Enzymatic Synthesis of S-Adenosylmethionine Using Immobilized Methionine Adenosyltransferase Variants on the 50-mM Scale

School of Life Sciences, Northwestern Polytechnical University, Xi’an 710072, China
*
Author to whom correspondence should be addressed.
Catalysts 2017, 7(8), 238; https://doi.org/10.3390/catal7080238
Submission received: 17 July 2017
/
Revised: 13 August 2017
/
Accepted: 14 August 2017
/
Published: 17 August 2017
(This article belongs to the Special Issue Biocatalysis and Biotransformations)
Abstract
:S-adenosylmethionine (SAM), an important metabolite in all living organisms, has been widely used to treat various diseases. To develop a simple and efficient method to produce SAM, an engineered variant of the methionine adenosyltransferase (MAT) from Escherichia coli was investigated for its potential use in the enzymatic synthesis of SAM due to its significantly decreased product inhibition. The recombinant I303V MAT variant was successfully produced at a high level (~800 mg/L) with approximately four-fold higher specific activity than the wild-type MAT. The recombinant I303V MAT was covalently immobilized onto the amino resin and epoxy resin in order to obtain a robust biocatalyst to be used in industrial bioreactors. The immobilized preparation using amino resin exhibited the highest activity coupling yield (~84%), compared with approximately 3% for epoxy resin. The immobilized enzyme was more stable than the soluble enzyme under the reactive conditions, with a half-life of 229.5 h at 37 °C. The KmATP value (0.18 mM) of the immobilized enzyme was ca. two-fold lower than that of the soluble enzyme. Furthermore, the immobilized enzyme showed high operational stability during 10 consecutive 8 h batches, with the substrate adenosine triphosphate (ATP) conversion rate above 95% on the 50-mM scale.
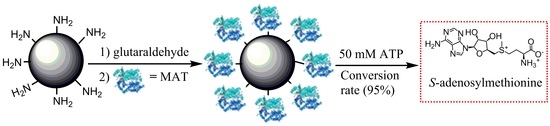
1. Introduction
S-adenosylmethionine (SAM), which is synthesized in vivo from adenosine triphosphate (ATP) and l-methionine by methionine adenosyltransferase (MAT, EC2.5.1.6), is an important metabolic intermediate in all living organisms because it acts as a methyl group donor in transmethylation reactions of nucleic acids, proteins, polysaccharides, phospholipids, and fatty acids, and as a precursor molecule in the transsulfuration pathway [1,2]. In clinical practice, SAM is widely used in treating alcoholic liver disease, depression and osteoarthritis [3,4,5,6]. However, the very high price of SAM might be a limitation for its wide application in the field of medicine. Thus, there has been increasing interest in developing an efficient and low-cost mean of producing SAM on the industrial scale.
At present, SAM is commercially prepared through the extraction of yeast cells that harbor high MAT activity after their cultivation in medium that is supplemented with l-methionine [7,8]. Most studies have been focused on the improvement of SAM production by altering Saccharomyces cerevisiae or Pichia pastoris strains by a genetic engineering approach or by optimization of culture conditions [9,10,11,12,13,14,15,16]. However, the production of SAM by yeast fermentation has some obvious drawbacks, such as a long fermentation period, low SAM content and yield, and low conversion rate of the substrate. Furthermore, the extraction of SAM from yeast cells is complicated, since the yeast cells have to be disrupted first and SAM is purified from the many other cell components [17]. In contrast, enzymatic synthesis of SAM using MAT appears quite attractive. Enzymatic production of SAM in vitro has many advantages over yeast fermentation methods, including a short reaction time (~8 h), a high conversion efficiency, a simple purification, and being environment friendly. Additionally, immobilization of MAT could improve the operational stability of the enzyme and reduce the cost of the relevant process at the same time. Among the available immobilization procedures, covalent attachment of enzymes to solid supports is more advantageous than other methods since restricted diffusion of the product or substrate is absent [18]. In this context, amino resin and epoxy-activated carriers have proven to be very suitable for covalent immobilization of enzymes for industrial applications. To date, however, few studies have focused on the biosynthesis of SAM catalyzed by an immobilized enzyme.
In addition, it is of great importance to choose the suitable enzyme for the biosynthesis of SAM. In contrast to the MAT isoenzymes derived from animal tissues and microorganisms [17,19,20], the MAT encoded by the metK gene in Escherichia coli has some advantages, such as high specific activity and conversion efficiency, a low Km value, and a high level of expression in E. coli [21,22]. The lysates that contain the recombinant E. coli MAT have been used to synthesize SAM on the 30-mM scale. However, in order to successfully overcome the product inhibition of E. coli MAT, addition of a high concentration of sodium p-toluenesulfonate to the incubation was required [22,23]. Therefore, the development of an engineered E. coli MAT variant with greatly-reduced product inhibition would be of great value in the enzymatic synthesis of SAM.
In the present study, a simple and efficient method for biosynthesis of SAM using the immobilized enzyme was developed. Both wild-type MAT and engineered MAT varieties with significantly-reduced product inhibition were highly expressed, purified, and characterized. Additionally, two different methods for the immobilization of the partially-purified enzyme onto amino resin and epoxy resin were explored, and the characteristics of the immobilized enzyme were studied. Furthermore, we have successfully used the recycled immobilized enzyme to catalyze the synthesis of SAM on the 50-mM scale.
2. Results and Discussion
2.1. Purification and Characterization of Wild-Type and I303V MAT
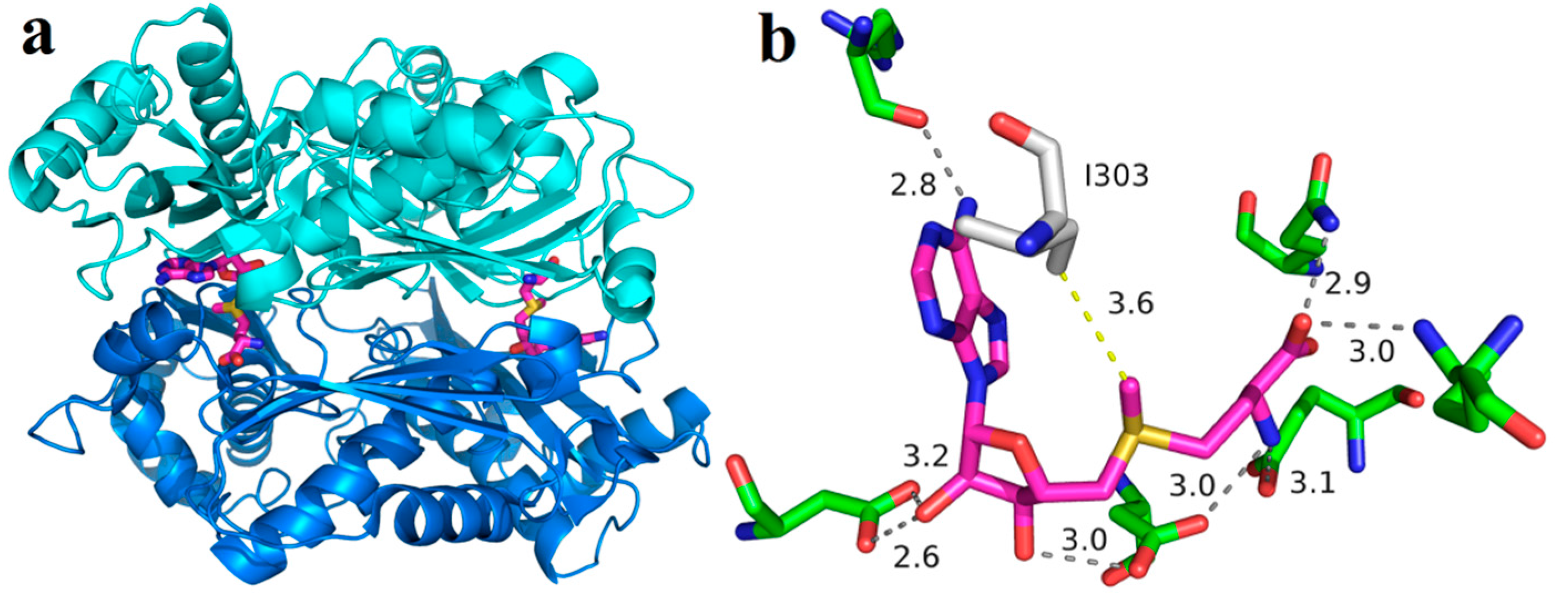
Due to product inhibition in incubations containing ≥1 mM SAM, the enzymatic synthesis of SAM using wild-type MAT from E. coli strain was not feasible unless a high concentration of sodium p-toluenesulfonate (~800 mM) was added to the reaction mixture to overcome the inhibition in the 30-mM scale reactions. A previous study showed that product inhibition by SAM may result from the prevention of ATP binding at the ATP binding site of the enzyme. When a certain concentration of sodium p-toluenesulfonate is added to the reaction solution it can directly bind with the cationic SAM to form SAM p-toluenesulfonate salt that does not have affinity anymore for the MAT, thus leading to the release of SAM from the active site [23]. Therefore, engineered variants of MAT with reduced product inhibition would be of great value. A previous study reported that the amino acid residue in the active site of SAM synthase from Bacillus subtilis, which directly interacts with the methyl group of SAM, is the major steric hindrance for the correct positioning of the substrate into the active site of the enzyme. When the position was exchanged with the less voluminous residue valine or alanine, the substrate spectrum of the enzyme to artificial S-alkylhomocysteines was extended [24]. We speculated that both hydrophobic interaction and steric hindrance likely hinder the release SAM product from the active site of the enzyme, thus leading to the observed product inhibition. Based on the information given above we analyzed the crystal structure of the MAT from Escherichia coli [25]. Figure 1a shows the ternary complex of the protein with its product SAM. A detailed view of the bound SAM molecule indicates that I303, which has the shortest distance to the methyl group of product SAM (3.6 Å), may contribute to product inhibition (Figure 1b). Therefore, the I303 residue was replaced with the less-voluminous residue valine or alanine by site-directed mutagenesis. The results showed that the specific activity of I303A MAT variant was ca. three-fold lower than wild-type MAT, whereas the specific activity of I303V MAT was ca. four-fold higher than wild-type MAT. Since the I → V substitution is the most conservative, it is not surprising that I303V MAT was less severely affected than substitution by alanine at this position. The very low specific activity of the I303A MAT precluded its further characterization. Then, the recombinant wild-type and I303V MAT proteins were expressed and purified.
