Human Defensins: Potential Tools for Clinical Applications

1
Department of Periodontology, Operative and Preventive Dentistry, University of Bonn, Welschnonnenstr. 17, D-53111 Bonn, Germany
2
Department of Oral & Maxillofacial Plastic Surgery, University of Bonn, Welschnonnenstr. 17, D-53111 Bonn, Germany
*
Author to whom correspondence should be addressed.
Polymers 2012, 4(1), 691-709; https://doi.org/10.3390/polym4010691
Submission received: 15 January 2012
/
Revised: 31 January 2012
/
Accepted: 10 February 2012
/
Published: 28 February 2012
(This article belongs to the Special Issue Antibacterial Polymers: From Natural Inspiration to Practical Applications)
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
: As components of the innate immune system, antimicrobial peptides in the form of human defensins play an important role in host defense by serving as the epithelial layer’s biochemical barrier against local infections. Recent studies have shown these molecules to have far more additional cellular functions besides their antimicrobial activity. Defensins play a role in cell division, attraction and maturation of immune cells, differentiation and reorganization of epithelial tissues, wound healing and tumor suppression. This multitude of function makes human defensins appear to be excellent tools for therapeutic approaches. These antimicrobial peptides may be used directly as a remedy against bacterial and viral infections. Furthermore, the application of human defensins can be used to promote wound healing and epithelial reorganization. In particular, human β-defensins have a strong impact on osteoblast proliferation and differentiation. Human β-defensins have already been applied as a vaccination against HIV-1. Another potentially useful characteristic of defensins is their suitability as diagnostic markers in cancer therapy. In particular, α-defensins have already been used for this purpose. Human α-defensin-3, for example, has been described as a tumor marker for lymphocytes. High gene expression levels of α-defensin-3 and -4 have been detected in benign oral neoplasia, α-defensin-6 is considered to be a tumor marker for colon cancer.
1. Introduction
Antimicrobial peptides (AMPs) are polypeptides of low molecular weight with less than 20 kDa. They are involved in host defense, but besides their antimicrobial activity they are involved in various cellular processes. One subfamiliy of mammalian AMPs are defensins. These polypeptides were first described in the 1980s and identified as components of the innate immune system. Shortly afterwards, they were found to have additional functions, other than just having antimicrobial properties. Human defensins are distributed in phagocytes and epithelial tissues. They share a framework of six disulphide-linked cysteine (cys) residues with a characteristic β-sheet structure and contain a number of positively charged amino acids, causing their typical cationic features under physiological conditions [1,2,3,4].
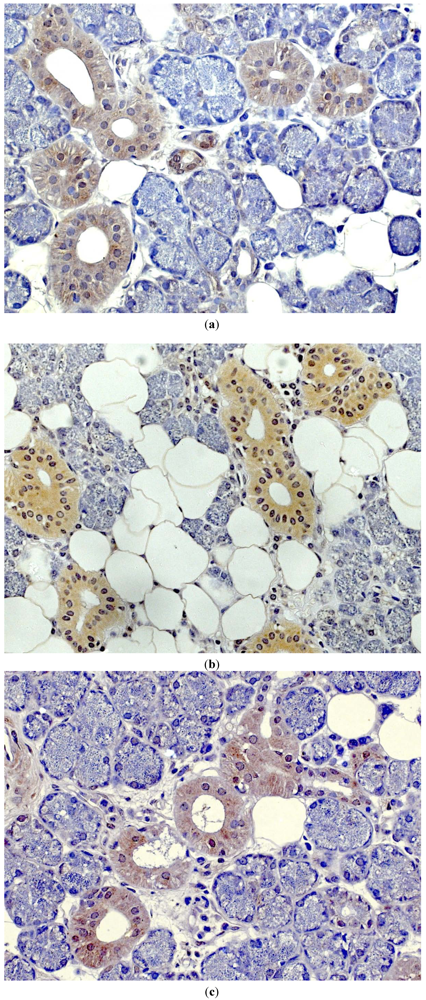
Human defensins are divided into two subgroups: α- and β-defensins. Both α- and β-defensins consist of a triple-stranded β-sheet structure, and have a molecular weight between 3 and 6 kDa. They mainly differ in the pairing of cystine-disulphide bridges. Cysteine residues in α-defensins are linked between 1–6/2–4/3–5, whereas β-defensins share a 1–5/2–4/3–6 cys-cys pattern. So far, four α-defensins have been isolated from granules of leukocytes. As these AMPs were found in granules of polymorphonuclear neutrophils, they were named human neutrophil peptides 1–4 (HNP1–4). Two α-defensins (DEFA), α-defensins-5 and -6 are distributed in intestinal Paneth cells. Human β-defensins (hBDs) are expressed in cell populations of various epithelia (skin, urogenital and respiratory tract, eye, oral tissues, salivary glands) (Figure 1a–c) and bone [1,2,4,5,6,7,8,9]. Three subtypes (hBD-1-3) have been isolated by biochemical means [10,11,12]. Human defensins are encoded as preproprecursor proteins, processed to mature peptides by specific cellular proteases. Defensin synthesis and release are regulated by bacterial and viral signals, inflammatory cytokines as well as growth factors [1,2,4]. Recently, it has been demonstrated that human β-defensins are involved in bisphosphonate-associated osteochemonecrosis of the jawbone [13]. Furthermore, these antimicrobial peptides are able to affect their own genes—either directly or indirectly by modulating gene activity and secretion of cytokines, which in consequence leads to feedback mechanisms on defensin genes [4,14,15,16].
Besides their antimicrobial activity, a number of cellular functions are triggered by defensins, when they bind as ligands to certain receptors. For instance, it has been demonstrated that hBD-2 is able to bind to the chemokine receptor 6 (CCR 6) [17]. HBD-2 also activates dendritic cells by binding to toll-like receptor 4 [18]. Furthermore, defensins play an active role in several growth-dependent processes, like wound healing and proliferation. HNP1–3, for example, enables wound closure in airway epithelial cell cultures [19]. Human β-defensins positively affect the rate of migration and proliferation of epidermal keratinocytes [20]. These cellular functions are dependent on an activated epidermal growth factor receptor [19,20]. HBDs also stimulate proliferation in various human cancer cells in vitro [15,16]. Human β-defensin-2 is involved in bone cell differentiation and mineralization of osteoblast-like cells [15]. Human defensins even seem to play a role in reproduction. Recently, male fertility has been described as dependent on active DEFB126 [21].
Figure 1.
Immunostaining (brownish color) for (a) hBD-1, (b) hBD-2 and (c) hBD-3 in healthy salivary gland tissue [7].
Figure 1.
Immunostaining (brownish color) for (a) hBD-1, (b) hBD-2 and (c) hBD-3 in healthy salivary gland tissue [7].

Furthermore, antimicrobial peptides strongly contribute to the functions of the innate immune system, when they act like local antibiotics.
Some defensins are constitutively expressed, like HNP1–3, DEFA5-6, or hBD-1. Gene expression of other defensins (hBD-2-3) is inducible by various inflammatory and microbial stimuli. The mechanism by which human defensins exert their biological, antimicrobial activity is based on their cationic and amphipathic molecular features. Antimicrobial peptides bind to negatively charged constituents of the bacterial cell surface and by forming pores in the lipid-bilayer, they permeabilize and disrupt the cytoplasma membrane—which leads in most cases to certain bacterial inactivation. Additionally, some defensins are also able to inhibit DNA, RNA and protein biosynthesis of intruding microbes [1,2,3,4].
Due to their wide variety of functions, human defensins are a promising tool for clinical application. For instance, these molecules could be directly applied on a lesion, in order to work as local antibiotics. Furthermore, their ability to influence and direct the adaptive immune system makes defensins promising immune modulators in systemic anti-inflammatory or anti-cancer treatment. In the treatment of guided tissue repair, defensins might be useful when applied locally after infection is under control, thus helping to overcome the problems of failed wound healing. Finally, their role in carcinogenesis makes them a fascinating target in the diagnosis and therapy of a growing number of cancers involving head and neck, bone (Figure 2), as well as the gastrointestinal and urogenitary tracts. Arguably, this is the most important role of human defensins, even though it is not yet completely understood and investigated.
