Biocatalytic Synthesis of Fungal β-Glucans



1
Biotechnology Laboratory, School of Chemical Engineering, National Technical University of Athens, 9, Heroon Polytechniou Str., Zographou Campus, 15780 Athens, Greece
2
Biochemical and Chemical Process Engineering, Division of Sustainable Process Engineering, Department of Civil, Environmental and Natural Resources Engineering, Luleå University of Technology, SE-97187 Luleå, Sweden
*
Author to whom correspondence should be addressed.
†
These authors have contributed equally in this work.
Catalysts 2018, 8(7), 274; https://doi.org/10.3390/catal8070274
Submission received: 4 May 2018
/
Revised: 25 June 2018
/
Accepted: 4 July 2018
/
Published: 6 July 2018
(This article belongs to the Special Issue Biocatalysis for Industrial Applications)
Abstract
:Glucans are the dominant polysaccharide constituents of fungal cell walls. Remarkably, these major bioactive polysaccharides account for the beneficial effects that have been observed by many mushrooms of medicinal interest. Accordingly, the prevailing tendency is the use of bioactive mushroom β-glucans mainly in pharmaceutical industries or as food additives, since it seems that they can be involved in meeting the overall growing demand for food in the future, but also in medical and material sectors. β-(1,3)-Glucan synthase (GLS) is the responsible enzyme for the synthesis of these important polysaccharides, which is a member of the glycosyl transferase (GT) family. For optimizing the production of such natural polymers of great interest, the comprehension of the fungal synthetic mechanism, as well as the biochemical and molecular characteristics of the key enzyme GLS and its expression seem to be crucial. Overall, in this review article, the fungal β-glucans biosynthesis by GLS is summarized, while the in vitro synthesis of major polysaccharides is also discussed, catalyzed by glycoside hydrolases (GHs) and GTs. Possible future prospects of GLS in medicine and in developing other potential artificial composite materials with industrial applications are also summarized.
1. Introduction
Polysaccharides are polymers of natural origin, consisting of sugar monomers connected with glycosidic linkages. Aside from their universal presence in all living organisms, several polysaccharides with specific properties are emerging as alternative bio-based products for various purposes, mainly in medicinal, pharmaceutical, cosmetic, as well as in chemical sectors. The exploitation of such renewable resources in relevant applications is consistent with the trend to minimize the use of petrochemical plastics [1]. Together with the aim of carbon neutrality these novel materials are considered environmentally friendly and sustainable [1]. Actually, bio-based polymers are considered an attractive renewable alternative to fossil fuel resources.
There are several reports regarding the enzymatic synthesis of natural polysaccharides in vitro. However, the polymerization procedure seems to be extremely complicated, as the starting monomers that have been used so far require specific modifications [2,3,4]. Therefore, a robust glucan synthesis system is not yet available, although the demand for a similar methodology is great, due to the research efforts to elucidate the biological role of carbohydrates in general. The controlled synthesis of carbohydrates could be of significant value in the efforts to study and exploit their natural function. In this respect, the standard chemical synthesis methods do not suffice, while the natural enzymes responsible for biological production of oligosaccharides, glycosyl transferases (GTs), have not been efficiently produced so far. Specifically, the widespread use of GTs is hindered at the moment, because of two reasons. Firstly, the expression of the membrane-bound enzymes is problematic. Secondly, the nucleotide–phosphate substrates, essential for transferases, are usually too costly for large-scale reactions. Meanwhile, the ideal process would take advantage of both the substrate specificity and the stereoselectivity of these enzymes.
It is interesting to report that β-(1,3)-glucans, which are the most prevalent polysaccharides observed in fungi, together with chitin, demonstrate a wide range of useful applications mainly in the industry of pharmaceuticals as they exhibit various biomedical functions, as well as in foods, cosmetics and chemical industries [5]. Alternative attempts to achieve high-yields of β-(1,3)-glucan synthesis include the in vitro synthesis of these polymers [6], or the adjustment of the culture medium that is used for the microorganism growth [7,8,9]. Along with the above, the increase of β-(1,3)-glucan synthase (GLS) expression which is the key enzyme in the β-(1,3)-glucan synthesis [10], as well as GLS gene overexpression using promoter swapping [11] seem to be promising alternatives for higher productivities. Therefore, the main objective of this review article is the relevant research on GLS, summarizing previously published studies, since it is regarded as a largely unexplored biotechnological tool. Specifically, an attempt to deeply comprehend the synthesis mechanism of the valuable β-(1,3)-glucans in fungi, catalyzed by GLS is carried out, by summarizing the available data regarding the biology of GLS, including the molecular architecture, together with all the available information regarding the biochemical characteristics of this enzyme; substrate specificity, optimum reaction conditions, effect of divalent metals, stimulators and inhibitors. We also present the biomedical applications of GLS which is of high importance, since GLS is a popular target for human antifungal therapy. Finally, we include data considering the in vitro synthesis of major polysaccharides by using genetically modified glycoside hydrolases (GHs) and GTs for synthesizing and improving β-glucan production in a controlled manner.
2. Glucans
The main structural polysaccharides in most fungal species are glucan and chitin [12]. There are two groups of glucans (α- and β-glucans; Figure 1), as specified by linkage type. Glucans are major cell wall polysaccharides, but they can also be found intracellularly, or they can be secreted into the medium.
The basic components of the mushroom cell walls are β-glucans. It is well reported that β-glucans can be of use in a wide variety of industrial applications including food supplements, pharmaceuticals, cosmetics, chemicals, food and feed industries [5]. Specifically, considering their biomedical attributes, β-glucans provide antioxidant properties [13], radical scavenging, antitumor, anti-inflammatory, antimicrobial, cardiovascular, but also antidiabetic and liver-protecting effects [5,14,15]. Consequently, research efforts are focusing on the elucidation of their biological roles, along with the design of new drugs or dietary supplements based on β-glucan as the active ingredient [16]. Recently, in the Review paper conducted by Zhu et al. [5], it has been reported that that these valuable polysaccharides are involved in present, as well as in future food and healthcare-medical industries globally. The amount of soluble β-glucan detected in different mushroom species is variable. For instance, the soluble β-glucan content in edible mushrooms species belonging to Pleurotus genus, as well as in the medicinal Shiitake mushroom (Lentinus edodes) varies between 0.6 and 1.5% (dry weight assessment) [17]. Several studies report a certain level of correlation between the glucan content of the mushrooms and the cultivation conditions, for example the carbon/nitrogen ratio and the pH of the medium, as well as the temperature of the process [18]. The polypore mushroom Sparassis crispa, which is a powerful producer of β-glucan has been found to contain up to 40% soluble β-glucan, on a dry matter basis [19].
Actually, β-(1,3)-d-glucan is used as a collective term, including two types of glucan molecules: (a) those with β-(1,3) linkages exclusively, and (b) those with β-(1,3) linkages together with a variable degree of β-(1,4) or β-(1,6) glycosidic bonds (Figure 1). Typically, six β-glucan types have been identified. β-(1,3)-Glucans with a variable but high degree of β-(1,6) branching are the most common [20]. The common name of these individual molecules is given according to their source they come from (i.e., lentinan, scleroglucan, curdlan) [21].
β-(1,3)-Glucans are the dominant components of the majority of fungal cell walls [22], accounting for approximately 65–90% of the overall β-glucan content [23]. This type of glucan can also appear bonded to β-(1,6)-glucan in branching form [24]. β-Glucans with branched structure can be either strongly bound to the cell wall or lightly connected, resulting in gelatinous material around the fruiting body [25,26]. The biological function of these compounds is varied, according to the branching degree, the molecular weight and the secondary structure of the molecule. In other words, the structural features, as well as the localization of the polysaccharides in the cell are highly important for defining their biological activity. Indeed, different physiological roles have been assigned for the intracellular β-glucans in comparison with the extracellular ones [27].
It is of special interest that the structural complexity of the molecules of β-glucans reflects the biological complexity of the sources from which they originate. Hence, mushrooms have higher complexity of β-glucan molecules than yeasts and bacteria [17]. This creates a requirement for further research regarding the action of β-glucans coming from different sources. Interestingly, many Basidiomycetes have grown and utilized agro-industrial wastes as substrates which, in some cases, could enhance the production of β-glucans by the induction of certain enzymes, such as GLS [10].
3. In Vivo Synthesis of Fungal β-Glucans by β-(1,3)-Glucan Synthases
The high level of variability in the glucan molecules corresponds to a high complexity, reflected to the synthetic mechanism of β-glucans at the fungal cell wall. The β-glucan synthesis mechanism is a topic under ongoing investigation, but the number of enzymes needed for the completion of the process is a major drawback towards this goal [24,28]. Moreover, the significant differences in structure that the GLS display compared with other β-GTs, indicate their distinct phylogenetic origin [29].
The overall process of β-glucan synthesis presents some similarities to chitin synthesis: first, the β-glucan chains (up to 1500 glucose monomers) synthesized in the cytoplasm, are transferred to the periplasmic space by a transmembrane enzyme complex [26,30]. Further modifications of the polymer structure take place in the periplasmic space. Specifically, for the construction of a resistant cell wall, β-(1,6)-glycosidic side branches are added, which account for approximately 3–10% of the total glucan bonds, connecting several β-(1,3)-glucan chains together [23,31]. These data have been derived from a very small number of glucan synthases of fungal origin purified in a certain degree [32].
As mentioned above, the completion of such a complex network of polymers, composed of β-(1,3) chains combined with a variable degree of β-(1,6) branching is a multi-step process. The main reaction steps are the initiation, the elongation of glucan chains and the branching step. Chain elongation is the only reaction investigated in-depth to date, where uridine diphosphate glucose (UDPG) is used as the glycosyl donor from glucan synthases [20]. In fungi, the GLS (EC.2.4.1.34, UDP-glucose:1,3-β-d-glucan 3-β-d-glucosyl transferase), a membrane protein complex, is the catalyst for the reaction of β-(1,3)-glucan synthesis in vivo, using UDPG as substrate (Figure 2) [33].
The elongation is proceeding with the addition of UDPG monomers to the non-reducing end of the growing glucan chain. In this process, each new polymer is driven from the plasma membrane to the cell wall, through a channel of transmembrane domains [34]. As GLS seems to be deeply involved in the process of fungal cell wall synthesis, it is constantly the focus of intensive study. In particular, this enzyme is crucial for cell wall structure because the inhibition of GLS activity affects irregularly the cell wall forming process and thus the morphology of hyphae [35].
GLS is a plasma membrane-associated enzyme with multiple transmembrane domains [36,37,38]. The enzyme catalyzing the β-(1,3)-glucan biosynthesis reaction in fungi utilizes cytoplasmic UDPG as a substrate and adds glucose residues to the growing linear glucan polymer [39]. Notably, the research group of Shematek [40] reported that the Saccharomyces cerevisiae GLS, similarly to chitin synthase, is placed in the cell membrane, towards the cytoplasm.
The GLS molecular weight in fungi has been reported to be around 200 kDa [30]. Τhe GLS complex is made up of two different subunits: a catalytic (FKS1 and FKS2) and a regulatory (RHO) subunit of lower molecular weight, with guanosine triphosphate (GTP)-ase activity [41]. FKS1 is expressed during the vegetative stage of growth under optimum conditions, while FKS2 is induced under environmental stress, such as carbon source limitation, sporulation [30], or when the cells are exposed to high temperatures. Recently, one more FKS gene (FKS3) has been reported in S. cerevisae. However, the exact FKS3 function remains unknown. Probably, FKS3 is involved in the regulation of glucan synthesis during the development of the ascospore wall [42]. Typically, FKS catalyzes the polymerization of glucose monomers. RHO is a regulatory G-protein, whose role is the activation of FKS through dephosphorylation of GTP (Figure 3). ROM, a wall GDP-GTP exchange factor protein, modulates RHO activity. In particular, ROM is activated when changes in the cell wall of fungi take place, resulted from conditions of environmental stress [10,20,43]. Generally, ROM is activated when alterations of the fungal cell wall occur, or when the cells are exposed to environmental stress factors [43]. The domain responsible for catalysis is one of the cytosolic domains of FKS, and thus the area where β-glucan synthesis takes place is the cytosol. Later, the biomolecules are driven to the periplasmic space, where they are either incorporated into the fungal cell wall or secreted outside the cell [41]. For example, CFks1p from Cordyceps militaris was predicted to possess a large domain of hydrophilic nature, in the center of the protein, facing the cytoplasm, which is probably related to catalysis [44]. The two transmembrane domains compose of six and seven transmembrane α-helices, respectively. The size of the Open Reading Frames (ORFs) coding for GLS is usually around 5–6 kb, corresponding to around 1600 amino acid residues. The fks together with rho genes are usually highly similar in most fungal genomes, where they can be present as one/two copies, though the available data regarding the regulation of their expression are insufficient.
3.1. Occurrence of β-1,3-Glucan Synthases
Many studies describe β-(1,3)-glucans biosynthesis in Phytophtora cinnamomi [45], S. cerevisiae [46,47], Aspergillus nidulans, Achlya ambisexualis [48] and Candida albicans [49].