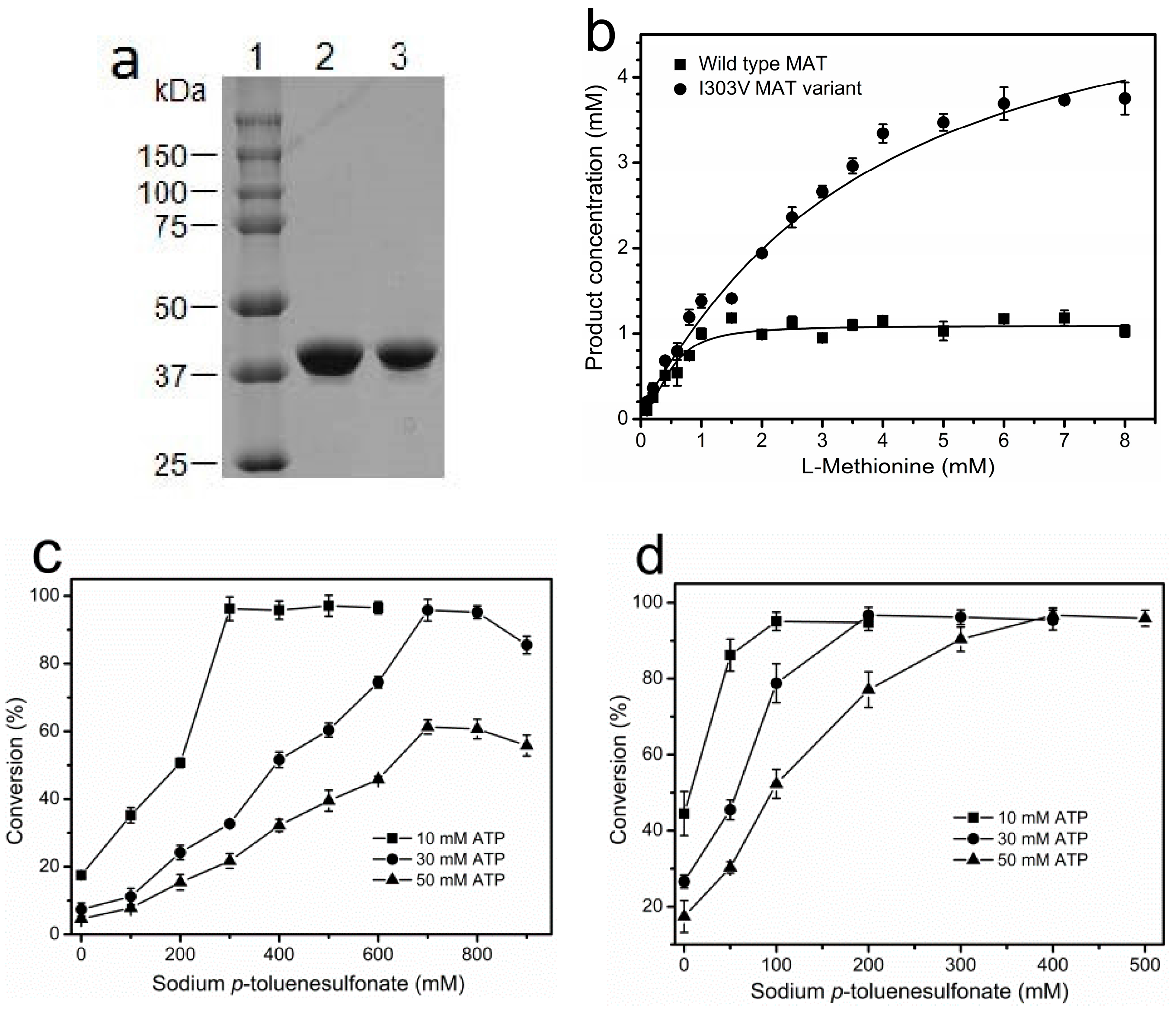
His6-tagged wild-type and I303V MAT proteins were purified in a single chromatography step using a HiTrap chelating nickel column. The purity of wild-type and I303V MAT proteins was judged to be >95% by sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE) analysis (Figure 2a). High expression of the recombinant enzyme in a 2-L shake flask was achieved (~150 mg/L wild-type MAT, ~100 mg/L I303V MAT). The expression level of the MAT gene from E. coli is much higher than that of the MAT gene from S. cerevisiae or B. subtilis [17,26]. Compared with the low specific activity of yeast MAT (~0.4 U/mg) [17], the specific activity of wild-type MAT and its variant was 4.1 ± 0.19 U/mg and 15.8 ± 0.84 U/mg, respectively. The I303V MAT enzyme shows a markedly-enhanced activity toward l-methionine. The Km values for both substrates (Kml-Methionine = 0.12 ± 0.005 mM; KmATP = 0.34 ± 0.013 mM) were slightly less than those of the wild-type MAT (Kml-Methionine = 0.18 ± 0.008 mM; KmATP = 0.43 ± 0.019 mM). The total activity of purified wild-type and I303V MAT that were obtained from the 2-L culture was 1230 ± 82 U and 3160 ± 152 U, respectively.
2.2. Product Inhibition of Wild-Type and I303V MAT
We next investigated the product inhibition of wild-type and I303V MAT. Both the purified wild-type and I303V MAT proteins were employed to synthesize SAM in the presence of 10 mM ATP and varying concentrations of l-methionine (0.1–8 mM). As shown in Figure 2b, in accordance with a previous report [23], the wild-type MAT is subject to severe product inhibition in reaction mixtures containing ≥1 mM SAM, and the inhibition constant (Ki) of SAM against ATP is 0.02 ± 0.001 mM. Expectedly, the product inhibition of I303V MAT is much reduced, and the product concentration reached 4 mM in the presence of 8 mM l-methionine. Compared with the wild-type MAT, I303V MAT has been a six-fold increase in the inhibition constant (Ki = 0.12 ± 0.01 mM). Furthermore, the suitability of wild-type and I303V MAT for the synthesis of SAM on the 10–50 mM scale was evaluated. As previously reported, the addition of a high concentration of sodium p-toluenesulfonate (700 mM) to a 30-mM scale incubation completely overcame the product inhibition of wild-type MAT (Figure 2c) [22,23]. However, the product inhibition during the enzymatic synthesis of SAM on a 50-mM scale was not removed even though 900 mM sodium p-toluenesulfonate was added to the reaction solution (Figure 2c). Compared with the wild-type MAT, as little as 200 mM sodium p-toluenesulfonate was required to completely overcome the product inhibition of I303V MAT on a 30-mM scale incubation. In total, a 95% conversion was achieved on a 50-mM scale in the presence of 400 mM sodium p-toluenesulfonate (Figure 2d). These results show that the I303V MAT variant from E. coli would be a suitable enzyme for producing SAM on the industrial scale.
2.3. Production of Recombinant I303V MAT in a Bioreactor
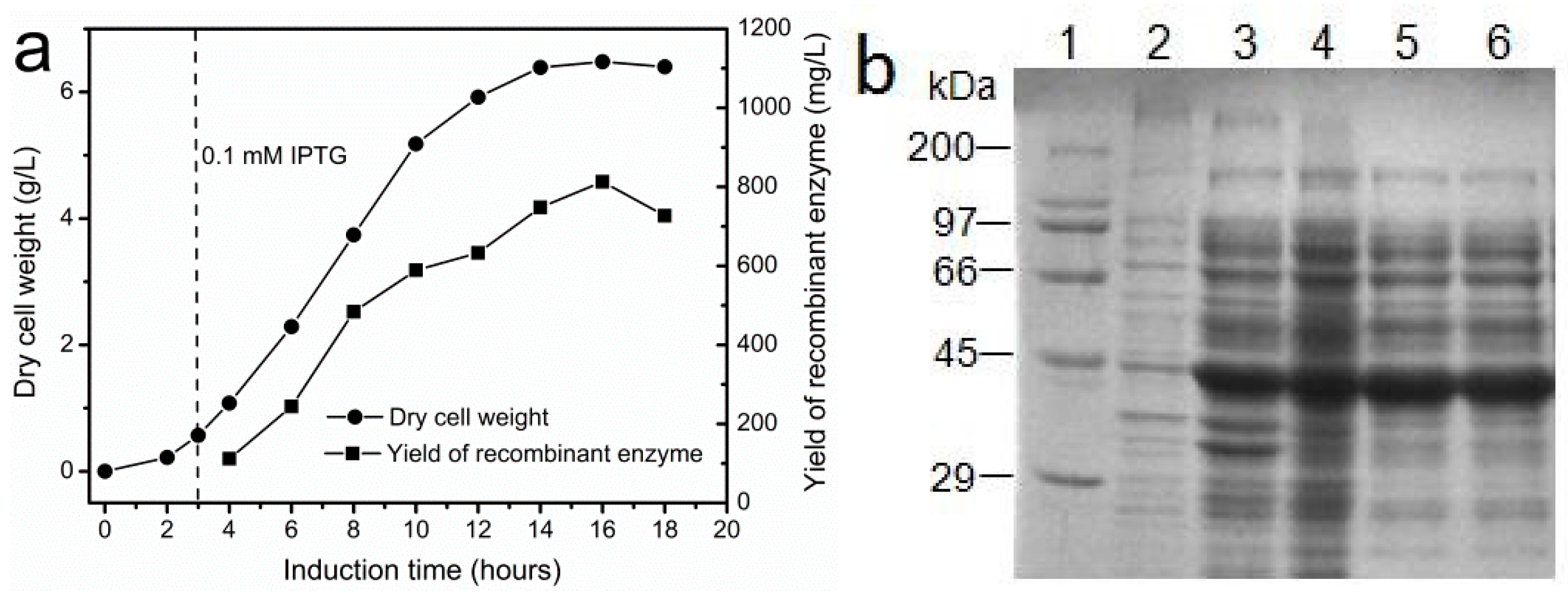
In order to obtain a large amount of I303V MAT for enzyme immobilization, the method of fermentation was employed in the preparation of the recombinant enzyme. The effect of induction time on the yield of recombinant I303V MAT is shown in Figure 3a. The enzyme accumulated as the induction time increased. The highest yield of the recombinant enzyme was ~800 mg/L after a 16 h induction, much higher than the highest yield of recombinant yeast MAT expressed in P. pastoris KM71 strains after a 160 h induction (~200 mg/L in a bioreactor) [17]. Moreover, in order to reduce the cost of producing the enzyme, the recombinant enzyme was partially purified by ammonium sulfate (1 M) precipitation (Figure 3b). Then, the supernatant containing the crude enzyme was collected and used to prepare the immobilized enzyme. The specific activity of the crude enzyme is 5.6 ± 0.4 U/mg.