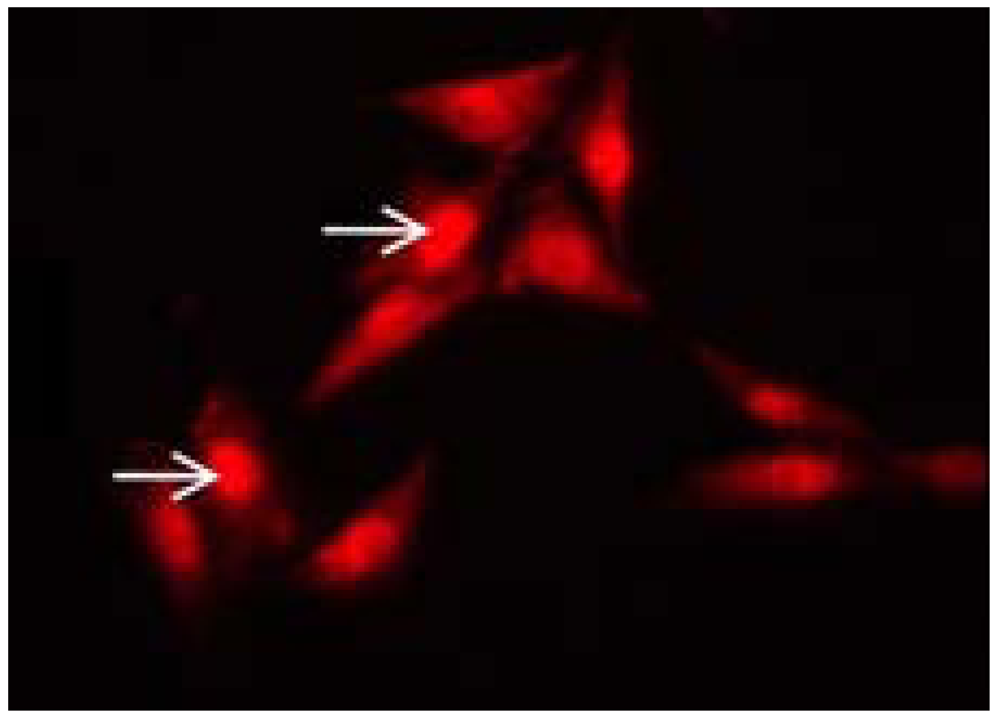
Figure 2.
Intracellular nuclear localization of hBD-1 (arrows) in MG63 cells using fluorescence microscopy.
Figure 2.
Intracellular nuclear localization of hBD-1 (arrows) in MG63 cells using fluorescence microscopy.

2. Antimicrobial Activity
Human α-defensins (DEFA) were first isolated from neutrophils. Therefore, they are also designated as human neutrophil peptides (HNP) [22]. Human polymorphonuclear leukocytes contain four members (HNP1–4) of this antimicrobial peptide subfamily [2]. Two more α-defensins, HD-5 and HD-6, also referred to as DEFA5 and DEFA6, have been detected in Paneth cells of the intestine [23,24].
HNP1–4 and HD-5 exhibit antimicrobial activities against Gram-positive bacteria, i.e., Staphylococcus aureus, and Gram-negative bacteria as Enterobacter aerogenes and Escherichia coli. No antibacterial activity could be detected for HD-6 [25]. HNP1 was highly effective at inactivating Mycobacterium tuberculosis [26,27]. HNP1–3 were able to bind to the lethal factor of the anthrax pathogen, Bacillus anthracis, thus causing the inhibition of its enzymatic activity [28].
In addition to their anti-bacterial capability, human α-defensins also exhibit a strong anti-viral potency. Human α-defensins, for example, inhibit infections of human immunodeficiency virus (HIV) and herpes simplex virus (HSV)in vitro [29] by binding to gp120 of HIV and CD4 receptors [30]. Furthermore, α-defensins can repress the replication of HIV-1 [31,32]. Their anti-HSV activity is caused by blocking viral adhesion to host cell surface molecules [33].
Human β-defensins (hBDs) are expressed by epithelial tissues [1,2]. To date, four hBDs were isolated (hBD-1–4) [2,34]. These antimicrobial peptides display their bactericidal activity against Gram-positive and -negative bacteria [11,12,35,36,37]. HBD-1 and hBD-3 share the ability to inactivate Candida species [38,39]. HBD-3 was shown to inhibit infection of HSV, whereas hBD-1 and -2 show no anti-viral effects [29]. HBD-2 and hBD-3 exhibit their antiviral activity against HIV by interacting with the virion particle and through modulation of the CXCR4 co-receptor [40]. Additionally, hBD-2 inhibits the synthesis of reverse transcribed DNA [41].
As several studies have shown that no bacterial or viral resistance against AMPs has developed so far [42,43,44], antimicrobial peptides could be a powerful tool in the therapy of acute or chronic infectious diseases in the future. As there is a growing number of treatment-resistant bacteria and viruses causing immense problems and exploding costs in public healthcare (Figure 3), it is a promising therapeutic concept to develop treatments involving the application of these molecules, sole or in combination with conventional antibiotics. In contrast with conventional antibiotics the allergic potential of AMP is very little.
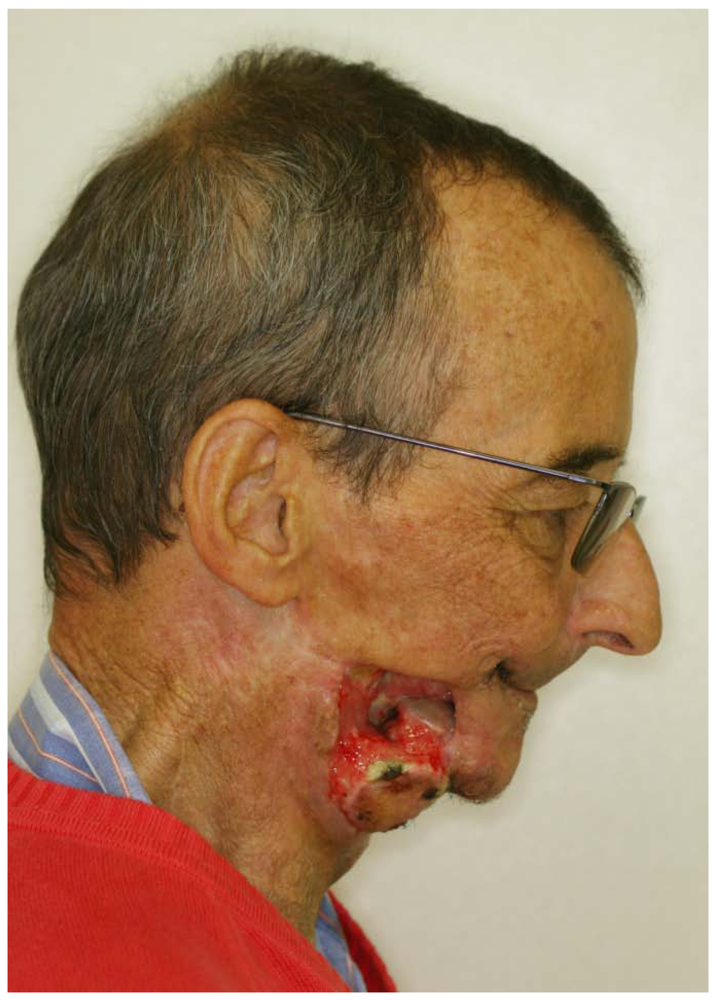
Figure 3.
Severe irradiation-induced osteomyelitis with destruction of the lower jaw and the surrounding soft tissue.
Figure 3.
Severe irradiation-induced osteomyelitis with destruction of the lower jaw and the surrounding soft tissue.

