
An Overview of the Top Ten Detergents Used for Membrane Protein Crystallization


Groningen Biomolecular Sciences and Biotechnology Institute, University of Groningen, Nijenborgh 4, 9747 AG Groningen, The Netherlands
*
Author to whom correspondence should be addressed.
Crystals 2017, 7(7), 197; https://doi.org/10.3390/cryst7070197
Submission received: 7 June 2017
/
Revised: 26 June 2017
/
Accepted: 28 June 2017
/
Published: 1 July 2017
(This article belongs to the Special Issue Recent Advances in Protein Crystallography)
Abstract
:To study integral membrane proteins, one has to extract them from the membrane—the step that is typically achieved by the application of detergents. In this mini-review, we summarize the top 10 detergents used for the structural analysis of membrane proteins based on the published results. The aim of this study is to provide the reader with an overview of the main properties of available detergents (critical micelle concentration (CMC) value, micelle size, etc.) and provide an idea of what detergents to may merit further study. Furthermore, we briefly discuss alternative solubilization and stabilization agents, such as polymers.
1. Introduction
Every cell is encircled by the semipermeable membrane (often termed as the lipid bilayer), not only to protect the cell content from the environment, but also to differentiate it externally from the other cells, and internally to form the dedicated organelles within the cell. However, this forced evolution to design a myriad of membrane-embedded and associated proteins, which are essential for the transport of charged and large chemicals in and out of the cell (since they either cannot pass or diffuse too slowly across the membrane) and also for communication between adjacent cells. Not surprisingly, malfunction of these proteins can be extremely detrimental, therefore there is a constantly growing interest to study these proteins in order to decipher the molecular basis of diseases. However they are also attractive from a more fundamental point of view—since many crucial processes occur in the membrane or associated with it, for example photosynthesis [1] or G-protein-coupled receptor (GPCR)-signalling [2] to name just a few. Unfortunately, membrane proteins turned out to be rather difficult to study, since one needs to extract a protein of interest from the membrane, but more importantly extracted protein requires a special environment mimicking the membrane to keep it stable. Historically, the most widely used agents for membrane protein extraction and stabilization are detergents, the amphiphatic molecules, bearing a hydrophilic headgroup (typically polar, sometimes charged) and a hydrophobic (apolar) tail. Due to this nature, detergents are capable of inserting their hydrophobic tails into the lipidic membrane, thus disrupting the latter and eventually (with an increasing concentration of detergent) extracting membrane-embedded proteins (Figure 1). Since the other part of detergent molecule is polar, detergent molecules spontaneously form micelles (pseudo-spherical assemblies) as soon as critical micelle concentration (CMC) is achieved and under condition that the sample is above critical micellar temperature (CMT). Below CMC value, detergent molecules are mostly present as monomers. Due to the micelle formation, a membrane protein becomes a part of the detergent (lipid)-protein complex, sometimes with the complete loss of surrounding lipids. In many cases, the importance of lipids for the function and/or getting crystals of membrane proteins has been highlighted, e.g., for photosynthetic proteins [3,4], transporters and channels [5,6,7,8], and GPCRs [9]. Furthermore, even the partial removal of lipids sometimes can be destabilizing, possibly due to the decrease in lateral pressure [10] or increase in hydrophobic mismatch [11]. Also, it is important not to forget that the formed detergent-protein complex is dynamic, thus there is a constant exchange of the detergents molecules between the complex and the free detergent pool. This implies a strict (and rather costly) condition to maintain the concentration of detergent above its CMC value during all the steps of protein study. Regardless of all these and other limitations, detergents still to remain the first choice agents in studies on membrane proteins. Despite all of the chemical variety of detergents, all are typically characterized by several parameters. In addition to the aforementioned CMC value, the aggregation number N, and the micelle size (in terms of Mw) are commonly reported. The latter value is extremely helpful during the concentration step of purified protein-detergent complex—the right cut-off value of a concentrator will prevent the overconcentration of a detergent (in free micelles), a potentially harmful event for crystallization [12]. The higher concentration of a detergent, the higher the chance of phase separation, which might ultimately lead to protein denaturation. Parameters such as temperature, salt, and precipitant also influence the detergent phase behavior [13,14]. However, the phase separation can be also used in a beneficial way, for example as a simple and cheap purification technique (reviewed in [15,16]) or to design crystallization experiments [17].
To estimate the amount of bound detergent to the protein of interest, several techniques can be used. The application of matrix-assisted laser desorption/ionization time of flight mass spectrometry (MALDI-TOF MS) is extremely powerful, since it allows both quantification and identification of bound detergents/lipids [18,19] and even analysis of (membrane) protein–ligand interactions [20]. In cases where the identity of the bound substance is known, one can also use multi-angle laser light scattering coupled with size-exclusion chromatography (SEC-MALLS) to quantify the amounts of bound material [21,22].
The most general classification of detergents divides them into three major classes: non-ionic, ionic and zwitter-ionic. Non-ionic detergents are the most commonly used for cases when native state of protein is critical. These detergents are considered mild—a qualitative indicator of the impact of detergent onto protein stability. Non-ionic detergents typically disrupt protein-lipid (and lipid-lipid) but not protein-protein interactions, thus maintaining (or better to phrase—not interfering with) the native state of a protein. On the contrary, ionic detergents are typically harsh, since they disrupt additionally protein–protein interactions, thus often bringing proteins into the denatured state. Ionic detergents have head groups charged either positively (cationic) or negatively (anionic). Due to this fact, the properties of these detergents (most importantly their CMC values) are affected by the ionic strength, thus special care should be taken during experiments. Zwitter-ionic detergents, as their name implies, have both charges present, rendering an electro-neutral molecule. These detergents are typically milder than the ionic ones, but somewhat harsher than non-ionic detergents. There are numerous comprehensive reviews on different aspects of detergents available [23,24,25], to which we would like to point out to the readers’ attention for more detailed information.
Apart from detergents, there are several other agents capable of extraction and stabilization of membrane proteins. In the past few years, the styrene maleic acid (SMA) polymer has gained much attention (see refs. [26,27] for recent reviews). There are a few benefits of SMA—it is a very cheap material, it extracts membrane proteins with the accompanying lipids (both intrinsic and annular), and also keeps them stable as a SMA-lipid-protein (SMALP) complex (Figure 1). Since SMA does not form micelles, there is no need to maintain the pool of freely available SMA molecules in buffers as in case of detergents (see above). Clearly, there are also some limitations of using SMAs, preventing them from the ubiquitous application, and those are a rather defined size of SMALPs of ~10 nm [28], thus large proteins will probably not fit in; pH sensitivity (at pH 6.5 and below SMA starts precipitating [29]), and limited compatibility with the crystallogenesis. However, very recently, the group of Oliver Ernst reported the first crystal structure of bacteriorhodopsin extracted and purified in SMA and crystallized in the lipidic cubic phase [30]. Additionally, SMAs might find their niche in the cryo-Electron Microscopy (Cryo-EM) field [31,32] and with the future generations of SMAs, (e.g., with the various lengths and ratios of polymers [33]) they might become a very useful tool for studies on membrane proteins.
Another polymer widely used in membrane protein research are amphipols (see ref. [34] for the detailed review). Amphipols are amphipathic (bearing both hydrophobic and hydrophilic units) and highly soluble, thus capable to wind around a membrane protein, shielding its hydrophobic patches away and providing the necessary solubility of the formed protein–amphipol complex (Figure 1). The key difference from the aforementioned SMA polymers is that amphipols cannot be used for membrane-protein extraction and can be only applied to the detergent-solubilized sample. However the amphipol–detergent exchange is simple and quick, and many proteins are more stable in the presence of amphipols (see ref. [35] and references therein). The most commonly used of the amphipols is A8-35, comprised of ~35 acrylate units with Mw of ~4.3 kDa. On its own, amphipols can form micelle-like spherical particles (with ~3.15 nm radius) at the concentrations exceeding their ‘CMC’ value (0.002% (w/v) for A8-35 [36]). Despite this, there is an exchange between the protein–amphipol complex and amphipol particles (there are almost no free amphipol molecules in solution); amphipols stay bound to membrane proteins strongly even in the amphipol-free buffers. However amphipols can be very easily washed away by detergents, thus upon an amphipol exchange, care should be taken to not expose a sample to detergent-containing buffers. Currently, amphipols gained significant popularity in the Cryo-EM field [37,38,39] but also in solution NMR studies [40,41]. For the X-ray crystallography field, it has been shown that amphipols are compatible with lipidic cubic phases [42], thus currently amphipols can only be used for in meso crystallization [43], similarly to SMALPs.
2. Results
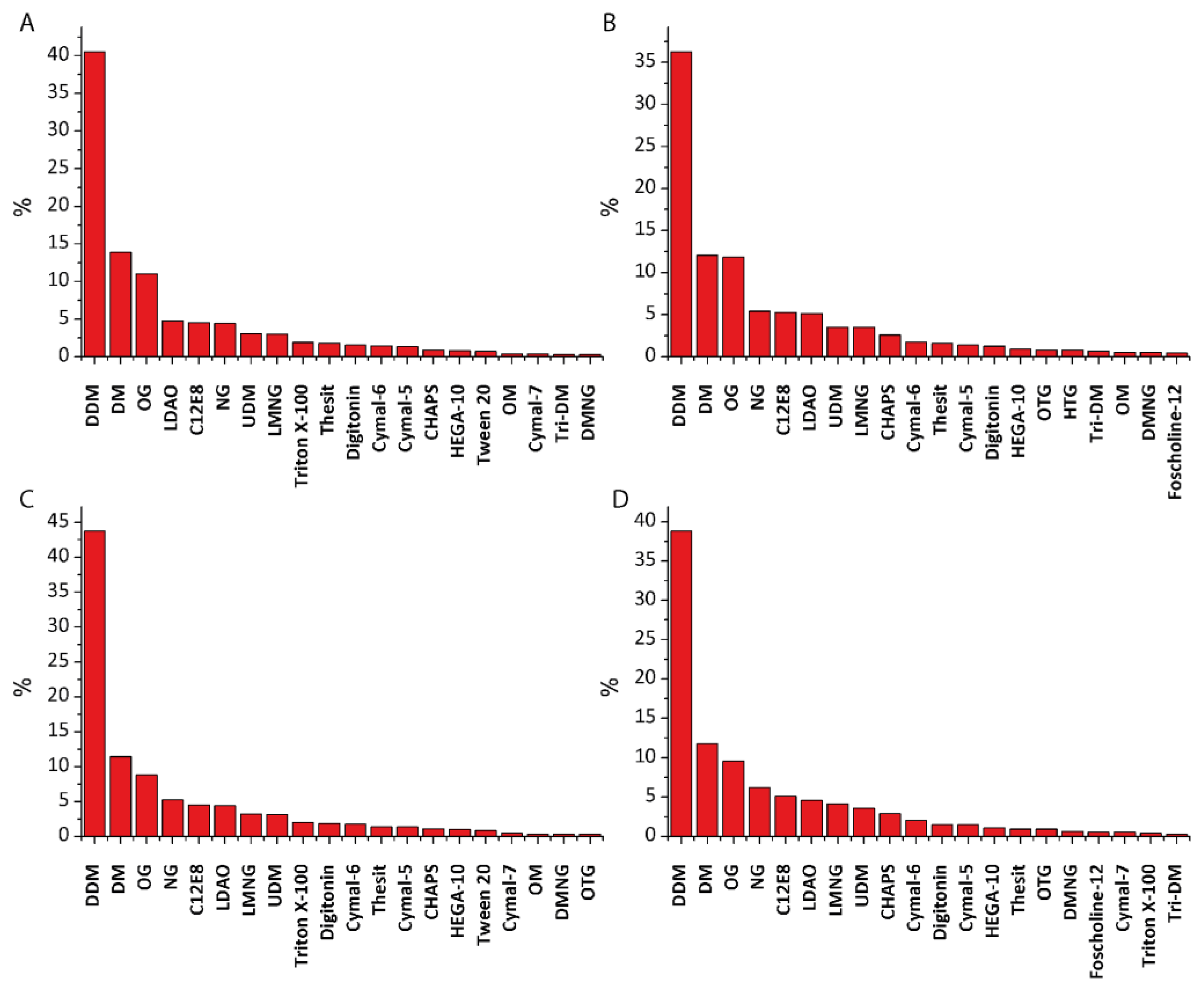
We have analyzed all the entries in the “Membrane Proteins of Known Structure” database (up to 31 December 2016) and manually extracted information regarding what kind of detergent was used for protein purification and crystallization. It is obvious that the increased interest in membrane proteins during the past 30 years caused not only the cumulative growth in use of detergents but also forced the development of novel detergents. The first membrane protein structure of the photosynthetic reaction center solubilized using zwitter-ionic detergent Lauryldimethylamine-N-oxide (LDAO) was solved in 1985 [47] and since then LDAO application has grown steadily. Another zwitter-ionic detergent 3-[(3-cholamidopropyl)dimethylammonio]-1-propanesulfonate (CHAPS) (and its hydroxylated form 3-[(3-cholamidopropyl)dimethylammonio]-2-hydroxy-1-propanesulfonate (CHAPSO)) started gaining popularity in the early 2000s with an increasing application in Cryo-EM. But in fact more than a half of proteins during these years were investigated using alkyl maltosides and glycosides. The most commonly used detergent in both categories (purification and crystallization) is n-Dodecyl-β-D-Maltopyranoside (DDM) (40.6% and 36.3% respectively), followed by n-Decyl-β-d-Maltopyranoside (DM) (13.9% and 12.1%), and n-Octyl-β-d-Glucopyranoside / n-Nonyl-β-d-Glucopyranoside (OG/NG) (11.0% and 11.9%/4.5% and 5.4% respectively) (Figure 2). The other detergents that scored more than 1% are LDAO (4.8% and 5.2%), C12E8 (4.6% and 5.3%), n-Undecyl-β-d-maltopyranoside (UDM) (3.1% and 3.5%), Lauryl maltose neopentyl glycol (LMNG) (3.0% and 3.5%), Triton X-100 (1.9% and 0.3%), Thesit (1.8% and 1.6%), Digitonin (1.6% and 1.3%), Cymal-5 (1.4% and 1.4%) and Cymal-6 (1.5% and 1.8%), and CHAPS (1.0% and 2.6%). In only about 50% cases the same detergent was used for both purification and crystallization.