2.4. Comparison of Covalent Immobilization Methods
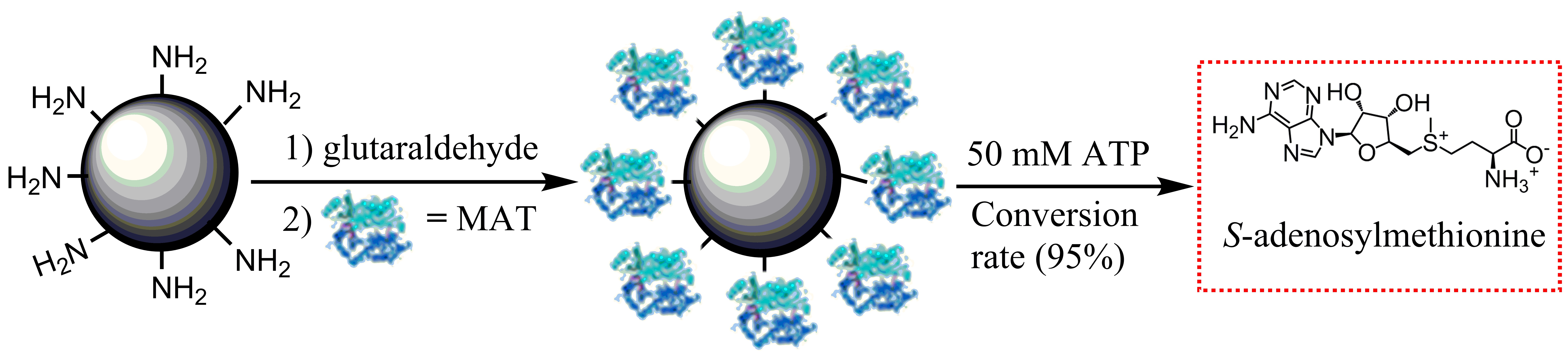
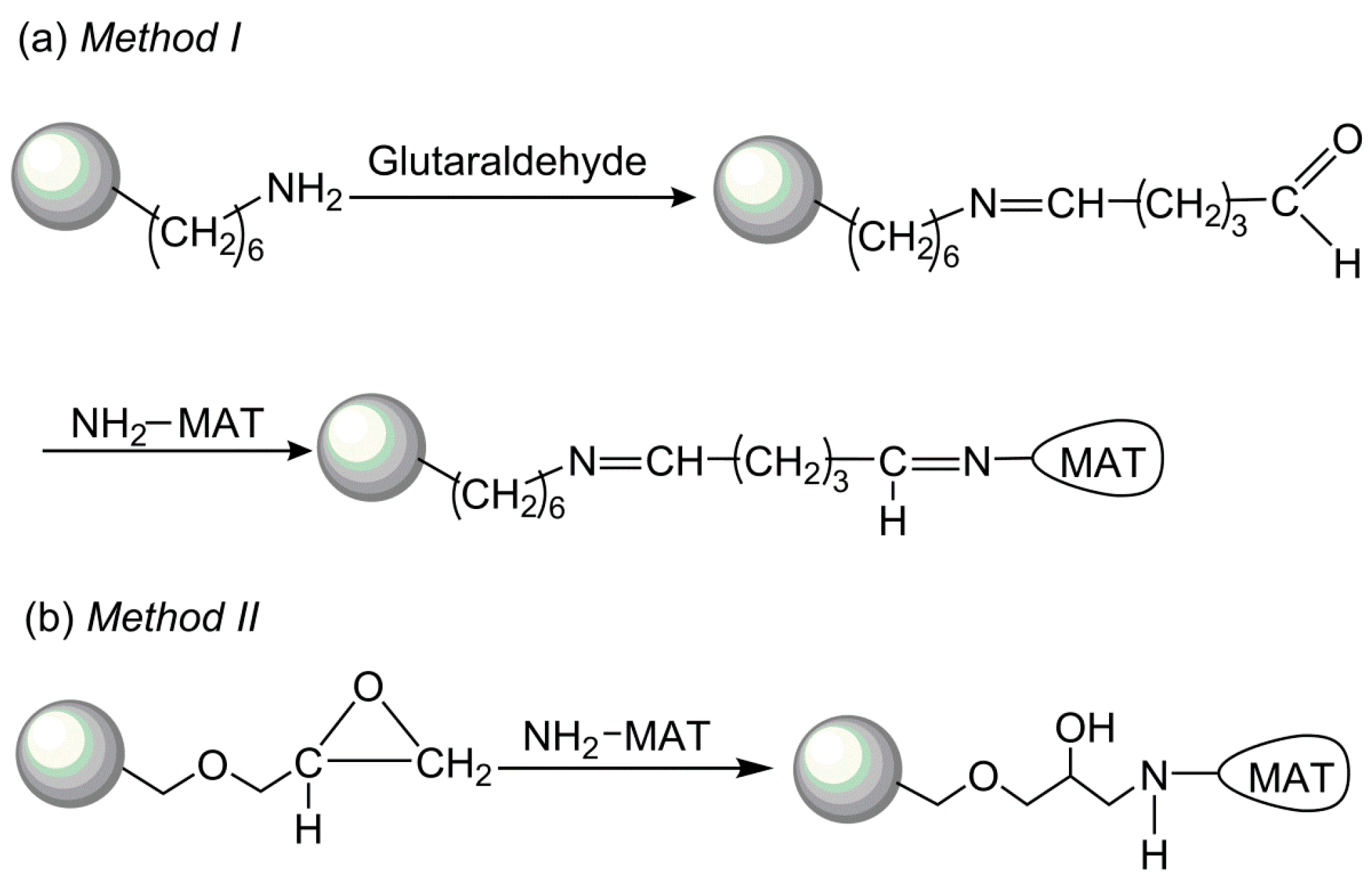
To select an optimal immobilization system for SAM production, two different methods to covalently immobilize I303V MAT have been explored: (a) enzyme binding to polymers via a spacer made from amino groups/glutaraldehyde; and (b) direct enzyme binding to polymers via the oxirane groups (Figure 4). The performances of these immobilized enzymes were compared with respect to enzyme loading, specific activity, and activity coupling yields. The results of the immobilization process with both supports are summarized in Table 1. A high enzyme loading was achieved with both amino resin LX-1000HA and epoxy resin LX-1000EP (~30 mg/g resin), indicating that the free enzyme covalently bound to the resins. However, very low activity coupling yields were obtained with epoxy resin (~3%), which could be related to multiple attachments and to reactions with groups that are involved in the active site of the enzyme and the tertiary structure of MAT. In comparison, a much higher activity coupling yield (~70%) was obtained with amino resin when the enzyme was bound via a longer spacer arm of amino groups/glutaraldehyde (Figure 4); this arm was considered to provide additional distance between the enzyme and the carrier and places the enzyme in a better position for catalysis after immobilization [27]. Due to the higher specific activity of the immobilized enzyme, further experiments were carried out with the enzyme immobilized on the amino resin.
We next investigated the effect of the quantity of the I303V MAT enzyme (10–50 mg) on the enzyme loading on the amino resin LX-1000HA. In each experiment, 0.5 g of resin was immersed in a certain volume of enzyme solution. Table 1 shows that enzyme loading on the amino resin LX-1000HA increased rapidly as the amount of I303V MAT increased, and the loading leveled off at 30 mg/g support. The enzyme coupling yield was in the range of 70.8–92.3% for amino resin LX-1000HA. However, the activity coupling yield is inversely related to the amount of enzyme that was bound. For example, increasing enzyme loading from 8.9 to 35.4 mg/g support resulted in decreasing the activity coupling yield from 83.6% to 54.5%, possibly due to packing of the immobilized enzyme on the support surface that could limit the access of substrates and, thus, reduce the enzyme activity. It is generally acknowledged that the activity coupling yield of immobilization processes decreases when enzyme loading exceeds a certain value and a yield that provides optimum activity should be selected [27]. In this study, the maximum activity of the immobilized enzyme that was attained was 109.6 ± 7.6 U/gsupport with 30 mg/gsupport of the enzyme exposed to the support, and the corresponding activity coupling yield was 70.7%.
2.5. Thermal Inactivation of Soluble and Immobilized Enzyme
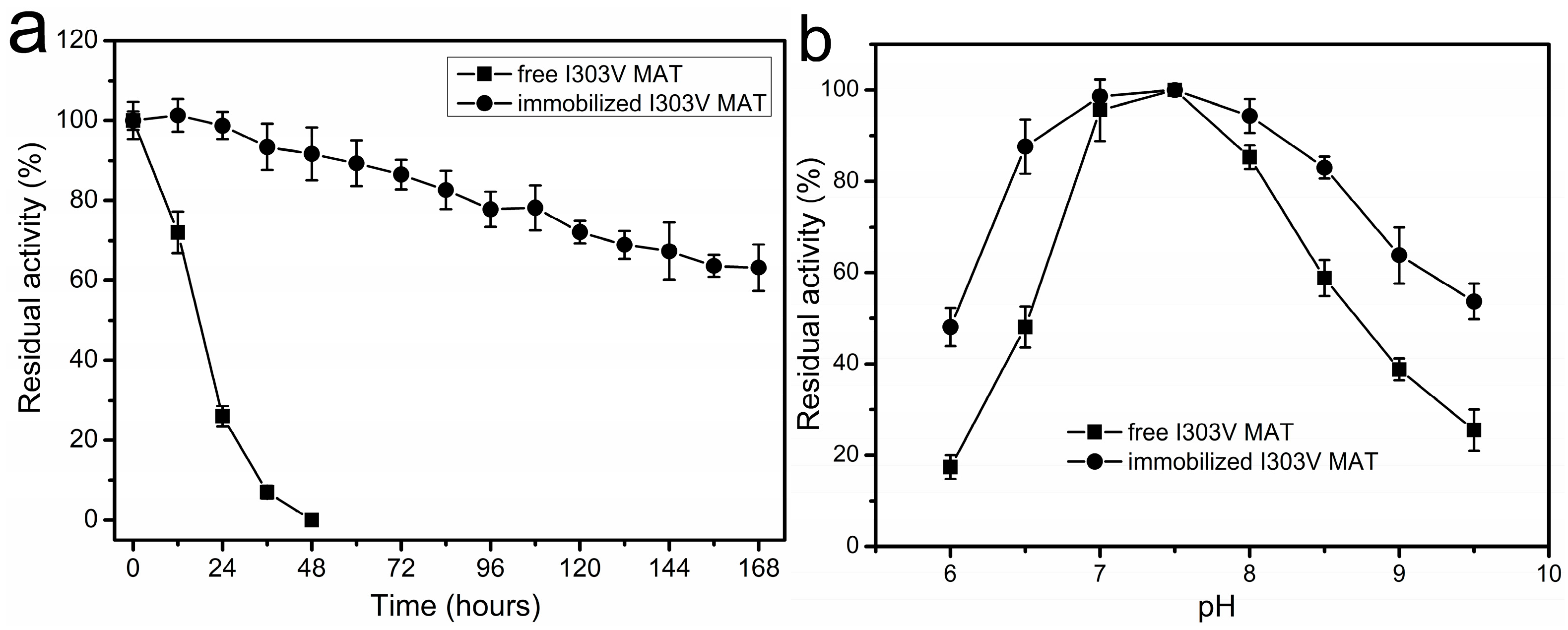
Since the thermal stability of free MAT enzymes derived from E. coli and S. cerevisiae is poor, these free enzymes are not suitable for use in synthesis of SAM on the industrial scale. For example, after incubation of the recombinant yeast MAT at 40 °C for 6 h the enzyme lost its activity completely [17]. The residual activity of the recombinant E. coli MAT incubated at 37 °C for 12 h was approximately 70%, and after 36–48 h of incubation at 37 °C the enzyme lost its activity completely (Figure 5a). Thus, the main objective of this study was to improve enzyme stability and develop an immobilized enzyme that is capable of synthesizing SAM for extended periods of time. Accordingly, the thermal stability of the free and immobilized enzyme on the amino resin LX-1000HA was evaluated at 37 °C for a 168 h incubation. As shown in Figure 5a, the stability of the immobilized enzyme was much higher than that of the soluble enzyme. After incubation of free MAT for 36–48 h the enzyme lost its activity completely. However, 63% of the initial enzyme activity of immobilized MAT was remained after 168 h of incubation. Plots of the natural log of relative enzyme activity as a function of time showed first-order kinetics for both free and immobilized MAT and were used to evaluate the deactivation constants (kd) and half-life values (t1/2). The estimated parameters were kd = 0.076 ± 0.006/h and t1/2 = 9.2 ± 0.7 h for soluble enzyme and kd = 0.003 ± 0.0002/h and t1/2 = 229.5 ± 18.8 h for immobilized enzyme at 37 °C. Overall, improvement in MAT thermal stability by covalent immobilization on an amino resin was achieved. Comparing the t1/2 values that were obtained at 37 °C, it can be concluded that the immobilized enzyme was almost 25-fold more stable than the free enzyme. Immobilized preparations resulted in enhanced thermal stability, which could be attributed to the rigidification of the enzyme’s tertiary structure.