3. Immunomodulatory and Chemotactic Effects
Human defensins respond to various inflammatory stimuli, like tumor necrosis factor-α (TNF-α), interleukin-1β (IL-1β), lipopolysaccharide (LPS) as well as a number of bacterial surface antigens and toxins [12,37,45,46,47,48,49,50,51,52,53]. Besides their direct antimicrobial activity, defensins prevent the activation of toll-like receptors (TLRs) by blocking binding of LPS to LPS binding protein [54]. Furthermore, defensins cross-regulate pro-inflammatory cytokines and chemokines [4,14]. Alpha-defensins released from necrotic neutrophils are able to repress the secretion of various cytokines from macrophages, thus exhibiting anti-inflammatory effects [55]. Enhanced secretion of TNF-α or interferon-γ (IFN-γ) from neutrophils has been demonstrated to be caused by HNP1–3, thereby inducing increased phagocytotic activity of human macrophages [56]. Studies on human epidermal keratinocytes revealed that hBD-2 to -4 positively affect cytokines and chemokines, like IL-6, IL-10 or macrophage inflammatory protein-3α. Furthermore, these AMPs stimulate cell migration and proliferation [20]. Human β-defensins have anti-apoptotic effects on polymorphonuclear leukocytes by activation of the chemokine receptor (CCR) 6, which means they can be simultaneously down-regulating a pro-apoptotic factor and up-regulating an anti-apoptotic protein [57]. Human defensins also have a strong impact on the adaptive immune system [58]. They exhibit non-specific chemo-attractant functions for various immune cells [2] and they selectively chemo-attract different types of leukocytes [59,60,61]. Alpha-defensins enhance the migration of T-cells [62,63], while human β-defensins have chemotactic effects on dendritic cells [17,62]. HNP1–3 and hBD-1 exhibit chemotactic functions for monocytes [34]. HBD-2 is a chemotactic peptide for human neutrophils stimulated with TNF-α [60] as well as macrophages [34] and it also acts as a chemotaxin for human mast cells [34,59]. In addition, hBDs function as important chemotactic agents for immature dendritic cells [64]. HBD-2 and hBD-3 have also been detected as ligands of CCR 2 [64] and CCR 6 [17]. Furthermore, hBD-2 activates dendritic cells via the toll-like receptor (TLR) 4 [18]. HBD-3 interacts with TLR 1 and TLR 2 of monocytes and dendritic cells, thus activating MyD88 signaling [65].
Promising clinical approaches have been described by using methods of gene therapy as the genetic transformation of the antimicrobial peptide cathelicidin in a cystic fibrosis xenograft model, which leads to a conservation of the ability to disintegrate bacteria [66]. Additionally, a transient adenoviral gene therapy with LL-37 has been demonstrated to be effective for the treatment of cutaneous burn wound infections [67].
3. Tissue Reorganization and Wound Healing
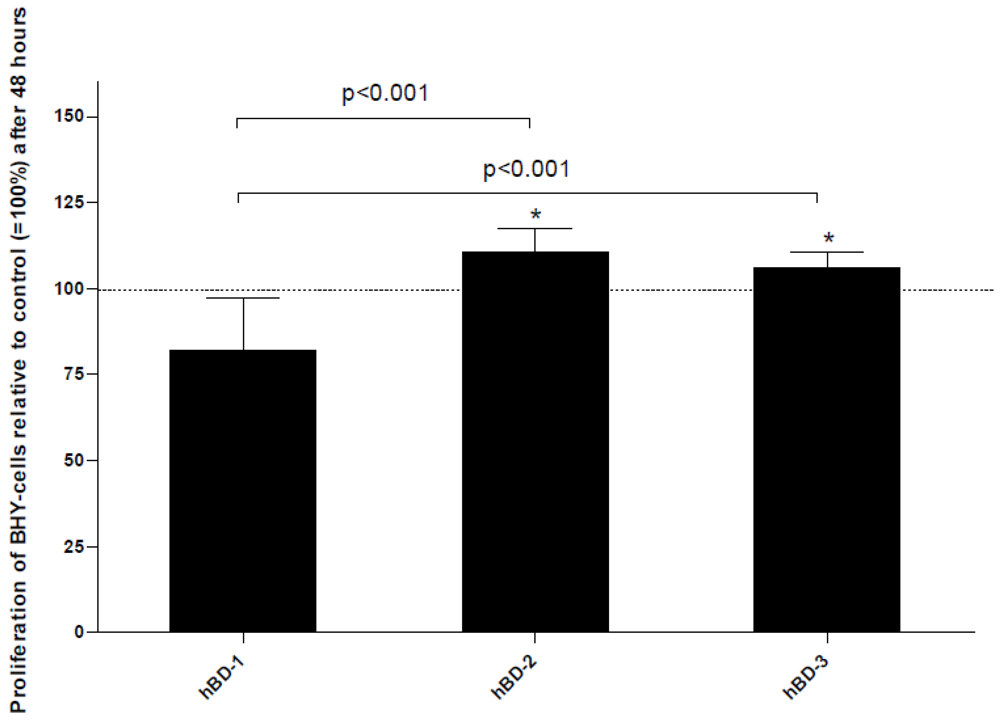
Besides their effects on host defense and immune modulation, human defensins also play an important role in wound healing. On the one hand, AMPs improve wound healing by inactivating bacteria, viruses and certain yeasts [67]. On the other, defensins participate in various cellular processes, like cell migration or proliferation, thus positively affecting wound healing [68]. HNP-1 and HNP-2 stimulate the proliferation and collagen synthesis of lung fibroblasts via β-catenin signaling. This observation might help to develop a therapy which could modulate the activity of fibroproliferative lesions as lung fibrosis [69]. HBD-1 to -4 enhance the migration of keratinocytes and show a positive effect on wound closure. Furthermore, they up-regulate the proliferation rate of human keratinocytes. It could be clearly demonstrated that these effects are based on the phosphorylation of the epidermal growth factor receptor [20]. This observation has been verified by other groups describing similar phenomena on different cells. HBD-2 has been shown to stimulate migration, proliferation and tube formation of endothelial cells [70]. In this context this antimicrobial peptide seems to be involved in the vascularization process which supports wound healing. HBD-2 and hBD-3 enhance the proliferation of osteoblast-like cells. Additionally, hBD-2 has a strong impact on the maturation of premature osteoblasts [15]. Furthermore, hBD-1, -2, and -3 have been demonstrated to stimulate the proliferation of tumor cells in vitro [16] (Figure 4). Apart from that, the gene products of human β-defensins cross-regulate their own gene expression [15,16].
Figure 4.
Significant changes in BHY cell proliferation after stimulation with 50 nM of hBD-1, -2 and -3 after 48 h are shown. Mean values of n = 12 experiments + standard deviation (SD) are depicted. Differences compared to the control are marked with * (p< 0.05).
Figure 4.
Significant changes in BHY cell proliferation after stimulation with 50 nM of hBD-1, -2 and -3 after 48 h are shown. Mean values of n = 12 experiments + standard deviation (SD) are depicted. Differences compared to the control are marked with * (p< 0.05).

Tissue regeneration is closely associated with wound healing. Human β-defensins exhibit various stimulatory effects on the proliferation and maturation of different cell types [15,16,20,69,70]. HBD-2 positively influences vascularization [70]. HBD-2 and hBD-3 might also be effective in bone tissue regeneration [15]. HBD-1 has been detected in the nuclei of keratinocytes and tumor cells [7,8,71]. It is suggested that hBD-1 acts as a transcription factor in differentiation processes, protecting keratinocytes from apoptosis during epithelial reorganization [71,72].
