.; Huan, S.Y.; Fu, T.; Zhang, X.B.; Shen, G.L.; Yu, R.Q. DNAzyme-based biosensors and nanodevices. Chem. Commun. 2015, 51, 979–995. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Lu, Y. Adenosine-dependent assembly of aptazyme-functionalized gold nanoparticles and its application as a colorimetric biosensor. Anal. Chem. 2004, 76, 1627–1632. [Google Scholar] [CrossRef] [PubMed]
- Win, M.N.; Smolke, C.D. A modular and extensible RNA-based gene-regulatory platform for engineering cellular function. Proc. Natl. Acad. Sci. USA 2007, 104, 14283–14288. [Google Scholar] [CrossRef] [PubMed]
- Sun, C.; Liu, X.; Feng, K.; Jiang, J.; Shen, G.; Yu, R. An aptazyme-based electrochemical biosensor for the detection of adenosine. Anal. Chim. Acta 2010, 669, 87–93. [Google Scholar] [CrossRef] [PubMed]
- Auslander, S.; Ketzer, P.; Hartig, J.S. A ligand-dependent hammerhead ribozyme switch for controlling mammalian gene expression. Mol. Biosyst. 2010, 6, 807–814. [Google Scholar] [CrossRef] [PubMed]
- Bevilacqua, P.C.; Brown, T.S.; Nakano, S.; Yajima, R. Catalytic roles for proton transfer and protonation in ribozymes. Biopolymers 2004, 73, 90–109. [Google Scholar] [CrossRef] [PubMed]
- Doudna, J.A.; Lorsch, J.R. Ribozyme catalysis: Not different, just worse. Nat. Struct. Mol. Biol. 2005, 12, 395–402. [Google Scholar] [CrossRef] [PubMed]
- Lilley, D.M. Structure, folding and mechanisms of ribozymes. Curr. Opin. Struct. Biol. 2005, 15, 313–323. [Google Scholar] [CrossRef] [PubMed]
- Wedekind, J.E. Metal ion binding and function in natural and artificial small RNA enzymes from a structural perspective. Met. Ions Life Sci. 2011, 9, 299–345. [Google Scholar] [PubMed]
- Auffinger, P.; Grover, N.; Westhof, E. Metal ion binding to RNA. Met. Ions Life Sci. 2011, 9, 1–35. [Google Scholar] [PubMed]
- Ward, W.L.; Plakos, K.; DeRose, V.J. Nucleic acid catalysis: Metals, nucleobases, and other cofactors. Chem. Rev. 2014, 114, 4318–4342. [Google Scholar] [CrossRef] [PubMed]
- Nakano, S.; Miyoshi, D.; Sugimoto, N. Effects of molecular crowding on the structures, interactions, and functions of nucleic acids. Chem. Rev. 2014, 114, 2733–2758. [Google Scholar] [CrossRef] [PubMed]
- Winship, P.R. An improved method for directly sequencing PCR amplified material using dimethyl sulphoxide. Nucl. Acids Res. 1989, 17, 1266. [Google Scholar] [CrossRef] [PubMed]
- Jensen, M.A.; Fukushima, M.; Davis, R.W. DMSO and betaine greatly improve amplification of GC-rich constructs in de novo synthesis. PLoS ONE 2010, 5, e11024. [Google Scholar] [CrossRef] [PubMed]
- Dave, N.; Liu, J. Fast molecular beacon hybridization in organic solvents with improved target specificity. J. Phys. Chem. B 2010, 114, 15694–15699. [Google Scholar] [CrossRef] [PubMed]