2.6. Effect of pH on Enzyme Stability
The pH stabilities of both the free and immobilized enzyme were compared by immersing them in 50 mM buffer at pH values that ranged from 6 to 9.5 at 4 °C for 12 h, and then determining their residual activity under the standard reaction conditions. The results are shown in Figure 5b. Both the immobilized and free enzymes were more stable in neutral or alkaline solution. A wider range of pH stability was obtained with the immobilized enzyme. For example, the free enzyme remained stable in the pH range from 7 to 8, while the immobilized MAT enzyme was found to be stable from pH 6.5–8.5.
2.7. Kinetic Analysis
Kinetics of the immobilized enzyme were investigated using varying concentrations of l-methionine and ATP substrates. The kinetic constant Km was calculated from the double reciprocal plot, and the values of Kml-methionine and KmATP were 0.22 ± 0.008 mM and 0.18 ± 0.006 mM, respectively. The immobilized enzyme exhibited a Kml-methionine value that was approximately two-fold higher than that of free MAT (Kml-methionine = 0.12 ± 0.005 mM). The increase in Kml-methionine value might be due to either tertiary structure changes in the MAT that were introduced by the immobilization procedure or the lower accessibility of the L-Met substrate to the active site of the immobilized enzyme [28]. In contrast, the Kml-ATP value of immobilized enzyme was approximately two-fold less than that of free MAT (KmATP = 0.34 ± 0.013 mM). The decrease in KmATP value is probably caused by the increased concentration of the ATP substrate around the immobilized enzyme that is caused by the interaction between the positively charged support (NH3+) and the negatively-charged ATP. Additionally, the values of Vmaxl-methionine and VmaxATP of the immobilized enzyme were 2.45 ± 0.089 mmol h−1 mg−1 and 2.81 ± 0.093 mmol h−1 mg−1, respectively. Compared with the Vmax values of the free enzyme (Vmaxl-methionine = 4.71 ± 0.196 mmol h−1 mg−1; VmaxATP = 4.41 ± 0.168 mmol h−1 mg−1), the Vmax values showed a decreasing trend on enzyme immobilization in the cases of all substrates, indicating a slight loss of enzyme activity.
2.8. Recycling of Immobilized Enzyme for SAM Synthesis
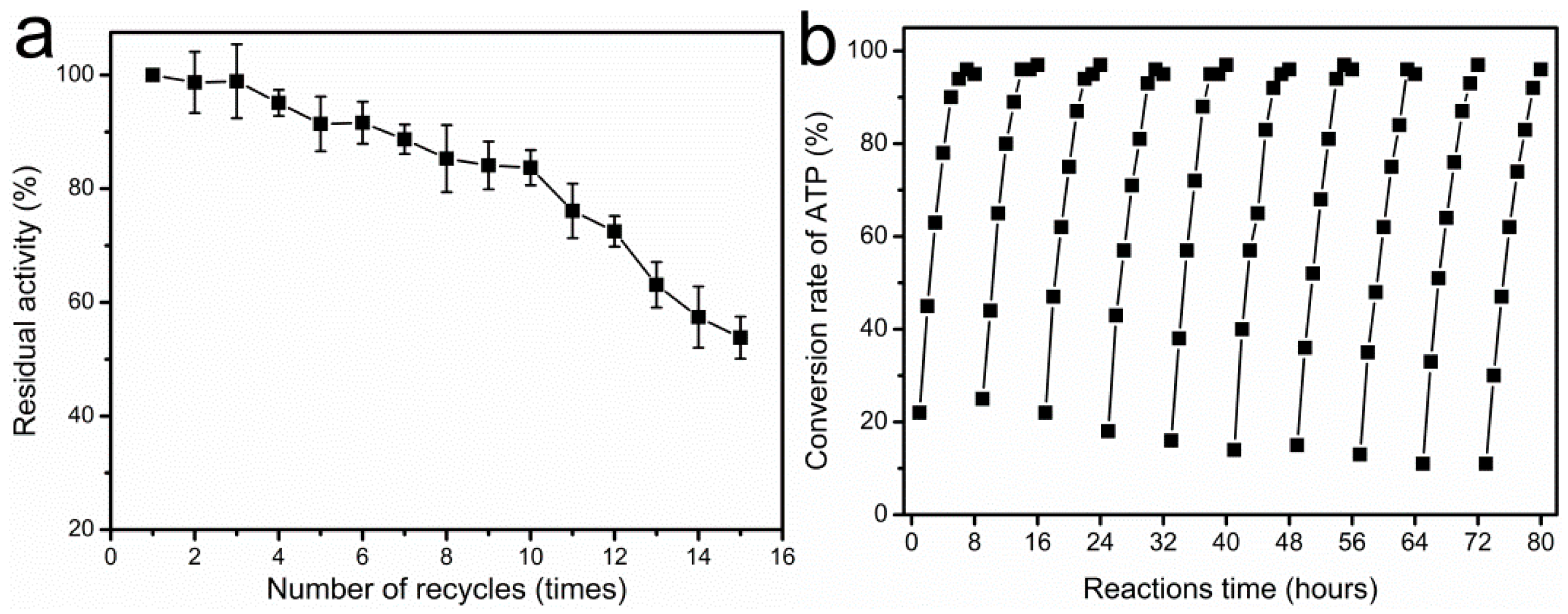
When immobilized enzymes are employed for preparative or industrial use, characterization of their operational stabilities is very important. In the present study, the operational stability of the immobilized enzyme was evaluated in a repeated batch process. Figure 6a shows the effect of recycled use on activity of immobilized enzyme. It was found that the immobilized enzyme maintained 84% of its initial activity after 10 cycles of 8 h and 54% after 15 cycles. No leakage of the enzyme from the support was observed during repeated batch processes, suggesting that the deactivation is essentially due to the activity loss of the enzyme, and not to its desorption from the support. In contrast, the activity retention of the immobilized yeast MAT after three reuses was only ~10% of its initial activity [17]. Therefore, the operational stability of the immobilized enzyme may indicate the applicability of immobilized enzyme for continuous production of SAM. Moreover, the immobilized enzyme could be recycled for 10 times in the synthesis of SAM on a 50-mM scale, with a high conversion rate of ATP (>95%). The 50 mM ATP was completely converted after 6 h from the first to the seventh cycle. With the reduced activity of immobilized enzyme it took 7–8 h to completely covert the 50-mM ATP from the eighth to the tenth cycle (Figure 6b). Additionally, due to the existence of sodium p-toluenesulfonate in reaction solution, the stability of SAM product (SAM p-toluenesulfonate salt) could be increased during the enzymatic synthesis of SAM. The purity of SAM in reaction mixture reached to approximately 85–90% by high performance liquid chromatography (HPLC) analysis. Generally, in comparison with the enzymatic synthesis of SAM in other reports [17,22,23], our work presented a significant improvement on the productivity of SAM and the short reaction time (8 h) by using immobilized enzyme.
3. Materials and Methods
3.1. Materials
DNA polymerase, restriction enzyme, and T4 DNA ligase were obtained from TaKaRa (Tokyo, Japan). E. coli DH5α was used as a host for DNA manipulation, and E. coli BL21 (DE3) was used for MAT gene expression. Vector pET-28a(+) was purchased from Novagen (Madison, WI, USA). Yeast extract and peptone were obtained from Oxoid (Hampshire, UK). Ni Sepharose 6 Fast Flow resin and HiTrap desalting column were purchased from GE Healthcare (Little Chalfont, Buckinghamshire, UK); glutaraldehyde solution, L-Met, isopropyl β-d-1-thiogalactopyranoside (IPTG), and kanamycin were obtained from Amresco (Solon, OH, USA); adenosine triphosphate (ATP) was purchased from Kaiping Genuine Biochemical Pharmaceutical Co., Ltd. (Jiangmen, China); S-adenosylmethionine was obtained from Sigma-Aldrich (St. Louis, MO, USA). Amino resin LX-1000HA (content of amino groups ~600 μmol/g; particle size ~300 μm) and epoxy resin LX-1000EP (content of epoxide groups ~600 μmol/g; particle size ~300 μm) were purchased from Sunresin (Xi’an, China).