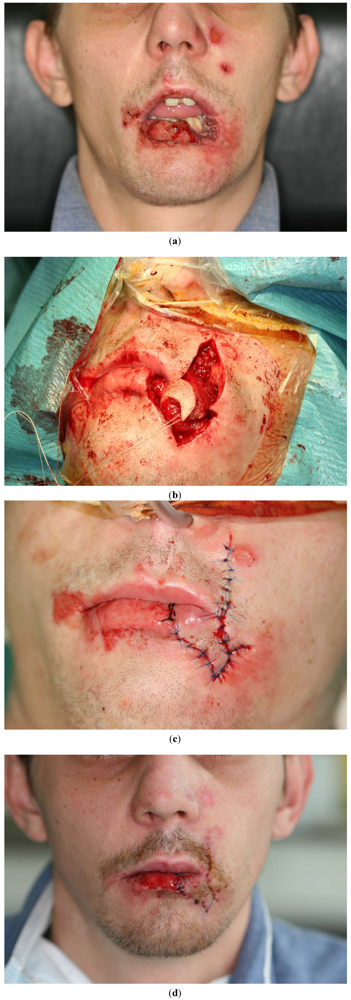
These observations indicate that the surgical treatment of extensive lesions could also benefit from a better understanding of AMPs. The application of human defensins could be useful to assist wound healing and epithelial reorganization. By local application, these antimicrobial peptides may be used as a first step in fighting bacterial and viral infections—and then, secondly, individually designed defensins could be applied to accelerate wound healing in microbially-traumatized lesions. The treatment of extensive lesions in modern surgery involves delicate local flap (Figure 5(a–c)) and microvascular procedures. Frequently, only time-consuming presurgical treatments allow these procedures to be administered. Such presurgical treatments comprise, for instance, the application of negative pressure via a vacuum pump (KCI) or occlusive hydrocolloid dressings, which help to increase early epithelial migration and accelerate the proliferation of granulation tissue. A better understanding of the AMP functions in tissue regeneration and wound healing could make them an additional option in presurgical wound preparation. This could help patients with extended and complicated soft tissue and bone lesions, and could also mean they have to spend less time in hospital.
Figure 5.
Plastic reconstruction after traumatic tissue loss in the lateral lower lip with sufficient reconstruction of the corner of the mouth (Operation performed by Dr. Marcus Teschke).
Figure 5.
Plastic reconstruction after traumatic tissue loss in the lateral lower lip with sufficient reconstruction of the corner of the mouth (Operation performed by Dr. Marcus Teschke).

4. Cancer
There seems to be a link between chronic infection and cancerogenesis, as epidemiological studies showed an association between inflammatory markers (CCL2, IL-6, IL-8, Cox-2) and tumor-formation [73,74,75]. In this context, interactions of the innate and adaptive immune system seem to play an important role, especially macrophages and natural killer cells [76]. Tumor-associated macrophages (TAM) represent the largest population of tumor-infiltrating inflammatory cells [76,77]. This type of macrophage leads to a tumor initiation by releasing IL-6 [77]. Tumor cells develop an immunosuppressive environment by secretion of IL-10 differentiating and recruiting M2-macrophages [77]. These macrophages enhance the immunosuppressive microenvironment by secretion of S100 proteins leading to the recruitment of immunosuppressive macrophages [76,77].
The correlation of chronic periodontitis with cancer has already been reported in the oral cavity, especially an association with tongue carcinoma and other epithelial head and neck tumors like oral squamous cell carcinoma [78,79].
In this putative sequence that leads from inflammation towards tumor, human defensins have moved into the focus of interest. A potential involvement of hBD-1 in carcinogenesis of epithelial tumors was first described in tumors of the urogenital tract, when a cancer-specific loss of hBD-1 gene expression was described in 90% of renal clear cell carcinomas and 82% of malignant prostate cancers, whereas in benign epithelium the expression of hBD-1 was intact [80,81]. A decreased hBD-1 expression has also been reported in oral squamous cell carcinoma [82]. As human defensins are involved in chronic periodontal inflammation as well as the carcinogenesis of oral tumors, it is an interesting new hypothesis that they might act as key-molecules in the inflammation-tumor-sequence [7,8,16,83,84,85,86]. On the one hand DEFB1 encoding for hBD-1 has been identified as a major periodontitis-associated gene [87] with important functions in local host defense. On the other, hBD-1 was found to be an important factor in proliferation control of oral cancers [7,8,16,83,85]. In regulating proliferation in oral squamous cell carcinoma cells, hBD-1 inhibits proliferation as a tumorsuppressor [80,81,82,83] whereas hBD-3 promotes proliferation as a potential protooncogene [83,84] (Figure 4). Exogenous administration of hBD-1, hBD-2 and hBD-3 has a strong impact on oral tumor cells (Figure 6). These AMPs cross-regulate their own gene expression and stimulate proliferation of oral squamous cell carcinoma cells in vitro [16]. HBD-2 and hBD-3 stimulate the proliferation of osteosarcoma cells. Additionally, hBD-2 enhances the maturation process of osteoblastic differentiation [15]. Apart from their anti-inflammatory capability of destroying various bacteria, viruses and yeasts, alpha-defensins also destroy the cell membranes of a number of different tumors in the mouse model and in human cancer cells in vitro: For instance stimulation with HNP1–3 leads to the disintegration of multiple myeloma cells [88].
One promising strategy to use human defensins in cancer therapy might be the topical application of hBD-1 as combined surface active protein and tumor suppressor: hBD-1’s destructive function on cell membranes would destroy tumor cells, and a high topic hBD-1 concentration would lead to a down-regulation of tumor cell proliferation. This could be supported by the application of specific antibodies against the oncogene hBD-3, thus causing an additional decrease of tumor cell proliferation. A number of adhesive dressings for the mucosa do exist. They can contain extracts from calf blood, including anorganic salts, amino acids, polypeptides and purines, but no proteins or antigenic substances or blood group characteristics and could therefore be loaded with recombined hBD-1.
Figure 6.
Relative hBD-3 gene expression of BHY cells after hBD-1 stimulation (50 nM). Statistically significant differences compared to the unstimulated control are shown with asterisks(*=p < 0.05). Mean values of n = 6 experiments + SD are depicted.
Figure 6.
Relative hBD-3 gene expression of BHY cells after hBD-1 stimulation (50 nM). Statistically significant differences compared to the unstimulated control are shown with asterisks(*=p < 0.05). Mean values of n = 6 experiments + SD are depicted.

Another possible method is the genetic transformation of cancer tissue by adenoviral vector systems. Recently, an in situ tumor vaccine based on human defensins has been developed [89]: An injection of adenovirus particles, encoding for hBD-2, directly into the tumor-tissue leads towards a cellular release of tumor antigens and hBD-2, which recruits dendritic cells, inducing local host defense against the presented antigens [89].
Besides the direct application of hBDs, another potential application could be as diagnostic markers in early cancer development and risk estimation for the malignant transformation of formerly benign lesions. Particularly, α-defensins have already been used for this purpose. Human α-defensin-3 has been described as a tumor marker for lymphocytes [90]. High gene expression levels of α-defensin-3 and -4 have been detected in benign oral neoplasia [85,86]. Also in the digestive tract, human α-defensin-6 is considered to be a tumor marker for benign epithelial colon cancers, such as colon adenoma [91].
5. Concluding Remarks
Human defensins are more than just antimicrobial peptides. These molecules are involved in various cellular processes. This multitude of functions makes AMPs a promising tool for specific clinical application. Antimicrobial peptides have been successfully applied in cystic fibrosis. They were applied via aerosolization [92]. Vector-based mediated delivery of gene encoding for antimicrobial peptides were used in cancer therapy [93]. However, systemic treatment with intravenous application of AMPs bears an immense risk to healthy cells due to the possibility of adverse reactions. To overcome this problem, diastereomers of antimicrobial peptides with D-configured amino acids were synthesized and successfully applied by topical and subcutaneous treatment [94,95]. Hence, many efforts have recently been made to focus on the pharmaceutical administration and modification of antimicrobial peptides as new leads for clinical therapeutic applications [96,97,98,99,100,101].
Acknowledgements
The authors are grateful to BONFOR research foundation of the Medical Faculty of the University of Bonn. The authors are supported by the German Research Foundation (KFO 208; TP10). Special Thanks to Marcus Teschke for the excellent photos depicted in Figure 5(a–c) and Jens Schröder for critically reading the manuscript.