Interestingly, if we take into account only the past decade, the results do not differ much (Figure 2)—DDM, DM, and OG dominate, both for purification and crystallization.
Remarkably, in the field of Nuclear magnetic resonance (NMR) spectroscopy, the distribution is totally different. DDM is used very rarely, since it forms large micelles that tumble slowly, causing the broadening of protein signal in NMR spectra, rendering this detergent not very suitable for NMR spectroscopy. However anionic detergents, such as sodium dodecyl sulfate (SDS) and different variants of phosphocholine (e.g., n-Dodecylphosphocholine (FC-12), 1,2-Dimyristoyl-sn-Glycero-3-Phosphocholine (DMPC), 1,2-Dioleoyl-sn-Glycero-3-Phosphocholine (DOPC)), considered very harsh for macromolecular crystallography studies, are widely used in NMR. The overview of detergents (and alternative membrane mimetics) explicitly used in NMR spectroscopy can be found in [48].
Below we provide the short descriptions of the detergents that are currently used the most in structural biology of membrane proteins.
2.1. n-Dodecyl-β-d-Maltopyranoside
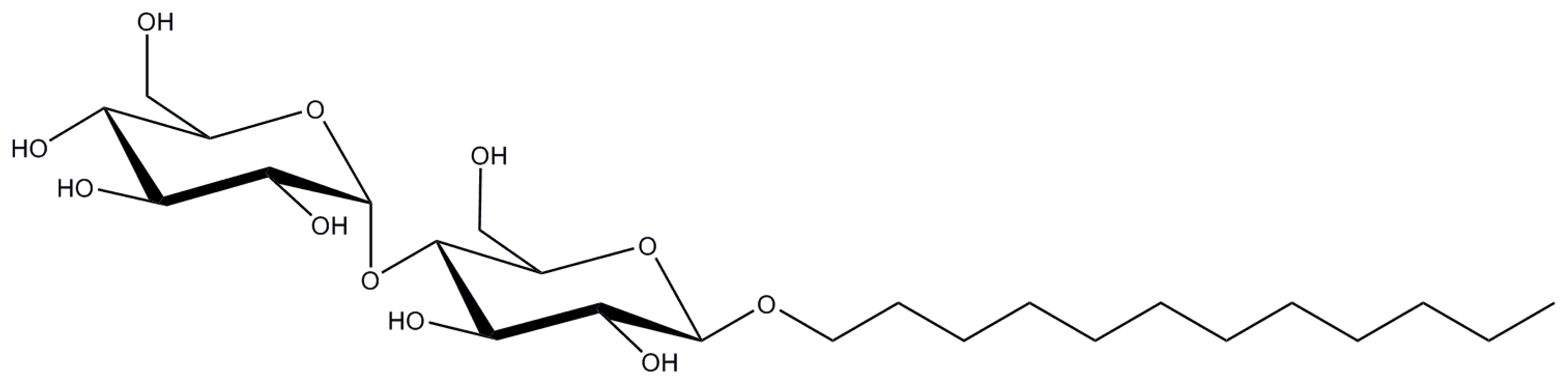
Also known as Layrul maltoside or most commonly, DDM, n-Dodecyl-β-d-maltopyranoside is currently the most used detergent (see above). It belongs to the class of alkyl maltosides, thus it has a hydrophilic maltose headgroup and a hydrophobic alkyl chain (Figure 3).
Its popularity can be explained by several factors: it was developed as a cheaper alternative to OG detergent basically at the dawn of membrane protein (structural) studies, in 1980 [49]. Furthermore it has a low CMC value (~0.0087%/0.17 mM [50]) that allows reducing the amount of detergent needed (though rendering it more difficult to remove). Thus both economically and historically, as well as partly due to ‘herd behavior’, it acquired the leading position for structural studies on membrane proteins. Nevertheless, it is also quite an efficient detergent for protein extraction and since it is non-ionic, it is mild enough to maintain the stable native state of many proteins. The main drawback of DDM is that the micelle it forms is relatively large (Mw ~65–70 kDa [51,52]) and it forms a substantial and rather mobile belt around proteins that can be detrimental during crystallogenesis. DDM seems to be a truly universal detergent (however with the limited application in NMR spectroscopy), since it has proved to be successful for virtually all the classes of α-helical proteins.
2.2. n-Decyl-β-d-Maltopyranoside
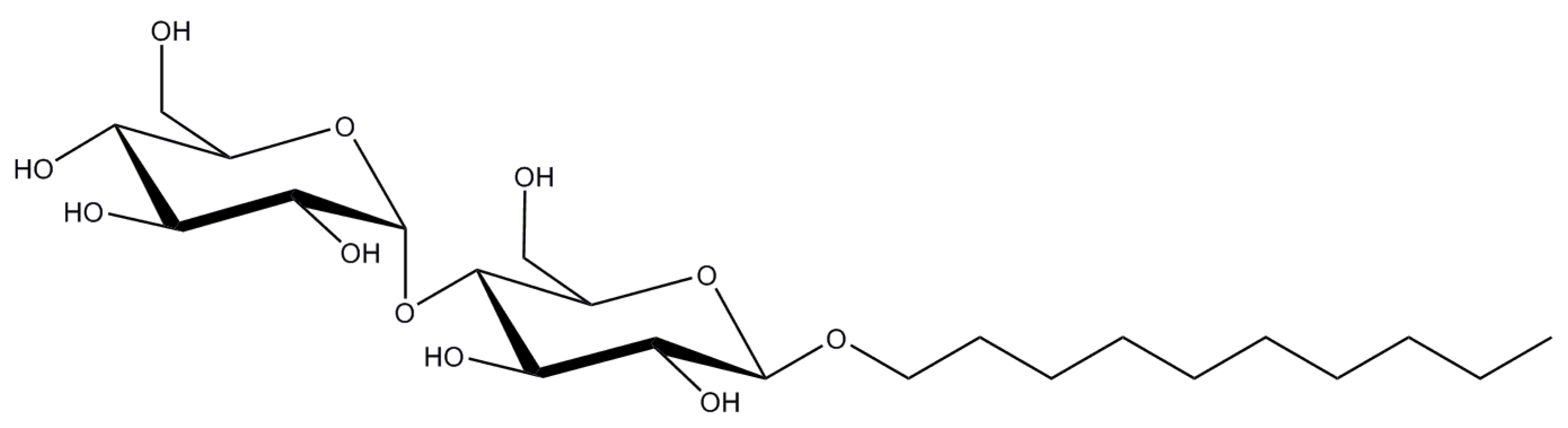
Also known as Decyl maltoside or DM, n-Decyl-β-d-maltopyranoside is a shortened version of DDM (two carbons less in the hydrophobic tail) (Figure 4). Due to this shortening, it forms considerably smaller micelles (Mw ~ 40 kDa [51]) but at the expense of 10 times higher CMC (0.087%/1.8 mM [50]). This higher CMC value might partially explain why this detergent is less popular than DDM (at least in economical reasons), but in general detergents with the shorter alkyl chains seem to be less stabilizing.
2.3. n-Octyl-β-d-Glucopyranoside
Also known as Octyl glycoside or OG, n-Octyl-β-d-glucopyranoside is the one of the two most used glucoside-based detergents. It has one sugar moiety less in its head group compared to maltosides (Figure 5), but similarly to them, OG is a non-ionic, albeit less milder detergent. It has a very high CMC of 0.53%/20 mM [53] with the compact micelle (Mw ~ 25 kDa [53]).
2.4. n-Nonyl β-d-Glucopyranoside
Also known as Nonyl glucoside or NG, n-Nonyl-β-d-glucopyranoside is an extended (one extra alkyl unit) version of OG (Figure 6). Such an extension rendered it to have smaller CMC (0.20%/6.5 mM) but much larger micelle (Mw ~90 kDa, this work).
Application of NG correlates well with the proteins studied with OG, with an addition of the intramembrane proteases family, where NG dominates among other detergents, and the Energy coupling factor (ECF) family of ATP-binding cassette (ABC) transporters, where NG was used in combination with (D)DM.
2.5. Lauryldimethylamine-N-Oxide
Lauryldimethylamine-N-oxide (LDAO), also known as dodecyldimethylamine oxide (DDAO), is a zwitter-ionic detergent, bearing two opposite charges in its head group and a long hydrophobic tail (Figure 7). Though considered rather a harsh detergent due to the charged head group, nevertheless it has landed on the fifth place of the most used detergents for membrane protein research. This might be explained by the evidence that proteins stable in LDAO often produce well diffracting crystals [8,54,55]. The latter is probably dictated by the compact micelle size (Mw ~ 21.5 kDa [52]) that might promote better packing within crystals. Interestingly LDAO was shown to form rather elongated and not the spherical micelles [56,57]. It has CMC value of 0.023% / 1–2 mM.
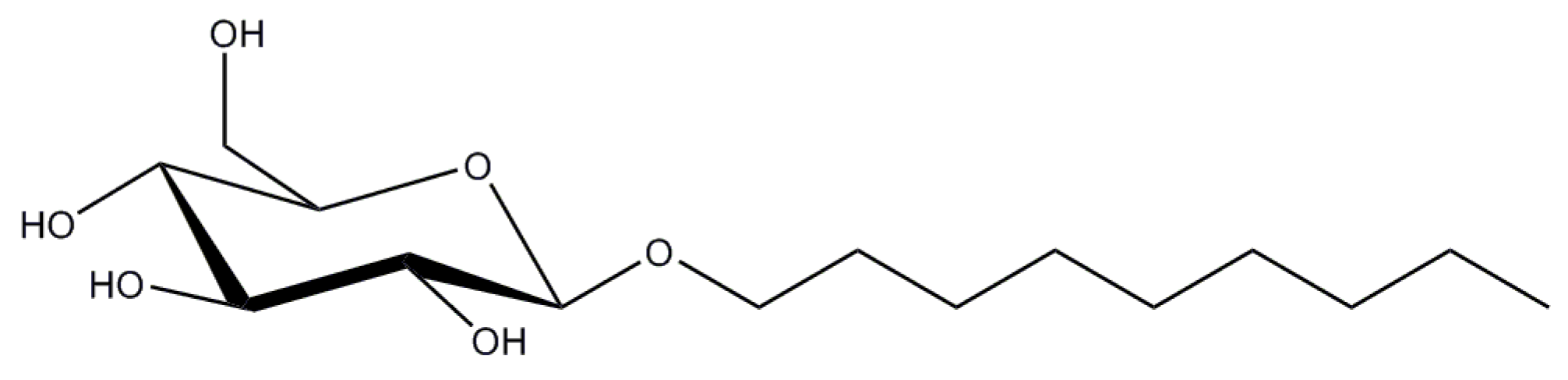
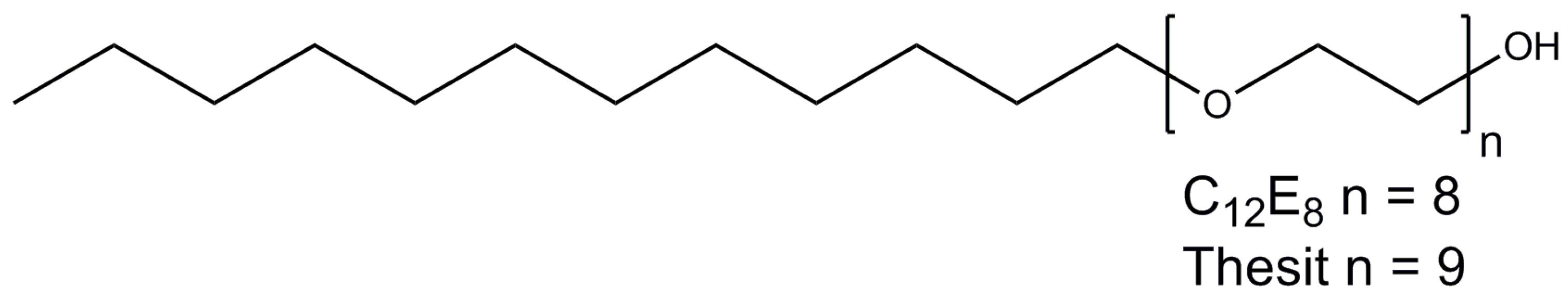
2.6. Polyoxyethylene 8 (9) dodecyl Ether
Polyoxyethylene 8 (9) dodecyl ether, or C12E8 (C12E9), are non-ionic detergents that belong to the family of alkyl polyoxyethelenes with the general formula CxEy, where y denotes oxyethelene units in the head group, and x describes the length of the alkyl tail (Figure 8). C12E9 is also named Thesit or polydocanol.
C12E8 has a very low CMC value of 0.005% (0.09 mM) [58] and the micelle size of ~66 kDa [59]. C12E9 has even lower CMC value of 0.003% (0.05 mM) [60], but considerably larger micelle (Mw of ~ 83 kDa [52].