- Zhang, T.; Shang, C.; Duan, R.; Hakeem, A.; Zhang, Z.; Lou, X.; Xia, F. Polar organic solvents accelerate the rate of DNA strand replacement reaction. Analyst 2015, 140, 2023–2028. [Google Scholar] [CrossRef] [PubMed]
- Abe, H.; Abe, N.; Shibata, A.; Ito, K.; Tanaka, Y.; Ito, M.; Saneyoshi, H.; Shuto, S.; Ito, Y. Structure formation and catalytic activity of DNA dissolved in organic solvents. Angew. Chem. Int. Ed. Engl. 2012, 51, 6475–6479. [Google Scholar] [CrossRef] [PubMed]
- Behera, A.K.; Schlund, K.J.; Mason, A.J.; Alila, K.O.; Han, M.; Grout, R.L.; Baum, D.A. Enhanced deoxyribozyme-catalyzed RNA ligation in the presence of organic cosolvents. Biopolymers 2013, 99, 382–391. [Google Scholar] [CrossRef] [PubMed]
- Zhou, W.; Saran, R.; Chen, Q.; Ding, J.; Liu, J. A new Na+-dependent RNA-cleaving DNAzyme with over 1000-fold rate acceleration by ethanol. ChemBioChem 2016, 17, 159–163. [Google Scholar] [CrossRef] [PubMed]
- Yu, T.; Zhou, W.; Liu, J. Ultrasensitive DNAzyme-based Ca2+ detection boosted by ethanol and a solvent compatible scaffold for aptazyme design. ChemBioChem 2017. [Google Scholar] [CrossRef] [PubMed]
- Leamy, K.A.; Assmann, S.M.; Mathews, D.H.; Bevilacqua, P.C. Bridging the gap between in vitro and in vivo RNA folding. Q. Rev. Biophys. 2016, 49, e10. [Google Scholar] [CrossRef] [PubMed]
- Nakano, S.; Sugimoto, N. Model studies of the effects of intracellular crowding on nucleic acid interactions. Mol. Biosyst. 2016, 13, 32–41. [Google Scholar] [CrossRef] [PubMed]
- Nakano, S.; Sugimoto, N. The structural stability and catalytic activity of DNA and RNA oligonucleotides in the presence of organic solvents. Biophys. Rev. 2016, 8, 11–23. [Google Scholar] [CrossRef] [PubMed]
- Osborne, E.M.; Schaak, J.E.; DeRose, V.J. Characterization of a native hammerhead ribozyme derived from Schistosomes. RNA 2005, 11, 187–196. [Google Scholar] [CrossRef] [PubMed]
- Nelson, J.A.; Uhlenbeck, O.C. Minimal and extended hammerheads utilize a similar dynamic reaction mechanism for catalysis. RNA 2008, 14, 43–54. [Google Scholar] [CrossRef] [PubMed]
- Nakano, S.; Karimata, H.T.; Kitagawa, Y.; Sugimoto, N. Facilitation of RNA enzyme activity in the molecular crowding media of cosolutes. J. Am. Chem. Soc. 2009, 131, 16881–16888. [Google Scholar] [CrossRef] [PubMed]
- Nakano, S.; Kitagawa, Y.; Yamashita, H.; Miyoshi, D.; Sugimoto, N. Effects of cosolvents on the folding and catalytic activities of the hammerhead ribozyme. ChemBioChem 2015, 16, 1803–1810. [Google Scholar] [CrossRef] [PubMed]