Additionally, GLS activity has been investigated in saprophytic mushrooms such as Agrocybe aegerita, Auricularia auricula-judae, species of the genus Pleurotus (e.g., P. eryngii, P. ostreatus, P. ferulae, P. nebrodensis and P. pulmonarius), as well as Stropharia aeruginosa and the Shiitake mushroom (L. edodes) [10,50]. Furthermore, GLS has been the object of research focus in many other cases. Particularly, the filamentous Ascomycetes including Neurospora crassa [51,52,53], Botrytis cinerea [54], Pyricularia oryzae [55], Paracoccidioides brasiliensis [56], A. nidulans [57], and Aspergillus fumigatus [58,59], are some examples. Besides, there has been relative reports regarding non-filamentous Ascomycetes (Schizosaccharomyces pombe [60] and C. albicans [39,61,62]). Finally, GLS has been investigated in Cryptococcus neoformans [38], Schizophyllum commune [53,63], Sclerotium glucanicum [64], which belong to Basidiomycetes, but also in Phytophthora spp. [65,66], Saprolengia monoica [67,68], Aphanomyces astaci [69] and A. ambisexualis [53,63], classified in Oomycetes, which are not considered true fungi.
3.2. Molecular Organization of β-(1,3)-Glucan Synthase Genes
While the available data about the molecular mechanisms that are involved in the synthetic process of β-(1-3)-d-glucans are sparse, the corresponding genes (gls) are well characterized. Actually, the significant progress in molecular and genetic biology has contributed greatly to the discovery of various genes with high homology to the yeast fks1 gene [36], probably encoding S. cerevisae’s catalytic subunit of GLS. In the majority of genomes of fungal origin, fks, as well as rho genes usually exist in one/two copies, and they are highly homologous [70].
In S. cerevisiae, the most well-studied microorganism in respect to GLS molecular biology, there are two fks genes, as well as in L. edodes, Auricularia auricula-judae and P. ferulae [10], while in the strains C. albicans, C. neoformans, A. fumigatus, Ustilago maydis, P. ostreatus and P. eryngii, a single gene copy has been found [10,11,38,41,70,71]. According to a recent classification from Yang et al. [72], Ascomycetes gls genes are widespread mostly in the subphylum Pezizomycotina, while for Basidiomycetes, gls genes are distributed to four subphyla, with the majority of them belonging to subphylum Agaricomycotina, class Agaricomycetes. This specific class of Basidiomycetes, including Agaricus bisporus, Trametes versicolor, S. commune and Coprinopsis cinerea, is the only class to include two different types of GLS. The restricted distribution of this feature, might reflect a recent duplication event in the common ancestor of several Agaricomycetes species, and divergent evolution of the resulting genes [72].
In the same study, a multiple sequence analysis has revealed similar and variable domains in the two gls gene types: while the transmembrane domains were highly similar in both types, the loops were found to contain signature sequence motifs that are different for the two types (Figure 4). For example, a similar sequence motif (AMFNGXNPFAXE/D) was found in loop L9, not present in Type II genes. Accordingly, a sequence motif discovered in L13 (WXK/R) was found to exist only in Type II gene family.
Regarding the transcriptional regulation of these genes, major differences can be found among different organisms. While in yeasts stress-causing agents has been observed to induce the expression of gls genes [73], in most fungi the expression of these genes has been found to be constitutive and irrelevant to stress induction and developmental stage. In this respect, the corresponding genes in U. maydis were not overexpressed neither during infection of maize, nor under ionic and osmotic stress [70], while most mushrooms studied by Reverberi et al. [10], like A. aegerita, S. aeruginosa, P. ferulae, A. auricula-judae, P. pulmonarius and P. nebrodensis, also did not show a different expression pattern in response to environmental stress. Moreover, growth in olive oil mill waste was not found to affect significantly the production of either intracellular or extracellular β-glucans from P. ostreatus, Pleurotus citrinopileatus and Ganoderma lucidum [74]. However, addition of olive oil mill waste to the growth medium seemed to enhance the production of GLS activity from the strains L. edodes and P. ostreatus in another study [10]. It seemed that the phenolics present in the medium are the enhancers of β-glucans production instead of the lipid content of the waste material. It was hypothesized that, in response to stressful environmental conditions such as the oxidative environment generated by phenolics and reactive oxygen species generated during the decomposition of the phenols by oxidoreductive enzymes, gls genes are activated in order to modify the cell wall structure. Therefore, β-glucans may act as an antioxidative protection for these mushrooms. This is in accordance with the findings regarding Beauveria bassiana [75], where the expression levels of gls genes were correlated to cell wall thickness. This entomopathogenic fungus seems to down-regulate gls gene expression during the infection of its host, in order to minimize possible antigen display on its surface.
Reports on molecular cloning and overexpression of gls genes are sparse, probably because of the very big size of the protein complex, together with the transmembrane topology. Both these attributes are damaging for most host systems. However, Chai et al. [11] succeeded in overexpressing the relevant genes of P. ostreatus by swapping the native promoter with the constitutive promoter of glyceraldehyde-3-phosphate dehydrogenase (gpd) from A. nidulans. Thus, they obtained from 32 to 131% higher β-glucan yield in the supernatants of six transformants. Heterologous expression of the entire fks gene of A. fumigatus to a baculovirus expression system has failed, but an internal hydrophilic protein domain was successfully expressed in E. coli [41]. In the same study, authors managed to express heterologously the RHO protein in Pichia pastoris host cells. Besides, Ha et al. [76] achieved the heterologous expression of the gls gene from Fusarium solani (FsFKS1) to a knockout strain of A. fumigatus. Their findings support the functional expression of FsFKS1 gene in A. fumigatus, due to the recovery of caspofungin resistance by the transformant. The authors explained their results with the hypothesis that the heterologous FsFKS1 was responsible for proper cell wall structure and was thus responsible for the recovery of the antibiotic resistance.
3.3. Biochemical Features of β-1,3-Glucan Synthases
According to CAZy Database, GLS are GTs of the family 48, [77], which is composed of various enzymes, such as fungal FKS homologues from S. cerevisiae and callose synthases. These enzymes are involved in the β-(1,3)-glucan biosynthesis in plants, algae and Chromista. The GLS structure has been analyzed in silico and the results showed that while they share high homology, they also seem to display significant structural dissimilarities to the other β-glycosyl tranferases, independently of their origin [26].
It is of paramount importance to address that most of the available data regarding the synthesis of glucans, corresponds to the chain elongation step, and most relevant studies have been performed with crude enzyme preparations. Additionally, it seems necessary to outline that among the different systems studied, a noteworthy variability is apparent, regarding the features of the different GLS [20,78].
Another point to report is that fungal GLS enzymes share only a few common biochemical properties, such as a variable substrate specificity [guanosine diphosphoglucose (GDPG), UDPG, adenosine diphosphoglucose (ADPG)], optimal pH (between 5.8 and 7.8) and temperature (between 20 °C and 37 °C), but also inhibition or activation by several divalent metal ions, including Mg+2, Ca+2 and Fe+2 [20]. Moreover, the value of the respective Michaelis constant (KM) for the substrate, seems to vary depending mostly on the microorganism studied. In addition, it has been observed that the experimental procedures applied play an important role on determining the biochemical characteristics of GLS studied. After all, when comparing the biochemical properties of GLS from different sources, it is apparent that they ultimately represent a quite heterogeneous enzyme group [20].
Three significant biochemical characteristics of the GLS (KM, pH, temperature) from different sources are briefly outlined in Table 1. The heterogeneity observed seems to be the result of the species variability, as well as the experimental procedure applied in each case. In the following paragraphs, biochemical characteristics of GLS from different sources are described in more detail, as reported previously.
3.3.1. Substrate Specificity
The first report considering the β-(1,3)-glucans synthesis in vitro was performed by the Sentandreu research group in 1975 [86]. Specifically, Sentandreau et al. [86], used S. cerevisae protoplasts permeabilized by toluene for permitting the entrance of the substrate. Two years later, Lopéz Romero and Ruiz Herrera [83], attempted the synthesis of large amounts of β-(1,3)-glucans, examining different experimental conditions, in cell-free extracts. Later studies regarding several fungal species concluded that UDPG represents the typical glycosyl donor. However, there are cases where GDPG or ADPG serve as sugar donors [56,87,88].
Typically, GLS from different systems are membrane-associated complexes, utilizing UDPG as the usual substrate for the linear polysaccharide synthesis, a reaction that seems to be processive and results to the production of increasing chains. In particular, the suggested mechanism of the reaction seems to be the addition of one mole of glucose for each mole of hydrolyzed UDPG [40,80]. This proposed mechanism is in direct contrast with that of non-processive GTs, where sugar residues (mono- or oligosaccharides) are transferred to acceptor substrates such as proteins, lipids or other sugars [89].
As it is obvious from Table 1, most GLS enzymes use UDPG as a substrate, exhibiting a KM range of 0.12–4.5 mM in crude enzyme preparations. Actually, all GLS possess a KM of a few millimoles, with the exception of GLS derived from P. brasilliensis, the KM of which is in the micromolar range. However, significantly higher KM values are determined in the cases of A. ambisexualis (7.1 mM), and Pleurotus sojae (10.6 mM, Table 1). Thereby, it seems that the GLS KM value regarding Oomycetes species stands as a point of difference compared to the rest fungal GLS.
Regarding this subject, Wang and Bartnicki-Garcia [82] managed to synthesize in vitro β-(1,3)-glucans. In this study, the synthesis of β-(1,3)-β-(1,6) branched glucan from UDPG was documented, with P. cinnamomi cell-free extracts as the catalyst. The affinity of GLS for UDPG seemed to be affected by three factors, namely the system under study, the fungal growth stage, and the reaction conditions. As shown in the Table 1, the lowest UDPG KM value detected is 1.25 × 10−4 mM, corresponding to the GLS of the pathogenic fungus P. brasiliensis [56].
It is concluded that the growth conditions, as well as the stage of development of the microorganism affects significantly the value of KM and therefore, there is a large degree of variance concerning this biochemical characteristic. The possible reason for this observation, is the level of stimulation or inhibition of the GLS activity that arises from the presence of certain compounds in the crude extracts [26]. For example, regarding the case of P. palmivora, there are significant differences in the KM values calculated for the different fractions derived from differential centrifugation of cell-free extracts. The presence or absence of cellobiose in the process also affected the calculated KM values [82].
3.3.2. Optimal Reaction Conditions
Optimal pH values for GLS studied in different systems are detected in the range of 5.8 to 7.8 (Table 1). As it has been demonstrated, these values depend, to a certain extent, on the buffer used. Particularly, phosphate buffer has been proven to be the best one comparing with other buffers, whereas the optimal temperature for GLS varied between 20 °C and 30 °C [26,40,56,88]. The lowest optimum pH determined was at pH 5.8 for the Oomycete S. monoica [68], while the highest one was found at pH 7.8 for the GLS of P. palmivora [82]. Concerning the optimal temperature, it is interesting to outline that 20 °C was found to be the most appropriate in the case of P. oryzae [55], whereas 30 °C was determined the most favorable for the GLS from the pathogenic yeast C. albicans [39].
3.3.3. Effect of Divalent Metals
GLS activity can be either stimulated or inhibited by divalent metals. Typically, the presence of divalent metals (e.g., Mg+2, Mn+2, Ca+2, and Fe+2) has been proven to stimulate GLS activity. This is mostly observed in the cases considering fungal species belonging to genus Phytopthora [81,82], S. monoica [90] and P. brasiliensis [56]. Mg+2 and Mn+2 ions seem to be the most efficient ones [26], but there is a large degree of variance concerning the response of those enzymatic systems in which particular metals seem to function as activators. Specifically, this depends on the particular species studied, its developmental phase, as well as the physicochemical characteristics of the final product. Considering the synthesis of glucans from the P. palmivora extracts, Mg+2 activated the alkali-insoluble glucan biosynthesis more strongly than Ca+2, while the opposite was observed during the synthesis of water-soluble polysaccharide products [82]. This is an example on how the nature of the synthesized products influences the degree of enzyme stimulation by metals. In regard to the effect of developmental stage of the microorganism on the level of GLS stimulation by metal ions, the best example is the study conducted by San Blas [56]. Specifically, in this study, Ca+2 has been the best stimulator of GLS from P. brasiliensis in yeast form, but Fe+2 served as the best metal ion for activating GLS in the mycelial stage of growth [56]. Nonetheless, regarding the case of P. sojae, the GLS was not activated by divalent metals, such as Mg+2 and Ca+2, even though some others (Cu+2 and Zn+2) seemed to act as inhibitors [65]. Finally, in the case of S. monoica, contradictory results were obtained regarding the effect of Mg+2 on the GLS activity, depending on the assay conditions. Particularly, when ATP or GTP were present in the assay, this metal ion had a stimulatory behavior. Otherwise Mg++ seemed to act in an opposite way, inhibiting the reaction [90].
3.3.4. Stimulators and Inhibitors
Typically, it is widely approved that GLS could be activated or inhibited by certain molecules [26], some of which are listed in Table 2. More specifically, studies dealing with yeasts and other fungi have demonstrated that nucleoside triphosphates could serve as GLS stimulators. Particularly adenosine triphosphate (ATP), GTP, as well as their analogues have shown stimulatory effect on GLS activity of different systems [40,53,55,61,69,85,91]. Studies of fungal GLS from different taxa agree on the necessity of GTP presence, in order to maximize GLS activity [53,63]. However, there are exceptions, such as the results of Billon-Grande et al. [67] on S. monoica, showing that GTP (or even ATP) had an inhibitory effect on GLS activity. On the other hand, considering the cases of P. sojae [65] and A. ambisexualis [63], both of which belong to Oomycetes, the GLS activity was found to be GTP-independent. Moreover, many researchers have indicated the variability of the GLS activation mechanism, based on the fact that different enzyme preparations derived from the microorganisms examined, differed to a great extent in their response to stimulation by GTP [53,64,91].