C12E8 has been successfully used for purification and crystallization of several ABC-transporters and is the number one detergent for studies on P-type ATPases, whereas C12E9 has proven to be successful in the studies on the electron-transfer chain complex II.
2.7. n-Undecyl-β-d-Maltopyranoside
On the next position is yet another maltoside—n-Undecyl-β-d-maltopyranoside, also known as UM or UDM. It is an intermediate between DDM and DM in terms of the length of the hydrophobic tail: 11 carbon atoms for UDM (Figure 9) versus 12 and 10 for DDM and DM respectively. Its CMC value is about 0.029%/0.59 mM with the micelle size of about 50 kDa [51].
UDM has been used in the studies of many different membrane protein families, with the highest occurrence in the studies of cytochrome bc1 and cytochrome b6f.
2.8. Lauryl Maltose Neopentyl Glycol
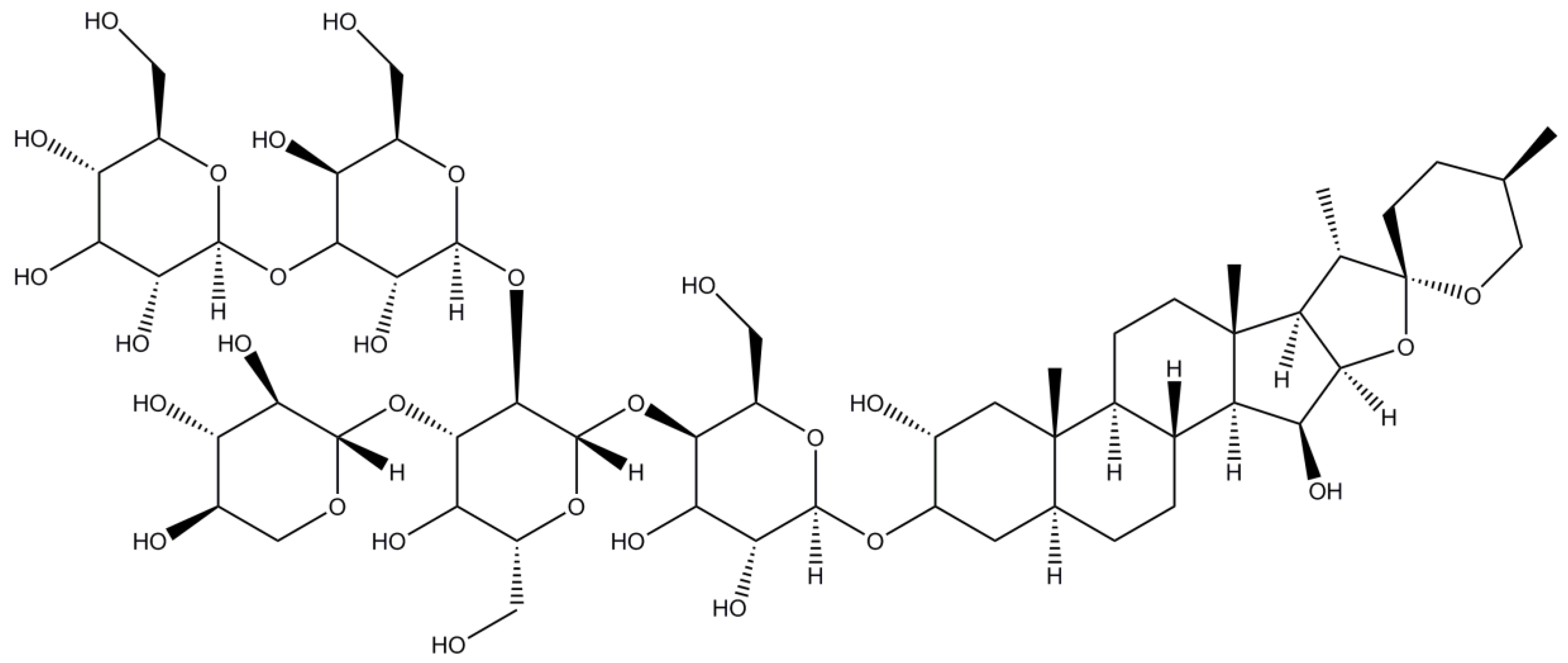
Lauryl maltose neopentyl glycol (LMNG or MNG-3) is a representative of the novel, recently developed maltose neopentyl glycol detergents [50]. The main feature of this class of detergents is the presence of quaternary carbon atom that allows incorporation of two hydrophilic head groups and two hydrophobic tails (Figure 10).
LMNG has a low CMC value (0.001%/0.01 mM [33]), with the size of micelle of Mw ~ 91 kDa (this work), which is in contrast with the recent result of ~ 393 kDa obtained with MALDI-TOF MS [19]. It has been reported to be efficient both at protein extraction and stabilization of several delicate membrane proteins [37,61,62]. So far it has been rather extensively used for studies on GPCRs and was also successful for the family of transient receptor potential (TRP) channels and the N-methyl-D-aspartate (NMDA) receptors among others.
2.9. Triton X-100
Triton X-100, or octyl phenol ethoxylate (Figure 11) is one of the oldest classical non-ionic detergents still in use. It has CMC value of 0.01%/0.2 mM, with the micelle size of 60 to 90 kDa (temperature-dependent) [63,64].
The main disadvantage of Triton X-100 is the presence of the aromatic ring that absorbs strongly in the UV region of the spectrum, thus interfering with the protein quantification. Furthermore, since Triton X-100 is obtained during the polymerization reaction of octylphenol with ethylene oxide, the final product is heterogeneous, containing on average 9.5 units of ethylene oxide. To aggravate that, polyethelene glycol is a typical by-product, and additionally peroxides were reported to be present in the detergent preparations as the result of aging [65]. This explains the significant difference between the number of cases, where Triton X-100 was used for extraction/purification (1.8%) and for crystallization (0.3%), where typically homogeneity and purity play a crucial role in crystallogenesis [66].
2.10. Digitonin
Digitonin is a truly natural (albeit toxic) detergent obtained from the purple foxglove plant Digitalis purpurea. It has a steroid-like saposin structure (Figure 12).
Digitonin has a CMC value of 0.02%–0.03% / 0.25–0.5 mM [67,68] with the micelle size of ~ 70–75 kDa [69]. Since it is not synthesized but extracted from the plant, there might be considerable batch-to-batch variations; therefore the alternative glyco-diosgenin (GDN) has been recently proposed as a suitable non-toxic replacement [70]. Nevertheless, during the past few years digitonin gained a considerable popularity for the structural studies on eukaryotic membrane proteins with Cryo-EM [71,72].
2.11. Cymal-5 (Cymal-6)
These detergents are the special case of maltoside detergents with the cyclohexyl aliphatic tail (Figure 13).
Cymal-5 has a very high CMC value of ~0.12% (~2.5 mM [73]) and the micelle size of ~ 23 kDa [74], whereas slightly longer Cymal-6 has a lowered CMC value of ~0.028% (0.56 mM) and a larger micelle size of ~32 kDa.
Cymal-5 and -6 were used for a broad range of channels and transporters, and in many cases as a second detergent or an additive.
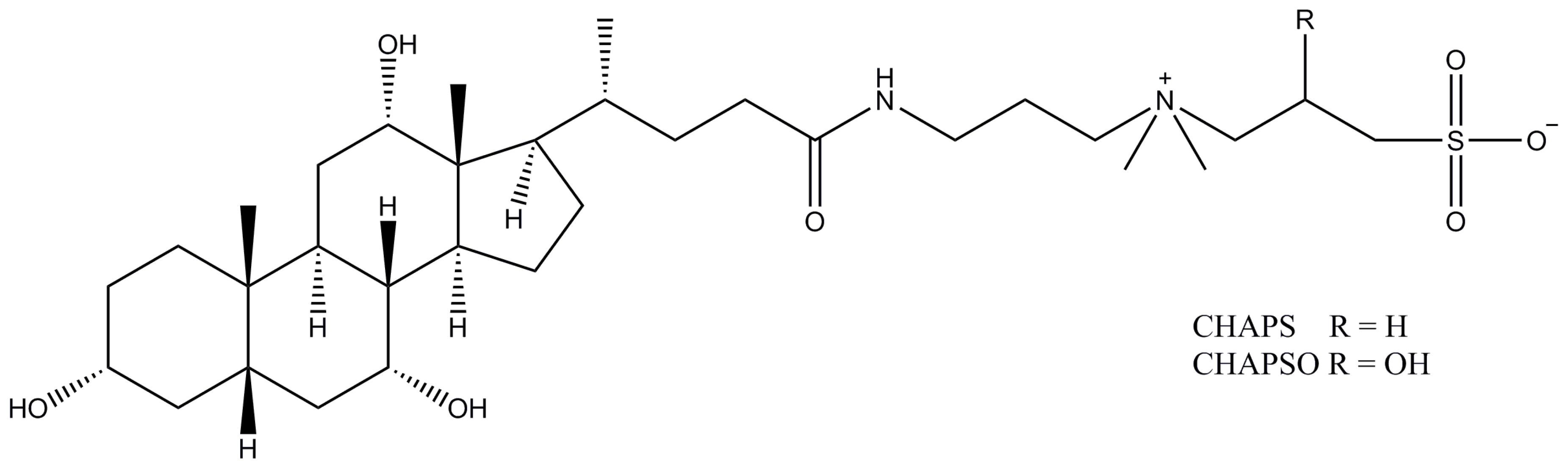
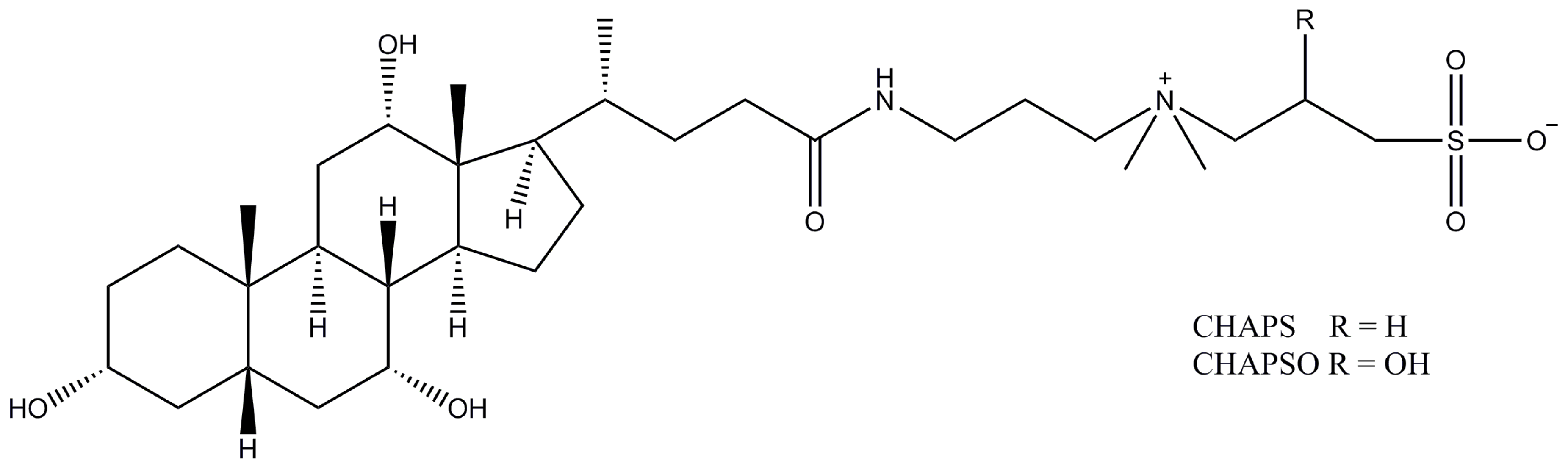
2.12. CHAPS (CHAPSO)
CHAPS (CHAPSO), also known as 3-[(3-cholamidopropyl)dimethylammonio]-1-propanesulfonate and 3-[(3-cholamidopropyl)dimethylammonio]-2-hydroxy-1-propanesulfonate respectively, are other representatives of zwitter-ionic detergents derived from the bile salts and having cholesterol-like structures (Figure 14). Both CHAPS and CHAPSO have a very high CMC value of ~0.5% (8–10 mM) with the very small micelles of just 6 and 7 kDa respectively. Such a combination of high CMC and small micelle size makes it extremely easy to remove CHAPS and CHAPSO by dialysis. Both detergents have been used in the structural studies on a variety of transporters, with the recently increased interest in their application for Cryo-EM.
For convenience, we compiled Table 1 to summarize the main characteristics of the aforementioned detergents.
3. Discussion and Outlook
Looking at the results of the performed analysis, one can confidently say that the sugar-based detergents rule the field of structural studies on membrane proteins. Just two maltoside-based detergents, DDM and DM, are reported as the detergents of choice for more than a half of deposited structures. With an addition of UDM, cyclic maltosides Cymal-5 and Cymal-6, and glycoside-based detergents OG and NG, the total fraction of sugar-based detergents is around 75%. Does it mean that sugar-based detergents are the best? Not necessarily so, but the fact that they are the most common helps them to maintain their popularity over the years. Clearly, any new detergent faces a problem of how to reach the market – to make it commercially available and successful, first it must prove to be useful for a broad range of targeted proteins. Furthermore, many scientists tend to utilize proven techniques and reagents, and not only because of reproducibility of results, but quite often also for economical reasons, since detergents are rather expensive chemicals. In this respect it is interesting and exciting to witness the extension of a current toolbox with cheaper alternatives, such as SMA polymer. Clearly it has numerous limitations, but with further improvements in the design of this polymer, coupled with the recent promising results in application of SMA in crystallization and Cryo-EM studies, it is feasible to say it will gradually gain more and more attention. Amphipols are also becoming more popular, again more in the field of Cryo-EM. They are easy to apply and provide a good stability; in addition, amphipols do not form large micelles like detergents do.