References
- Ganz, T. Defensins: Antimicrobial peptides of innate immunity. Nat. Rev. Immunol. 2003, 3, 710–720. [Google Scholar] [CrossRef]
- Lehrer, R.I. Primate defensins. Nat. Rev. Microbiol. 2004, 2, 727–738. [Google Scholar]
- Brogden, K.A. Antimicrobial peptides: Pore formers or metabolic inhibitors in bacteria? Nat. Rev. Microbiol. 2005, 3, 238–250. [Google Scholar] [CrossRef]
- Hazlett, L.; Wu, M. Defensins in innate immunity. Cell Tissue Res. 2011, 343, 175–188. [Google Scholar] [CrossRef]
- Mathews, M.; Jia, H.P.; Guthmiller, J.M.; Losh, G.; Graham, S.; Johnson, G.K.; Tack, B.F.; McCray, P.B. Production of β-defensin antimicrobial peptides by the oral mucosa and salivary glands. Infect. Immun. 1999, 67, 2740–2745. [Google Scholar]
- Abiko, Y.; Saitoh, M.; Nishimura, M.; Yamazaki, M.; Sawamura, D.; Kaku, T. Role of β-defensins in oral epithelial health and disease. Med. Mol. Morphol. 2007, 40, 179–184. [Google Scholar] [CrossRef]
- Wenghoefer, M.; Pantelis, A.; Dommisch, H.; Götz, W.; Reich, R.; Bergé, S.; Martini, M.; Allam, J.P.; Jepsen, S.; Merkelbach-Bruse, S.; Fischer, HP.; Novak, N.; Winter, J. Nuclear hBD-1 accumulation in malignant salivary gland tumors. BMC Cancer 2008, 8. [Google Scholar]
- Pantelis, A.; Wenghoefer, M.; Haas, S.; Merkelbach-Bruse, S.; Pantelis, D.; Jepsen, S.; Bootz, F.; Winter, J. Down regulation and nuclear localization of human beta-defensin-1 in pleomorphic adenomas of salivary glands. Oral Oncol. 2009, 45, 526–530. [Google Scholar] [CrossRef]
- Warnke, P.H.; Springer, I.N.; Russo, P.A.J.; Wiltfang, J.; Essig, H.; Kosmahl, M.; Sherry, E.; Acil, Y. Innate immunity in human bone. Bone 2006, 38, 400–408. [Google Scholar]
- Bensch, K.W.; Raida, M.; Mägert, H.J.; Schulz-Knappe, P.; Forssmann, W.G. hBD-1: A novel beta-defensin from human plasma. FEBS Lett. 1995, 368, 331–335. [Google Scholar] [CrossRef]
- Harder, J.; Bartels, J.; Christophers, E.; Schröder, J.M. A peptide antibiotic from human skin. Nature 1997, 387, 861. [Google Scholar]
- Harder, J.; Bartels, J.; Christophers, E.; Schröder, J.M. Isolation and characterization of human β-defensin-3, a novel human inducible peptide antibiotic. J. Biol. Chem. 2001, 276, 5707–5713. [Google Scholar]
- Stockmann, P.; Wehrhan, F.; Schwarz-Furlan, S.; Stelzle, F.; Trabert, S.; Neukam, F.W.; Nkenke, E. Increased human defensine levels hint at an inflammatory etiology of bisphosphonate-associated osteonecrosis of the jaw: An immunohistological study. J. Transl. Med. 2011, 9. [Google Scholar]
- Presicce, P.; Giannelli, S.; Taddeo, A.; Villa, M.L.; Della Bella, S. Human defensins activate monocyte-derived dendritic cells, promote the production of proinflammatory cytokines, and up-regulate the surface expression of CD91. J. Leukoc. Biol. 2009, 86, 941–948. [Google Scholar] [CrossRef]
- Kraus, D.; Deschner, J.; Jäger, A.; Wenghoefer, M.; Bayer, S.; Jepsen, S.; Allam, J.; Novak, N.; Meyer, R.; Winter, J. Human β-defensins differently affect proliferation, differentiation, and mineralization of osteoblast-like MG63 cells. J. Cell. Physiol. 2011. [Google Scholar]
- Winter, J.; Pantelis, A.; Reich, R.; Martini, M.; Kraus, D.; Jepsen, S.; Allam, J.P.; Novak, N.; Wenghoefer, M. Human beta-defensin-1, -2, and -3 exhibit opposite effects on oral squamous cell carcinoma cell proliferation. Cancer Invest. 2011, 29, 196–201. [Google Scholar]
- Yang, D.; Chertov, O.; Bykovskaia, S.N.; Chen, Q.; Buffo, M.J.; Shogan, J.; Anderson, M.; Schröder, J.M.; Wang, J.M.; Howard, O.M.; Oppenheim, J.J. Beta-defensins: Linking innate and adaptive immunity through dendritic and T cell CCR6. Science 1999, 286, 525–528. [Google Scholar]
- Biragyn, A.; Ruffini, P.A.; Leifer, C.A.; Klyushnenkova, E.; Shakhov, A.; Chertov, O.; Shirakawa, A.K.; Farber, J.M.; Segal, D.M.; Oppenheim, J.J.; Kwak, L.W. Toll-like receptor 4-dependent activation of dendritic cells by beta-defensin 2. Science 2002, 298, 1025–1029. [Google Scholar]
- Aarbiou, J.; Verhoosel, R.M.; van Wetering, S.; De Boer, W.I.; van Krieken, J.H.; Litvinov, S.V.; Rabe, K.F.; Hiemstra, P.S. Neutrophil defensins enhance lung epithelial wound closure and mucin gene expression in vitro. Am. J. Respir. Cell Mol. Biol. 2004, 30, 193–201. [Google Scholar]
- Niyonsaba, F.; Ushio, H.; Nakano, N.; Ng, W.; Sayama, K.; Hashimoto, K.; Nagaoka, I.; Okumura, K.; Ogawa, H. Antimicrobial peptides human beta-defensins stimulate epidermal keratinocyte migration, proliferation and production of proinflammatory cytokines and chemokines. J. Invest. Dermatol. 2007, 127, 594–604. [Google Scholar]
- Tollner, T.L.; Venners, S.A.; Hollox, E.J.; Yudin, A.I.; Liu, X.; Tang, G.; Xing, H.; Kays, R.J.; Lau, T.; Overstreet, J.W.; Xu, X.; Bevins, C.L.; Cherr, G.N. A common mutation in the defensin DEFB126 causes impaired sperm function and subfertility. Sci. Transl. Med. 2011, 3. [Google Scholar]
- Ganz, T.; Selsted, M.E.; Szklarek, D.; Harwig, S.S.L.; Daher, K.; Bainton, D.F.; Lehrer, R.I. Defensins. J. Clin. Invest. 1985, 76, 1427–1435. [Google Scholar]
- Selsted, M.E.; Miller, S.I.; Henschen, A.H.; Quellette, A.J. Enteric defensins: Antibiotic peptide components of intestinal host defense. J. Cell Biol. 1992, 118, 929–936. [Google Scholar]
- Jones, D.E.; Bevins, C.L. Defensin-6 mRNA in human Paneth cells: Implication for antimicrobial peptides in host defense of the human bowel. FEBS Lett. 1993, 315, 187–192. [Google Scholar]
- Ericksen, B.; Wu, Z.; Lu, W.; Lehrer, R.I. Antibacterial activity and specificity of the six human alpha-defensins. Antimicrob. Agents Chemother. 2005, 49, 269–275. [Google Scholar] [CrossRef]
- Sharma, S.; Verma, I.; Khuller, GK. Antibacterial activity of human neutrophil peptide 1 against Mycobacterium tuberculosis H37Rv: In vitro and ex vivo study. Eur. Respir. J. 2000, 16, 112–117. [Google Scholar]
- Sharma, S.; Verma, I.; Khuller, GK. Therapeutic potential of human neutrophil peptide 1 against experimental tuberculosis. Antimicrob. Agents Chemother. 2001, 45, 639–640. [Google Scholar] [CrossRef]
- Verma, C.; Seebah, S.; Low, S.M.; Zhou, L.; Liu, S.P.; Li, J.; Beuerman, R.W. Defensins: Antimicrobial peptides for therapeutic development. Biotechnol. J. 2007, 2, 1353–1359. [Google Scholar]