Based on the results obtained by the research group of Beauvais [58], considering the membrane extracts of A. fumigatus, five factors have been shown to stimulate GLS activity: the age of the culture, GTP, and the compounds NaF, sucrose and ethylenediaminetetraacetic acid (EDTA), while the highest levels of GLS activity were observed in the early exponential (log) phase of growth. However, this could be the result of increase of protease activity to the ageing mycelium, but in any case, the drop in GLS activity with ageing is not a common property among fungal GLS. Requirement of GTP [53] is common among fungal GLS, including Hansenula anomala, N. crassa, Cryptococcus laurentii, S. commune and Wangiella dermatitidis. It is also important to add that GTP seems to be an allosteric activator of the GLS as suggested for H. anomala and N. crassa. The evidence from the studies implying that nucleoside triphosphates could serve as GLS stimulators [40], provides the framework for understanding the mechanism engaged with the enzyme catalysis. S. cerevisae represents a model system for the elucidation of the mode of action of nucleoside triphosphates (ATP and GTP) regarding the stimulation of GLS activity [40,85]. Accordingly, it has been evidenced that the stimulation mechanism of these two nucleotides is not the same: ATP participates in the phosphorylation step, while GTP binds to the enzyme and it can be replaced by non-phosphorylating analogs. Other investigators have also contributed to clarify the probable mechanism of activation, and they concluded that phosphorylation of the enzyme is not necessary [92].
There are several studies supporting that the inorganic compound NaF could stimulate GLS activity as well. This has been observed in crude extracts from S. cerevisae cultures [47,98], in accordance with the results of Beauvais et al. [58]. This basic salt also enhanced GLS activity of a membrane fraction from the mycelium of A. fumigatus.
In this regard, prior research generally confirms that fungal GLS could be also stimulated in vitro by several disaccharides, but also BSA has been found to activate significantly GLS derived from S. cerevisae [40], C. albicans [61], N. crassa [95] and other fungi [78]. BSA could be substituted with β-lactoglobulin in the case of S. cerevisae [97].
On the other hand, previous studies revealed that the β-glucan synthesis in S. cerevisiae was competitively inhibited by the synthesis byproduct UDP [84]. This seems to be the case also for most other microorganisms studied so far [54,96]. Finally, a number of studies [54,96] have found that gluconolactone that has been described as a known inhibitor of glucanases [84], could also inhibit the activity of GLS.
3.4. Biomedical Significance
Douglas et al. [30] reported that GLS enzyme has become an attractive target for human antifungal therapy, not to mention that this enzyme is a highly similar constituent of fungal morphogenetic machinery. In addition, the same team reported that GLS enzyme is a major component of the complex control network of fungal adaptation to environmental stress and to growth signals. Particularly, amphiphilic antibiotics, such as papulacandins, echinocandins and aculeacins are the most studied inhibitors of GLS. These molecules include variable hydrophilic domains, together with fatty acid moieties, corresponding to the hydrophobic domains of the molecule. Interestingly, this hydrophobic domain is of paramount importance for activity. Echinocandins are the newest class of antifungals. As it has been suggested [100,101], echinocandins have strong effect on cell wall biosynthesis, which arises from the noncompetitive inhibition of GLS.
On this subject, a most recent study was performed, on the effects of antifungal drugs on cell wall synthesis [102]. As it has been discovered, treatment with echinocandin B (EB) gave rise to numerous effects including defects to cell polarity, high variability in size, reduction of actin levels, as well as important changes in the cell nucleus, such as increase and irregularities. Additionally, mutations in different domains of FKS result in cells with morphological differences. EB has been found to interfere with the FKS1 domains carrying the mutations. Hence, decline in β-(1,3) glucan synthesis was noticed, together with several modifications regarding cell morphology and polarity [102]. Consequently, echinocandin antifungal drugs could cause significant irregularities in cell wall integrity/structure. Typically, the above antifungal agents are administered for candidiasis and aspergillosis therapy [103]. On this matter, SCY-078, a derivative of enfumafungin, represents an innovative class of triterpenoid GLS inhibitors, structurally distinct from traditional echinocandin inhibitors. Taking into consideration that SCY-078 is an orally-available inhibitor of GLS, approved with QIDP designation, it is regarded as a first-in-class GLS inhibitor. Also, it is outstanding to mention that this novel drug exhibits in vitro inhibitory effects on β-(1,3) glucan synthesis in pathogenic fungal species, such as Candida and Aspergillus species, including several fluconazole-resistant strains [104,105]. Furthermore, SCY-078 has been observed to retain its inhibitory activity on some Candida and Aspergillus strains, regarded as echinocandin-resistant [106,107]. In regard to the case of examining SCY-078 in vitro against other molds, it has been observed that this GLS inhibitor was the only one functioning against Scedosporium prolificans, a pan-resistant fungal pathogen causing a wide variety of infections [108]. SCY-078 was recently reported to be in phase II clinical trials of drug development process for invasive candidiasis therapy, for oral administration [109]. Meanwhile, the IV formulation of the same drug has been reported to be in phase I clinical development [109].
Fungal β-glucans are an important mediator of fungal virulence, and there is vast literature concerning the role of fungal β-glucans during infection, as well as the mediation of immune response (for selected reviews, see [13,110,111,112]). Fungal β-1,3-glucans bind to C-type, lectin-like dectin 1 receptor, displayed by macrophages and dendritic cells, inducing an innate immunity response, involving the secretion of cytokines and tumor necrosis factor-α. Cytokines activate the adaptive immune response (B and T lymphocytes and natural killer cells), and resulting to a systemic activation of the whole immune system as a response to fungal attack. Most previous studies on the interaction of β-glucan–dectin-1 have been performed with β-glucans isolated from natural sources (mainly yeasts and mushrooms), but this strategy poses some obvious issues: the heterogeneity of natural β-glucan preparations in terms of composition, chemical and physical properties, can lead to contradictory results and prohibits the development of medicinal formulations of consistent composition. Nonetheless, there are a few recent reports in the literature, regarding the effects of synthetic β-glucan molecules to mammal immunity: for example, synthetic β-glucan oligosaccharides (chemically synthesized) revealed the specificity of dectin-1 receptor in regard to the β-glucan structure [113], while the conjugation of natural [114] and synthetic [115,116] β-glucan oligopeptides to protein antigens was found to greatly enhance the immune response by the host, paving the way for the design of novel vaccine formulations. As a result, the development of novel tools for controlled, custom-made synthesis of β-glucan oligo- and polysaccharides is expected to shed light on the pathogen-host interaction mechanisms, and possibly lead to the design and manufacturing of novel therapeutic agents.
4. In vitro Synthesis of β-Glucans
The prospect of synthesizing tailor-made oligo- and polysaccharides would undoubtedly bring great benefits to the fundamental and applied research in the scientific areas of Biology and Nanobiotechnology. These compounds have been reported to exhibit different biological activities. In addition, they could also have industrial applications, such as the production of artificial composites. At present, the complexity of the β-glucans synthetic machinery in vivo is hindering their application to cell-free systems, and as a result, the only economically feasible approach to large scale production of β-glucans to date is the fermentation of the producing microorganisms. However, biocatalytic approaches offer the added advantage of custom-made production of selected saccharide molecules, with controlled and consistent structure, together with the possibility of manufacturing non-natural oligo- or polysaccharide molecules. Therefore, saccharide synthesis has been gathering research attention for almost 20 years. The relevant literature includes various different approaches towards this goal, including the use of a wide array of biocatalysts, among them glucan synthases, GHs, and glycosyl phosphorylases. However, all the available enzyme systems to this date suffer from significant drawbacks, and therefore, β-glucan synthesis to this date is still far from commercialization.
4.1. Glycosyltranferases (GTs)
In contrast with polysaccharide molecules isolated from natural sources, biocatalytic synthetic approaches permit the production of homogeneous saccharide polymers with defined structure. Ideally, this technology could be developed by implementing GT enzymes from natural sources. However, their use to date is limited, due to stability issues and the necessity for costly donors. The relevant literature includes only a few reports on the in vitro synthesis of polysaccharides mediated by GTs. One example is the work of Pelosi et al. [117], implementing S. monoica and blackberry (Rubus fruticosus) detergent extracts as catalysts for the production of β-(1,3)-d-glucans, and their subsequent characterization in terms of morphology and structure. UPDG was used as the donor, and the yields obtained were in line with the requirements of analytical techniques (in the range of mg) [118]. From the results of this work, it became apparent that enzyme extracts from the same organism can produce variable polymers according to the reaction conditions, as shown by the two synthase isoenzymes of S. monoica, requiring different pH for activity [67]. All the products were characterized in detail, revealing a microfibrillar structure, however, products from S. monoica extracts were different in morphology according to the pH of the reaction. The products of R. fruticosus extracts also showed significant morphological differences.
The development of a feasible process for β-glucans biocatalytic production is presently hindered by two major impediments: the instability of the bulky transmembrane β-glucan synthase complexes, and their need for costly sugar nucleotide donors. Both these issues are unresolved to date, however, elegant novel techniques are increasingly becoming available for the tackling of these issues. For example, the need for activated donor molecules, i.e., sugar nucleotides, can be circumvented with the use of modern biocatalytic approaches, such as the production of activated donors from sucrose [119]. Leloir GTs (sucrose synthases) are gaining rising attention regarding the production of a great variety of glycosylated compounds. This system has been implemented for the production of geranyl glucoside, i.e., the glycosylation of a lipophilic molecule [120], while Schmolzer et al. [121] have recently succeeded in producing UDP-glucose from sucrose, with high yield, and in preparative scale. The co-immobilization, or the design of a cascade reaction of sucrose synthase, together with a β-glucan synthase of choice, could be a major step towards realization of a feasible process for β-glucan controlled synthesis. On the other hand, β-glucan synthases, as most transmembrane proteins, are especially difficult to isolate and study. This fact poses obvious issues on the industrialization of a β-glucan synthase—mediated glucan production. However, cutting-edge approaches from various groups attempt to develop methodologies for stabilization and/or immobilization of transmembrane proteins. There are various alternative approaches to the same problem: from the overexpression of β-glucan synthases to their native hosts, and cell lysis to obtain enzymes immobilized to the cell membranes, a technique applied to E. coli cells so far [122], to the immobilization of the membrane proteins in hydrogels [123]. To date, none of these approaches have been applied to fungal β-glucan synthases, but the consistently high research interest in the transmembrane enzyme’s biology and applications is very encouraging for future developments.
4.2. Glycoside Hydrolases (GHs)
GHs serve as a pool of glucan processing enzymes. In more detail, they are responsible for the hydrolysis of the glycosidic bonds between sugar moieties or between a sugar and another biomolecule. The Carbohydrate Active Enzymes Database (CAZy, http://www.cazy.org/) categorizes GHs to over 120 families, according to sequence identity [78]. Considering GHs function, there are two options. In native form, they can perform transglucosylation synthetic reactions, but they can also be genetically engineered to acquire “glucosynthase” activity, and thus assemble glucoside moieties towards the production of higher molecular weight saccharides.
4.2.1. Glycosynthases
The in vitro synthesis of β-glucans has also been performed with the use of GHs that are genetically modified, known as glycosynthases. One of the classic strategies includes the use of GH mutated variants, with modifications of the nucleophile amino acid responsible for catalysis, to allow for the high-yield (90%) production of oligosaccharides, according to the research conducted by Malet and Planas [124]. “Glycosynthases” is the prevailing term for these GHs that are genetically engineered, in contrast to glucan synthases (which generally belong to GT family), and it will be used here to describe this particular enzyme function.
Glycosynthases are regarded as valuable enzymes as they allow the synthesis of glucans of high interest, with potential to be used as sensors in protein-ligand interaction studies (e.g., antigen display). Due to the major medicinal significance of cell-surface displayed glucan molecules, glucosynthases may be used also as inhibitors and treatment agents on their own. Therefore, these enzymes could provide a multifunctional platform for controlled synthesis of glucan molecules. The rearrangement of glycosylation patterns in proteins with therapeutic properties using GHs and their mutated variants has been considered an astonishing frontier in both fundamental and applied research fields, with significant industrial potential [125]. Taking into account the substrate versatility that these GH families exhibit, regarding the synthesis of glycosides, they may be considered valuable instruments for the production of a great variety of glucan molecules. However, the range of glycosynthases available at present, is not considered adequate for synthesizing the whole spectrum of potential oligosaccharide research targets.
The use of glycosynthases was extended to endo-glycosidases with success [124], even though at first, they were designed on an exo-β-glucosidase [126]. It is worthy to mention that glycosynthase reactions are more preferable than transglycosylation reactions, in which wild-type enzymes are used as catalysts. The main advantage is that in glycosynthase reactions, hydrolysis of the final product by the hydrolytically inactive variant is not observed. Indeed, glycosynthases are widely regarded as innovative tools aimed at the production of novel saccharide molecules, carrying specific, as well as controlled structures [127,128,129]. They also represent another alternative for wild-type synthases, to be used in research efforts aiming at the elucidation of the polymerization mechanism and the structure of high-molecular weight polysaccharides, as well.
A first example to report is the mutant glycosynthase derived from Bacillus β-(1,3)-(1,4)-d-glucanase, previously isolated and studied by the research teams of Mallet and Planas [124] and Faijes et al. [130]. As it was revealed, this glycosynthase was capable to catalyze successfully condensation reactions of donor and acceptor sugar moieties bearing different polymerization degrees. The data gathered by Faijes et al. [131], showed that glycosynthases could serve as promising enzymatic systems, valuable for the structure-controlled production of novel polysaccharides.