In the case of a detergent solubilized protein, one can face the situation that the most of the protein is obscured by large detergent belt, therefore impeding the analysis of images, whereas amphipols form a relatively small belt, thus not interfering with classification of images. Furthermore, other alternatives exist, such as bicelles for crystallization [76]—commercial kits available (in the form of lipid-detergent mixture, typically DMPC with CHAPSO); fluorinated surfactants [46] and nanodiscs (a patch of the lipid bilayer encircled by membrane scaffolding proteins) [44] for protein stabilization. Fluorinated surfactants have not gained much attention so far, probably due to its nature—it has a hydrophobic tail with the incorporation of several fluorine atoms, rendering it lipophobic and preventing effective interactions with a membrane protein, typically causing aggregation of the latter. The insertion of a hydrogenated tip reduced lypophobicity [77] and improved protein stability, but nevertheless these agents are still far from being widely used. In contrast, reconstitution in nanodiscs has become one of the standard methods, especially for functional studies on membrane proteins. However, it was recently found that they also were useful for structural studies using Cryo-EM [78]. Recently, more tunable and size-controlled covalently circularized nanodiscs with enhanced stability have been engineered [79].
Nevertheless, there is also considerable research being conducted in the design and development of novel detergents. One of the very successful examples are neopentyl glycol maltoside and glucoside detergents [50]. Introduced less than a decade ago, it has confidently landed in top 10 of the most used detergents in our analysis. The further design of this class of detergents is still ongoing; very recently the same group presented so-called tandem-neopentyl glycol maltosides [80]. Another example is the development of calixarenes [45]—this novel class of detergents is built on a rigid calixarene scaffold to which hydrophobic tail and hydrophilic and charged headgroups are attached. The charge present on their headgroups promotes salt-bridge interactions with the patches of basic residues readily available on surfaces of membrane proteins at the cytosol-membrane interface [81]. Furthermore the steroid-based facial amphiphiles (FA) are another type of solubilization/stabilization agents being actively developed. They utilize the cholate scaffold, to which (typically) sugar headgroups are attached [82,83]. Such design proved to be useful to not only to stabilize membrane proteins (via tight interactions between a membrane proteins and FA owning to the large hydrophobic surface of the latter), but to also improve their crystallization (via the formation of more compact protein–detergent complexes, which can pack more tightly in a crystal, owing to the ability of FA to mediate intermolecular contacts between molecules of membrane proteins) [84]. Additionally, the longer tandem version of FA is available, where two steroid moieties are connected via a linker of varying length to match the width of a lipid bilayer [85].
Taking into account a large collection of classical detergents and continuously appearing new ones, the valid question of any researcher dealing with the membrane proteins is which detergent to use. Unfortunately, there is no magic single detergent that would work for all of the proteins, therefore usually some screening is required. In cases where the high throughput is not supreme, the small-scale solubilization and purification tests can be run with about three to five detergents from the aforementioned list (in our lab we generally test DDM, NG, OG, LMNG, and LDAO) to get the initial idea about protein stability. The screening can be optimized via the introduction of GFP-fusion for a rapid assessment of detergent solubilization efficiency [86] and protein stability in a certain detergent [86,87]. In cases where the high throughput is desirable, other protocols such as BMSS (Biotinylated Membranes Solubilization & Separation) [88] or fully-automated detergent screening [89] can be applied for fast-screening of 96 different detergents at once.
From a very naïve point of view, it might appear that alternative solubilization and stabilization agents will gradually seize the Cryo-EM branch of structural biology, whereas detergents will dominate in the X-ray crystallography field. Whether it is correct or not, time will tell. However, it is exciting to witness the current developments both in the design of new detergents and further improvements in the alternative agents. Coupled with the technological improvements in their structural biology, it is feasible to say that the amount of new membrane proteins deposited in the protein data bank will steadily grow in coming years thus producing a larger array of data to analyze preferences of membrane proteins for different detergents.
4. Materials and Methods
We used a public database (Membrane Proteins of Known Structure) maintained by Dr. Stephen White’s laboratory (UC Irvine, USA).
The determination of micelles size formed by NG and LMNG was performed using size-exclusion chromatography coupled with multi-angle laser light scattering (SEC-MALLS) as described in [51].
Acknowledgments
This work has been funded by Netherlands Organisation for Scientific Research NWO Vidi grant 723.014.002 to A.G. We are grateful to Marina Guskova for help during figures preparation.
Author Contributions
A.S. extracted the data and performed SEC-MALLS; A.S. and A.G. analyzed data and wrote the manuscript.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Flügge, U.-I.; Westhoff, P.; Leister, D. Recent advances in understanding photosynthesis. F1000Research 2016, 5, 2890. [Google Scholar] [CrossRef] [PubMed]
- Ellisdon, A.M.; Halls, M.L. Compartmentalization of GPCR signalling controls unique cellular responses. Biochem. Soc. Trans. 2016, 44, 562–567. [Google Scholar] [CrossRef] [PubMed]
- Kern, J.; Guskov, A. Lipids in photosystem II: Multifunctional cofactors. J. Photochem. Photobiol. B Biol. 2011, 104, 19–34. [Google Scholar] [CrossRef] [PubMed]
- Mazor, Y.; Borovikova, A.; Caspy, I.; Nelson, N. Structure of the plant photosystem I supercomplex at 2.6 Å resolution. Nat. Plants 2017, 3, 17014. [Google Scholar] [CrossRef] [PubMed]
- Hunte, C.; Richers, S. Lipids and membrane protein structures. Curr. Opin. Struct. Biol. 2008, 18, 406–411. [Google Scholar] [CrossRef] [PubMed]
- Valiyaveetil, F.I.; Zhou, Y.; MacKinnon, R. Lipids in the structure, folding, and function of the KcsA K+ channel. Biochemistry 2002, 41, 10771–10777. [Google Scholar] [CrossRef] [PubMed]
- Lemieux, M.J.; Song, J.; Kim, M.J.; Huang, Y.; Villa, A.; Auer, M.; Li, X.-D.; Wang, D.-N. Three-dimensional crystallization of the Escherichia coli glycerol-3-phosphate transporter: A member of the major facilitator superfamily. Protein Sci. 2003, 12, 2748–2756. [Google Scholar] [CrossRef] [PubMed]
- Sonoda, Y.; Newstead, S.; Hu, N.-J.; Alguel, Y.; Nji, E.; Beis, K.; Yashiro, S.; Lee, C.; Leung, J.; Cameron, A.D.; et al. Benchmarking Membrane Protein Detergent Stability for Improving Throughput of High-Resolution X-ray Structures. Structure 2011, 19, 17–25. [Google Scholar] [CrossRef] [PubMed]
- Cherezov, V.; Rosenbaum, D.M.; Hanson, M.A.; Rasmussen, S.G.F.; Thian, F.S.; Kobilka, T.S.; Choi, H.-J.; Kuhn, P.; Weis, W.I.; Kobilka, B.K.; et al. High-resolution crystal structure of an engineered human beta2-adrenergic G protein-coupled receptor. Science 2007, 318, 1258–1265. [Google Scholar] [CrossRef] [PubMed]
- Van den Brink-van der Laan, E.; Chupin, V.; Killian, J.A.; de Kruijff, B. Stability of KcsA tetramer depends on membrane lateral pressure. Biochemistry 2004, 43, 4240–4250. [Google Scholar] [CrossRef] [PubMed]
- Parton, D.L.; Klingelhoefer, J.W.; Sansom, M.S.P. Aggregation of model membrane proteins, modulated by hydrophobic mismatch, membrane curvature, and protein class. Biophys. J. 2011, 101, 691–699. [Google Scholar] [CrossRef] [PubMed]
- Newby, Z.E.R.; O’Connell, J.D.; Gruswitz, F.; Hays, F.A.; Harries, W.E.C.; Harwood, I.M.; Ho, J.D.; Lee, J.K.; Savage, D.F.; Miercke, L.J.W.; et al. A general protocol for the crystallization of membrane proteins for X-ray structural investigation. Nat. Protoc. 2009, 4, 619–637. [Google Scholar] [CrossRef] [PubMed]
- Hitscherich, C.; Aseyev, V.; Wiencek, J.; Loll, P.J. Effects of PEG on detergent micelles: Implications for the crystallization of integral membrane proteins. Acta Crystallogr. D Biol. Crystallogr. 2001, 57, 1020–1029. [Google Scholar] [CrossRef] [PubMed]
- Bordier, C. Phase separation of integral membrane proteins in Triton X-114 solution. J. Biol. Chem. 1981, 256, 1604–1607. [Google Scholar] [PubMed]
- Sánchez-Ferrer, A.; Bru, R.; García-Carmona, F. Phase separation of biomolecules in polyoxyethylene glycol nonionic detergents. Crit. Rev. Biochem. Mol. Biol. 1994, 29, 275–313. [Google Scholar] [CrossRef] [PubMed]
- Arnold, T.; Linke, D. Phase separation in the isolation and purification of membrane proteins. BioTechniques 2007, 43, 427–442. [Google Scholar] [CrossRef] [PubMed]
- Koszelak-Rosenblum, M.; Krol, A.; Mozumdar, N.; Wunsch, K.; Ferin, A.; Cook, E.; Veatch, C.K.; Nagel, R.; Luft, J.R.; DeTitta, G.T.; et al. Determination and application of empirically derived detergent phase boundaries to effectively crystallize membrane proteins. Protein Sci. 2009, 18, 1828–1839. [Google Scholar] [CrossRef] [PubMed]