- Hazrati, E.; Galen, B.; Lu, W.; Wang, W.; Ouyang, Y.; Keller, M.J.; Lehrer, R.I.; Herold, B.C. Human alpha- and beta-defensins block multiple steps in herpes simplex virus infection. J. Immunol. 2006, 177, 8658–8666. [Google Scholar]
- Wang, W.; Owen, S.M.; Rudolph, D.L.; Cole, A.M.; Hong, T.; Waring, A.J.; Lal, R.B.; Lehrer, R.I. Activity of alpha- and theta-defensins against primary isolates of HIV-1. J. Immunol. 2004, 173, 515–520. [Google Scholar]
- Mackewicz, C.E.; Yuan, J.; Tran, P.; Diaz, L.; Mack, E.; Selsted, M.E.; Levy, J.A. Alpha-defensins can have anti-HIV activity but are not CD8 cell anti-HIV factors. AIDS 2003, 26, F23–F32, Erratum in: AIDS 2003, 26, F31.. [Google Scholar]
- Chang, T.L.; Vargas, J., Jr.; DelPortillo, A.; Klotman, M.E. Dual role of alpha-defensin-1 in anti-HIV-1 innate immunity. J. Clin. Invest. 2005, 115, 765–773. [Google Scholar]
- Yasin, B.; Wang, W.; Pang, M.; Cheshenko, N.; Hong, T.; Waring, A.J.; Herold, B.C.; Wagar, E.A.; Lehrer, R.I. Theta defensins protect cells from infection by herpes simplex virus by inhibiting viral adhesion and entry. J. Virol. 2004, 78, 5147–5156. [Google Scholar]
- McDermott, A.M. Defensins and other antimicrobial peptides at the ocular surface. Ocul. Surf. 2004, 2, 229–247. [Google Scholar]
- Valore, E.V.; Ganz, T. Posttranslational processing of defensins in immature human myeloid cells. Blood 1992, 79, 1538–1544. [Google Scholar]
- Schröder, J.M.; Harder, J. Human β-defensin-2. Int. J. Cell Biol. 1999, 31, 645–651. [Google Scholar] [CrossRef]
- García, J.R.; Krause, A.; Schulz, S.; Rodríguez-Jiménez, F.J.; Klüver, E.; Adermann, K.; Forssmann, U.; Frimpong-Boateng, A.; Bals, R.; Forssmann, W.G. Human beta-defensin 4: A novel inducible peptide with a specific salt-sensitive spectrum of antimicrobial activity. FASEB J. 2001, 15, 1819–1821. [Google Scholar]
- Feng, Z.; Jiang, B.; Chandra, J.; Ghannoum, M.; Nelson, S.; Weinberg, A. Human beta-defensins: Differential activity against Candidal species and regulation by Candida albicans. J. Dent. Res. 2005, 84, 445–450. [Google Scholar]
- Joly, S.; Organ, C.C.; Johnson, G.K.; McCray, P.B.; Guthmiller, J.M. Correlation between beta-defensin expression and induction profiles in gingival keratinocytes. Mol. Immunol. 2005, 42, 1073–1084. [Google Scholar]
- Quiñones-Mateu, M.E.; Lederman, M.M.; Feng, Z.; Chakraborty, B.; Weber, J.; Rangel, H.R.; Marotta, M.L.; Mirza, M.; Jiang, B.; Kiser, P.; Medvik, K.; Sieg, S.F.; Weinberg, A. Human epithelial beta-defensins 2 and 3 inhibit HIV-1 replication. AIDS 2003, 17, F39–F48. [Google Scholar]
- Sun, L.; Finnegan, C.M.; Kish-Catalone, T.; Blumenthal, R.; Garzino-Demo, P.; La Terra Maggiore, G.M.; Berrone, S.; Kleinman, C.; Wu, Z.; Abdelwahab, S.; Lu, W.; Garzino-Demo, A. Human beta-defensins suppress human immunodeficiency virus infection: Potential role in mucosal protection. J. Virol. 2005, 79, 14318–14329. [Google Scholar]
- Steinberg, D.A.; Hurst, M.A.; Fujii, C.A.; Kung, A.H.; Ho, J.F.; Cheng, C.; Loury, D.J.; Fiddess, J.C. Protegrin-1: A broad-spectrum, rapidly microbicidal peptide with in vivo activity. Antimicrob. Agents Chemother. 1997, 41, 1738–1742. [Google Scholar]
- Ge, Y.D.; MacDonald, D.L.; Holroyd, K.J.; Thornsberry, C.; Wexler, H.; Zasloff, M. In vitro antibacterial properties of pexiganan, an analog of magainin. Antimicrob. Agents Chemother. 1999, 43, 782–788. [Google Scholar]
- Zhang, L.; Parente, J.; Harris, S.M.; Woods, D.E.; Hancock, E.W.; Falla, T.J. Antimicrobial peptide therapeutics for cystic fibrosis. Antimicrob. Agents Chemother. 2005, 49, 2921–2927. [Google Scholar]
- Greenwald, G.I.; Ganz, T. Defensins mediate the microbicidal activity of human neutrophil granule extract against Acinetobacter calcoaceticus. Infect. Immun. 1987, 55, 1365–1368. [Google Scholar]
- Harder, J.; Meyer-Hoffert, U.; Teran, L.M.; Schwichtenberg, L.; Bartels, J.; Maune, S.; Schröder, J.M. Mucoid Pseudomonas aeroginosa, TNF-alpha, and IL-1beta, but not IL-6, induce human beta-defensin-2 in respiratory epithelia. Am. J. Respir. Cell Mol. Biol. 2000, 22, 714–721. [Google Scholar]
- Jia, H.P.; Schutte, B.C.; Schudy, A.; Linzmeier, R.; Guthmiller, J.M.; Johnson, G.K.; Tack, B.F.; Mitros, J.P.; Rosenthal, A.; Ganz, T. Discovery of new human b-defensins using a genomics-based approach. Gene 2001, 263, 211–218. [Google Scholar] [CrossRef]
- Duits, L.A.; Ravensbergen, B.; Rademaker, M.; Hiemstra, P.S.; Nibbering, P.H. Expression of beta-defensin 1 and 2 mRNA by human monocytes, macrophages and dendritic cells. Immunology 2002, 106, 517–525. [Google Scholar]
- Sumikawa, Y.; Asada, H.; Hoshino, K.; Azukizawa, H.; Katayama, I.; Akira, S.; Itami, S. Induction of beta-defensin 3 in keratinocytes stimulated by bacterial lipopeptides through toll-like receptor 2. Microbes Infect. 2006, 8, 1513–1521. [Google Scholar] [CrossRef]
- Chalifour, A.; Jeannin, P.; Gauchat, J.F.; Blaecke, A.; Malissard, M.; N’Guyen, T.; Thieblemont, N.; Delneste, Y. Direct bacterial protein PAMP recognition by human NK cells involves TLRs and triggers alpha-defensin production. Blood 2004, 104, 1778–1783. [Google Scholar]
- Wehkamp, J.; Harder, J.; Weichenthal, M.; Schwab, M.; Schäffeler, E.; Schlee, M.; Herrlinger, K.R.; Stallmach, A.; Noack, F.; Fritz, P.; Schröder, J.M.; Bevins, C.L.; Fellermann, K.; Stange, E.F. NOD2 (CARD15) mutations in Crohn’s disease are associated with diminished mucosal alpha-defensin expression. Gut 2004, 53, 1658–1664. [Google Scholar]
- Lu, Q.; Darveau, R.P.; Samaranayake, L.P.; Wang, C.Y.; Jin, L. Differential modulation of human beta-defensins expression in human gingival epithelia by Porphyromonas gingivalis lipopolysaccharide with tetra- and penta-acylated lipid A structures. Innate Immun. 2009, 15, 325–335. [Google Scholar] [CrossRef]
- Varoga, D.; Wruck, C.J.; Tohidnezhad, M.; Brandenburg, L.; Paulsen, F.; Mentlein, R.; Seekamp, A.; Besch, L.; Pufe, T. Osteoblasts participate in the innate immunity of the bone by producing human beta defensin-3. Histochem. Cell Biol. 2009, 131, 207–218. [Google Scholar] [CrossRef]