3.2. Expression and Purification of Wild-Type and I303V MAT
The E. coli MAT gene was amplified by PCR using the genome of E. coli DH5α as the template with the following primers. 5′-ACGTGAATTCATGGCAAAACACCTTTTTAC-3′ and 5′-ATGTAAGCTTTTACTTCAGACCGGCAGCATC-3′. In the sense primer, the Eco RI restriction site (underlined) was added. In the antisense primer, the HindIII restriction site (underlined) was inserted. The MAT gene was subcloned into the pET-28a(+) vector to construct the recombinant plasmid pET-28a-MAT. For construction of a I303V MAT variant, the wild-type gene was modified using the QuikChange II Site-Directed Mutagenesis Kit (Agilent Technologies, Santa Clara, CA, USA) according to the instructions. The plasmids were then transformed into E. coli BL21 (DE3) for expression of the wild-type and I303V MAT. The recombinant strains were grown in LB medium containing 50 mg/L kanamycin to an optical density (OD600) of 0.6 at 30 °C, and then IPTG was added to a final concentration of 0.1 mM. After induction at 30 °C for 12–16 h in a 2 L flask, cells were harvested by centrifugation at 6000 rpm for 15 min at 4 °C. Cell pellets obtained from a 2 L culture were resuspended in 200 mL of phosphate-buffered saline (PBS) that was supplemented with 10 mg of lysozyme and an “EDTA-free” protease inhibitor tablet (Roche). The cell lysate was prepared by sonication and centrifuged at 12,000 rpm for 30 min at 4 °C, and the supernatant was obtained. The recombinant wild-type MAT or I303V MAT was purified on a HiTrap chelating nickel column (elution buffer: 50 mM sodium phosphate, 300 mM NaCl, 500 mM imidazole, pH 7.4), followed by a HiTrap desalting column using gel filtration buffer (50 mM Tris-HCl, pH 7.5). Fractions containing the wild-type MAT or I303V MAT were pooled, and the aliquoted enzyme was stored at −80 °C.
To obtain a large amount of the recombinant I303V MAT, the fermentation of the recombinant E. coli was performed at 30 °C in a 10 L bioreactor (Baoxing bio-engineering equipment Co., Ltd., Shanghai, China) with 6 L of TB medium (24 g/L yeast extract, 12 g/L peptone, 72 mM K2HPO4, 17 mM KH2PO4, and 4 mL/L glycerol) that was supplemented with 50 mM kanamycin. The inoculum was grown at 30 °C in shake flasks to an OD600 of 0.6 in LB medium, and then 300 mL of inoculum was added to the bioreactor. The recombinant strains were grown in TB medium to an OD600 of ~1.5 at 30 °C, at which point IPTG was added to a final concentration of 0.1 mM. Dissolved oxygen was maintained at more than 20% saturation, and the stirring rate was maintained between 400 and 800 rpm. The fermentation was operated in batch mode until all of the glycerol was consumed (for approximately 18 h), which was indicated by a rapid increase in dissolved oxygen concentration. Biomass was determined by measuring the OD600 and the dry cell weight.
To reduce the enzyme cost, the recombinant I303V MAT protein was partially purified in one step by ammonium sulfate precipitation. The cells were collected by centrifugation at 6000 rpm for 15 min at 4 °C. Cell pellets were resuspended in 50 mM Tris-HCl buffer (pH 7.5), followed by high-pressure homogenization (100 MPa). The recombinant enzyme was partially purified by adding the ammonium sulfate to the E. coli cell lysate to a final concentration of 1 M. Then, the supernatant containing the recombinant I303V MAT protein was collected and used to prepare the immobilized enzyme.
3.3. Immobilization of I303V MAT
Two different methods were studied to evaluate their suitability for covalent immobilization of recombinant I303V MAT on amino resin LX-1000HA and epoxy resin LX-1000EP. The procedure that was used to immobilize the enzyme on amino resin LX-1000HA is as follows: The amino group of the LX-1000A resin reacts with glutaraldehyde, and the amino group of the enzyme is then coupled to the carriers via a free carbonyl group of the support. The resin was first washed three times with water, which was followed by reaction for 5 h with 2% (w/v) glutaraldehyde solution buffered at pH 7.5 (50 mM Tris-HCl buffer) at room temperature with gentle shaking. Next, the resin was washed five times with 50 mM Tris-HCl buffer. Activated resin was then incubated with the partially purified enzyme solution in 50 mM Tris-HCl buffer (5 mg/mL, pH 7.5) and 4 °C for 24 h with gentle shaking. Finally, unbound enzyme was removed by washing the resin five times with 50 mM Tris-HCl buffer at pH 7.5 and stored at 4 °C until use. The method for enzyme immobilization on epoxy resin LX-1000EP involves the enzyme directly binding to polymers via oxirane groups. Resin LX-1000EP was incubated with enzyme solution in 50 mM Tris-HCl buffer (5 mg/mL, pH 7.5) at 4 °C for 24 h. Subsequently, the resin was collected by filtration, washed with 50 mM Tris-HCl buffer, pH 7.5 and stored at 4 °C until use.
The efficiency of immobilization was evaluated based on enzyme coupling yield and activity coupling yield. The enzyme coupling yield, ηenz (%), and activity coupling yield, ηact (%), can be calculated as follows:
where M1 is the amount of immobilized enzyme, M0 the initial amount of enzyme, SA2 the specific activity of immobilized enzyme, and SA1 is the specific activity of the free enzyme.
3.4. Determination and Analysis
3.4.1. Spectrophotometric Assay for MAT Activity
The activity of recombinant MAT was determined in 96-well plate at 37 °C according to a previous report [24]. Briefly, the 50 μL assay mixture contained 10 mM ATP, 0–10 mM L-Met, 50 mM K2SO4, 20 mM MgSO4, and 100 mM Tris-HCl, pH 8.0. The reaction mixture was incubated at 37 °C for 5 min, and the reaction was started by the addition of free or immobilized enzyme. After incubation for 30 min at 37 °C, the concentration of phosphate that had been released from ATP was measured. In total, 100 μL of malachite green-molybdate solution was added, and the absorbance at 620 nm was measured after 5 min of incubation. A standard solution of sodium phosphate (0–75 μM Na2HPO4 and Na2H2P2O7 in 10 mM ATP) was used for calculation of the phosphate concentration. The enzyme activity was calculated based on a linear fit to the phosphate concentration. One unit (U) of enzyme activity was defined as the amount of MAT that catalyzes the formation of 1 μM phosphate per minute at 37 °C, pH 8.0. For the kinetic analysis, the reaction system is the same as for the enzyme activity determination except that the substrate concentration of l-methionine or ATP was varied from 0.01–1.0 mM.
3.4.2. High-Performance Liquid Chromatography (HPLC) Analysis
Synthesis of SAM that was catalyzed by free or immobilized enzyme was stopped by adding trichloroacetic acid to a final concentration of 15% (w/v). The reaction mixture was centrifuged, and the supernatant was analyzed by HPLC (Waters, Milford, MA, USA) using a Kromasil C18 column (4.6 mm × 250 mm, 5 μM) and monitoring the absorption of the products at 254 nm. The mobile phase that was used for the SAM analysis consisted of 10 mM sodium 1-hexanesulfonate, 1% acetic acid (v/v) and 15% acetonitrile (v/v) with the flow rate at 1 mL/min. Peak area analysis was performed based on the standard calibration curves of SAM.
3.4.3. Protein Assay
The protein concentrations in the enzyme solutions and on the resins were determined by the Coomassie Blue method [29]. The amount of immobilized enzyme was calculated according to the difference between the amount of protein that was added to the immobilized solution and the amount of residual protein that was present in the supernatant after immobilization. During enzyme purification, active proteins were analyzed with a 12% SDS-PAGE gel under reducing conditions. The gels were stained using Coomassie Brilliant Blue R250, and apparent molecular masses were estimated using molecular markers from TaKaRa (Tokyo, Japan).
3.4.4. Thermal Stability of the Soluble and Immobilized Enzymes
Thermal stability assays were carried out at 37 °C in solution (50 mM Tris-HCl, pH 7.5). The free MAT enzyme (1 mg/mL) was incubated for a various time periods. Then, aliquots (1 mL) were taken, and the residual activity was measured using a spectrophotometric assay. For the immobilized MAT, the same amount of biocatalyst was incubated at 37 °C in 50 mM Tris-HCl buffer, pH 7.5. After various incubation times, the residual activity was determined according to the method above. No enzyme loss was detected in the reaction mixture containing the immobilized enzyme, suggesting that the deactivation is due to the activity loss of the immobilized enzyme. Relative activity is presented as the percentage of the enzyme activity that was retained compared to the activity that was exhibited before the incubation, which was taken as 100%. Data from experiments were expressed as the mean ± SD (n = 3).
3.4.5. Effect of pH on the Stability of the Soluble and Immobilized Enzymes
The effect of pH on the stability of soluble and immobilized enzyme was determined using two buffer systems: sodium phosphate (pH 6–7.5) and Tris-HCl (pH 8–9.5). Partially-purified soluble or immobilized enzyme (1 mg soluble enzyme or 50 mg immobilized support) was added to 5 mL of 50 mM buffer, followed by incubation for 12 h at 4 °C. The residual activity of the treated enzymes was measured using the spectrophotometric assay described above. The activity of soluble and immobilized enzymes that were treated at pH 7.5 was defined as 100%, respectively.
3.5. Reusability of Immobilized Enzyme on Scaled-Up Production of SAM
Biosynthesis of SAM was catalyzed by the immobilized enzyme on a 50-mM scale. Due to the instability of SAM under the alkaline conditions, the reaction was carried out at pH 7.0. Briefly, the reaction mixture consisted of 50 mM ATP, 65 mM l-methionine, 50 mM K2SO4, 100 mM MgSO4, 400 mM sodium p-toluenesulfonate, and 100 mM Tris-HCl, pH 7.0. After the addition of immobilized enzyme (10%, w/v), the reaction mixture was incubated for 8 h at 37 °C. The operational stability of the immobilized enzyme was tested in 15 consecutive 8 h batches. After the completion of each batch, the immobilized enzyme was separated from the reaction mixture by filtration and washed with 100 mM Tris-HCl, pH 7.0. The leaked MAT activity in the reaction mixture was measured using the spectrophotometric assay described above. The consumption of ATP and the amount of SAM in the reaction mixture were analyzed by HPLC. The residual activity of the immobilized enzyme was measured using spectrophotometric assay. The immobilized enzyme was reused in the following batch. The activity of immobilized enzyme in the first batch was assumed to 100%. The residual activity after each recycle was calculated by dividing the enzyme activity after each batch by the enzyme activity in the first batch and multiplying by 100.
4. Conclusions
The recombinant I303V MAT variant from E. coli significantly reduced the product inhibition in the synthesis of SAM and was expressed at high levels in a 10 L bioreactor. The specific activity of I303V MAT increased four times with respect to the wild-type MAT. The recombinant I303V MAT was effectively immobilized onto the amino resin LX-1000HA with the highest activity coupling yield (~84%). Moreover, the immobilized enzyme retained 84% activity after being used for 10 batches, while converting more than 95% of the substrate ATP on the 50-mM scale. The method presented in this work would be practical for large-scale SAM synthesis in the future.