- Laganowsky, A.; Reading, E.; Allison, T.M.; Ulmschneider, M.B.; Degiacomi, M.T.; Baldwin, A.J.; Robinson, C.V. Membrane proteins bind lipids selectively to modulate their structure and function. Nature 2014, 510, 172–175. [Google Scholar] [CrossRef] [PubMed]
- Chaptal, V.; Delolme, F.; Kilburg, A.; Magnard, S.; Montigny, C.; Picard, M.; Prier, C.; Monticelli, L.; Bornert, O.; Agez, M.; et al. Quantification of Detergents Complexed with Membrane Proteins. Sci. Rep. 2017, 7, 41751. [Google Scholar] [CrossRef] [PubMed]
- Allison, T.M.; Reading, E.; Liko, I.; Baldwin, A.J.; Laganowsky, A.; Robinson, C.V. Quantifying the stabilizing effects of protein-ligand interactions in the gas phase. Nat. Commun. 2015, 6, 8551. [Google Scholar] [CrossRef] [PubMed]
- Guskov, A.; Slotboom, D.J. Size exclusion chromatography with multi-angle laser light scattering (SEC-MALLS) to determine protein oligomeric states. In From Molecules to Living Organisms: An Interplay between Biology and Physics; Lecture Notes of the Les Houches School of Physics; Volume 102, July 2014; Oxford University Press: New York, NY, USA, 2016; pp. 169–183. [Google Scholar]
- Folta-Stogniew, E. Oligomeric states of proteins determined by size-exclusion chromatography coupled with light scattering, absorbance, and refractive index detectors. Methods Mol. Biol. 2006, 328, 97–112. [Google Scholar] [PubMed]
- Linke, D. Detergents: An overview. Methods Enzymol. 2009, 463, 603–617. [Google Scholar] [PubMed]
- Smith, S.M. Strategies for the Purification of Membrane Proteins. In Protein Chromatography: Methods and Protocols; Walls, D., Loughran, S., Eds.; Springer: Berlin, Germany, 2017; pp. 389–400. [Google Scholar]
- Lin, S.-H.; Guidotti, G. Purification of membrane proteins. Methods Enzymol. 2009, 463, 619–629. [Google Scholar] [PubMed]
- Lee, S.C.; Pollock, N.L. Membrane proteins: Is the future disc shaped? Biochem. Soc. Trans. 2016, 44, 1011–1018. [Google Scholar] [CrossRef] [PubMed]
- Dörr, J.M.; Scheidelaar, S.; Koorengevel, M.C.; Dominguez, J.J.; Schäfer, M.; van Walree, C.A.; Killian, J.A. The styrene-maleic acid copolymer: A versatile tool in membrane research. Eur. Biophys. J. 2016, 45, 3–21. [Google Scholar] [CrossRef] [PubMed]
- Jamshad, M.; Grimard, V.; Idini, I.; Knowles, T.J.; Dowle, M.R.; Schofield, N.; Sridhar, P.; Lin, Y.; Finka, R.; Wheatley, M.; et al. Structural analysis of a nanoparticle containing a lipid bilayer used for detergent-free extraction of membrane proteins. Nano Res. 2015, 8, 774–789. [Google Scholar] [CrossRef]
- Lee, S.C.; Knowles, T.J.; Postis, V.L.G.; Jamshad, M.; Parslow, R.A.; Lin, Y.-P.; Goldman, A.; Sridhar, P.; Overduin, M.; Muench, S.P.; et al. A method for detergent-free isolation of membrane proteins in their local lipid environment. Nat. Protoc. 2016, 11, 1149–1162. [Google Scholar] [CrossRef] [PubMed]
- Broecker, J.; Eger, B.T.; Ernst, O.P. Crystallogenesis of Membrane Proteins Mediated by Polymer-Bounded Lipid Nanodiscs. Structure 2017, 25, 384–392. [Google Scholar] [CrossRef] [PubMed]
- Dörr, J.M.; Koorengevel, M.C.; Schäfer, M.; Prokofyev, A.V.; Scheidelaar, S.; van der Cruijsen, E.A.W.; Dafforn, T.R.; Baldus, M.; Killian, J.A. Detergent-free isolation, characterization, and functional reconstitution of a tetrameric K+ channel: The power of native nanodiscs. Proc. Natl. Acad. Sci. USA 2014, 111, 18607–18612. [Google Scholar] [CrossRef] [PubMed]
- Postis, V.; Rawson, S.; Mitchell, J.K.; Lee, S.C.; Parslow, R.A.; Dafforn, T.R.; Baldwin, S.A.; Muench, S.P. The use of SMALPs as a novel membrane protein scaffold for structure study by negative stain electron microscopy. Biochim. Biophys. Acta 2015, 1848, 496–501. [Google Scholar] [CrossRef] [PubMed]
- Craig, A.F.; Clark, E.E.; Sahu, I.D.; Zhang, R.; Frantz, N.D.; Al-Abdul-Wahid, M.S.; Dabney-Smith, C.; Konkolewicz, D.; Lorigan, G.A. Tuning the size of styrene-maleic acid copolymer-lipid nanoparticles (SMALPs) using RAFT polymerization for biophysical studies. Biochim. Biophys. Acta 2016, 1858, 2931–2939. [Google Scholar] [CrossRef] [PubMed]
- Zoonens, M.; Popot, J.-L. Amphipols for Each Season. J. Membr. Biol. 2014, 247, 759–796. [Google Scholar] [CrossRef] [PubMed]
- Popot, J.L.; Althoff, T.; Bagnard, D.; Banéres, J.L.; Bazzacco, P.; Billon-Denis, E.; Catoire, L.J.; Champeil, P.; Charvolin, D.; Cocco, M.J.; et al. Amphipols from a to Z. Annu. Rev. Biophys. 2011, 40, 379–408. [Google Scholar] [CrossRef] [PubMed]
- Giusti, F.; Popot, J.-L.; Tribet, C. Well-defined critical association concentration and rapid adsorption at the air/water interface of a short amphiphilic polymer, amphipol A8–35: A study by Förster resonance energy transfer and dynamic surface tension measurements. Langmuir 2012, 28, 10372–10380. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Clarke, O.B.; Kim, J.; Stowe, S.; Kim, Y.-K.; Assur, Z.; Cavalier, M.; Godoy-Ruiz, R.; Desiree, C.; Manzini, C.; et al. Structure of the STRA6 receptor for retinol uptake. Science 2016, 353, aad8266. [Google Scholar] [CrossRef] [PubMed]
- Zubcevic, L.; Herzik, M.A.; Chung, B.C.; Liu, Z.; Lander, G.C.; Lee, S.-Y. Cryo-electron microscopy structure of the TRPV2 ion channel. Nat. Struct. Mol. Biol. 2016, 23, 180–186. [Google Scholar] [CrossRef] [PubMed]
- Bai, X.-C.; Rajendra, E.; Yang, G.; Shi, Y.; Scheres, S.H.W. Sampling the conformational space of the catalytic subunit of human γ-secretase. Elife 2015, 4, e11182. [Google Scholar] [CrossRef] [PubMed]
- Elter, S.; Raschle, T.; Arens, S.; Viegas, A.; Gelev, V.; Etzkorn, M.; Wagner, G. The use of amphipols for NMR structural characterization of 7-TM proteins. J. Membr. Biol. 2014, 247, 957–964. [Google Scholar] [CrossRef] [PubMed]
- Planchard, N.; Point, É.; Dahmane, T.; Giusti, F.; Renault, M.; Le Bon, C.; Durand, G.; Milon, A.; Guittet, É.; Zoonens, M.; et al. The use of amphipols for solution NMR studies of membrane proteins: Advantages and constraints as compared to other solubilizing media. J. Membr. Biol. 2014, 247, 827–842. [Google Scholar] [CrossRef] [PubMed]
- Polovinkin, V.; Gushchin, I.; Sintsov, M.; Round, E.; Balandin, T.; Chervakov, P.; Schevchenko, V.; Utrobin, P.; Popov, A.; Borshchevskiy, V.; et al. High-resolution structure of a membrane protein transferred from amphipol to a lipidic mesophase. J. Membr. Biol. 2014, 247, 997–1004. [Google Scholar] [CrossRef] [PubMed]
- Caffrey, M. A comprehensive review of the lipid cubic phase or in meso method for crystallizing membrane and soluble proteins and complexes. Acta Crystallogr. F Struct. Biol. Commun. 2015, 71, 3–18. [Google Scholar] [CrossRef] [PubMed]
- Denisov, I.G.; Sligar, S.G. Nanodiscs for structural and functional studies of membrane proteins. Nat. Struct. Mol. Biol. 2016, 23, 481–486. [Google Scholar] [CrossRef] [PubMed]
- Matar-Merheb, R.; Rhimi, M.; Leydier, A.; Huché, F.; Galián, C.; Desuzinges-Mandon, E.; Ficheux, D.; Flot, D.; Aghajari, N.; Kahn, R.; et al. Structuring detergents for extracting and stabilizing functional membrane proteins. PLoS ONE 2011, 6, e18036. [Google Scholar] [CrossRef] [PubMed]
- Popot, J.-L. Amphipols, nanodiscs, and fluorinated surfactants: Three nonconventional approaches to studying membrane proteins in aqueous solutions. Annu. Rev. Biochem. 2010, 79, 737–775. [Google Scholar] [CrossRef] [PubMed]
- Deisenhofer, J.; Epp, O.; Miki, K.; Huber, R.; Michel, H. Structure of the protein subunits in the photosynthetic reaction centre of Rhodopseudomonas viridis at 3Å resolution. Nature 1985, 318, 618–624. [Google Scholar] [CrossRef] [PubMed]
- Warschawski, D.E.; Arnold, A.A.; Beaugrand, M.; Gravel, A.; Chartrand, É.; Marcotte, I. Choosing membrane mimetics for NMR structural studies of transmembrane proteins. Biochim. Biophys. Acta 2011, 1808, 1957–1974. [Google Scholar] [CrossRef] [PubMed]
- Rosevear, P.; VanAken, T.; Baxter, J.; Ferguson-Miller, S. Alkyl glycoside detergents: A simpler synthesis and their effects on kinetic and physical properties of cytochrome c oxidase. Biochemistry 1980, 19, 4108–4115. [Google Scholar] [CrossRef] [PubMed]
- Chae, P.S.; Rasmussen, S.G.F.; Rana, R.R.; Gotfryd, K.; Chandra, R.; Goren, M.A.; Kruse, A.C.; Nurva, S.; Loland, C.J.; Pierre, Y.; et al. Maltose–neopentyl glycol (MNG) amphiphiles for solubilization, stabilization and crystallization of membrane proteins. Nat. Methods 2010, 7, 1003–1008. [Google Scholar] [CrossRef] [PubMed]
- Slotboom, D.J.; Duurkens, R.H.; Olieman, K.; Erkens, G.B. Static light scattering to characterize membrane proteins in detergent solution. Methods 2008, 46, 73–82. [Google Scholar] [CrossRef] [PubMed]
- Strop, P.; Brunger, A.T. Refractive index-based determination of detergent concentration and its application to the study of membrane proteins. Protein Sci. 2005, 14, 2207–2211. [Google Scholar] [CrossRef] [PubMed]
- Lorber, B.; Bishop, J.B.; DeLucas, L.J. Purification of octyl beta-D-glucopyranoside and re-estimation of its micellar size. Biochim. Biophys. Acta 1990, 1023, 254–265. [Google Scholar] [CrossRef]
- Sonoda, Y.; Cameron, A.; Newstead, S.; Omote, H.; Moriyama, Y.; Kasahara, M.; Iwata, S.; Drew, D. Tricks of the trade used to accelerate high-resolution structure determination of membrane proteins. FEBS Lett. 2010, 584, 2539–2547. [Google Scholar] [CrossRef] [PubMed]
- Parker, J.L.; Newstead, S. Current trends in α-helical membrane protein crystallization: An update. Protein Sci. 2012, 21, 1358–1365. [Google Scholar] [CrossRef] [PubMed]
- Thiyagarajan, P.; Tiede, D.M. Detergent micelle structure and micelle-micelle interactions determined by small-angle neutron scattering under solution conditions used for membrane protein crystallization. J. Phys. Chem. 1994, 98, 10343–10351. [Google Scholar] [CrossRef]