Obviously, the glycosynthase technology offers an interesting prospect in regard to the control of regio- and stereo-selectivity of saccharide polymerization reactions. Indeed, endoglycosynthases could have interesting synthetic functions, such as the synthesis of oligosaccharides by coupling of donor/acceptor sugars, as well as the polysaccharide synthesis through polycondensation reaction carried out by the sugar donor without any acceptor. In the research performed by Faijes et al. [131], the second strategy has been utilized, for the structurally-controlled polysaccharide synthesis in vitro. Additionally, the technology of transglycosylation with glucosidase mutants [132,133] has been used successfully in the production process of several polysaccharides. For instance, amylose, xylan, β-glucans bearing mixed links in their molecule as well as hybrid molecules of cellulose and chitin or xylan, have been synthesized with moderate yields, due to residual hydrolase activity of the mutants. The glycosynthase strategy was extended to the polysaccharides formation, proceeding through donor self-condensation [132]. As mentioned earlier, Faijes et al. used a glucosynthase variant of a Bacillus β-(1,3)-(1,4)-glucanase for the polymerization of α-laminaribiosyl fluoride monomer to yield a glucan molecule with alternating mixed β-(1,3)- and β-(1,4)-bonds [131]. The final product is a non-natural polysaccharide, and it was characterized using nuclear magnetic resonance spectroscopy (NMR), as well as methylation analysis. Morphology analysis was also conducted with microscopy techniques. The results revealed an unfamiliar crystalline structure. Cellulose [134], laminarin [135], xylans [136], β-(1,3)-(1,4)-glucans [137], as well as non—linear xyloglucans [138], are some other polysaccharides that have been prepared with great success. Furthermore, the research team of Fort [134], studied the properties of a glycosynthase (HiCel7B E197A), obtained from a cellulase named Cel7B from Humicola insolens. This enzyme apparently was able to use various functionalized cellobiosyl donors, showing significant potential in various synthetic applications, such as xyloglucan oligosaccharide production [139]. Moreover, it has been suggested that this glycosynthase could be also used for the production of a polysaccharide with excellently alternating side chains, which after a few functionalization steps, could give rise to novel products with controlled structure and a regular side-chain pattern. Hence, polymerization reactions of simple glucosides catalyzed by glucosidases are regarded feasible, offering the additional advantage of regio- and stereospecificity for the assembling glycosidic bonds, through transglycosylation that is kinetically regulated [140].
Aside from engineering approaches, natural novel enzymes with hybrid properties might be useful in the synthesis of target compounds. For example, the research team of Takakashi et al. [141], identified and characterized a Magnaporthe oryzae endotransglucosylase (ETG), with the ability to transfer oligomers from a β-1,4-glucan molecule to an acceptor substrate. This enzyme (MoCel7B) was found to belong to family GH7. This novel ETG was capable of removing oligomers from large β-glucan molecules, and transfer them to other donors, in order to disrupt the structure of plant cell walls. In addition, depending on substrate molecular size, the reaction of endotransglucosylation was found to cause depolymerization and polymerization of β-glucans. Τhe findings of this survey provide significant information considering the de-polymerization of β-1,3-1,4-glucan caused by M. oryzae infection, as well as important insights on the application of the in vitro synthesis of cellulose, in an efficient manner.
Overall, the engineered glycosynthases can be efficient tools for in vitro analysis of the biosynthetic mechanisms of polysaccharides, as well as the structure of the final product, in contrast to wild-type synthases, which are usually membrane-associated. Moreover, glycosynthases are also very promising catalytic systems in the specific, structure-controlled synthesis of novel glucoside polymers, as well as novel biomaterials. Heterogeneous biomaterials synthesis, mediated by glycosynthases, could be achieved by including other polymers in the in vitro reaction. Thus, the production of novel composites could be obtained, due to self-assembly of the various oligomers. These novel biomaterials could subsequently be characterized in detail in terms of their structure, using techniques such as NMR spectroscopy, due to the high product yields provided by glycosynthases [131].
Despite their significant biocatalytic potential, the applicability of mutant glycosynthases on polysaccharide synthesis is still impaired by the need for glycosyl fluoride donors. Such donors are of crucial importance in the catalytic cycle performed by mutant glycosynthases because they mimic the enzyme-substrate intermediate that nucleophile knockout mutants are unable to form. Therefore, the need for costly activated donors is a major issue to the design of a large-scale process for glucosynthase-mediated polysaccharide production, as is the case for GTs. An interesting approach to overcome this issue has been developed by Pozzo et al. [142], implementing a chemical rescue technique to eliminate the need for fluoride donors. The authors engineered a thermostable GH3 β-glucosidase from Thermotoga neapolitana TnBgl3B to a β-glucosynthase, by introducing double nucleophile mutations. The mutant enzyme was able to produce a β-1,3-disaccharide product both by using a fluoride donor, and by using formate as an added nucleophile, to obtain donor regeneration. These results are very promising in regard to future developments of glucosynthases as cost-efficient catalysts for large-scale polysaccharide production.
4.2.2. Glycoside Phosphorylases (GPs)
Enzymatic glycosylation carried out by glycoside phosphorylases has been considered as another approach under study [143,144,145]. In contrast to other glycoside hydrolases, glycoside phosphorylases remove mainly glucose monomers from the non-reducing end of an oligo- or polysaccharide and transfer the monomer to inorganic phosphate, yielding a glycosyl phosphate product. The reaction catalyzed by glycoside phosphorylases is reversible, therefore these enzymes have been used widely for the synthesis of various oligo- and polysaccharides, from glycosyl phosphate and a suitable acceptor as starting material. GPs can be either inverting or retaining enzymes in regard to the anomeric configuration of the donor and the product. The characterized GPs in the literature cover the whole spectrum of glycosidic linkages, except from β-1,1 and α- and β-1,6. They are soluble enzymes, and the donors they require for synthesis are low-cost substrates; hence, they are considered especially attractive for synthetic applications. However, most of them use disaccharide acceptors, with high regio-specificity, a property not always desirable for polymerization applications. This characteristic creates the need for extensive mutagenesis and genetic engineering studies, in order to broaden the substrate spectrum of GPs [146,147], to include glucose as well as larger oligosaccharides.
Previous synthetic efforts with the use of glycoside phosphorylases have been extensively reviewed by O’Neill and Field [144]. Regarding the formation of β-(1,3) linkages, a small number of respective phosphorylases has been found, including β-1,3-glucan phosphorylase [148] and laminarin phosphorylase, from algae [148,149,150], bacterial [151,152] and plant sources [153], able to form small β-1,3-disaccharides or oligosaccharides [152]. β-1,3-Glucans of higher polymerization degrees have also been obtained, such as the highly crystalline glucan with polymerization degree of 30 [154], or the β-glucan precipitate formed by β-glucan phosphorylase OdBGP by the algae Ochromonas danica [148], using glucose-1-phosphate and laminaribiose. These efforts are of special importance towards the controlled in vitro synthesis of β-glucans. Nonetheless, it should be noted that, despite the abundance of β-1,3-glucans in the cell walls of Basidiomycetes fungi, a fungal β-1,3-glucan phosphorylase is yet to be discovered, together with a phosphorylase acting on β-1,6-glycosyl bonds, also present in many fungal cell walls.
5. Conclusions and Future Prospects
The present paper has focused on reviewing the literature concerning the mechanism of fungal β-glucans biosynthesis, aiming on the biochemical and molecular characteristics of the responsible enzyme systems, such in case of GLS. The biotechnological value of these catalysts is apparent, in view of the significant properties of the polysaccharide products regarding their potential uses in the industry of pharmaceuticals, foods and chemicals.
Aside from the designing of drugs with antifungal activity targeting the GLS action, as well as the key role of these enzymes in the growth and morphogenesis of fungal cell-walls, fundamental/applied research in a biological/nanobiotechnological scope of catalysts could also lead to the in vitro synthesis of custom-made oligo- and polysaccharides. In this respect, GHs and GTs have been described as highly promising enzymatic tools with regard to structure-controlled production of novel saccharide molecules, and novel biomaterials. While there are numerous questions still unresolved considering the biology of these enzymes, it seems that mechanisms of β-glucan synthesis will continue to attract research interest. One of the most prominent research targets is the development of in vitro technological tools for the separate examination of the different steps of β-(1,3)-glucan biosynthesis. Examples of the desired tools would be novel cloning and expression systems, enabling researchers to overcome the inherent difficulties in separating and purifying the individual catalysts participating in the β-glucan biosynthesis. Such technological advances will probably aid in the complete elucidation of the synthetic machinery, as well as the synthetic mechanism, of these significant fungal polysaccharides.
Author Contributions
A.Z. and E.T. conceived the outline of the review and, along with L.-M.P., selected the material. All authors contributed to the writing of the paper.
Funding
This research was funded by IKY Fellowships of Excellence for Postgraduate Studies in Greece-Siemens Programme.
Acknowledgments
Lefki-Maria Papaspyridi thanks the State Scholarships’ Foundation of Greece for a grant (IKY Fellowships of Excellence for Postgraduate Studies in Greece-Siemens Programme).
Conflicts of Interest
The authors declare no conflict of interest.
References
- Iwata, Τ. Biodegradable and bio-based polymers: Future prospects of eco-friendly plastics. Angew. Chem. Int. Ed. 2015, 54, 3210–3215. [Google Scholar] [CrossRef] [PubMed]
- Kobayashi, S. New developments of polysaccharide synthesis via enzymatic polymerization. Proc. Jpn. Acad. Ser. B 2007, 83, 215–247. [Google Scholar] [CrossRef] [Green Version]
- Kobayashi, S.; Ohmae, Μ. Enzymatic polymerization to polysaccharides. Adv. Polym. Sci. 2006, 194, 159–210. [Google Scholar]
- Kobayashi, S.; Sakamoto, J.; Kimura, S. In vitro synthesis of cellulose and related polysaccharides. Prog. Polym. Sci. 2001, 26, 1525–1560. [Google Scholar] [CrossRef]