- Scott, M.G.; Vreugdenhil, A.C.; Buurman, W.A.; Hancock, R.E.; Gold, M.R. Cutting edge: Cationic antimicrobial peptides block the binding of lipopolysaccharide (LPS) to LPS binding protein. J. Immunol. 2000, 164, 549–553. [Google Scholar]
- Miles, K.; Clarke, D.J.; Lu, W.; Sibinska, Z.; Beaumont, P.E.; Davidson, D.J.; Barr, T.A.; Campopiano, D.J.; Gray, M. Dying and necrotic neutrophils are anti-inflammatory secondary to the release of alpha-defensins. J. Immunol. 2009, 183, 2122–2132. [Google Scholar]
- Soehnlein, O.; Kai-Larsen, Y.; Frithiof, R.; Sorensen, O.E.; Kenne, E.; Scharffetter-Kochanek, K.; Eriksson, E.E.; Herwald, H.; Agerberth, B.; Lindblom, L. Neutrophil primary granule proteins HBP and HNP1-3 boost bacterial phagocytosis by human and murine macrophages. J. Clin. Invest. 2008, 118, 3491–3502. [Google Scholar] [CrossRef]
- Nagaoka, I.; Niyonsaba, F.; Tsutsumi-Ishii, Y.; Tamura, H.; Hirata, M. Evaluation of the effect of human beta-defensins on neutrophil apoptosis. Int. Immunol. 2008, 20, 543–553. [Google Scholar]
- Oppenheim, J.J.; Yang, D. Alarmins: Chemotatctic activators of immune response. Curr. Opin. Immunol. 2005, 17, 359–365. [Google Scholar]
- Niyonsaba, F.; Iwabuchi, K.; Matsuda, H.; Ogawa, H.; Nagaoka, I. Epithelial cell-derived human beta-defensin-2 acts as a chemotaxin for mast cells through a pertussis toxin-sensitive and phospholipase C-dependent pathway. Int. Immunol. 2002, 14, 421–426. [Google Scholar] [CrossRef]
- Niyonsaba, F.; Ogawa, H.; Nagaoka, I. Human beta-defensin-2 functions as a chemotactic agent for tumor necrosis factor-alpha-treated human neutrophils. Immunology 2004, 111, 273–281. [Google Scholar] [CrossRef]
- Territo, M.C.; Ganz, T.; Selsted, M.E.; Lehrer, R.I. Monocyte-chemotactic activity of defensins from human neutrophils. J. Clin. Invest. 1989, 84, 2017–2020. [Google Scholar]
- Yang, D.; Chen, Q.; Chertov, O.; Oppenheim, J.J. Human neutrophil defensins selectively chemoattract naive T and immature dendritic cells. J. Leukoc. Biol. 2000, 68, 9–14. [Google Scholar]
- Yang, D.; Chertov, O.; Oppenheim, J.J. Participation of mammalian defensins and cathelicidins in anti-microbial immunity: Receptors and activities of human defensins and cathelicidin (LL-37). J. Leukoc. Biol. 2001, 69, 691–697. [Google Scholar]
- Rohrl, J.; Yand, D.; Oppenheim, J.J.; Hehlgans, T. Human β-defensins 2 and 3 and their mouse orthologs induce chemotaxis through interaction with CCR2. J. Immunol. 2010, 184, 6688–6694. [Google Scholar] [CrossRef]
- Funderburg, N.; Lederman, M.M.; Feng, Z.; Drage, M.G.; Jadlowsky, J.; Harding, C.V.; Weinberg, A.; Sieg, S.F. Human -defensin-3 activates professional antigen-presenting cells via Toll-like receptors 1 and 2. Proc. Natl. Acad. Sci. USA 2007, 104, 18631–18635. [Google Scholar]
- Bals, R.; Weiner, D.J.; Meegalla, R.L.; Wilson, J.M. Transfer of a cathelicidin peptide antibiotic gene restores bacterial killing in a cystic fibrosis xenograft model J. Clin. Invest. 1999, 103, 1113–1117. [Google Scholar] [CrossRef]
- Jacobsen, F.; Mittler, D.; Hirsch, T.; Gerhards, A.; Lehnhardt, M.; Voss, B.; Steinau, H.U.; Steinstraesser, L. Transient cutaneous adenoviral gene therapy with human host defense peptide hCAP-18/LL-37 is effective fort the treatment of burn wound infections. Gene Ther. 2005, 12, 1494–1502. [Google Scholar]
- Lai, Y.; Gallo, R.L. AMPed up immunity: How antimicrobial peptides have multiple roles in immune defense. Trends Immunol. 2009, 30, 131–141. [Google Scholar] [CrossRef]
- Han, W.; Wang, W.; Mohammed, K.A.; Su, Y. Defensins increase lung fibroblast proliferation and collagen synthesis via β-catenin signaling pathway. FEBS J. 2009, 276, 6603–6614. [Google Scholar] [CrossRef]
- Baroni, A.; Donnarumma, G.; Paoletti, I.; Longanesi-Cattani, I.; Bifulco, K.; Tufano, M.A.; Carriero, M.V. Antimicrobial human beta-defensin-2 stimulates migration, proliferation and tube formation of human umbilical vein endothelial cells. Peptides 2009, 30, 267–272. [Google Scholar]
- Bick, R.J.; Poindexter, B.J.; Buja, L.M.; Lawyer, C.H.; Milner, S.M.; Bhat, S. Nuclear lokalization of hBD-1 in human keratinocytes. J. Burn Wounds 2007, 7, 25–32. [Google Scholar]
- Prado-Montes de Orca, E. Human β-defensin 1: A restless warrior against allergies, infections and cancer. Int. J. Biochem. Cell Biol. 2010, 42, 800–804. [Google Scholar] [CrossRef]
- St John, M.A.; Li, Y.; Zhou, X.; Denny, P.; Ho, C.M.; Montemagno, C.; Shi, W.; Qi, F.; Wu, B.; Sinha, U.; Jordan, R.; Wolinsky, L.; Park, N.H.; Liu, H.; Abemayor, E.; Wong, D.T. Interleukin 6 and interleukin 8 as potential biomarkers for oral cavity and oropharyngeal squamous cell carcinoma. Arch. Otolaryngol. Head Neck Surg. 2004, 130, 929–935. [Google Scholar]
- Michiels, K.; Schutyser, E.; Conings, R.; Lenaerts, J.P.; Put, W.; Nuyts, S.; Delaere, P.; Jacobs, R.; Struyf, S.; Proost, P.; van Damme, J. Carcinoma cell-derived chemokines and their presence in oral fluid. Eur. J. Oral Sci. 2009, 117, 362–368. [Google Scholar] [CrossRef]
- Pitiyage, G.; Tilakaratne, W.M.; Tavassoli, M.; Warnakulasuriya, S. Molecular markers in oral epithelial dysplasia: Review. J. Oral Pathol. Med. 2009, 38, 737–752. [Google Scholar] [CrossRef]
- Porta, C.; Larghi, P.; Rimoldi, M.; Totaro, M.G.; Allavena, P.; Mantovani, A.; Sica, A. Cellular and molecular pathways linking inflammation and cancer. Immunobiology 2009, 214, 761–777. [Google Scholar] [CrossRef]
- Sica, A.; Allavena, P.; Mantovani, A. Cancer related inflammation: The macrophage connection. Cancer Lett. 2008, 267, 204–215. [Google Scholar] [CrossRef]
- Tezal, M.; Sullivan, M.A.; Reid, M.E.; Marshall, J.R.; Hyland, A.; Loree, T.; Lillis, C.; Hauck, L.; Wactawski-Wende, J.; Scannapieco, F.A. Chronic periodontitis and the risk of tongue cancer. Arch. Otolaryngol. Head Neck Surg. 2007, 133, 450–454. [Google Scholar]
- Tezal, M.; Sullivan, M.A.; Hyland, A.; Marshall, J.R.; Stoler, D.; Reid, M.E.; Loree, T.R.; Rigual, N.R.; Merzianu, M.; Hauck, L.; Lillis, C.; Wactawski-Wende, J.; Scannapieco, F.A. Chronic periodontitis and the incidence of head and neck squamous cell carcinoma. Cancer Epidemiol. Biomarkers Prev. 2009, 18, 2406–2412. [Google Scholar]