- Psachoulia, E.; Bond, P.J.; Sansom, M.S.P. MD Simulations of Mistic: Conformational Stability in Detergent Micelles and Water. Biochemistry 2006, 45, 9053–9058. [Google Scholar] [CrossRef] [PubMed]
- Le Maire, M.; Champeil, P.; Moller, J.V. Interaction of membrane proteins and lipids with solubilizing detergents. Biochim. Biophys. Acta 2000, 1508, 86–111. [Google Scholar] [CrossRef]
- Alonso, H.; Roujeinikova, A. Characterization and two-dimensional crystallization of membrane component AlkB of the medium-chain alkane hydroxylase system from Pseudomonas putida GPo1. Appl. Environ. Microbiol. 2012, 78, 7946–7953. [Google Scholar] [CrossRef] [PubMed]
- Mast, R.C.; Haynes, L.V. The use of the fluorescent probes perylene and magnesium 8-anilinonaphthalene-1-sulfonate to determine the critical micelle concentration of surfactants in aqueous solution. J. Colloid Interface Sci. 1975, 53, 35–41. [Google Scholar] [CrossRef]
- Kang, Y.; Zhou, X.E.; Gao, X.; He, Y.; Liu, W.; Ishchenko, A.; Barty, A.; White, T.A.; Yefanov, O.; Han, G.W.; et al. Crystal structure of rhodopsin bound to arrestin by femtosecond X-ray laser. Nature 2015, 523, 561–567. [Google Scholar] [CrossRef] [PubMed]
- White, J.F.; Noinaj, N.; Shibata, Y.; Love, J.; Kloss, B.; Xu, F.; Gvozdenovic-Jeremic, J.; Shah, P.; Shiloach, J.; Tate, C.G.; et al. Structure of the agonist-bound neurotensin receptor. Nature 2012, 490, 508–513. [Google Scholar] [CrossRef] [PubMed]
- Hasko Paradies, H. Shape and size of a nonionic surfactant micelle. Triton X-100 in aqueous solution. J. Phys. Chem. 1980, 84, 599–607. [Google Scholar] [CrossRef]
- Streletzky, K.; Phillies, G.D.J. Temperature dependence of triton X-100 micelle size and hydration. Langmuir 1995, 11, 42–47. [Google Scholar] [CrossRef]
- Ashani, Y.; Catravas, G.N. Highly reactive impurities in Triton X-100 and Brij 35: Partial characterization and removal. Anal. Biochem. 1980, 109, 55–62. [Google Scholar] [CrossRef]
- McPherson, A.; Gavira, J.A. Introduction to protein crystallization. Acta Crystallogr. F Struct. Biol. Commun. 2014, 70, 2–20. [Google Scholar] [CrossRef] [PubMed]
- Baker, M.R.; Fan, G.; Serysheva, I.I. Single-particle cryo-EM of the ryanodine receptor channel in an aqueous environment. Eur. J. Transl. Myol. 2015, 25, 35–48. [Google Scholar] [CrossRef] [PubMed]
- Abeyrathne, P.D.; Arheit, M.; Kebbel, F.; Castano-Diez, D.; Goldie, K.N.; Chami, M.; Stahlberg, H.; Renault, L.; Kühlbrandt, W. Chapter 119–1.15 Analysis of 2-D Crystals of Membrane Proteins by Electron Microscopy. In Comprehensive Biophysics; Elsevier: Amsterdam, The Netherlands, 2012; pp. 277–310. [Google Scholar]
- Smith, E.L.; Pickels, E.G. Micelle Formation in Aqueous Solutions of Digitonin. Proc. Natl. Acad. Sci. USA 1940, 26, 272–277. [Google Scholar] [CrossRef] [PubMed]
- Chae, P.S.; Rasmussen, S.G.F.; Rana, R.R.; Gotfryd, K.; Kruse, A.C.; Manglik, A.; Cho, K.H.; Nurva, S.; Gether, U.; Guan, L.; et al. A new class of amphiphiles bearing rigid hydrophobic groups for solubilization and stabilization of membrane proteins. Chemistry 2012, 18, 9485–9490. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Chen, J. Atomic Structure of the Cystic Fibrosis Transmembrane Conductance Regulator. Cell 2016, 167, 1586–1597. [Google Scholar] [CrossRef] [PubMed]
- Letts, J.A.; Fiedorczuk, K.; Sazanov, L.A. The architecture of respiratory supercomplexes. Nature 2016, 537, 644–648. [Google Scholar] [CrossRef] [PubMed]
- Kaufmann, T.C.; Engel, A.; Rémigy, H.-W. A novel method for detergent concentration determination. Biophys. J. 2006, 90, 310–317. [Google Scholar] [CrossRef] [PubMed]
- D’Avanzo, N.; McCusker, E.C.; Powl, A.M.; Miles, A.J.; Nichols, C.G.; Wallace, B.A. Differential lipid dependence of the function of bacterial sodium channels. PLoS ONE 2013, 8, e61216. [Google Scholar] [CrossRef] [PubMed]
- Zana, R.; Weill, C. Effect of temperature on the aggregation behaviour of nonionic surfactants in aqueous solutions. J. Phys. Lett. 1985, 46, 953–960. [Google Scholar] [CrossRef]
- Poulos, S.; Morgan, J.L.W.; Zimmer, J.; Faham, S. Bicelles coming of age: An empirical approach to bicelle crystallization. Methods Enzymol. 2015, 557, 393–416. [Google Scholar] [PubMed]
- Breyton, C.; Chabaud, E.; Chaudier, Y.; Pucci, B.; Popot, J.-L. Hemifluorinated surfactants: A non-dissociating environment for handling membrane proteins in aqueous solutions? FEBS Lett. 2004, 564, 312–318. [Google Scholar] [CrossRef]
- Efremov, R.G.; Leitner, A.; Aebersold, R.; Raunser, S. Architecture and conformational switch mechanism of the ryanodine receptor. Nature 2015, 517, 39–43. [Google Scholar] [CrossRef] [PubMed]
- Nasr, M.L.; Baptista, D.; Strauss, M.; Sun, Z.-Y.J.; Grigoriu, S.; Huser, S.; Plückthun, A.; Hagn, F.; Walz, T.; Hogle, J.M.; et al. Covalently circularized nanodiscs for studying membrane proteins and viral entry. Nat. Methods 2017, 14, 49–52. [Google Scholar] [CrossRef] [PubMed]
- Bae, H.E.; Mortensen, J.S.; Ribeiro, O.; Du, Y.; Ehsan, M.; Kobilka, B.K.; Loland, C.J.; Byrne, B.; Chae, P.S. Tandem neopentyl glycol maltosides (TNMs) for membrane protein stabilisation. Chem. Commun. 2016, 52, 12104–12107. [Google Scholar] [CrossRef] [PubMed]
- Von Heijne, G. Membrane protein structure prediction. Hydrophobicity analysis and the positive-inside rule. J. Mol. Biol. 1992, 225, 487–494. [Google Scholar] [CrossRef]
- Zhang, Q.; Ma, X.; Ward, A.; Hong, W.-X.; Jaakola, V.-P.; Stevens, R.C.; Finn, M.G.; Chang, G. Designing facial amphiphiles for the stabilization of integral membrane proteins. Angew. Chem. Int. Ed. Engl. 2007, 46, 7023–7025. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.C.; Bennett, B.C.; Hong, W.-X.; Fu, Y.; Baker, K.A.; Marcoux, J.; Robinson, C.V.; Ward, A.B.; Halpert, J.R.; Stevens, R.C.; et al. Steroid-based facial amphiphiles for stabilization and crystallization of membrane proteins. Proc. Natl. Acad. Sci. USA 2013, 110, E1203–E1211. [Google Scholar] [CrossRef] [PubMed]
- He, X.; Szewczyk, P.; Karyakin, A.; Evin, M.; Hong, W.-X.; Zhang, Q.; Chang, G. Structure of a cation-bound multidrug and toxic compound extrusion transporter. Nature 2010, 467, 991–994. [Google Scholar] [CrossRef] [PubMed]
- Chae, P.S.; Gotfryd, K.; Pacyna, J.; Miercke, L.J.W.; Rasmussen, S.G.F.; Robbins, R.A.; Rana, R.R.; Loland, C.J.; Kobilka, B.; Stroud, R.; et al. Tandem facial amphiphiles for membrane protein stabilization. J. Am. Chem. Soc. 2010, 132, 16750–16752. [Google Scholar] [CrossRef] [PubMed]
- Drew, D.; Lerch, M.; Kunji, E.; Slotboom, D.J.; de Gier, J.-W. Optimization of membrane protein overexpression and purification using GFP fusions. Nat. Methods 2006, 3, 303–313. [Google Scholar] [CrossRef] [PubMed]
- Kawate, T.; Gouaux, E. Fluorescence-detection size-exclusion chromatography for precrystallization screening of integral membrane proteins. Structure 2006, 14, 673–681. [Google Scholar] [CrossRef] [PubMed]
- Desuzinges-Mandon, E.; Agez, M.; Pellegrin, R.; Igonet, S.; Jawhari, A. Novel systematic detergent screening method for membrane proteins solubilization. Anal. Biochem. 2017, 517, 40–49. [Google Scholar] [CrossRef] [PubMed]
- Lantez, V.; Nikolaidis, I.; Rechenmann, M.; Vernet, T.; Noirclerc-Savoye, M. Rapid automated detergent screening for the solubilization and purification of membrane proteins and complexes. Eng. Life Sci. 2015, 15, 39–50. [Google Scholar] [CrossRef]
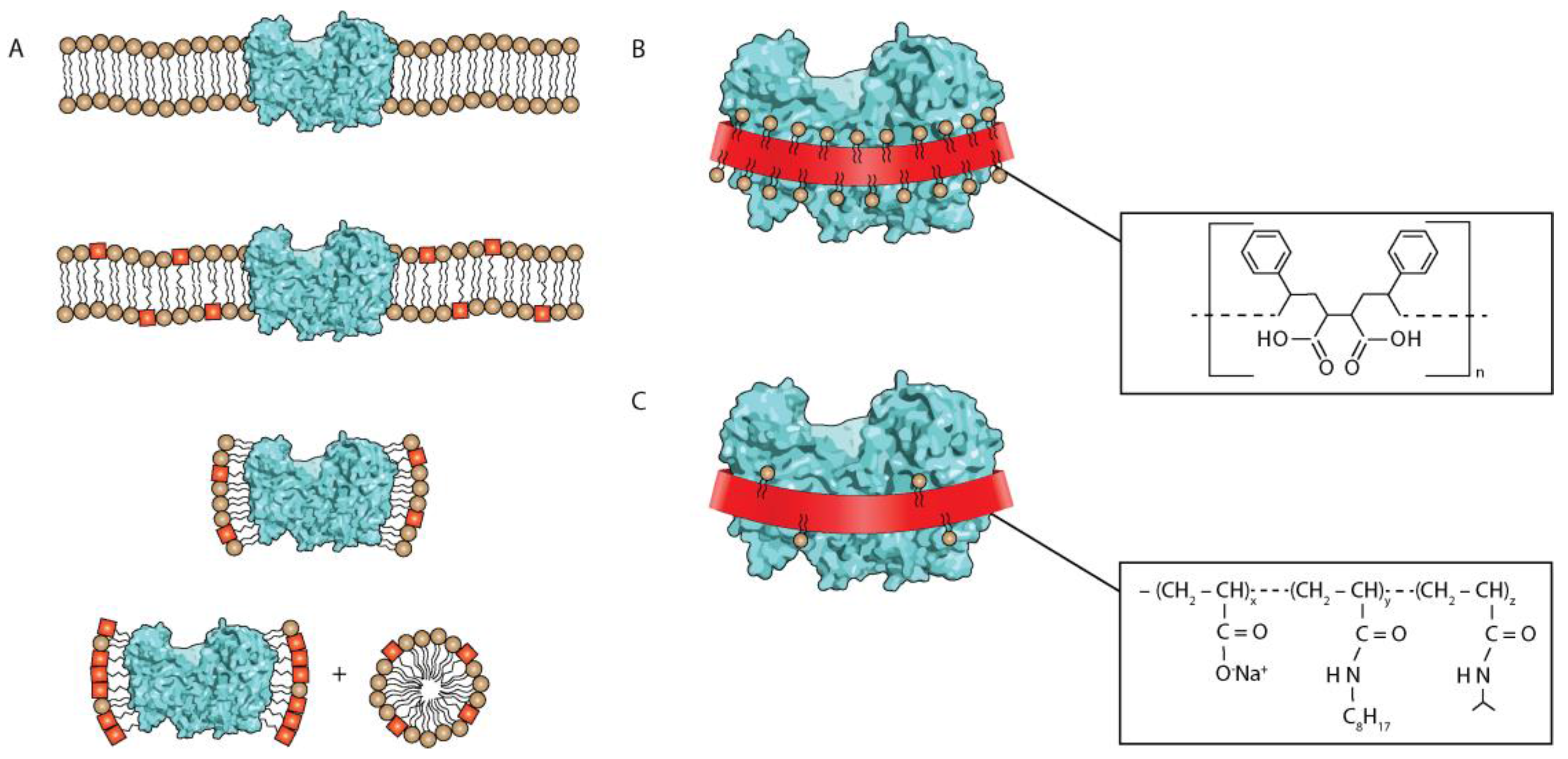
Figure 1.
(A) Extraction of a membrane protein (cyan) from the lipid bilayer (lipid heads as circles, tails as lines and detergent molecules as red squares). (From top to bottom) the concentration of detergent is increased, until the protein is extracted in the form of the complex with detergent (and some residual lipids), surrounded by free detergent-lipid micelles. Note that detergent/lipid molecules are omitted in the plane towards a viewer. (B) A membrane protein encircled with the styrene maleic acid (SMA) belt (the chemical structure is shown in the insert). Note that SMA cuts out a fraction of membrane with the protein, thus maintaining the endogenous lipids. (C) A membrane protein encircled with the amphipol belt (the chemical structure of the most commonly used amphipol A8-35 is shown in the insert). Since amphipols are applied only after extraction with a detergent, there are far fewer lipid molecules bound to the protein.
Figure 1.
(A) Extraction of a membrane protein (cyan) from the lipid bilayer (lipid heads as circles, tails as lines and detergent molecules as red squares). (From top to bottom) the concentration of detergent is increased, until the protein is extracted in the form of the complex with detergent (and some residual lipids), surrounded by free detergent-lipid micelles. Note that detergent/lipid molecules are omitted in the plane towards a viewer. (B) A membrane protein encircled with the styrene maleic acid (SMA) belt (the chemical structure is shown in the insert). Note that SMA cuts out a fraction of membrane with the protein, thus maintaining the endogenous lipids. (C) A membrane protein encircled with the amphipol belt (the chemical structure of the most commonly used amphipol A8-35 is shown in the insert). Since amphipols are applied only after extraction with a detergent, there are far fewer lipid molecules bound to the protein.
Figure 2.
The bar representation (in %) of different detergents used for (A) purification and (B) crystallization. Panels (C) and (D) include data only for the time period of 2006–2016.
Figure 2.
The bar representation (in %) of different detergents used for (A) purification and (B) crystallization. Panels (C) and (D) include data only for the time period of 2006–2016.
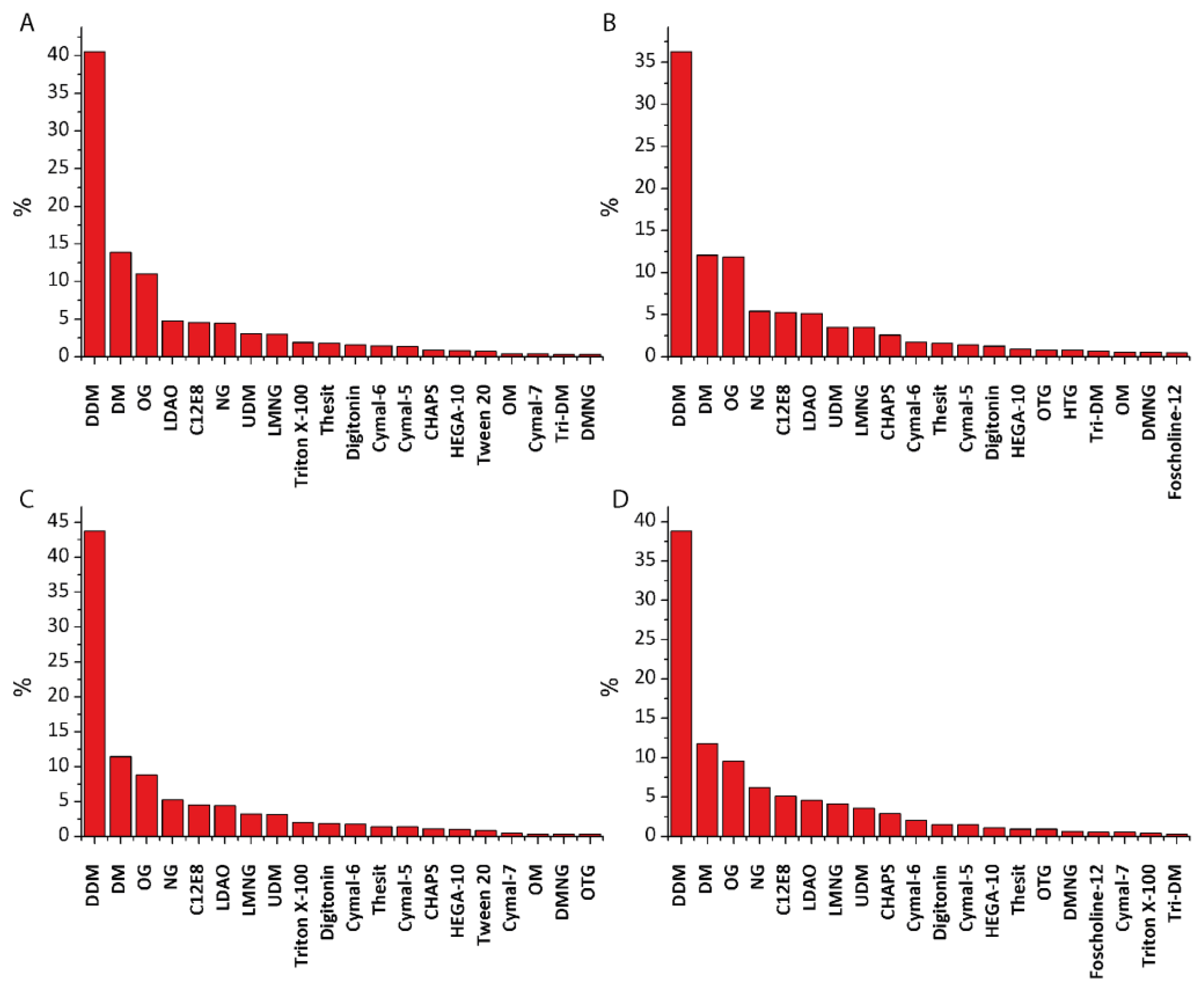
Figure 3.