- Zhu, F.; Du, B.; Xu, B. A critical review on production and industrial applications of beta-glucans. Food Hydrocoll. 2016, 52, 275–288. [Google Scholar] [CrossRef]
- Tomazett, P.K.; Félix, C.R.; Lenzi, H.L.; de Paula, F.F.; de Almeida Soares, C.M.; Pereira, M. 1,3-β-d-Glucan synthase of Paracoccidioides brasiliensis: Recombinant protein, expression and cytolocalization in the yeast and mycelium phases. Fungal Biol. 2010, 114, 809–816. [Google Scholar] [CrossRef] [PubMed]
- Bzducha-Wróbel, A.; Błażejak, S.; Molenda, M.; Reczek, L. Biosynthesis of β(1,3)/(1,6)-glucans of cell wall of the yeast Candida utilis ATCC 9950 strains in the culture media supplemented with deproteinated potato juice water and glycerol. Eur. Food Res. Technol. 2015, 240, 1023–1034. [Google Scholar] [CrossRef]
- Gern, R.M.M.; Wisbeck, E.; Rampinelli, J.R.; Ninow, J.L.; Furlan, S.A. Alternative medium for production of Pleurotus ostreatus biomass and potential antitumor polysaccharides. Bioresour. Technol. 2008, 99, 76–82. [Google Scholar] [CrossRef] [PubMed]
- Papaspyridi, L.-M.; Katapodis, P.; Gonou-Zagou, Z.; Kapsanaki-Gotsi, E.; Christakopoulos, P. Optimization of biomass production with enhanced glucan and dietary fibres content by Pleurotus ostreatus ATHUM 4438 under submerged culture. Biochem. Eng. J. 2010, 50, 131–138. [Google Scholar] [CrossRef]
- Reverberi, M.; Di Mario, F.; Tomati, U. β-glucan synthase induction in mushrooms grown on olive mill wastewaters. Appl. Microbiol. Biotechnol. 2004, 66, 217–225. [Google Scholar] [CrossRef] [PubMed]
- Chai, R.; Qiu, C.; Liu, D.; Qi, Y.; Gao, Y.; Shen, J.; Qiu, L. β-Glucan Synthase Gene Overexpression and β-Glucans Overproduction in Pleurotus ostreatus Using Promoter Swapping. PLoS ONE 2013, 8, e61693. [Google Scholar] [CrossRef] [PubMed]
- Ruiz-Herrera, J.; Sentandreu, R. Fungal cell wall synthesis and assembly. In Current Topics in Medical Mycology; McGinnis, M.R., Borgers, M., Eds.; Springer: Berlin/Heidelberg, Germany, 1989; Volume 3, pp. 169–217. [Google Scholar]
- Chen, J.; Seviour, R. Medicinal importance of fungal beta-(1->3), (1->6)-glucans. Mycol. Res. 2007, 111, 635–652. [Google Scholar] [CrossRef] [PubMed]
- Du, B.; Lin, C.Y.; Bian, Z.X.; Xu, B.J. An insight into anti-inflammatory effects of fungal beta-glucan. Trends Food Sci. Technol. 2015, 41, 49–59. [Google Scholar] [CrossRef]
- Wasser, S.P. Medicinal mushroom science: History, current status, future trends, and unsolved problems. Int. J. Med. Mushrooms 2010, 12, 1–16. [Google Scholar] [CrossRef]
- De Silva, D.D.; Rapior, S.; Fons, F.; Bahkali, A.H.; Hyde, K.D. Medicinal mushrooms in supportive cancer therapies: An approach to anti-cancer effects and putative mechanisms of action. Fungal Divers 2012, 55, 1–35. [Google Scholar] [CrossRef]
- Otakar, R.; Mlcek, J.; Jurikova, T. Beta-glucans in higher fungi and their health effects. Nutr. Rev. 2009, 67, 624–631. [Google Scholar]
- Wang, J.C.; Hu, S.H.; Liang, Z.C.; Yeh, C.J. Optimization for the production of water-soluble polysaccharide from Pleurotus citrinopileatus in submerged culture and its antitumor effect. Appl. Microbiol. Biotechnol. 2005, 67, 759–766. [Google Scholar] [CrossRef] [PubMed]
- Kimura, T. Natural products and biological activity of the pharmacologically active cauliflower mushroom Sparassis crispa. BioMed Res. Int. 2013. [Google Scholar] [CrossRef] [PubMed]
- Ruiz-Herrera, J. Biosynthesis of β-glucans in fungi. Antonie van Leeuwenhoek 1991, 60, 73–81. [Google Scholar] [CrossRef]
- Stone, B.A.; Clarke, A.E. Chemistry and Biology of (1->3)-β-Glucans; La Trobe University Press: Melbourne, Australia, 1992. [Google Scholar]
- Wessels, J.G.H. Cell wall synthesis in apical growth. Int. Rev. Cytol. 1986, 104, 37–97. [Google Scholar]
- Bowman, S.M.; Free, S.J. The structure and synthesis of the fungal cell wall. BioEssays 2006, 28, 799–808. [Google Scholar] [CrossRef] [PubMed]
- Shahinian, S.; Bussey, H. β-1,6-Glucan synthesis in Saccharomycetes cerevisae. Mol. Microbiol. 2000, 35, 477–489. [Google Scholar] [CrossRef] [PubMed]
- Ruel, K.; Joseleau, J.P. Involvement of an extracellular glucan sheath during degradation of Populus wood by Phanerochaete chrysosporium. Appl. Environ. Microbiol. 1991, 57, 374–384. [Google Scholar] [PubMed]
- Ruiz-Herrera, J. Fungal Cell Wall: Structure, Synthesis, and Assembly, 2nd ed.; CRC Press: Boca Raton, FL, USA, 2012; pp. 73–95. [Google Scholar]
- Reshetnikov, S.V.; Wasser, S.P.; Nevo, E.; Duckman, I.; Tsukor, K. Medicinal value of the genus Tremella Pers. (Heterobasidiomycetes), (Review). Int. J. Med. Mushrooms 2000, 2, 169–193. [Google Scholar] [CrossRef]
- Vink, E.; Rodriquez-Suarez, R.J.; Gérand-Vincent, M.; Ribas, J.C.; de Nobel, H.; van den Ende, H.; Durán, A.; Klis, F.M.; Bussey, H. An in vitro assay for (1->6)-beta-d-glucan synthesis in Saccharomycetes cerevisae. Yeast 2004, 21, 1121–1131. [Google Scholar] [CrossRef] [PubMed]
- Campbell, J.A.; Davies, G.J.; Bulone, V.; Henrissat, B. A classification of nucleotide-diphospho-sugar glycosyltransferases based on amino acid sequence similarities. Biochem. J. 1997, 326, 929–939. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Douglas, C.M. Fungal β(1,3)-d-glucan synthesis. Med. Mycol. 2001, 39, 55–66. [Google Scholar] [CrossRef] [PubMed]
- Latgé, J.P. The cell wall: A carbohydrate armour for the fungal cell. Mol. Microbiol. 2007, 66, 279–290. [Google Scholar] [CrossRef] [PubMed]
- Ruiz-Herrera, J.; Elorza, M.V.; Alvarez, P.E.; Sentandreu, R. Bioynthesis of the fungal cell wall. In Pathogenic Fungi: Structural Biology and Taxonomy; San-Blas, G., Calderone, R., Eds.; Caister Academic Press: Norfolk, UK, 2004; pp. 41–99. [Google Scholar]
- Perlin, D.S. Current perspectives on echinocandin class drugs. Future Microbiol. 2011, 6, 441–457. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Free, S.J. Fungal cell wall organization and biosynthesis. In Advances in Genetics, 1st ed.; Friedmann, T., Dunlap, J.C., Goodwin, S.F., Eds.; Elsevier: New York, NY, USA, 2013; pp. 33–82. [Google Scholar]
- Fukushima, Y.; Sakagami, Y.; Marumo, S. β-Glucan biosynthesis inhibitors isolated from fungi as hyphal malformation inducer. Bioorg. Med. Lett. 1993, 3, 1219–1222. [Google Scholar] [CrossRef]
- Douglas, C.M.; Foor, F.; Marriman, J.A.; Morin, N.; Nielsen, J.B.; Dahl, A.M.; Mazur, P.; Baginsky, W.; Li, W.; el-Sherbeini, M.; et al. The Saccharomyces cerevisiae FKS1 (ETG1) gene encodes an integral membrane protein which is a subunit of 1,3-β-d-glucan synthase. Proc. Natl. Acad. Sci. USA 1994, 91, 12907–12911. [Google Scholar] [CrossRef] [PubMed]
- Lesage, G.; Bussey, H. Cell wall assembly in Saccharomyces cerevisiae. Microbiol. Mol. Biol. Rev. 2006, 70, 317–343. [Google Scholar] [CrossRef] [PubMed]
- Thompson, J.R.; Douglas, C.M.; Li, W.; Jue, C.K.; Pramanik, B.; Yuan, X.; Rude, T.H.; Toffaletti, D.L.; Perfect, J.R.; Kurtz, M. A glucansynthase FKS homolog in Cryptococcus neoformansis is single copy and encodes an essential function. J. Bacteriol. 1999, 181, 444–453. [Google Scholar] [PubMed]
- Frost, D.J.; Brandt, K.; Capobianco, J.; Goldman, R. Characterization of (1,3)-β-glucan synthase in Candida albicans: Microsomal assay from the yeast or mycelia morphological forms and a permeabilized whole-cell assay. Microbiology 1994, 140, 2239–2246. [Google Scholar] [CrossRef] [PubMed]
- Shematek, E.M.; Braatz, J.A.; Cabib, E. Biosynthesis of the yeast cell wall. 1. Preparation and properties of β-(1-3)glucan synthetase. J. Biol. Chem. 1980, 255, 888–894. [Google Scholar] [PubMed]
- Beauvais, A.; Bruneau, J.M.; Mol, P.C.; Buitrago, M.J.; Legrand, R.; Latge, J.P. Glucan synthase complex of Aspergillus fumigatus. J. Bacteriol. 2001, 183, 2273–2279. [Google Scholar] [CrossRef] [PubMed]
- Teparić, R.; Mrsa, V. Proteins involved in building, maintaining and remodeling of yeast cell walls. Curr. Genet. 2013, 59, 171–185. [Google Scholar] [CrossRef] [PubMed]
- Bickle, M.; Delley, P.A.; Schmidt, A.; Hall, M.N. Cell wall integrity modulates RHO1 activity via exchange factor ROM2. EMBO J. 1998, 17, 2235–2245. [Google Scholar] [CrossRef] [PubMed]
- Ujita, M.; Katsuno, Y.; Szuki, K.; Sugiyama, K.; Takeda, E.; Hara, A.; Yokoyama, E. Molecular cloning and sequence analysis of the β-1,3-glucan synthase catalytic subunit gene from a medicinal fungus, Cordyceps militaris. Mycoscience 2006, 47, 98–105. [Google Scholar] [CrossRef]
- Zevenhuizen, L.P.T.M.; Bartnicki-Garcia, S. Chemical structure of the insoluble hyphal wall glucan of Phytophthora cinnamomi. Biochemistry 1968, 8, 1496–1502. [Google Scholar] [CrossRef]
- Kopecka, M.; Kreger, D.R. Assembly of microfibrils in vivo and in vitro from (1→3)-β-d-glucan synthesised by proptoplasts of Saccharomyces cerevisiae. Arch. Microbiol. 1986, 143, 387–395. [Google Scholar] [CrossRef] [PubMed]
- Larriba, G.; Morales, M.; Ruiz-Herrera, J. Biosynthesis of β-Glucans Microfibriles by cell-free extracts from Saccharomycetes cerevisiae. J. Gen. Microbiol. 1981, 124, 375–383. [Google Scholar]
- Reiskind, J.B.; Mullins, J.T. Molecular architecture of the hyphal wall of Achlya ambisexualis Raper. I. Chemical analyses. Can. J. Microbiol. 1981, 27, 1092–1099. [Google Scholar] [CrossRef]
- Surarit, R.; Gopal, P.K.; Shepherd, M.G. Evidence for a glycosidic linkage between chitin and glucan in the cell wall of Candida albicans. J. Gen. Microbiol. 1988, 134, 1723–1730. [Google Scholar] [CrossRef] [PubMed]
- Sanchez-Hernandez, E.; Garcia-Mendoza, C.; Novaes-Ledieu, M. Characterization of the hyphal walls of the basidiomycete Armillaria mellea. Exp. Mycol. 1990, 14, 178–183. [Google Scholar] [CrossRef]
- Hrmova, M.; Taft, C.; Selitrennikoff, C.P. β-1,3-d-glucan synthase of Neurospora crasssa: Partial purification and characterization of solubilized enzyme activity. Exp. Mycol. 1989, 13, 129–139. [Google Scholar] [CrossRef]
- Mishra, N.C.; Tatum, E.L. Effect of L-sorbose on polysaccharide synthetases of Neurospora crassa. Proc. Natl. Acad. Sci. USA 1972, 69, 313–317. [Google Scholar] [CrossRef] [PubMed]
- Szaniszlo, P.J.; Kang, M.S.; Cabib, E. Stimulation of β(1-3)glucan synthetase of various fungi by nucleoside triphosphates: Generalized regulatory mechanism for cell wall biosynthesis. J. Bacteriol. 1985, 161, 1188–1194. [Google Scholar] [PubMed]
- Monschau, N.; Stahmann, K.-P.; Pielken, P.; Sahm, H. In vitro synthesis of β-(1-3)-glucan with a membrane fraction of Botrytis cinerea. Mycol. Res. 1997, 101, 97–101. [Google Scholar] [CrossRef]
- Kominato, M.; Kamimiya, S.; Tanaka, H. Preparation and properties of β-glucan synthase of Pyricularia oryzae P2. Agric. Biol. Chem. 1987, 51, 755–761. [Google Scholar] [CrossRef]
- San Blas, G. Biosynthesis of glucans by subcellular fractions of Paracoccidioides brasiliensis. Exp. Mycol. 1979, 3, 249–258. [Google Scholar] [CrossRef]
- Kelly, R.; Register, E.; Hsu, M.-J.; Kurtz, M.B.; Nielsen, J. Isolation of a gene involved in 1,3-beta-glucan synthesis in Aspergillus nidulans and purification of the corresponding protein. J. Bacteriol. 1996, 178, 4381–4391. [Google Scholar] [CrossRef] [PubMed]
- Beauvais, A.; Drake, R.; Ng, K.; Diaquin, M.; Latgé, J.P. Characterization of the β 1,3-glucan synthase of Aspergillus fumigatus. J. Gen. Microbiol. 1993, 139, 3071–3078. [Google Scholar] [CrossRef] [PubMed]