Acknowledgments
This work was supported by the National Natural Science Foundation of China (grant number: 31471086), the Key Science and Technology Program of Shaanxi Province, China (grant number: 2017GY-143), and the Seed Foundation of Innovation and Creation for Graduate Students in Northwestern Polytechnical University (grant number: Z2017233).
Author Contributions
W.N. conceived and designed the experiments; W.N., S.C., M.Y., and L.X. performed the experiments; W.N., S.C., M.Y., and L.X. analyzed the data; and W.N. wrote the paper.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Fontecave, M.; Atta, M.; Mulliez, E. S-adenosylmethionine: Nothing goes to waste. Trends Biochem. Sci. 2004, 29, 243–249. [Google Scholar] [CrossRef] [PubMed]
- Grillo, M.A.; Colombatto, S. S-adenosylmethionine and its products. Amino Acids 2008, 34, 187–193. [Google Scholar] [CrossRef] [PubMed]
- Hosea Blewett, H.J. Exploring the mechanisms behind S-adenosylmethionine (SAMe) in the treatment of osteoarthritis. Crit. Rev. Food Sci. Nutr. 2008, 48, 458–463. [Google Scholar] [CrossRef] [PubMed]
- Lu, S.C.; Mato, J.M. S-adenosylmethionine in liver health, injury, and cancer. Physiol. Rev. 2012, 92, 1515–1542. [Google Scholar] [CrossRef] [PubMed]
- Noureddin, M.; Mato, J.M.; Lu, S.C. Nonalcoholic fatty liver disease: Update on pathogenesis, diagnosis, treatment and the role of S-adenosylmethionine. Exp. Biol. Med. 2015, 240, 809–820. [Google Scholar] [CrossRef] [PubMed]
- Shippy, R.A.; Mendez, D.; Jones, K.; Cergnul, I.; Karpiak, S.E. S-adenosylmethionine (SAM-e) for the treatment of depression in people living with HIV/AIDS. BMC Psychiatry 2004, 4, 38. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, H.; Wang, Z.; Cai, H.; Zhou, C. Progress in the microbial production of S-adenosyl-l-methionine. World J. Microbiol. Biotechnol. 2016, 32, 153. [Google Scholar] [CrossRef] [PubMed]
- Chu, J.; Qian, J.; Zhuang, Y.; Zhang, S.; Li, Y. Progress in the research of S-adenosyl-l-methionine production. Appl. Microbiol. Biotechnol. 2013, 97, 41–49. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Yang, Y.; Wang, Z.; Dou, J.; Wang, H.; Zhou, C. Elevated intracellular acetyl-CoA availability by ACS2 overexpression and MLS1 deletion combined with metK1 introduction enhanced SAM accumulation in Saccharomyces cerevisiae. Biochem. Eng. J. 2016, 107, 26–34. [Google Scholar] [CrossRef]
- He, J.; Deng, J.; Zheng, Y.; Gu, J. A synergistic effect on the production of S-adenosyl-l-methionine in Pichia pastoris by knocking in of S-adenosyl-l-methionine synthase and knocking out of cystathionine-beta synthase. J. Biotechnol. 2006, 126, 519–527. [Google Scholar] [CrossRef] [PubMed]
- Hu, X.; Chu, J.; Zhang, S.; Zhuang, Y.; Wang, Y.; Zhu, S.; Zhu, Z.; Yuan, Z. A novel feeding strategy during the production phase for enhancing the enzymatic synthesis of S-adenosyl-l-methionine by methylotrophic Pichia pastoris. Enzyme Microb. Technol. 2007, 40, 669–674. [Google Scholar] [CrossRef]
- Hu, H.; Qian, J.; Chu, J.; Wang, Y.; Zhuang, Y.; Zhang, S. Optimization of l-methionine feeding strategy for improving S-adenosyl-l-methionine production by methionine adenosyltransferase overexpressed Pichia pastoris. Appl. Microbiol. Biotechnol. 2009, 83, 1105–1114. [Google Scholar] [CrossRef] [PubMed]
- Kanai, M.; Mizunuma, M.; Fujii, T.; Iefuji, H. A genetic method to enhance the accumulation of S-adenosylmethionine in yeast. Appl. Microbiol. Biotechnol. 2017, 101, 1351–1357. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Wang, D.; Wei, G.; Shao, N. Enhanced co-production of S-adenosylmethionine and glutathione by an ATP-oriented amino acid addition strategy. Bioresour. Technol. 2012, 107, 19–24. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Wang, X.; Su, E.; Fang, G.; Ren, Y.; Wei, D. A new fermentation strategy for S-adenosylmethionine production in recombinant Pichia pastoris. Biochem. Eng. J. 2008, 41, 74–78. [Google Scholar] [CrossRef]
- Zhao, W.; Hang, B.; Zhu, X.; Wang, R.; Shen, M.; Huang, L.; Xu, Z. Improving the productivity of S-adenosyl-l-methionine by metabolic engineering in an industrial Saccharomyces cerevisiae strain. J. Biotechnol. 2016, 236, 64–70. [Google Scholar] [CrossRef] [PubMed]
- Luo, Y.; Yuan, Z.; Luo, G.; Zhao, F. Expression of secreted His-tagged S-adenosylmethionine synthetase in the methylotrophic yeast Pichia pastoris and its characterization, one-step purification, and immobilization. Biotechnol. Prog. 2008, 24, 214–220. [Google Scholar] [CrossRef] [PubMed]
- Hormigo, D.; de la Mata, I.; Acebal, C.; Arroyo, M. Immobilized aculeacin A acylase from Actinoplanes utahensis: Characterization of a novel biocatalyst. Bioresour. Technol. 2010, 101, 4261–4268. [Google Scholar] [CrossRef] [PubMed]
- Sakata, S.F.; Shelly, L.L.; Ruppert, S.; Schutz, G.; Chou, J.Y. Cloning and expression of murine S-adenosylmethionine synthetase. J. Biol. Chem. 1993, 268, 13978–13986. [Google Scholar] [PubMed]
- Zhou, J.; Chu, J.; Wang, Y.H.; Zhang, S.L.; Zhuang, Y.P.; Yuan, Z.Y. Purification and properties of Saccharomyces cerevisiae S-adenosylmethionine synthetase expressed in recombinant Pichia pastoris. World J. Microbiol. Biotechnol. 2008, 24, 789–796. [Google Scholar] [CrossRef]
- Markham, G.D.; Hafner, E.W.; Tabor, C.W.; Tabor, H. S-adenosylmethionine synthetase from Escherichia coli. J. Biol. Chem. 1980, 255, 9082–9092. [Google Scholar] [PubMed]
- Park, J.; Tai, J.; Roessner, C.A.; Scott, A.I. Enzymatic synthesis of S-adenosyl-l-methionine on the preparative scale. Bioorg. Med. Chem. 1996, 4, 2179–2185. [Google Scholar] [CrossRef]
- Park, J.; Tai, J.; Roessner, C.A.; Scott, A.I. Overcoming product inhibition of S-adenosyl-l-methionine (SAM) synthetase: preparation of SAM on the 30 mM scale. Bioorg. Med. Chem. Lett. 1995, 5, 2203–2206. [Google Scholar] [CrossRef]
- Dippe, M.; Brandt, W.; Rost, H.; Schmidt, J.; Wessjohann, L.A. Rationally engineered variants of S-adenosylmethionine (SAM) synthase: Reduced product inhibition and synthesis of artificial cofactor homologues. Chem. Commun. 2015, 51, 3637–3640. [Google Scholar] [CrossRef] [PubMed]
- Komoto, J.; Yamada, T.; Takata, Y.; Markham, G.D.; Takusaqawa, F. Crystal structure of the S-adenosylmethionine synthetase ternary complex: A novel catalytic mechanism of S-adenosylmethionine synthesis from ATP and Met. Biochemistry 2004, 43, 1821–1831. [Google Scholar] [CrossRef] [PubMed]
- Kamarthapu, V.; Rao, K.V.; Srinivas, P.N.; Reddy, G.B.; Reddy, V.D. Structural and kinetic properties of Bacillus subtilis S-adenosylmethionine synthetase expressed in Escherichia coli. Biochim. Biophys. Acta 2008, 1784, 1949–1958. [Google Scholar] [CrossRef] [PubMed]
- Knezevic, Z.; Milosavic, N.; Bezbradica, D.; Jakovljevic, Z.; Prodanovic, R. Immobilization of lipase from Candida rugosa on Eupergit® C supports by covalent attachment. Biochem. Eng. J. 2006, 30, 269–278. [Google Scholar] [CrossRef]
- Chiou, S.H.; Wu, W.T. Immobilization of Candida rugosa lipase on chitosan with activation of the hydroxyl groups. Biomaterials 2004, 25, 197–204. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
Figure 1.
Tertiary structure of the methionine adenosyltransferase (MAT) from E. coli (PDB: 1RG9). (a) The dimeric MAT with S-adenosylmethionine (SAM) bound between two monomer units; and (b) the active site with the bound product SAM (purple carbon atoms). The methyl group of product SAM directly interacts with the isoleucine I303 which, therefore, became the target of site-directed mutagenesis.
Figure 1.
Tertiary structure of the methionine adenosyltransferase (MAT) from E. coli (PDB: 1RG9). (a) The dimeric MAT with S-adenosylmethionine (SAM) bound between two monomer units; and (b) the active site with the bound product SAM (purple carbon atoms). The methyl group of product SAM directly interacts with the isoleucine I303 which, therefore, became the target of site-directed mutagenesis.

Figure 2.
The purification and product inhibition of wild-type MAT and I303V MAT variant. (a) SDS-PAGE (12%) analysis of the purified wild-type and I303V MAT proteins from recombinant E. coli that was cultured in a 2-l shake flask. Lane 1: molecular weight marker. Lane 2: the purified wild-type MAT. Lane 3: the purified I303V MAT variant; (b) conversion rates of l-methionine catalyzed by the wild-type and I303V MAT proteins as a function of the substrate concentration. The reaction mixture, which contained 10 mM ATP and varying concentrations of l-methionine, was incubated for 5 h at 37 °C in the presence of excess enzyme. The reaction yield was determined by measuring the phosphate that was released from the substrate ATP during the reaction. Curves were fitted by using the logistic model. (c,d) The effect of varying concentrations of sodium p-toluenesulfonate on the ATP conversion rate using wild-type (c) and I303V MAT (d) in the presence of a 1.3-fold molar excess of l-methionine after 8 h incubation at 37 °C, pH 7.0. The consumption of ATP and the production of SAM were determined by high performance liquid chromatography (HPLC) as described in Section 3.
Figure 2.
