
Impact of Grass Cover Management with Herbicides on Biodiversity, Soil Cover and Humidity in Olive Groves in the Southern Iberian

 ,
,  , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
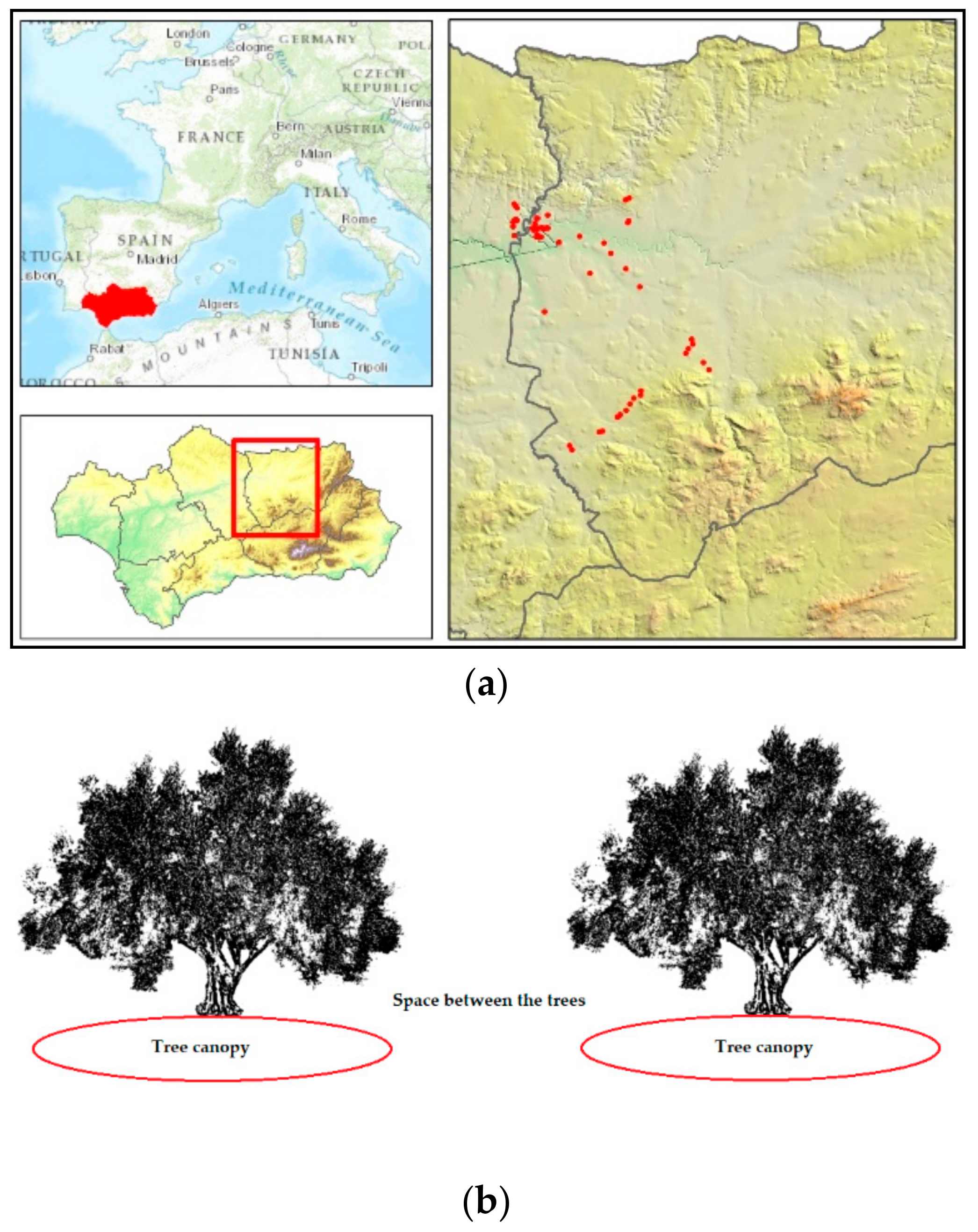
2.1. Study Area
2.2. Sampling and Resampling Points
- —
- Immediately under the olive tree canopy
- —
- In the middle of the road (outside the canopy)
- —
- On the fringe of the olive grove
- —
- Dehesa
2.3. Climatic and Bioclimatic Data
- T: Mean annual temperature in degrees centigrade
- M: Mean temperature of the maximums
- m: Mean temperature of the minimums
- Tmax: Mean temperature of the warmest month of the year