Structure of Layrul maltoside (DDM) (chemical formula is C24H46O11).
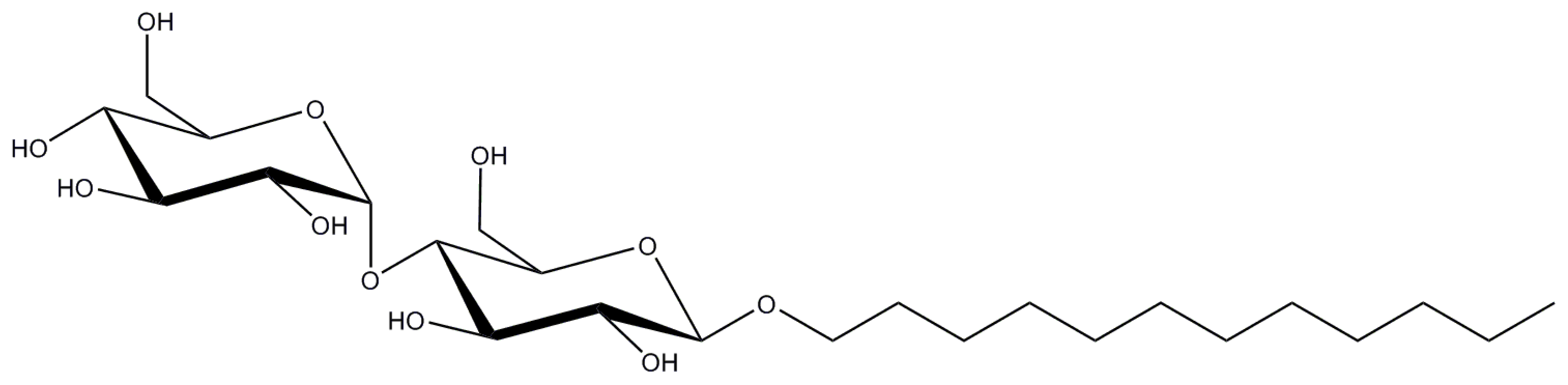
Figure 4.
Structure of Decyl maltoside (DM) (chemical formula is C22H44O11).
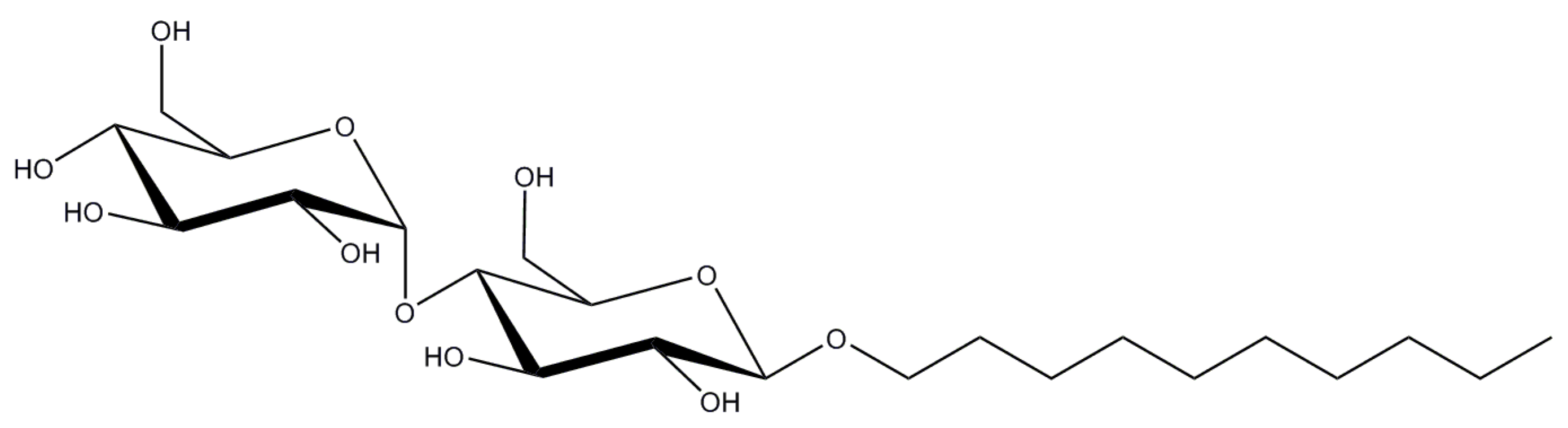
Figure 5.
Structure of Octyl glycoside (OG) (chemical formula is C14H28O6). OG has gained its popularity for studies on (bacterio-) rhodopsins, photosynthetic complexes and aquaporins.
Figure 5.
Structure of Octyl glycoside (OG) (chemical formula is C14H28O6). OG has gained its popularity for studies on (bacterio-) rhodopsins, photosynthetic complexes and aquaporins.

Figure 6.
Structure of Nonyl glucoside (NG) (chemical formula is C15H30O6).
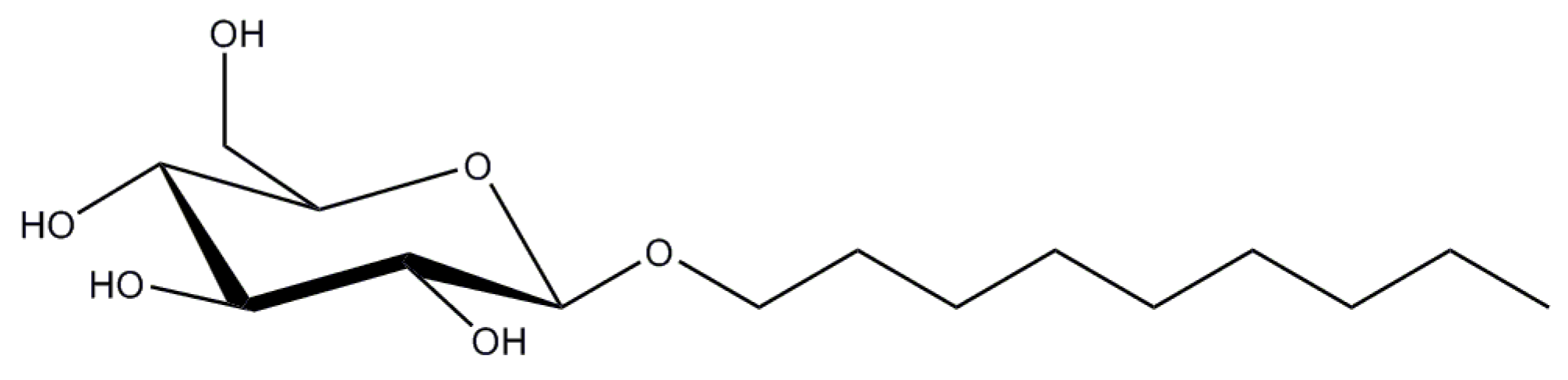
Figure 7.
Structure of dodecyldimethylamine oxide (LDAO) (chemical formula is C14H31NO).
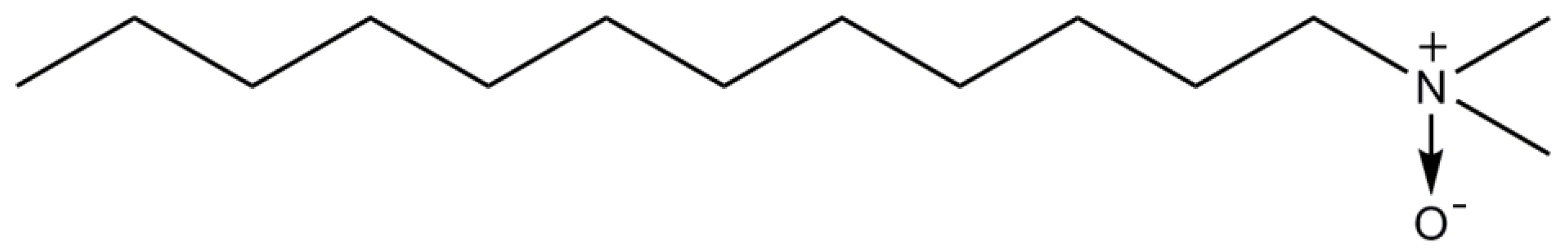
Figure 8.
Structure of C12E8 and C12E9 (chemical formula is (C2H4O)nC12H26O, n ~ 8 or 9).
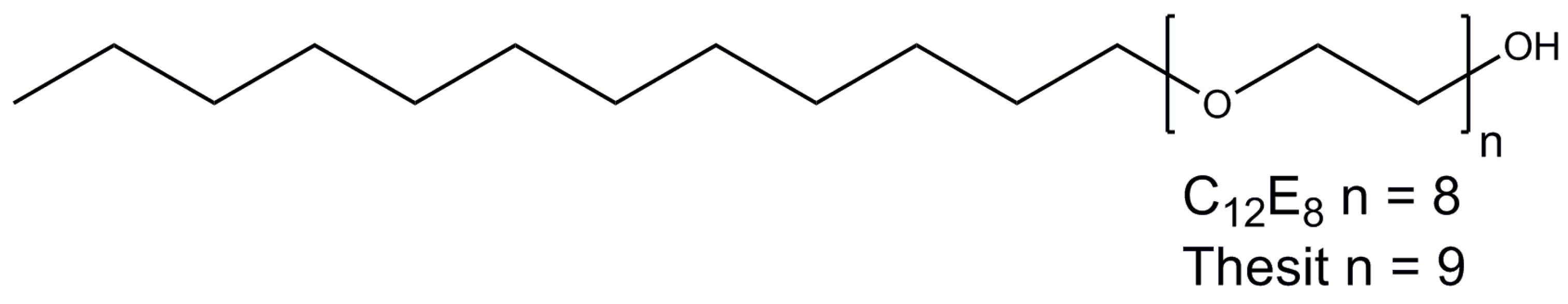
Figure 9.
Structure of n-Undecyl-β-d-maltopyranoside (UDM) (chemical formula is C23H44O11).
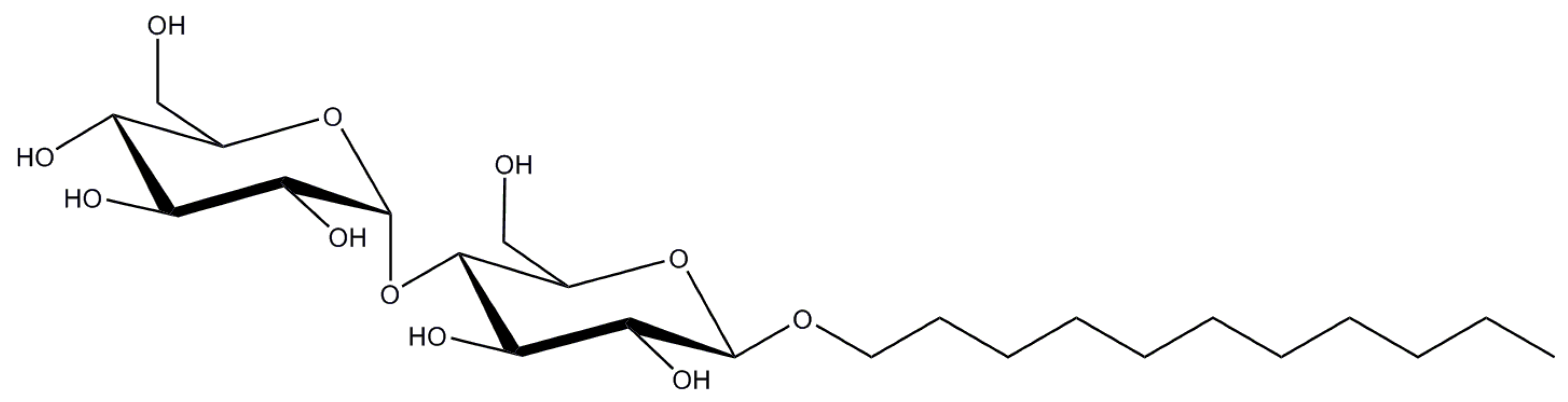
Figure 10.
Structure of Lauryl maltose neopentyl glycol (LMNG) (chemical formula is C47H88O26).