- Santoro, S.W.; Joyce, G.F. A general purpose RNA-cleaving DNA enzyme. Proc. Natl. Acad. Sci. USA 1997, 94, 4262–4266. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Zheng, W.; Kwon, A.H.; Lu, Y. In vitro selection and characterization of a highly efficient Zn(II)-dependent RNA-cleaving deoxyribozyme. Nucl. Acids Res. 2000, 28, 481–488. [Google Scholar] [CrossRef] [PubMed]
- Hwang, K.; Wu, P.; Kim, T.; Lei, L.; Tian, S.; Wang, Y.; Lu, Y. Photocaged DNAzymes as a general method for sensing metal ions in living cells. Angew. Chem. Int. Ed. Engl. 2014, 53, 13798–13802. [Google Scholar] [CrossRef] [PubMed]
- Zhou, W.; Chen, Q.; Huang, P.J.; Ding, J.; Liu, J. DNAzyme hybridization, cleavage, degradation, and sensing in undiluted human blood serum. Anal. Chem. 2015, 87, 4001–4007. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.Y.; Niu, C.G.; Guo, L.J.; Hu, L.Y.; Wu, S.Q.; Zeng, G.M.; Li, F. A fluorescence sensor for lead(II) ions determination based on label-free gold nanoparticles (GNPs)-DNAzyme using time-gated mode in aqueous solution. J. Fluoresc. 2017, 27, 643–649. [Google Scholar] [CrossRef] [PubMed]
- Nakano, S.; Karimata, H.; Ohmichi, T.; Kawakami, J.; Sugimoto, N. The effect of molecular crowding with nucleotide length and cosolute structure on DNA duplex stability. J. Am. Chem. Soc. 2004, 126, 14330–14331. [Google Scholar] [CrossRef] [PubMed]
- Knowles, D.B.; LaCroix, A.S.; Deines, N.F.; Shkel, I.; Record, M.T., Jr. Separation of preferential interaction and excluded volume effects on DNA duplex and hairpin stability. Proc. Natl. Acad. Sci. USA 2011, 108, 12699–12704. [Google Scholar] [CrossRef] [PubMed]
- Nakano, S.; Yamaguchi, D.; Tateishi-Karimata, H.; Miyoshi, D.; Sugimoto, N. Hydration changes upon DNA folding studied by osmotic stress experiments. Biophys. J. 2012, 102, 2808–2817. [Google Scholar] [CrossRef] [PubMed]
- Nakano, S.; Kitagawa, Y.; Miyoshi, D.; Sugimoto, N. Hammerhead ribozyme activity and oligonucleotide duplex stability in mixed solutions of water and organic compounds. FEBS Open Bio 2014, 4, 643–650. [Google Scholar] [CrossRef] [PubMed]
- Rogers, J.; Joyce, G.F. The effect of cytidine on the structure and function of an RNA ligase ribozyme. RNA 2001, 7, 395–404. [Google Scholar] [CrossRef] [PubMed]
- Kurihara, E.; Uchida, S.; Umehara, T.; Tamura, K. Development of a functionally minimized mutant of the R3C ligase ribozyme offers insight into the plausibility of the RNA world hypothesis. Biology 2014, 3, 452–465. [Google Scholar] [CrossRef] [PubMed]
- Purtha, W.E.; Coppins, R.L.; Smalley, M.K.; Silverman, S.K. General deoxyribozyme-catalyzed synthesis of native 3′-5′ RNA linkages. J. Am. Chem. Soc. 2005, 127, 13124–13125. [Google Scholar] [CrossRef] [PubMed]
- Bevers, S.; Xiang, G.; McLaughlin, L.W. Importance of specific adenosine N3-nitrogens for efficient cleavage by a hammerhead ribozyme. Biochemistry 1996, 35, 6483–6490. [Google Scholar] [CrossRef] [PubMed]