The purification and product inhibition of wild-type MAT and I303V MAT variant. (a) SDS-PAGE (12%) analysis of the purified wild-type and I303V MAT proteins from recombinant E. coli that was cultured in a 2-l shake flask. Lane 1: molecular weight marker. Lane 2: the purified wild-type MAT. Lane 3: the purified I303V MAT variant; (b) conversion rates of l-methionine catalyzed by the wild-type and I303V MAT proteins as a function of the substrate concentration. The reaction mixture, which contained 10 mM ATP and varying concentrations of l-methionine, was incubated for 5 h at 37 °C in the presence of excess enzyme. The reaction yield was determined by measuring the phosphate that was released from the substrate ATP during the reaction. Curves were fitted by using the logistic model. (c,d) The effect of varying concentrations of sodium p-toluenesulfonate on the ATP conversion rate using wild-type (c) and I303V MAT (d) in the presence of a 1.3-fold molar excess of l-methionine after 8 h incubation at 37 °C, pH 7.0. The consumption of ATP and the production of SAM were determined by high performance liquid chromatography (HPLC) as described in Section 3.
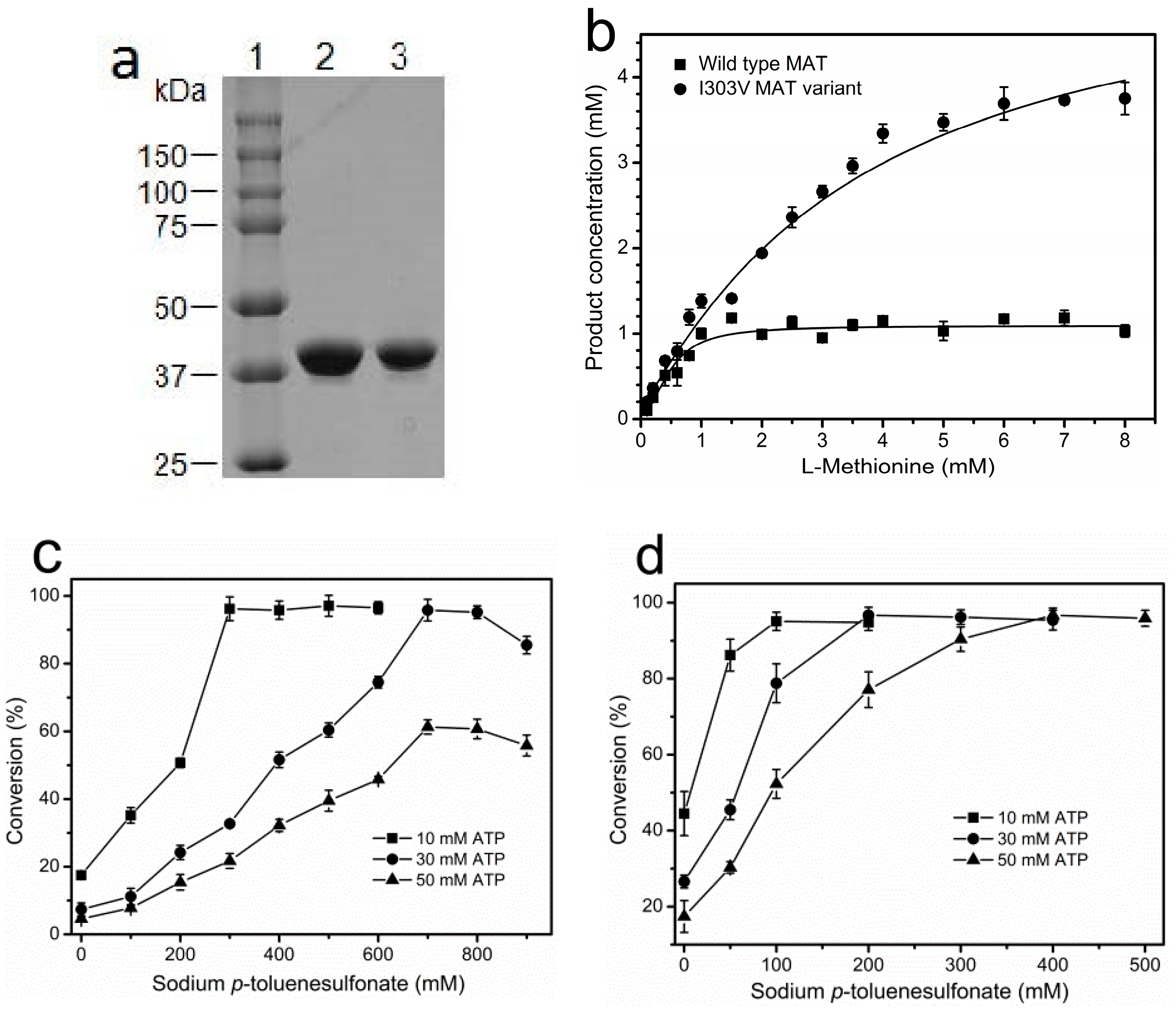
Figure 3.
Fermentation and partial purification of recombinant I303V MAT overexpressed in E. coli. (a) The effect of induction time on cell growth and yield of the recombinant enzyme. The recombinant E. coli was induced by 0.1 mM IPTG; and (b) SDS-PAGE (12%) analysis of the purification of the I303V MAT variant. Lane 1: molecular weight marker. Lane 2: total protein of recombinant E. coli before IPTG induction. Lane 3: total protein of recombinant E. coli after IPTG induction. Lane 4: the supernatant of recombinant E. coli disrupted by sonication. Lanes 5 and 6: I303V MAT partially purified by fractionation with 1 M ammonium sulfate. The position of I303V MAT protein was marked with a red arrow.
Figure 3.
Fermentation and partial purification of recombinant I303V MAT overexpressed in E. coli. (a) The effect of induction time on cell growth and yield of the recombinant enzyme. The recombinant E. coli was induced by 0.1 mM IPTG; and (b) SDS-PAGE (12%) analysis of the purification of the I303V MAT variant. Lane 1: molecular weight marker. Lane 2: total protein of recombinant E. coli before IPTG induction. Lane 3: total protein of recombinant E. coli after IPTG induction. Lane 4: the supernatant of recombinant E. coli disrupted by sonication. Lanes 5 and 6: I303V MAT partially purified by fractionation with 1 M ammonium sulfate. The position of I303V MAT protein was marked with a red arrow.
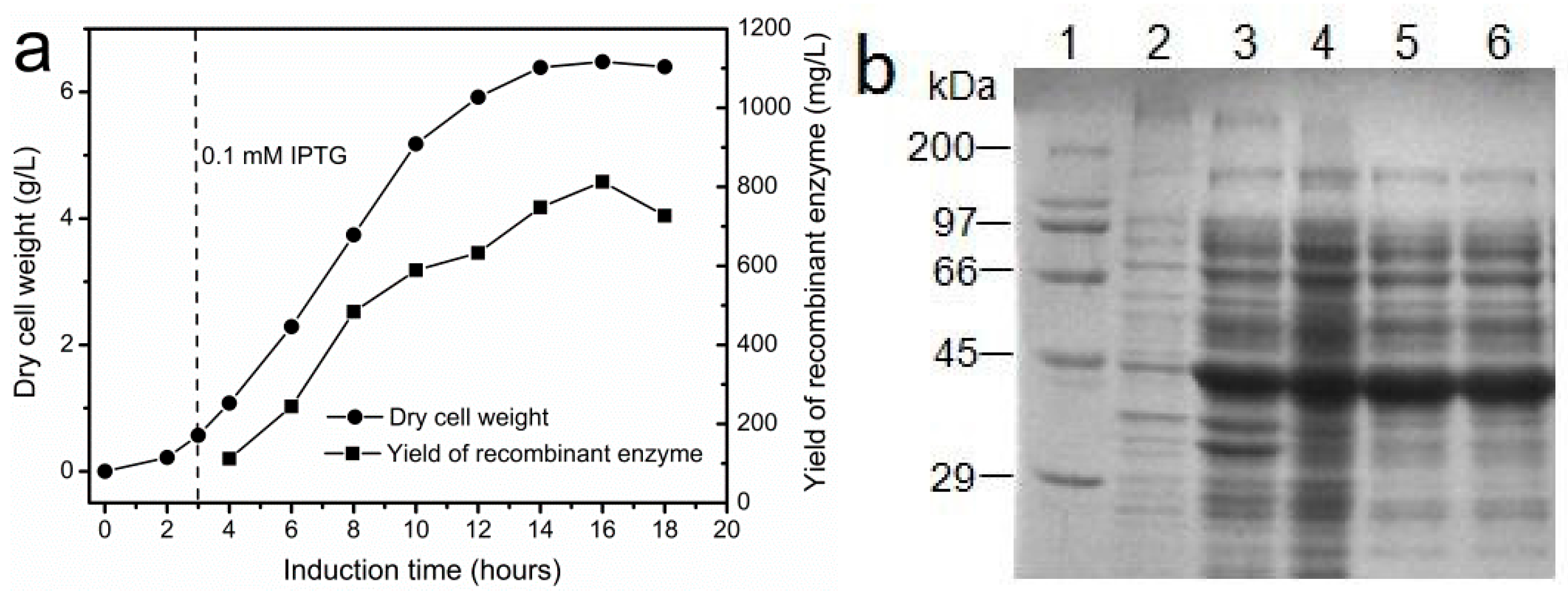
Figure 4.
Schematic illustration of the two different covalent methods of the methionine adenosyltransferase (MAT) immobilization.
Figure 4.
Schematic illustration of the two different covalent methods of the methionine adenosyltransferase (MAT) immobilization.