- Kurtz, M.B.; Heath, I.B.; Marrinan, J.; Dreikorn, S.; Onishi, J.; Douglas, C. Morphological effects of lipopeptides against Aspergillus fumigatus correlate with activities against 1,3-beta-d-glucan synthase. Antim. Agents Chemother. 1994, 38, 1480–1489. [Google Scholar] [CrossRef]
- Ribas, J.C.; Diaz, M.; Duran, A.; Perez, P. Isolation and characterization of Schizosaccharomyces pombe mutants defective in cell wall (1,3)beta-d-glucan. J. Bacteriol. 1991, 173, 3456–3462. [Google Scholar] [CrossRef] [PubMed]
- Orlean, P.A. (1,3)-β-d-Glucan synthase from budding and filamentous cultures of the dimorphic fungus Candida albicans. Eur. J. Biochem. 1982, 127, 397–403. [Google Scholar] [CrossRef] [PubMed]
- Tang, J.; Parr, T.R. W-1 Solubilization and kinetics of inhibition by cilofungin of Candida albicans (1,3)-β-d-glucan synthase. Antim. Agents Chemother. 1991, 35, 99–103. [Google Scholar] [CrossRef]
- Cabib, E.; Kang, M.S. Fungal 1,3-β-glucan synthase. Meth. Enzymol. 1987, 138, 637–642. [Google Scholar] [PubMed]
- Kottutz, E.; Rapp, P. 1,3-β-glucan synthase in cell free extracts from mycelium and protoplasts of Sclerotium glucanicum. J. Gen. Microbiol. 1990, 136, 1517–1523. [Google Scholar] [CrossRef] [PubMed]
- Antelo, L.; Cosio, E.G.; Hertkorn, N.; Ebel, J. Partial purification of a GTP-insensitive (1->3)-β-glucan synthase from Phytophthora sojae. FEBS Lett. 1998, 433, 191–195. [Google Scholar] [CrossRef]
- Wang, M.C.; Bartnicki-Garcia, S. Synthesis of β-1,3-glucan microfibrils by a cell-free extract from Phytophthora cinnamomi. Arch. Biochem. Biophys. 1976, 175, 351–354. [Google Scholar] [CrossRef]
- Billon-Grand, G.; Marais, M.F.; Joseleau, J.P.; Girard, V.; Gay, L.; Févre, M. A novel (1→3)-β–glucan synthase from the Oomycete Saprolegnia monoica. Microbiology 1997, 143, 3175–3183. [Google Scholar] [CrossRef]
- Fevre, M.; Dumas, C. β-Glucan synthetase from Saprolegnia monoica. J. Gen. Microbiol. 1977, 103, 297–306. [Google Scholar] [CrossRef]
- Cerenius, L.; Söderhäll, K. Isolation and properties of β-glucan synthetase from the aquatic fungus, Aphanomyces astaci. Physiol. Plant 1984, 60, 247–252. [Google Scholar] [CrossRef]
- Robledo-Briones, M.; Ruiz-Herrera, J. Transcriptional regulation of the genes encoding chitin and β-1,3-glucan synthases from Ustilago maydis. Curr. Microbiol. 2012, 65, 85–90. [Google Scholar] [CrossRef] [PubMed]
- Douglas, C.M.; D’Ippolito, J.A.; Shei, G.J.; Meinz, M.; Onishi, J.; Marrinan, J.A.; Li, W.; Abruzzo, G.K.; Flattery, A.; Bartizal, K.; et al. Identification of the FKS1 gene of Candida albicans as the essential target of 1,3-beta-d-glucan synthase inhibitors. Antim. Agents Chemother. 1997, 41, 2471–2479. [Google Scholar]
- Yang, Y.H.; Kang, H.-W.; Ro, H.-S. Cloning and Molecular Characterization of β-1,3-Glucan Synthase from Sparassis crispa. Mycobiology 2014, 42, 167–173. [Google Scholar] [CrossRef] [PubMed]
- Smits, G.J.; Van der Ende, H.; Klis, F.M. Differential regulation of cell wall biogenesis during growth and development in yeast. Microbiology 2001, 147, 781–794. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zerva, A.; Papaspyridi, L.-M.; Christakopoulos, P.; Topakas, E. Valorization of Olive Mill Wastewater for the Production of β-glucans from Selected Basidiomycetes. Waste Biomass Valoriz. 2017, 8, 1721–1731. [Google Scholar] [CrossRef]
- Tartar, A.; Shapiro, A.M.; Scharf, D.W.; Boucias, D.G. Differential expression of chitin synthase (CHS) and glucan synthase (FKS) genes correlates with the formation of a modified, thinner cell wall in in vivo-produced Beauveria bassiana cells. Mycopathologia 2005, 160, 303–314. [Google Scholar] [CrossRef] [PubMed]
- Ha, Y.; Covert, S.F.; Momany, M. FsFKS1, the 1,3-β-Glucan Synthase from the Caspofungin-Resistant Fungus Fusarium solani. Eukaryot. Cell 2006, 5, 1036–1042. [Google Scholar] [CrossRef] [PubMed]
- Lombart, V.; Golaconda Ramulu, H.; Drula, E.; Coutinho, P.M.; Henrissat, B. The carbohydrate-active enzymes database (CAZy) in 2013. Nucleic Acids Res. 2013, 42, D490-5. [Google Scholar] [CrossRef] [PubMed]
- Ruiz-Herrera, J. Fungal Cell Wall: Structure, Synthesis and Assembly; CRC Press: Boca Raton, FL, USA, 1992. [Google Scholar]
- Beauvais, A.; Latgé, J.P. Chitin and β(1-3) glucan synthases in the protoplastic entomophthorales. Arch. Microbiol. 1989, 152, 229–236. [Google Scholar] [CrossRef]
- Quigley, D.R.; Selitrennikoff, C.P. β-Linked disaccharides stimulate, but do not act as a primer for, β (1,3) glucan synthase activity of Neurospora crassa. Curr. Microbiol. 1987, 15, 181–184. [Google Scholar] [CrossRef]
- Wang, M.C.; Bartinicki-Garcia, A. Biosynthesis of beta-1,3- and beta-1,6-linked glucan by Phytophthora cinnamomi cell walls. Biochem. Biophys. Res. Commun. 1966, 24, 832–837. [Google Scholar] [CrossRef]
- Wang, M.C.; Bartinicki-Garcia, A. Synthesis of noncellulosic cell-wall β-glucan by cell-free extracts from zoospores and cysts of Phytophthora palmivora. Exp. Mycol. 1982, 6, 125–135. [Google Scholar] [CrossRef]
- Lopéz-Romero, E.; Ruiz-Herrera, J. Biosynthesis of β-glucans by cell free extracts from Saccharomyces cerevisiae. Biochim. Biophys. Acta 1977, 500, 372–384. [Google Scholar] [CrossRef]
- Lopéz-Romero, E.; Ruiz-Herrera, J. Properties of β-glucan synthetase from Saccharomyces cerevisiae. Antonie van Leeuwenhoek 1978, 44, 329–339. [Google Scholar] [CrossRef] [PubMed]
- Shematek, E.M.; Cabib, E. Biosynthesis of the yeast cell wall. II. Regulation of β-(1-3) glucan synthetase by ATP and GTP. J. Biol. Chem. 1980, 255, 895–902. [Google Scholar] [PubMed]
- Sentandreu, R.; Elorza, M.V.; Villanueva, J.R. Synthesis of yeast wall glucan. J. Gen. Microbiol. 1975, 90, 13–20. [Google Scholar] [CrossRef] [PubMed]
- Balint, S.; Farkas, B.; Bauer, S. Biosynthesis of β-glucans catalyzed by a particulate enzyme preparation from yeast. FEBS Lett. 1976, 64, 44–47. [Google Scholar] [CrossRef]
- Finkelmamn, A.J.; Vardanis, A. Synthesis of β-glucan by cell-free extracts of Aureobasidium pullulans. Can. J. Microbiol. 1987, 33, 123–127. [Google Scholar] [CrossRef]
- Paulson, J.C.; Colley, K.J. Glycosyltransferases. Structure, localization, and control of cell type-specific glycosylation. J. Biol. Chem. 1989, 264, 17615–17618. [Google Scholar] [PubMed]
- Fevre, M. Nucleotide effects on glucan-synthesis activities of particulate enzymes from Saprolegnia. Planta 1983, 159, 130–135. [Google Scholar] [CrossRef] [PubMed]
- Kang, M.S.; Cabib, E. Regulation of fungal cell wall growth: A guanine nucleotide-binding, proteinaceous component required for activity of (1-3)-β-d-glucan synthase. Proc. Natl. Acad. Sci. USA 1986, 83, 5808–5812. [Google Scholar] [CrossRef] [PubMed]
- Notario, V.; Kawai, H.; Cabib, E. Interaction between yeast beta-(1 goes to 3) glucan synthetase and activating phosphorylated compounds. A kinetic study. J. Biol. Chem. 1982, 257, 1902–1905. [Google Scholar] [PubMed]
- Orlean, P.A.B.; Ward, S.M. Sodium fluoride stimulates (1,3)-β-d-glucan synthase from Candida albicans. FEMS Microbiol. Lett. 1983, 18, 31–35. [Google Scholar] [CrossRef]
- Sawitowska-Schroder, E.T.; Kerridge, D.; Perry, H. Echinocandin inhibition of 1,3-β-d-glucan sythase from Candida albicans. FEBS Lett. 1984, 173, 134–138. [Google Scholar] [CrossRef]
- Quigley, D.R.; Selitrennikoff, C.P. β(1-3)Glucan synthase activity of Neurospora crassa: Stabilization and partial characterization. Exp. Mycol. 1984, 8, 202–214. [Google Scholar] [CrossRef]
- Quigley, D.R.; Selitrennikoff, C.P. β(1-3)Glucan synthase activity of Neurospora crassa: Kinetic analysis of negative effectors. Exp. Mycol. 1984, 8, 320–333. [Google Scholar] [CrossRef]
- Frost, D.J.; Drake, R.R.; Wasserman, B.P. (1,3)-β-Glucan synthase from S. cerevisae: In vitro activation by β-lactoglobulin or Brij-35, and photoaffinity labeling of enriched microsomal fractions with 5-azido-UDP-Glc and 8-azido-GTP. Curr. Microbiol. 1992, 24, 295–300. [Google Scholar] [CrossRef]
- Leal, F.; Ruiz-Herrera, J.; Villanueva, J.R.; Larriba, J. An examination of factors affecting the instability of Saccharomyces cerevisiae glucan synthetase in cell free extracts. Arch. Microbiol. 1984, 137, 209–214. [Google Scholar] [CrossRef] [PubMed]
- Varona, R.; Perez, P.; Duran, A. Effect of papulacandin B on β-glucan synthesis in Schizosaccharomyces pombe. FEMS Microbiol. Lett. 1983, 20, 243–247. [Google Scholar]
- Maligie, M.A.; Selitrennikoff, C.P. Cryptococcus neoformans resistance to echinocandins: (1,3)-β-Glucan synthase activity is sensitive to echinocandins. Antim. Agents Chemother. 2005, 49, 2851–2856. [Google Scholar] [CrossRef] [PubMed]
- Taft, C.S.; Stark, T.; Selitrennikoff, C.P. Cilofungin (LY121019) Inhibits Candida albicans (1-3)-β-d-Glucan Synthase Activity. Antim. Agents Chemother. 1988, 32, 1901–1903. [Google Scholar] [CrossRef]
- Okada, H.; Ohnuki, S.; Roncero, C.; Konopka, J.B.; Ohya, Y. Distinctroles of cell wall biogenesis in yeast morphogenesis as revealed by multivariate analysis of high-dimensional morphometric data. Mol. Biol. Cell 2014, 25, 222–233. [Google Scholar] [CrossRef] [PubMed]
- Denning, D.W. Echinocandin antifungal drugs. Lancet 2003, 362, 1142–1151. [Google Scholar] [CrossRef]
- Pfaller, M.A.; Messer, S.A.; Motyl, M.R.; Jones, R.N.; Castanheira, M. In vitro activity of a new oral glucan synthase inhibitor (MK-3118) tested against Aspergillus spp. by CLSI and EUCAST broth microdilution methods. Antim. Agents Chemother. 2013, 57, 1065–1068. [Google Scholar] [CrossRef] [PubMed]
- Lepak, A.J.; Marchillo, K.; Andes, D.R. Pharmacodynamic target evaluation of a novel oral glucan synthase inhibitor, SCY-078 (MK-3118), using an in vivo murine invasive candidiasis model. Antim. Agents Chemother. 2015, 59, 1265–1272. [Google Scholar] [CrossRef] [PubMed]
- Jiménez-Ortigosa, C.; Paderu, P.; Motyl, M.R.; Perlin, D.S. Enfumafungin derivative MK-3118 shows increased in vitro potency against clinical echinocandin-resistant Candida species and Aspergillus species isolates. Antim. Agents Chemother. 2014, 58, 1248–1251. [Google Scholar] [CrossRef] [PubMed]
- Pfaller, M.A.; Messer, S.A.; Motyl, M.R.; Jones, R.N.; Castanheira, M. Activity of MK-3118, a new oral glucan synthase inhibitor, tested against Candida spp. by two international methods (CLSI and EUCAST). Antim. Agents Chemother. 2013, 68, 858–863. [Google Scholar] [CrossRef] [PubMed]
- Lamoth, F.; Alexander, B.D. Antifungal activities of SCY-078 (MK-3118) and standard antifungal agents against clinical non-Aspergillus mold isolates. Antim. Agents Chemother. 2015, 59, 4308–4311. [Google Scholar] [CrossRef] [PubMed]
- Pianalto, K.M.; Alspaugh, J.A. Review: New Horizons in Antifungal Therapy. J. Fungi 2016, 2, 26. [Google Scholar] [CrossRef] [PubMed]
- Brown, G.D.; Gordon, S. A new receptor for β-glucans. Nature 2001, 413, 36. [Google Scholar] [CrossRef] [PubMed]
- Camilli, G.; Tabouret, G.; Quintin, J. The Complexity of Fungal β-Glucan in Health and Disease: Effects on the Mononuclear Phagocyte System. Front. Immunol. 2018, 9, 673. [Google Scholar] [CrossRef] [PubMed]
- Goodridge, H.S.; Wolf, A.J.; Underhill, D.M. Beta-glucan recognition by the innate immune system. Immunol. Rev. 2009, 230, 38–50. [Google Scholar] [CrossRef] [PubMed]
- Adams, E.L.; Rice, P.J.; Graves, B.; Ensley, H.E.; Yu, H.; Brown, G.D.; Gordon, S.; Monteiro, M.A.; Papp-Szabo, E.; Lowman, D.W.; et al. Differential high-affinity interaction of dectin-1 with natural or synthetic glucans is dependent upon primary structure and is influenced by polymer chain length and side-chain branching. J. Pharmacol. Exp. Ther. 2008, 325, 115–123. [Google Scholar] [CrossRef] [PubMed]