- Lee, T.S.; Lopez, C.S.; Giambasu, G.M.; Martick, M.; Scott, W.G.; York, D.M. Role of Mg2+ in hammerhead ribozyme catalysis from molecular simulation. J. Am. Chem. Soc. 2008, 130, 3053–3064. [Google Scholar] [CrossRef] [PubMed]
- Asami, K.; Hanai, T.; Koizumi, N. Dielectric properties of yeast cells. J. Membr. Biol. 1976, 28, 169–180. [Google Scholar] [CrossRef] [PubMed]
- Cuervo, A.; Dans, P.D.; Carrascosa, J.L.; Orozco, M.; Gomila, G.; Fumagalli, L. Direct measurement of the dielectric polarization properties of DNA. Proc. Natl. Acad. Sci. USA 2014, 111, E3624–E3630. [Google Scholar] [CrossRef] [PubMed]
- Grubbs, R.D. Intracellular magnesium and magnesium buffering. Biometals 2002, 15, 251–259. [Google Scholar] [CrossRef] [PubMed]
- Raz, M.H.; Hollenstein, M. Probing the effect of minor groove interactions on the catalytic efficiency of DNAzymes 8–17 and 10–23. Mol. Biosyst. 2015, 11, 1454–1461. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.K.; Rasnik, I.; Liu, J.; Ha, T.; Lu, Y. Dissecting metal ion-dependent folding and catalysis of a single DNAzyme. Nat. Chem. Biol. 2007, 3, 763–768. [Google Scholar] [CrossRef] [PubMed]
- Nakano, S.; Watabe, T.; Sugimoto, N. Modulation of the ribozyme and deoxyribozyme activities using tetraalkylammonium ions. ChemPhysChem 2017. [Google Scholar] [CrossRef] [PubMed]
- Ponce-Salvatierra, A.; Wawrzyniak-Turek, K.; Steuerwald, U.; Hobartner, C.; Pena, V. Crystal structure of a DNA catalyst. Nature 2016, 529, 231–234. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
(a) Structures of the hammerhead ribozymes and their substrates used in this study. The cleavage sites are indicated by arrowheads. (b) Kinetic traces for substrate cleavage catalyzed by HH(L) under single-turnover conditions in the presence of 10 mM MgCl2 and in the absence (black circles) and presence of PEG8000 (red squares) or PEG200 (blue triangles). The data relative to the first 5 min of the reaction are enlarged in the inset. (c) Kinetic traces for substrate cleavage catalyzed by HH(S) (closed symbols) and HH(L) (open symbols) under multiple-turnover conditions in the presence of 10 mM MgCl2 and in the absence (black circles) and presence of PEG8000 (red squares) or PEG200 (blue triangles). (d) Increments of the turnover rate of the reactions catalyzed by HH(S) (closed symbols) or HH(L) (open symbols) in the presence of 10 mM MgCl2 plotted against the amount of PEG8000 (red squares) or PEG200 (blue triangles) in solution. (e) Plot of the increments of the turnover rate of the HH(S)-catalyzed reaction in the presence of 10 mM MgCl2 against the inverse of the relative dielectric constant of solutions. The data obtained using 10% or 30% PEG solutions (PEG8000 and PEG200) are also included (triangles). The correlation coefficient of a linear fit is 0.94.