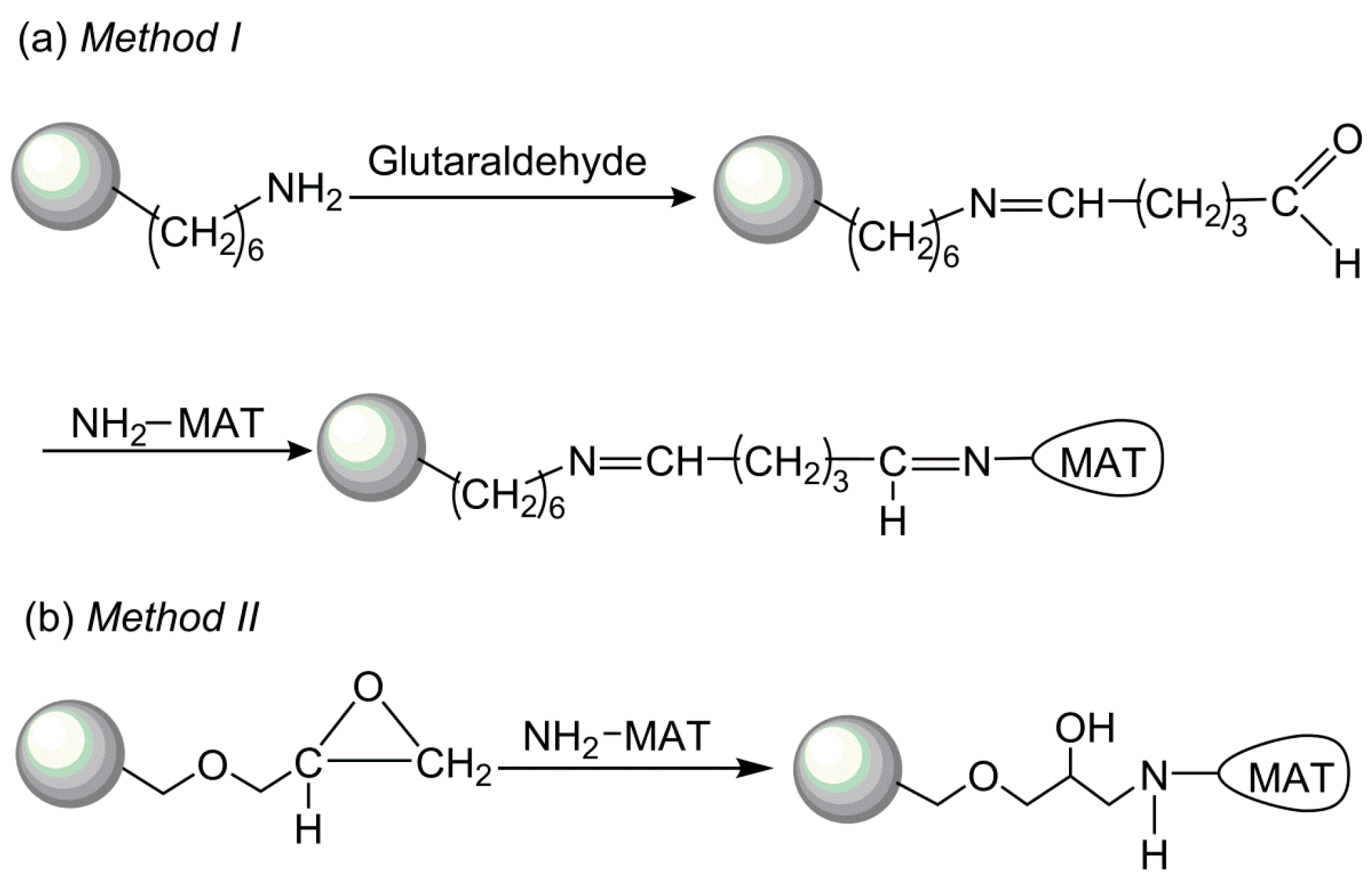
Figure 5.
The effects of temperature and pH on the stability of the free and immobilized I303V MAT. (a) Percent enzymatic activity remaining as a function of time for the free and immobilized I303V MAT. The thermal stability of the free and immobilized I303V MAT for a 168 h incubation at 37 °C. Assay conditions: 50 mM Tris-HCl buffer, pH 7.5. Initial activities were defined as 100%; and (b) the effect of pH on the stability of the free and immobilized I303V MAT. The activity of free and immobilized enzyme treated at pH 7.5 was defined as 100%.
Figure 5.
The effects of temperature and pH on the stability of the free and immobilized I303V MAT. (a) Percent enzymatic activity remaining as a function of time for the free and immobilized I303V MAT. The thermal stability of the free and immobilized I303V MAT for a 168 h incubation at 37 °C. Assay conditions: 50 mM Tris-HCl buffer, pH 7.5. Initial activities were defined as 100%; and (b) the effect of pH on the stability of the free and immobilized I303V MAT. The activity of free and immobilized enzyme treated at pH 7.5 was defined as 100%.
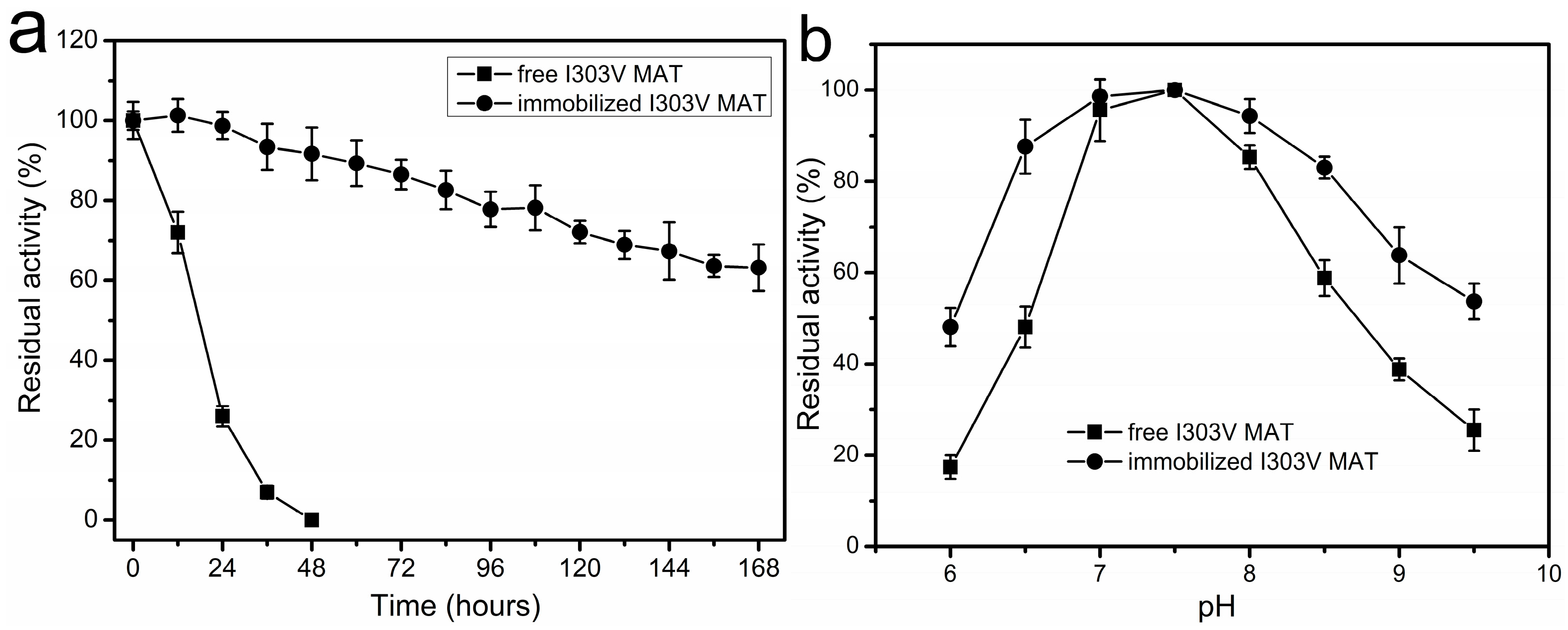
Figure 6.
Recycling of immobilized I303V MAT biocatalyst in the synthesis of SAM. (a) The effect of repeated use on activity of immobilized I303V MAT enzyme. The spectrophotometric assay was used to measure the residual activity; and (b) the synthesis of SAM with immobilized enzyme in 10 repeated batches. The production of SAM and the substrate ATP were determined by HPLC analysis. The reaction was carried out in 100 mM Tris-HCl buffer containing 50 mM K2SO4, 100 mM MgSO4, 50 mM ATP, 65 mM l-methionine, and 0.4 M sodium p-toluenesulfonate, pH 7.0 with an 8 h incubation at 37 ℃.
Figure 6.
Recycling of immobilized I303V MAT biocatalyst in the synthesis of SAM. (a) The effect of repeated use on activity of immobilized I303V MAT enzyme. The spectrophotometric assay was used to measure the residual activity; and (b) the synthesis of SAM with immobilized enzyme in 10 repeated batches. The production of SAM and the substrate ATP were determined by HPLC analysis. The reaction was carried out in 100 mM Tris-HCl buffer containing 50 mM K2SO4, 100 mM MgSO4, 50 mM ATP, 65 mM l-methionine, and 0.4 M sodium p-toluenesulfonate, pH 7.0 with an 8 h incubation at 37 ℃.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Influence of the quantity of enzyme in contact with the support for the different immobilization procedures. 0.5 g support, 50 mM Tris-HCl buffer, pH 7.5 at 4 °C.
Table 1.
Influence of the quantity of enzyme in contact with the support for the different immobilization procedures. 0.5 g support, 50 mM Tris-HCl buffer, pH 7.5 at 4 °C.
| Support | Enzyme Added (mg/g Support) | Enzyme Loading (mg/g Support) | Enzyme Coupling Yield (%) | Enzyme Activity (U/g Support) | Activity Coupling Yield (%) |
|---|---|---|---|---|---|
| LX-1000 HA (Method I) | 10 | 8.9 ± 0.5 | 89 | 41.7 ± 3.6 | 83.6 |
| 20 | 18.3 ± 1.1 | 91.5 | 76.4 ± 6.4 | 74.5 | |
| 30 | 27.7 ± 0.8 | 92.3 | 109.6 ± 7.6 | 70.7 | |
| 40 | 33.2 ± 1.2 | 83 | 104.9 ± 8.9 | 56.4 | |
| 50 | 35.4 ± 1.6 | 70.8 | 108 ± 7.7 | 54.5 | |
| LX-1000 EP (Method II) | 10 | 9.1 ± 0.4 | 91 | n.d. | n.d. |
| 20 | 17.5 ± 1.2 | 87.5 | 3.3 ± 0.36 | 3.3 | |
| 30 | 25.4 ± 0.5 | 84.7 | 3.9 ± 0.29 | 2.7 | |
| 40 | 30.8 ± 0.8 | 77 | 2.7 ± 0.21 | 1.5 | |
| 50 | 32.7 ± 1.2 | 65.4 | 3 ± 0.23 | 1.6 |
Specific activity of partially-purified free I303V MAT is 5.6 ± 0.4 U/mg protein. n.d.: not detectable.
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Niu, W.; Cao, S.; Yang, M.; Xu, L. Enzymatic Synthesis of S-Adenosylmethionine Using Immobilized Methionine Adenosyltransferase Variants on the 50-mM Scale. Catalysts 2017, 7, 238. https://doi.org/10.3390/catal7080238
AMA Style
Niu W, Cao S, Yang M, Xu L. Enzymatic Synthesis of S-Adenosylmethionine Using Immobilized Methionine Adenosyltransferase Variants on the 50-mM Scale. Catalysts. 2017; 7(8):238. https://doi.org/10.3390/catal7080238
Chicago/Turabian StyleNiu, Weining, Shanshan Cao, Menglin Yang, and Le Xu. 2017. "Enzymatic Synthesis of S-Adenosylmethionine Using Immobilized Methionine Adenosyltransferase Variants on the 50-mM Scale" Catalysts 7, no. 8: 238. https://doi.org/10.3390/catal7080238
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.