- Xie, J.; Guo, L.; Ruan, Y.; Zhu, H.; Wang, L.; Zhou, L.; Yun, X.; Gu, J. Laminarin-mediated targeting to Dectin-1 enhances antigen-specific immune responses. Biochem. Biophys. Res. Commun. 2010, 391, 958–962. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Wang, J.; Wang, W.; Liu, C.; Sun, S.; Gu, J.; Wang, X.; Boraschi, D.; Huang, Y.; Qu, D. Immunomodulatory activity of a novel, synthetic beta-glucan (beta-glu6) in murine macrophages and human peripheral blood mononuclear cells. PLoS ONE 2013, 8, e80399. [Google Scholar] [CrossRef] [PubMed]
- Donadei, A.; Gallorini, S.; Berti, F.; O’Hagan, D.T.; Adamo, R.; Baudner, B.C. Rational Design of Adjuvant for Skin Delivery: Conjugation of Synthetic beta-Glucan Dectin-1 Agonist to Protein Antigen. Mol. Pharm. 2015, 12, 1662–1672. [Google Scholar] [CrossRef] [PubMed]
- Pelosi, L.; Imai, T.; Chanzy, H.; Heux, L.; Buhler, E.; Bulone, V. Structural and morphological Diversity of (1->3)-β-d-Glucans Synthesized in vitro by Enzymes from Saprolengia monoïca. Comparison with a Corresponding in vitro Product from Blackbberry (Rubus fruticosus). Biochemistry 2003, 42, 6264–6274. [Google Scholar] [CrossRef] [PubMed]
- Lai-Kee-Him, J.; Pelosi, L.; Chanzy, H.; Putaux, J.-L.; Bulone, V. Biosynthesis of (1->3)-β-glucans (callose) by detergent extracts of a microsomal fraction from Arabidopsis thaliana. Eur. J. Biochem. 2001, 268, 4628–4638. [Google Scholar]
- Terasaka, K.; Mizutani, Y.; Nagatsu, A.; Mizukami, H. In situ UDP-glucose regeneration unravels diverse functions of plant secondary product glycosyltransferases. FEBS Lett. 2012, 586, 4344–4350. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, F.C.; Hinkelmann, J.; Hermenau, A.; Schwab, W. Enhanced production of beta-glucosides by in-situ UDP-glucose regeneration. J. Biotechnol. 2016, 224, 35–44. [Google Scholar] [CrossRef] [PubMed]
- Schmölzer, K.; Lemmerer, M.; Gutmann, A.; Nidetzky, B. Integrated process design for biocatalytic synthesis by a Leloir Glycosyltransferase: UDP-glucose production with sucrose synthase. Biotechnol. Bioeng. 2017, 114, 924–928. [Google Scholar] [CrossRef] [PubMed]
- Sührer, I.; Langemann, T.; Lubitz, W.; Weuster-Botz, D.; Castiglione, K. A novel one-step expression and immobilization method for the production of biocatalytic preparations. Microb. Cell Fact. 2015, 14, 180. [Google Scholar] [CrossRef] [PubMed]
- Kusters, I.; Mukherjee, N.; de Jong, M.R.; Tans, S.; Koçer, A.; Driessen, A.J.M. Taming Membranes: Functional Immobilization of Biological Membranes in Hydrogels. PLoS ONE 2011, 6, e20435. [Google Scholar] [CrossRef] [PubMed]
- Malet, C.; Planas, A. From β-glucanase to β-glucansynthase: Glycosyl transfer to α-glycosyl fluorides catalyzed by a mutant endoglucanase lacking its catalytic nucleophile. FEBS Lett. 1998, 440, 208–212. [Google Scholar] [CrossRef]
- Danbe, P.M.; Withers, S.G. Advances in Enzymatic Glycoside Synthesis. ACS Chem. Biol. 2016, 11, 1784–1794. [Google Scholar] [CrossRef] [PubMed]
- Mackenzie, L.F.; Wang, Q.; Warren, R.A.J.; Withers, S.G. Glycosynthases: Mutant glycosidases for oligosaccharide synthesis. J. Am. Chem. Soc. 1998, 120, 5583–5584. [Google Scholar] [CrossRef]
- Moracci, M.; Trincone, A.; Rossi, M. Glycosynthases: New enzymes for oligosaccharide synthesis. J. Mol. Catal. B Enzym. 2001, 11, 155–163. [Google Scholar] [CrossRef]
- Planas, A.; Faijes, M. Glycosidases and glycosynthases in enzymatic synthesis of oligosaccharides: An overview. Afinidad 2002, 59, 295–313. [Google Scholar]
- Williams, S.J.; Withers, S.G. Glycosynthases: Mutant glycosidases for glycoside synthesis. Aust. J. Chem. 2002, 55, 3–12. [Google Scholar] [CrossRef]
- Faijes, M.; Pérez, X.; Pérez, O.; Planas, A. Glycosynthase activity of Bacillus licheniformis 1,3-1,4-β-glucanase mutants: Specificity and mechanism. Biochemistry 2003, 42, 13304–13318. [Google Scholar] [CrossRef] [PubMed]
- Faijes, M.; Imai, T.; Bulone, V.; Planas, A. In vitro synthesis of a Crystalline (1->3, 1->4)-β-d-Glucan by a Mutaded (1->3, 1->4)-β-d-Glucanase from Bacillus. Biochem. J. 2004, 380, 635–641. [Google Scholar] [CrossRef] [PubMed]
- Faijes, M.; Planas, A. In vitro synthesis of artificial polysaccharides by glycosidases and glycosynthases. Carbohydr. Res. 2007, 342, 1581–1594. [Google Scholar] [CrossRef] [PubMed]
- Kadokawa, J.; Kobayashi, S. Polymer synthesis by enzymatic catalysis. Curr. Opin. Chem. Biol. 2010, 14, 145–153. [Google Scholar] [CrossRef] [PubMed]
- Fort, S.; Boyer, V.; Greffe, L.; Davies, G.J.; Moroz, O.; Christiansen, L.; Schülein, M.; Cottaz, S.; Driguez, H. Highly Efficient Synthesis of β-(1,4)-oligo- and -Polysaccharides Using a Mutant Cellulase. J. Am. Chem. Soc. 2000, 122, 5429–5437. [Google Scholar] [CrossRef]
- Hrmova, M.; Imai, T.; Rutten, S.J.; Fairweather, J.K.; Pelosi, L.; Bulone, V.; Driguez, H.; Fincher, G.B. Mutated barley (1,3)-beta-d-glucan endohydrolases synthesize crystalline (1,3)-beta-d-glucans. J. Biol. Chem. 2002, 277, 30102–30111. [Google Scholar] [CrossRef] [PubMed]
- Sugimura, M.; Nishimoto, M.; Kitaoka, M. Characterization of Glycosynthase Mutants Derived from Glycoside Hydrolase Family to Xylanases. Biosci. Biotechnol. Biochem. 2006, 70, 1210–1217. [Google Scholar] [CrossRef] [PubMed]
- Pérez, X.; Faijes, M.; Planas, A. Artificial mixed-linked β-glucans produced by glycosythase-catalyzed polymerization: Tuning morphology and degree of polymerization. Biomacromolecules 2011, 12, 494–501. [Google Scholar] [CrossRef] [PubMed]
- Spadiut, O.; Ibatullin, F.M.; Peart, J.; Gullfot, M.; Martinez-Fleites, C.; Ruda, M.; Xu, C.; Sundqvist, G.; Davies, G.J.; Brumer, H. Building custom polysaccharides in vitro with an efficient, broad-specificity xyloglucan glycosynthase and a fucosyltransferase. J. Am. Chem. Soc. 2011, 133, 10892–10900. [Google Scholar] [CrossRef] [PubMed]
- Gulfott, F.; Ibatullin, F.M.; Sundqvist, G.; Davies, G.J.; Brumer, H. Functional characterization of xyloglucan glycosynthase from GH7, GH12 and GH16 scaffolds. Biomacromolecules 2009, 10, 1782–1788. [Google Scholar] [CrossRef] [PubMed]
- Codera, V.; Edgar, K.; Faijes, M.; Planas, A. Functionalized Celluloses with regular substitution Pattern by Glycosynthase—Catalyzed Polymerization. Biomacromolecules 2016, 17, 1272–1279. [Google Scholar] [CrossRef] [PubMed]
- Takakashi, M.; Yoshioka, K.; Imai, T.; Miyoshi, Y.; Yamashita, T.; Furuta, Y.; Yamashita, T.; Futura, Y.; Watanobe, T.; Suqiyama, S.; et al. Degradation and Synthesis of β-glucans by a Magnaporthe oryzae endotransglucosylase, a member of the Glycoside Hydrolase 7 Family. J. Biol. Chem. 2013, 288, 13821–13830. [Google Scholar] [CrossRef] [PubMed]
- Pozzo, T.; Romero-Garcia, J.; Faijes, M.; Planas, A.; Nordberg Karlsson, E. Rational design of a thermostable glycoside hydrolase from family 3 introduces beta-glycosynthase activity. Glycobiology 2017, 27, 165–175. [Google Scholar] [CrossRef] [PubMed]
- Nakai, H.; Kitaoka, M.; Svensson, B.; Ohtsubo, K. Recent development of phosphorylases possessing large potential for oligosaccharide synthesis. Curr. Opin. Chem. Biol. 2013, 17, 301–309. [Google Scholar] [CrossRef] [PubMed]
- O’Neill, E.C.; Field, R.A. Enzymatic synthesis using glycoside phosphorylases. Carbohydr. Res. 2015, 403, 23–37. [Google Scholar] [CrossRef] [PubMed]
- Puchart, V. Glycoside phosphorylases: Structure, catalytic properties and biotechnological potential. Biotechnol. Adv. 2015, 33, 261–276. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, T.; Yamashita, H.; Mukai, K.; Watanabe, H.; Kubota, M.; Chaen, H.; Fukuda, S. Construction and characterization of chimeric enzymes of kojibiose phosphorylase and trehalose phosphorylase from Thermoanaerobacter brockii. Carbohydr. Res. 2006, 341, 2350–2359. [Google Scholar] [CrossRef] [PubMed]
- De Groeve, M.R.; Remmery, L.; Van Hoorebeke, A.; Stout, J.; Desmet, T.; Savvides, S.N.; Soetaert, W. Construction of cellobiose phosphorylase variants with broadened acceptor specificity towards anomerically substituted glucosides. Biotechnol. Bioeng. 2010, 107, 413–420. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, Y.; Kawashima, D.; Hashizume, A.; Hisamatsu, M.; Isono, N. Purification and characterization of 1,3-beta-d-glucan phosphorylase from Ochromonas danica. Biosci. Biotechnol. Biochem. 2013, 77, 1949–1954. [Google Scholar] [CrossRef] [PubMed]
- Kauss, H.; Kriebitzsch, C. Demonstration and partial purification of A β-(1→3)-glucan phosphorylase. Biochem. Bioph. Res. Commun. 1969, 35, 926–930. [Google Scholar] [CrossRef]
- Kitaoka, M.; Sasaki, T.; Taniguchi, H. Purification and properties of laminaribiose phosphorylase (EC 2.4 1.31) from Euglena gracilis. Z. Arch. Biochem. Biophys. 1993, 304, 508–514. [Google Scholar] [CrossRef] [PubMed]
- Kitaoka, M.; Matsuoka, Y.; Mori, K.; Nishimoto, M.; Hayashi, K. Characterization of a bacterial laminaribiose phosphorylase. Biosci. Biotechnol. Biochem. 2012, 76, 343–348. [Google Scholar] [CrossRef] [PubMed]
- Nihira, T.; Saito, Y.; Kitaoka, M.; Nishimoto, M.; Otsubo, K.; Nakai, H. Characterization of a laminaribiose phosphorylase from Acholeplasma laidlawii PG-8A and production of 1,3-beta-d-glucosyl disaccharides. Carbohydr. Res. 2012, 361, 49–54. [Google Scholar] [CrossRef] [PubMed]
- Liénart, Y.; Comtat, J.; Barnoud, F. Wall-bound 1,3-β-d-glucan: Orthophosphate glucosyltransferase activity from acacia cultured cells. Plant Sci. 1988, 58, 165–170. [Google Scholar] [CrossRef]
- Ogawa, Y.; Noda, K.; Kimura, S.; Kitaoka, M.; Wada, M. Facile preparation of highly crystalline lamellae of (1→3)-β-d-glucan using an extract of Euglena gracilis. Int. J. Biol. Macromol. 2014, 64, 415–419. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Chemical structure of fungal glucans. (a) α-(1,4) glucan, (b) β-(1,3) glucan, (c) β-(1,3) glucan with β-(1,6) side chain.
Figure 1.
Chemical structure of fungal glucans. (a) α-(1,4) glucan, (b) β-(1,3) glucan, (c) β-(1,3) glucan with β-(1,6) side chain.

Figure 2.
β-(1,3) glucan chain elongation reaction catalyzed by GLS.

Figure 3.
Schematic representation of β-glucan synthesis in the fungal cell wall by GLS protein complex. FKS 1/2 subunit catalyzes the elongation of the growing β-glucan chains by adding UDP-glucose monomers. The endoplasmic, hydrophilic loop, is one of the domains considered responsible for catalysis. RHO is a GTP-dephosphorylase protein, regulating the action of FKS 1/2 by GDP. ROM is a cell wall associated, GDP-GTP exchange protein, responsible for the regeneration of GTP.
Figure 3.
Schematic representation of β-glucan synthesis in the fungal cell wall by GLS protein complex. FKS 1/2 subunit catalyzes the elongation of the growing β-glucan chains by adding UDP-glucose monomers. The endoplasmic, hydrophilic loop, is one of the domains considered responsible for catalysis. RHO is a GTP-dephosphorylase protein, regulating the action of FKS 1/2 by GDP. ROM is a cell wall associated, GDP-GTP exchange protein, responsible for the regeneration of GTP.

Figure 4.
Diagram of the predicted topology of β-glucan synthase, based on Basidiomycetes sequence predictions. Extra- and intracellular loops are numbered L1–L15. α-helices, corresponding to transmembrane domains (TM1, TM2) are numbered H1–H14. Type 1 and 2 signal sequence motifs are also shown [44,72].