- Donald, C.D.; Sun, C.Q.; Lim, S.D.; Macoska, J.; Cohen, C.; Amin, M.B.; Young, A.N.; Ganz, T.A.; Marshall, F.F.; Petros, J.A. Cancer-specific loss of beta-defensin 1 in renal and prostatic carcinomas. Lab. Invest. 2003, 83, 501–505. [Google Scholar]
- Sun, C.Q.; Arnold, R.; Fernandez-Golarz, C.; Parrish, A.B.; Almekinder, T.; He, J.; Ho, S.M.; Svoboda, P.; Pohl, J.; Marshall, F.F.; Petros, J.A. Human beta-defensin-1, a potential chromosome 8p tumor suppressor: Control of transcription and induction of apoptosis in renal cell carcinoma. Cancer Res. 2006, 66, 8542–8549. [Google Scholar]
- Joly, S.; Compton, L.M.; Pujol, C.; Kurago, Z.B.; Guthmiller, J.M. Loss of human beta-defensin 1, 2, and 3 expression in oral squamous cell carcinoma. Oral Microbiol. Immunol. 2009, 24, 353–360. [Google Scholar] [CrossRef]
- Wenghoefer, M.; Pantelis, A.; Dommisch, H.; Reich, R.; Martini, M.; Allam, J.P.; Novak, N.; Bergé, S.; Jepsen, S.; Winter, J. Decreased gene expression of human beta-defensin-1 in the development of squamous cell carcinoma of the oral cavity. Int. J. Oral Maxillofac. Surg. 2008, 37, 660–663. [Google Scholar] [CrossRef]
- Kesting, M.R.; Loeffelbein, D.J.; Hasler, R.J.; Wolff, K.D.; Rittig, A.; Schulte, M.; Hirsch, T.; Wagenpfeil, S.; Jacobsen, F.; Steinstraesser, L. Expression profile of human beta-defensin 3 in oral squamous cell carcinoma. Cancer Invest. 2009, 27, 575–581. [Google Scholar] [CrossRef]
- Wenghoefer, M.; Pantelis, A.; Najafi, T.; Deschner, J.; Allam, J.P.; Novak, N.; Reich, R.; Martini, M.; Bergé, S.; Fischer, HP.; Jepsen, S.; Winter, J. Gene expression of oncogenes, antimicrobial peptides, and cytokines in the development of oral leukoplakia. Oral Surg. Oral Med. Oral Pathol. Oral Radiol. Endod. 2010, 110, 351–356. [Google Scholar]
- Winter, J.; Pantelis, A.; Allam, J.P.; Novak, N.; Reich, R.; Martini, M.; Bergé, S.; Deschner, J.; Jepsen, S.; Wenghoefer, M. High α-defensin and S100A7 expression and missing DOC-1 down-regulation characterize irritation fibromas of the oral cavity and may counteract malignant transformation. J. Craniofac. Surg. 2011, 22, 100–104. [Google Scholar]
- Schaefer, A.S.; Richter, G.M.; Nothnagel, M.; Laine, M.L.; Rühling, A.; Schäfer, C.; Cordes, N.; Noack, B.; Folwaczny, M.; Glas, J.; Dörfer, C.; Dommisch, H.; Groessner-Schreiber, B.; Jepsen, S.; Loos, B.G.; Schreiber, S. A 3' UTR transition within DEFB1 is associated with chronic and aggressive periodontitis. Genes Immun. 2010, 11, 45–54. [Google Scholar]
- Lichtenstein, A.; Ganz, T.; Selsted, M.E.; Lehrer, R.I. In vitro tumor cell cytolysis mediated by peptide defensins of human and rabbit granulocytes. Blood 1986, 68, 1407–1410. [Google Scholar]
- Lapteva, N.; Aldrich, M.; Rollins, L.; Ren, W.; Goltsova, T.; Chen, S.Y.; Huang, X.F. Attraction and activation of dendritic cells at the site of tumor elicits potent antitumor immunity. Mol. Ther. 2009, 17, 1626–1636. [Google Scholar]
- Escher, N.; Spies-Weisshart, B.; Kaatz, M.; Melle, C.; Bleul, A.; Driesch, D.; Wollina, U.; von Eggeling, F. Identification of HNP3 as a tumor marker in CD4+ and CD4- lymphocytes of patients with cutaneous cell lymphoma. Eur. J. Cancer 2006, 42, 249–255. [Google Scholar] [CrossRef]
- Radeva, M.Y.; Jahns, F.; Wilhelm, A.; Glei, M.; Settmacher, U.; Greulich, K.O.; Mothes, H. Defensins alpha 6 (DEFA 6) overexpression threshold of over 60 fold can distinguish between adenoma and fully blown colon carcinoma in individual patients. BMC Cancer 2010, 10. [Google Scholar]
- Zhang, L.; Parente, J.; Harris, S.M.; Woods, D.E.; Hancock, R.E.W.; Falla, T.J. Antimicrobial peptide therapeutics for cystic fibrosis. Antimicrob. Agents Chemother 2005, 49, 2921–2927. [Google Scholar]
- Winder, D.; Gunzburg, W.H.; Erfle, V.; Salmons, B. Expression of antimicrobial peptides has an antitumour effect in human cells. Biochem. Biophys. Res. Commun. 1998, 242, 608–612. [Google Scholar] [CrossRef]
- Papo, N.; Shahar, M.; Eisenbach, L.; Shai, Y. A novel lytic peptide composed of D, L amino acids selectively kills cancer cells in culture and in mice. J. Biol. Chem. 2003, 278, 21018–21023. [Google Scholar]
- Chen, Y.; Xu, X.; Hong, S.; Chen, J.; Liub, N.; Underhill, C.B.; Creswell, K.; Zhang, L. RGD-tachyplesin inhibits tumor growth. Cancer Res. 2001, 61, 2434–2438. [Google Scholar]
- Papo, N.; Shai, Y. Host defense peptides as new weapons in cancer treatment. Cell. Mol. Life Sci. 2005, 62, 784–790. [Google Scholar] [CrossRef]
- Jenssen, H.; Hamill, P.; Hancock, R.E.W. Peptide antimicrobial agents. Clin. Microbiol. Rev. 2006, 19, 491–511. [Google Scholar] [CrossRef]
- Hancock, R.E.W.; Sahl, H.G. Antimicrobial and host defense peptides as new anti-infective therapeutic strategies. Nat. Biotechnol. 2006, 24, 1551–1557. [Google Scholar] [CrossRef]
- Rotem, S.; Mor, A. Antimicobial peptide mimics for improved therapeutic properties. Biochim. Biophys. Acta 1788, 1582–1592. [Google Scholar]
- Findlay, B.; Zhanel, G.G.; Schweizer, F. Cationic amphiphiles, a new generation of antimicrobials inspired by the natural antimicrobial peptide scaffold. Antimicrob. Agents Chemother 2010, 54, 4049–4058. [Google Scholar] [CrossRef]
- Yeung, A.T.Y.; Gellatly, S.L.; Hancock, R.E.W. Multifunctional cationic host defense peptides and their clinical applications. Cell. Mol. Life Sci. 2011, 68, 2161–2176. [Google Scholar]
© 2012 by the authors; licensee MDPI, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
MDPI and ACS Style
Winter, J.; Wenghoefer, M. Human Defensins: Potential Tools for Clinical Applications. Polymers 2012, 4, 691-709. https://doi.org/10.3390/polym4010691
AMA Style
Winter J, Wenghoefer M. Human Defensins: Potential Tools for Clinical Applications. Polymers. 2012; 4(1):691-709. https://doi.org/10.3390/polym4010691
Chicago/Turabian StyleWinter, Jochen, and Matthias Wenghoefer. 2012. "Human Defensins: Potential Tools for Clinical Applications" Polymers 4, no. 1: 691-709. https://doi.org/10.3390/polym4010691