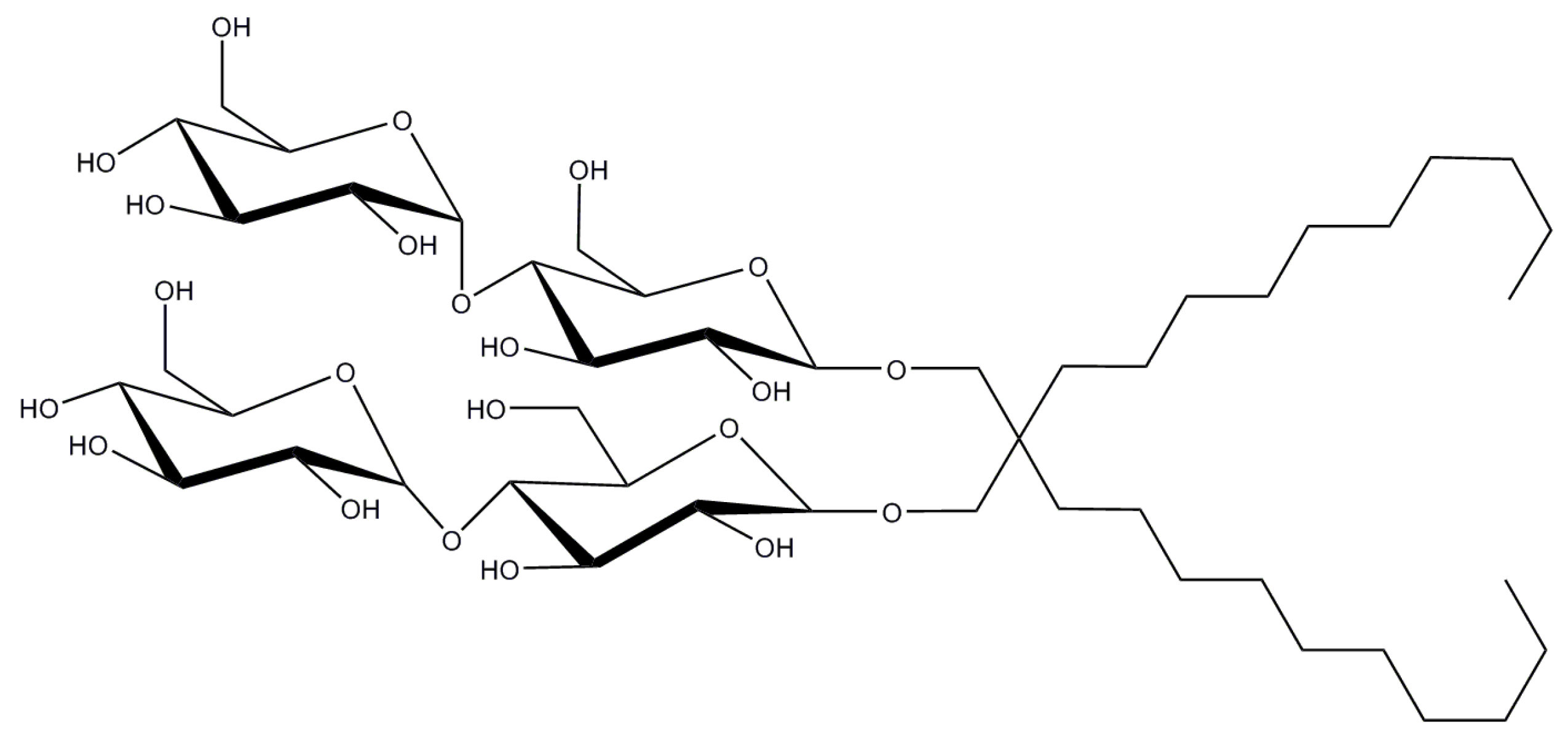
Figure 11.
Structure of Triton X-100 (chemical formula is C14H22O(C2H4O)n where n = 9, 10).
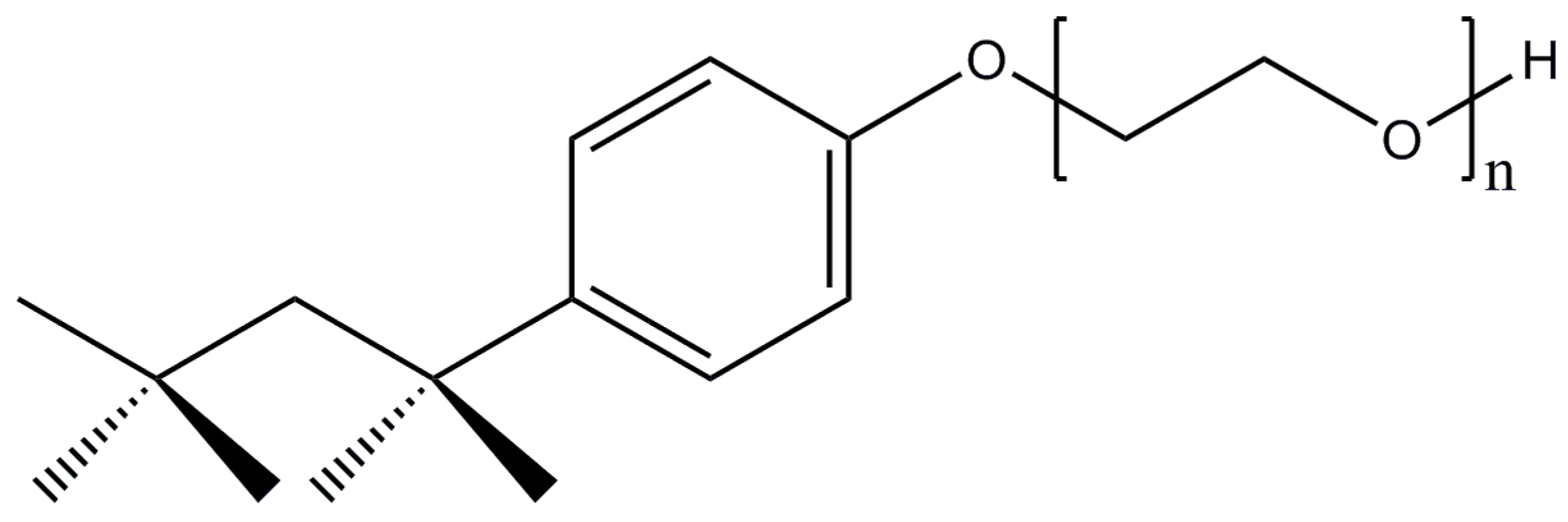
Figure 12.
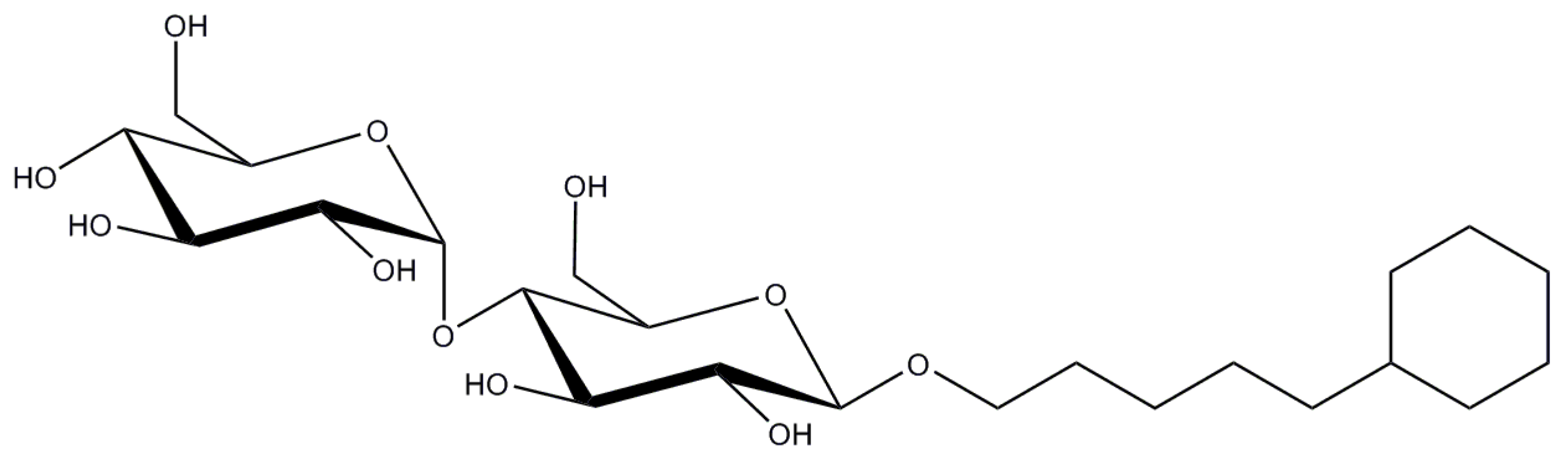
Structure of digitonin (chemical formula is C₅₆H₉₂O₂₉).
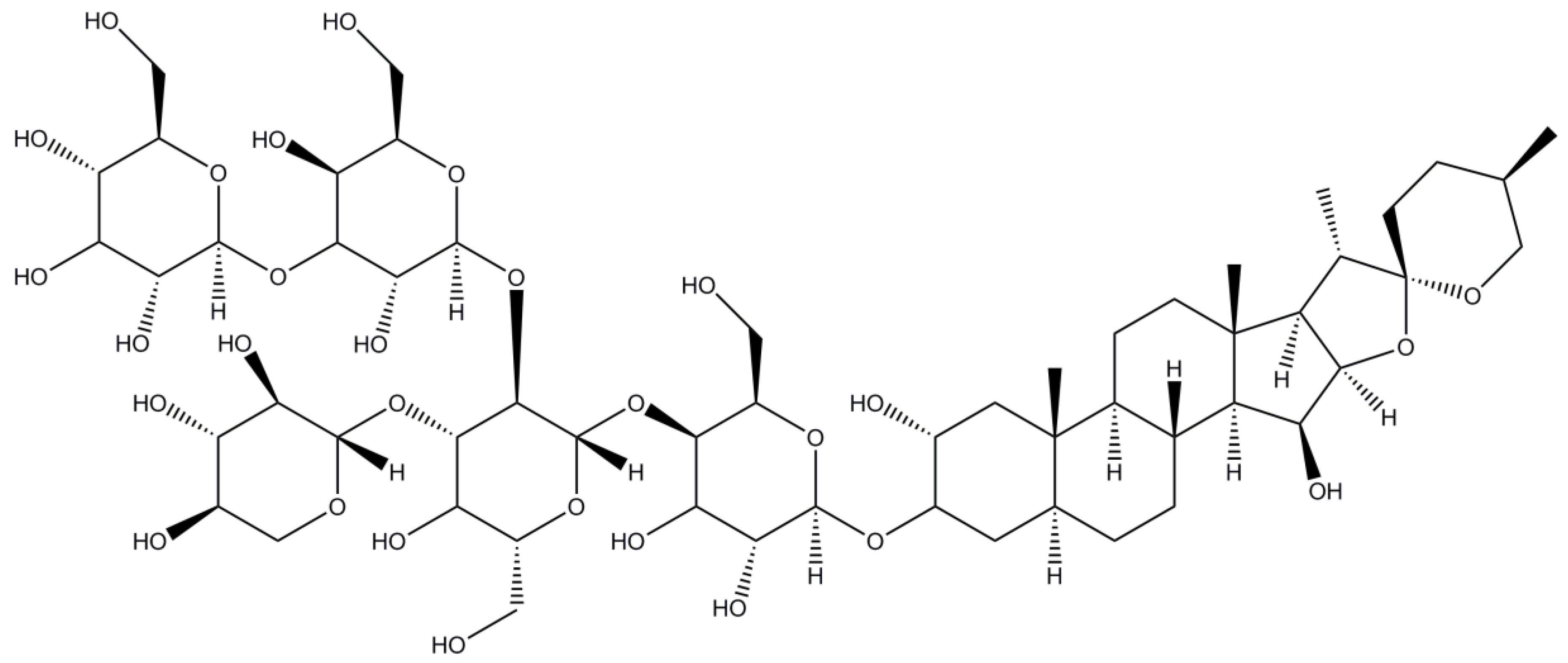
Figure 13.
Structure of Cymal-5 (chemical formula is C23H42O11; Cymal-6 has an extra carbon atom in its tail with formula C24H44O11).
Figure 13.
Structure of Cymal-5 (chemical formula is C23H42O11; Cymal-6 has an extra carbon atom in its tail with formula C24H44O11).

Figure 14.
Structure of CHAPS and CHAPSO (chemical formula is C32H58N2O7S / C32H58N2O8S).

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
The properties of the most commonly used detergents.
| Detergent | CMC (%/mM) | Aggregation Number, N 1 | Micelle Size Mw (kDa) | Mw (Da) |
|---|---|---|---|---|
| DDM | 0.0087/0.17 | 80–150 | 65–70 | 510.6 |
| DM | 0.087/1.8 | 69 | 40 | 482.6 |
| OG | 0.53/20 | 30–100 | 25 | 292.4 |
| NG | 0.20/6.5 | 133 | 85 | 306.4 |
| LDAO | 0.023/1-2 | 76 | 21.5 | 229.4 |
| C12E8 | 0.005/0.09 | 90–120 | 66 | 538.7 |
| C12E9 | 0.003/0.05 | 90 2 | 83 | 582.8 |
| UDM | 0.029/0.59 | 71 | 50 | 496.6 |
| LMNG | 0.001/0.01 | ~400 3 | 91–393 | 1069.2 |
| Triton X-100 | 0.01/0.2 | 75–165 | 60–90 | 624.8 (av.) |
| Digitonin | 0.002/0.5 | 60 4 | 70 | 1229.3 |
| Cymal-5 | 0.12/2.5 | 47 | 23 | 494.6 |
| Cymal-6 | 0.028/0.56 | 91 | 32 | 508.6 |
| CHAPS | 0.5/8–10 | 10 | 6 | 614.9 |
| CHAPSO | 0.5/8–10 | 11 | 7 | 630.9 |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Stetsenko, A.; Guskov, A. An Overview of the Top Ten Detergents Used for Membrane Protein Crystallization. Crystals 2017, 7, 197. https://doi.org/10.3390/cryst7070197
AMA Style
Stetsenko A, Guskov A. An Overview of the Top Ten Detergents Used for Membrane Protein Crystallization. Crystals. 2017; 7(7):197. https://doi.org/10.3390/cryst7070197
Chicago/Turabian StyleStetsenko, Artem, and Albert Guskov. 2017. "An Overview of the Top Ten Detergents Used for Membrane Protein Crystallization" Crystals 7, no. 7: 197. https://doi.org/10.3390/cryst7070197
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.