Figure 4.
Diagram of the predicted topology of β-glucan synthase, based on Basidiomycetes sequence predictions. Extra- and intracellular loops are numbered L1–L15. α-helices, corresponding to transmembrane domains (TM1, TM2) are numbered H1–H14. Type 1 and 2 signal sequence motifs are also shown [44,72].


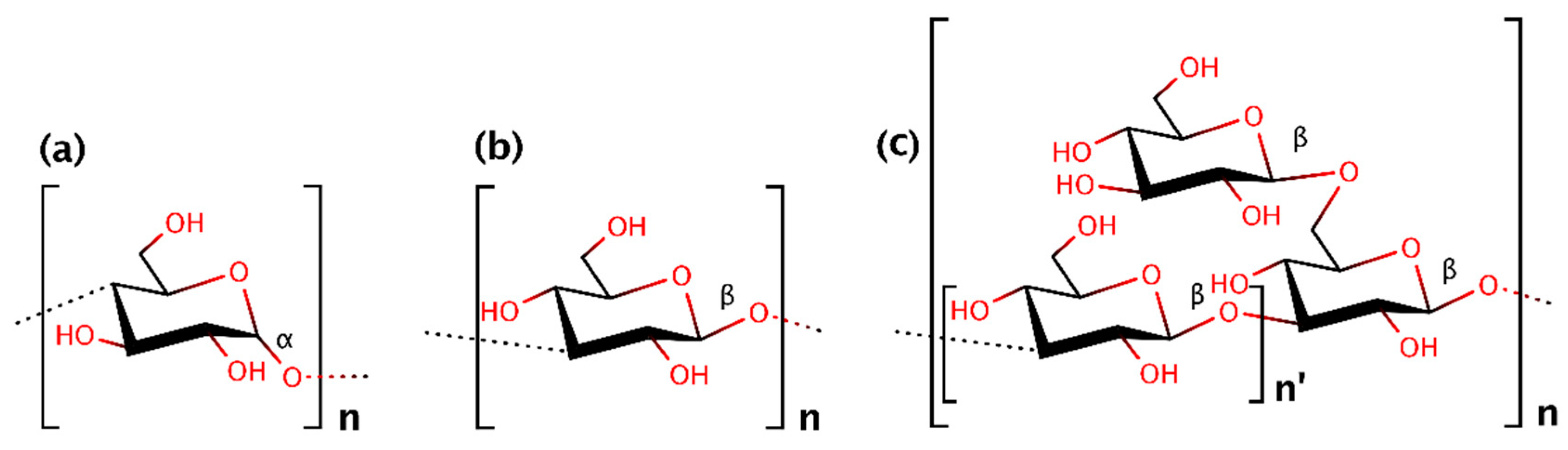{kind=link}
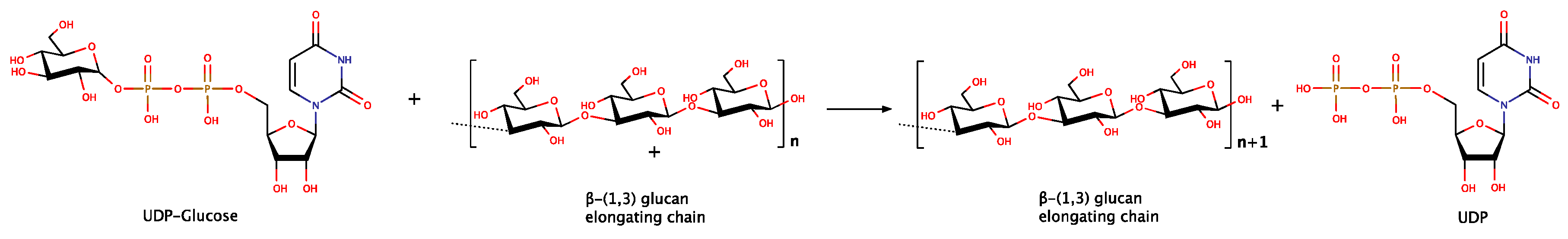{kind=link}
{kind=link}
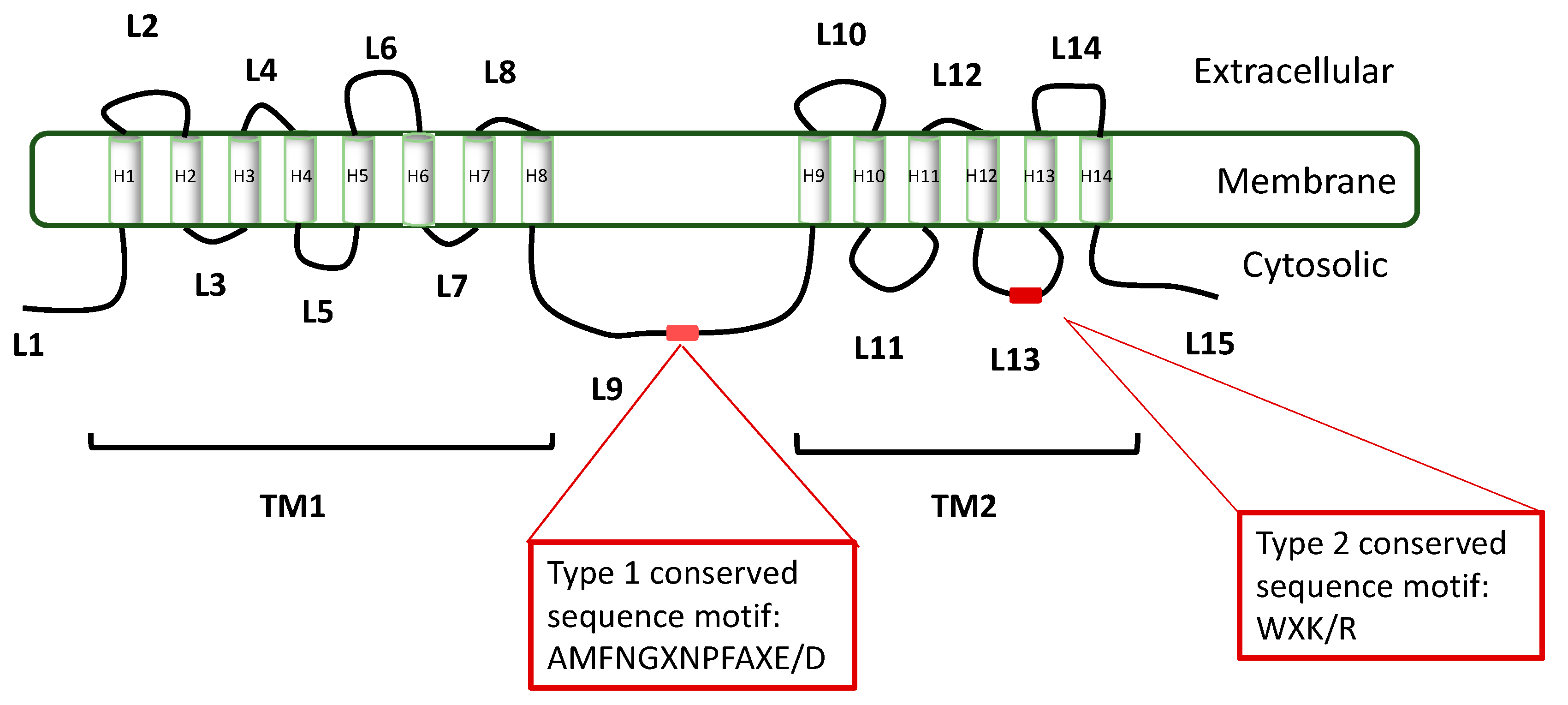{kind=link}
Table 1.
Values of Michaelis constant (KM) for UDPG, optimum pH and temperature (T °C) regarding GLS reported from different sources.
Table 1.
Values of Michaelis constant (KM) for UDPG, optimum pH and temperature (T °C) regarding GLS reported from different sources.
| Microorganism | KM (mM) | pH | T (°C) | Reference |
|---|---|---|---|---|
| Achlya ambisexualis | 7.1 | [53,63] | ||
| Aspergillus fumigatus | 1.92 | [58] | ||
| Aphanomyces astacii | 4.5 | [69] | ||
| Botrytis cinerea | 0.8 | 7.2 | 22 | [54] |
| Candida albicans | 1.5–1.9 | [61] | ||
| Candida albicans | 0.2–0.25 | [62] | ||
| Candida albicans | 1.1 | 7.75 | 30 | [39] |
| Cryptococcus laurentii | 0.86 | [53,63] | ||
| Entomophaga aulicae | 1.7 | [79] | ||
| Hansenula anomala | 0.67 | [53,63] | ||
| Neurospora crassa | 2.9 | [53,63] | ||
| Neurospora crassa | 7.0 | 23 | [52] | |
| Neurospora crassa | 0.71 | 7.4 | [80] | |
| Paracoccidiodes brasiliensis | 1.25–2.0 × 10-4 | [56] | ||
| Phytopthora cinnamoni | 0.09–1.61 | 7.8 | [81,82] | |
| Phytopthora sojae | 10.6 | [65] | ||
| Pyricularia oryzae | 1.2 | 7.0 | 20 | [55] |
| Saccharomyces cerevisae | 0.12 | 6.7 | 24 | [83,84] |
| Saccharomyces cerevisae | 3.8 | [53,63] | ||
| Saccharomyces cerevisae | 1.7 | [85] | ||
| Saprolengia monoica | 0.63–1.25 | 5.8 | [68] | |
| Schizophyllum commune | 0.8 | [53,63] | ||
| Sclerotium glucanicum | 0.54 | 7.5 | 25 | [64] |
| Wangiella dermatitidis | 1.8 | [53,63] |
Table 2.
Compounds that inhibit, activate or do not affect (independent) the enzymatic activity of GLS from different sources.
Table 2.
Compounds that inhibit, activate or do not affect (independent) the enzymatic activity of GLS from different sources.
| Microorganism | Substance | Activity | Reference |
|---|---|---|---|
| Achlya ambisexualis | GTP, NG, SG, ATP, ITP | IND | [53,63] |
| Achlya ambisexualis | BSA | ACT | [53] |
| Aphanomyces astaci | GTP-EDTA, cellobiose | ACT | [69] |
| Aspergillus fumigates | NaF, sucrose, EDTA, GTP | ACT | [58] |
| Botrytis cinerea | GTP, cellobiose, BSA, EGTA | ACT | [54] |
| Botrytis cinerea | UDP | INH | [54] |
| Candida albicans | GTP, ATP, BSA | ACT | [61] |
| Candida albicans | Detergent-W1 | ACT | [62] |
| Candida albicans | NaF | ACT | [93] |
| Candida albicans | TDPG | INH | [61,94] |
| Cryptococcus laurentii | BSA, GTP, NG, SG, ATP, ITP | ACT | [53] |
| Hansenula anomala | GTP | ACT | [91] |
| Hansenula anomala | BSA, GTP, NG, SG, ATP, ITP | ACT | [53] |
| Neurospora crassa | GTP | ACT | [91] |
| Neurospora crassa | cellobiose, BSA | ACT | [95] |
| Neurospora crassa | UDP, UMP, sorbose, gluconolactone | INH | [96] |
| Neurospora crassa | Sorbose | INH | [52] |
| Neurospora crassa | BSA, GTP, NG, SG, ATP, ITP | ACT | [53] |
| Phytopthora palmivora | gentobiose, laminaribiose | ACT | [82] |
| Phytopthora sojae | GTP | IND | [65] |
| Puricularia oryzae | GTP, ATP, UTP, CTP | ACT | [55] |
| Saccharomyces cerevisae | ATP, GTP | ACT | [85] |
| Saccharomyces cerevisae | β-lactoglobullin | ACT | [97] |
| Saccharomyces cerevisae | UDP, gluconolactone | INH | [84] |
| Saccharomyces cerevisae | NaF | ACT | [47,98] |
| Saprolengia monoica | ATP, GTP | INH | [67] |
| Saprolengia monoica | cellobiose | INH | [68] |
| Schizophyllum commune | BSA, GTP, NG, SG, ATP, ITP | ACT | [53] |
| Schizosaccharomyces pombe | GTP | ACT | [99] |
| Sclerotium glucanicum | ATP, GTP | ACT | [64] |
| Wangiella dermatitidis | BSA, GTP, NG, SG, ATP, ITP | ACT | [53] |
Abbreviations: ACT (activation); INH (inhibition); IND (independent) ATP (adenosine triphosphate); GTP (guanosine triphosphate); UDP (uridine diphosphate); UMP (uridine monophosphate); NG [guanosine 5′-(β,γ)-imino triphosphate]; SG [guanosine 5′-(γ-thio) triphosphate]; ITP (inosine triphosphate); BSA (bovine serum albumin); EGTA (egtazic acid); EDTA (ethylenediaminetetraacetic acid).
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Papaspyridi, L.-M.; Zerva, A.; Topakas, E. Biocatalytic Synthesis of Fungal β-Glucans. Catalysts 2018, 8, 274. https://doi.org/10.3390/catal8070274
AMA Style
Papaspyridi L-M, Zerva A, Topakas E. Biocatalytic Synthesis of Fungal β-Glucans. Catalysts. 2018; 8(7):274. https://doi.org/10.3390/catal8070274
Chicago/Turabian StylePapaspyridi, Lefki-Maria, Anastasia Zerva, and Evangelos Topakas. 2018. "Biocatalytic Synthesis of Fungal β-Glucans" Catalysts 8, no. 7: 274. https://doi.org/10.3390/catal8070274
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.