Figure 1.
(a) Structures of the hammerhead ribozymes and their substrates used in this study. The cleavage sites are indicated by arrowheads. (b) Kinetic traces for substrate cleavage catalyzed by HH(L) under single-turnover conditions in the presence of 10 mM MgCl2 and in the absence (black circles) and presence of PEG8000 (red squares) or PEG200 (blue triangles). The data relative to the first 5 min of the reaction are enlarged in the inset. (c) Kinetic traces for substrate cleavage catalyzed by HH(S) (closed symbols) and HH(L) (open symbols) under multiple-turnover conditions in the presence of 10 mM MgCl2 and in the absence (black circles) and presence of PEG8000 (red squares) or PEG200 (blue triangles). (d) Increments of the turnover rate of the reactions catalyzed by HH(S) (closed symbols) or HH(L) (open symbols) in the presence of 10 mM MgCl2 plotted against the amount of PEG8000 (red squares) or PEG200 (blue triangles) in solution. (e) Plot of the increments of the turnover rate of the HH(S)-catalyzed reaction in the presence of 10 mM MgCl2 against the inverse of the relative dielectric constant of solutions. The data obtained using 10% or 30% PEG solutions (PEG8000 and PEG200) are also included (triangles). The correlation coefficient of a linear fit is 0.94.
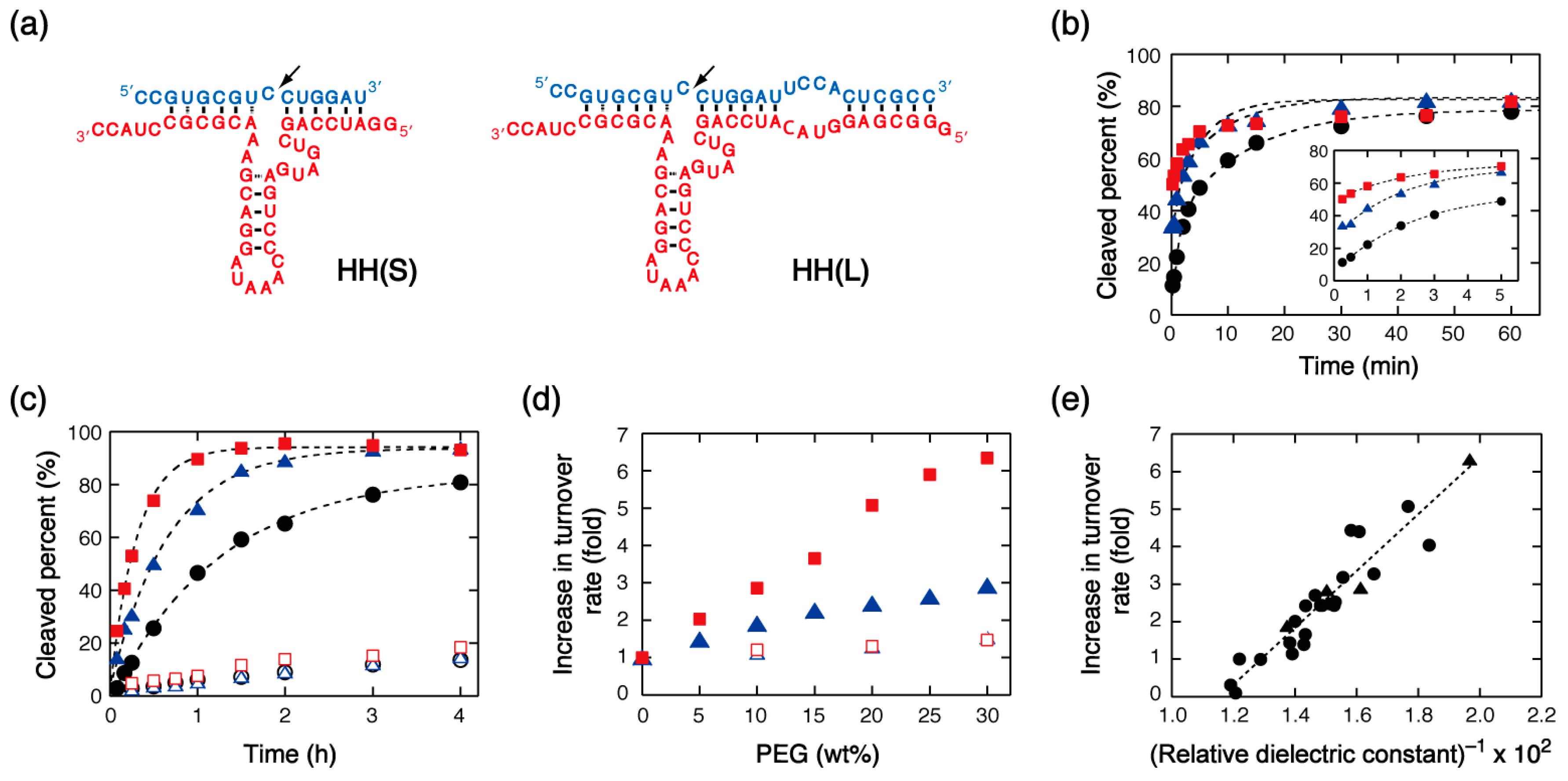
Figure 2.
(a) Structures of the 17E DNAzymes and their substrates used in this study. The cleavage sites are indicated by arrowheads. (b) Kinetic traces for substrate cleavage catalyzed by 17E(L) under single-turnover conditions in the presence of 10 mM MgCl2 and in the absence (black circles) and presence of PEG8000 (red squares) or PEG200 (blue triangles). (c) Kinetic traces for substrate cleavage catalyzed by 17E(S) (closed symbols) and 17E(L) (open symbols) under multiple-turnover conditions in the presence of 10 mM MgCl2 and in the absence (black circles) and presence of PEG8000 (red squares) or PEG200 (blue triangles). (d) Tm values of the DNA structure presented in the inset evaluated in solutions containing 10 mM MgCl2 (black) or 1 mM MgCl2 (gray), with and without EG, PEGs, or Dex. (e) Plot of the increments of the turnover rate of the 17E(S)-catalyzed reaction against ΔTm in the presence of 10 mM MgCl2 (black) or 1 mM MgCl2 (gray), excluding the data for reactions characterized by a very slow rate. The data obtained using 10% or 30% PEG solutions (PEG8000 and PEG200) and EtOH solution are also included (triangles). The correlation coefficient of a linear fit is 0.97.
Figure 2.
(a) Structures of the 17E DNAzymes and their substrates used in this study. The cleavage sites are indicated by arrowheads. (b) Kinetic traces for substrate cleavage catalyzed by 17E(L) under single-turnover conditions in the presence of 10 mM MgCl2 and in the absence (black circles) and presence of PEG8000 (red squares) or PEG200 (blue triangles). (c) Kinetic traces for substrate cleavage catalyzed by 17E(S) (closed symbols) and 17E(L) (open symbols) under multiple-turnover conditions in the presence of 10 mM MgCl2 and in the absence (black circles) and presence of PEG8000 (red squares) or PEG200 (blue triangles). (d) Tm values of the DNA structure presented in the inset evaluated in solutions containing 10 mM MgCl2 (black) or 1 mM MgCl2 (gray), with and without EG, PEGs, or Dex. (e) Plot of the increments of the turnover rate of the 17E(S)-catalyzed reaction against ΔTm in the presence of 10 mM MgCl2 (black) or 1 mM MgCl2 (gray), excluding the data for reactions characterized by a very slow rate. The data obtained using 10% or 30% PEG solutions (PEG8000 and PEG200) and EtOH solution are also included (triangles). The correlation coefficient of a linear fit is 0.97.
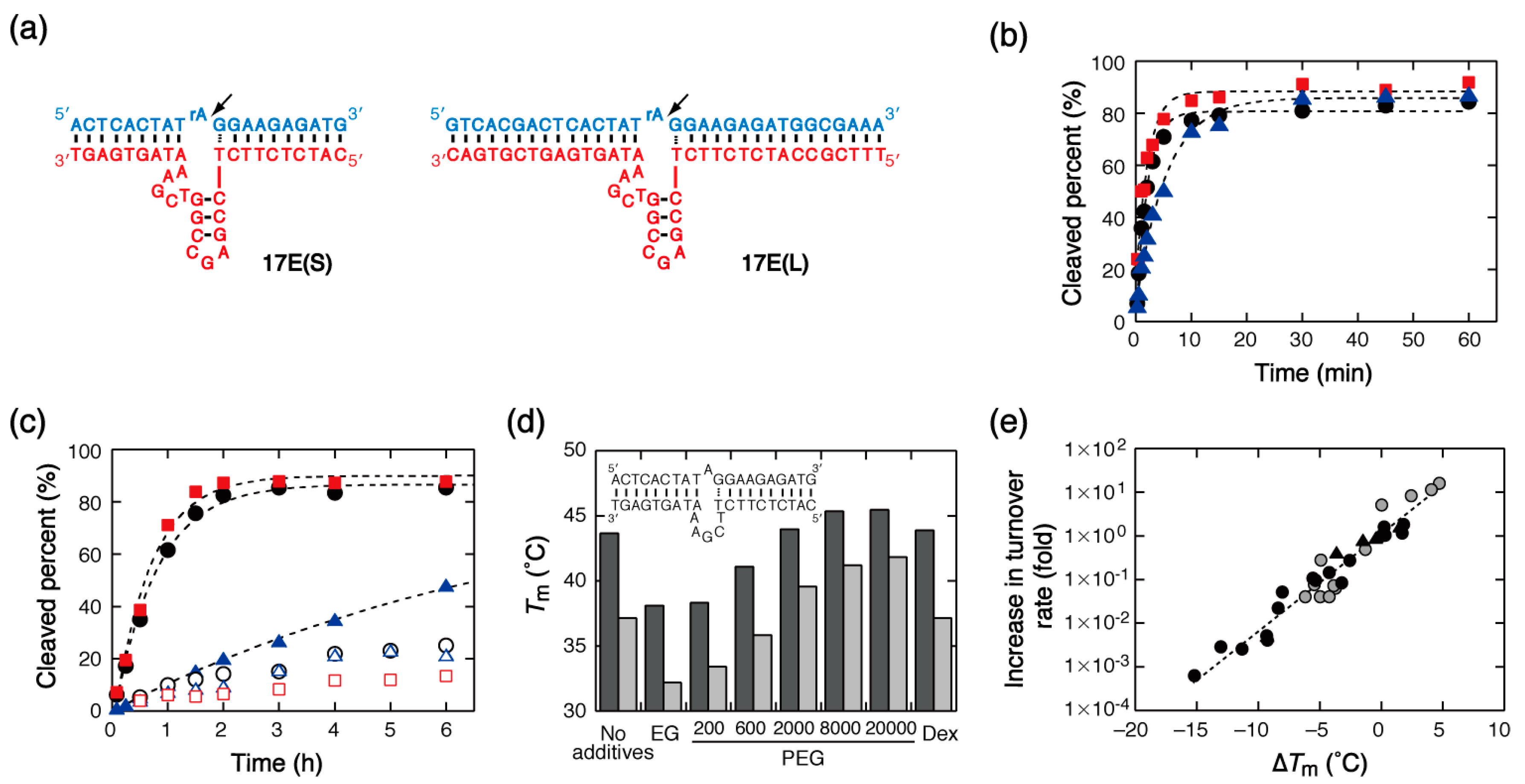
Figure 3.
(a) Structures of the ligase ribozymes and DNAzyme and their substrates used in this study. The ligation sites are indicated by arrowheads. (b) Kinetic traces for substrate ligation catalyzed by R3C(H) under single-turnover conditions in the presence of 10 mM MgCl2 and in the absence (black circles) and presence of PEG8000 (red squares) or PEG200 (blue triangles). (c) Kinetic traces for substrate ligation catalyzed by 9DB1 under single-turnover conditions in the presence of 10 mM MgCl2 and in the absence (black circles) and presence of PEG8000 (red squares) or PEG200 (blue triangles). (d) Kinetic traces for substrate ligation catalyzed by R3C(T) (closed symbols) and 9DB1 (open symbols) under multiple-turnover conditions in the presence of 10 mM MgCl2 and in the absence (black circles) and presence of PEG8000 (red squares) or PEG200 (blue triangles).
Figure 3.
(a) Structures of the ligase ribozymes and DNAzyme and their substrates used in this study. The ligation sites are indicated by arrowheads. (b) Kinetic traces for substrate ligation catalyzed by R3C(H) under single-turnover conditions in the presence of 10 mM MgCl2 and in the absence (black circles) and presence of PEG8000 (red squares) or PEG200 (blue triangles). (c) Kinetic traces for substrate ligation catalyzed by 9DB1 under single-turnover conditions in the presence of 10 mM MgCl2 and in the absence (black circles) and presence of PEG8000 (red squares) or PEG200 (blue triangles). (d) Kinetic traces for substrate ligation catalyzed by R3C(T) (closed symbols) and 9DB1 (open symbols) under multiple-turnover conditions in the presence of 10 mM MgCl2 and in the absence (black circles) and presence of PEG8000 (red squares) or PEG200 (blue triangles).

{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Fold increase in the rate of the HH(S)-catalyzed turnover reaction in mixed solutions over the rate in the absence of organic additives.
Table 1.
Fold increase in the rate of the HH(S)-catalyzed turnover reaction in mixed solutions over the rate in the absence of organic additives.
| Solution 1 | 10 mM MgCl2 | 1 mM MgCl2 | Solution 1 | 10 mM MgCl2 | 1 mM MgCl2 | Solution 1 | 10 mM MgCl2 | 1 mM MgCl2 |
|---|---|---|---|---|---|---|---|---|
| PEG20000 | 3.4 | 13 | PDO | 1.7 | 2.8 | FA | 0.32 | <0.02 |
| PEG8000 | 5.1 | 17 | MME | 2.4 | 3.9 | DMF | 1.3 | <0.02 |
| PEG2000 | 3.3 | 10 | DME | 4.4 | 11 | AcAm | 1.0 | <0.02 |
| PEG600 | 2.5 | 6.8 | MeOH | 2.4 | 4.9 | AcCN | 2.7 | 6.5 |
| PEG200 | 2.4 | 5.5 | EtOH | 2.5 | 4.8 | DMSO | 1.4 | 1.2 |
| EG | 2.0 | 1.3 | PrOH | 3.2 | 6.0 | DOX | 1.9 | 4.6 |
| Glyc | 1.1 | 1.3 | Urea | 0.10 | <0.02 | Dex | 1.4 | 2.5 |
1 Abbreviations: PEG (polyethylene glycol with an average molecular weight ranging from 2 × 104 to 2 × 102); EG (ethylene glycol); Glyc (glycerol); POD (1,3-propanediol); MME (2-methoxyethanol); DME (1,2-dimethoxyethane); MeOH (methanol); EtOH (ethanol); PrOH (1-propanol); FA (formamide); DMF (N,N-dimethylformamide); AcAm (acetamide); AcCN (acetonitrile); DMSO (dimethyl sulfoxide); DOX (1,4-dioxane); and Dex (dextran with an average molecular weight of 1 × 104).
Table 2.
Fold increase in the rate of the 17E(S)-catalyzed turnover reaction in mixed solutions over the rate in the absence of organic additives.
Table 2.
Fold increase in the rate of the 17E(S)-catalyzed turnover reaction in mixed solutions over the rate in the absence of organic additives.
| Solution | 10 mM MgCl2 | 1 mM MgCl2 | Solution | 10 mM MgCl2 | 1 mM MgCl2 | Solution | 10 mM MgCl2 | 1 mM MgCl2 |
|---|---|---|---|---|---|---|---|---|
| PEG20000 | 1.8 | 16 | PDO | 0.14 | 0.073 | FA | <0.001 | <0.03 |
| PEG8000 | 1.2 | 11 | MME | 0.0041 | <0.03 | DMF | <0.001 | <0.03 |
| PEG2000 | 1.0 | 8.2 | DME | 0.052 | 0.28 | AcAm | <0.001 | <0.03 |
| PEG600 | 0.27 | 0.49 | MeOH | 0.022 | 0.076 | AcCN | <0.001 | <0.03 |
| PEG200 | 0.095 | 0.064 | EtOH | 0.0026 | 0.041 | DMSO | 0.0051 | <0.03 |
| EG | 0.11 | 0.040 | PrOH | <0.001 | <0.03 | DOX | 0.0029 | <0.03 |
| Glyc | 0.083 | 0.040 | Urea | <0.001 | <0.03 | Dex | 1.6 | 5.1 |
Table 3.
Fold increase in the rates of the R3C(H)-catalyzed and 9DB1-catalyzed single-turnover reactions in mixed solutions in the presence of 10 mM MgCl2 over the rates in the absence of organic additives.
Table 3.
Fold increase in the rates of the R3C(H)-catalyzed and 9DB1-catalyzed single-turnover reactions in mixed solutions in the presence of 10 mM MgCl2 over the rates in the absence of organic additives.
| Solution | R3C(H) | 9DB1 | Solution | R3C(H) | 9DB1 | Solution | R3C(H) | 9DB1 |
|---|---|---|---|---|---|---|---|---|
| PEG20000 | 1.6 | 4.2 | PDO | 1.2 | 2.6 | FA | <0.001 | <0.001 |
| PEG8000 | 2.0 | 4.6 | MME | 1.9 | 3.3 | DMF | 0.26 | <0.001 |
| PEG2000 | 1.6 | 6.8 | DME | 1.2 | 1.9 | AcAm | 0.30 | <0.001 |
| PEG600 | 1.3 | 4.2 | MeOH | 1.5 | 4.5 | AcCN | 1.8 | 4.5 |
| PEG200 | 2.0 | 4.1 | EtOH | 2.2 | 2.4 | DMSO | 1.7 | 2.5 |
| EG | 1.0 | 2.8 | PrOH | 1.8 | 3.9 | DOX | 1.3 | 3.8 |
| Glyc | 1.2 | 1.1 | Urea | <0.001 | <0.001 | Dex | 1.3 | 0.89 |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Nakano, S.-i.; Horita, M.; Kobayashi, M.; Sugimoto, N. Catalytic Activities of Ribozymes and DNAzymes in Water and Mixed Aqueous Media. Catalysts 2017, 7, 355. https://doi.org/10.3390/catal7120355
AMA Style
Nakano S-i, Horita M, Kobayashi M, Sugimoto N. Catalytic Activities of Ribozymes and DNAzymes in Water and Mixed Aqueous Media. Catalysts. 2017; 7(12):355. https://doi.org/10.3390/catal7120355
Chicago/Turabian StyleNakano, Shu-ichi, Masao Horita, Miku Kobayashi, and Naoki Sugimoto. 2017. "Catalytic Activities of Ribozymes and DNAzymes in Water and Mixed Aqueous Media" Catalysts 7, no. 12: 355. https://doi.org/10.3390/catal7120355
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.