- Tmin: Mean temperature of the coldest month of the year
- Tp: Positive annual temperature (sum of the months with a mean temperature above 0 °C in tenths of degrees centigrade)
- Pp: Positive annual precipitation (of the months in Ti above 0 °C)
- PE: Thornthwaite’s potential annual evapotranspiration index [33]
- PEs: Potential evapotranspiration index of the summer quarter
- PE1–12: Potential monthly evapotranspiration index 1 = January, 2 = February, … 12 = December
2.4. Soil Data and Moisture
2.5. Determination of the Flora and Delimitation of the Plant Communities
2.6. Data Analysis
3. Results
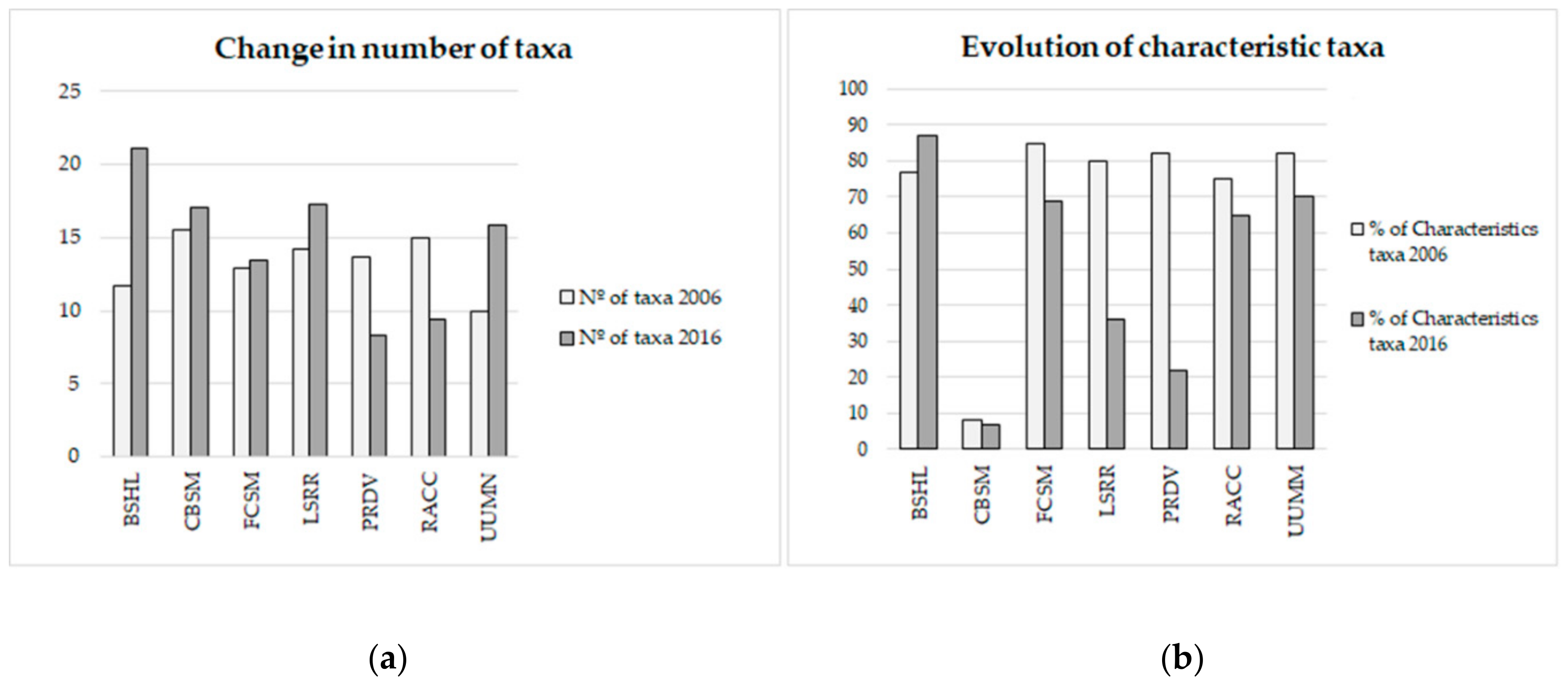
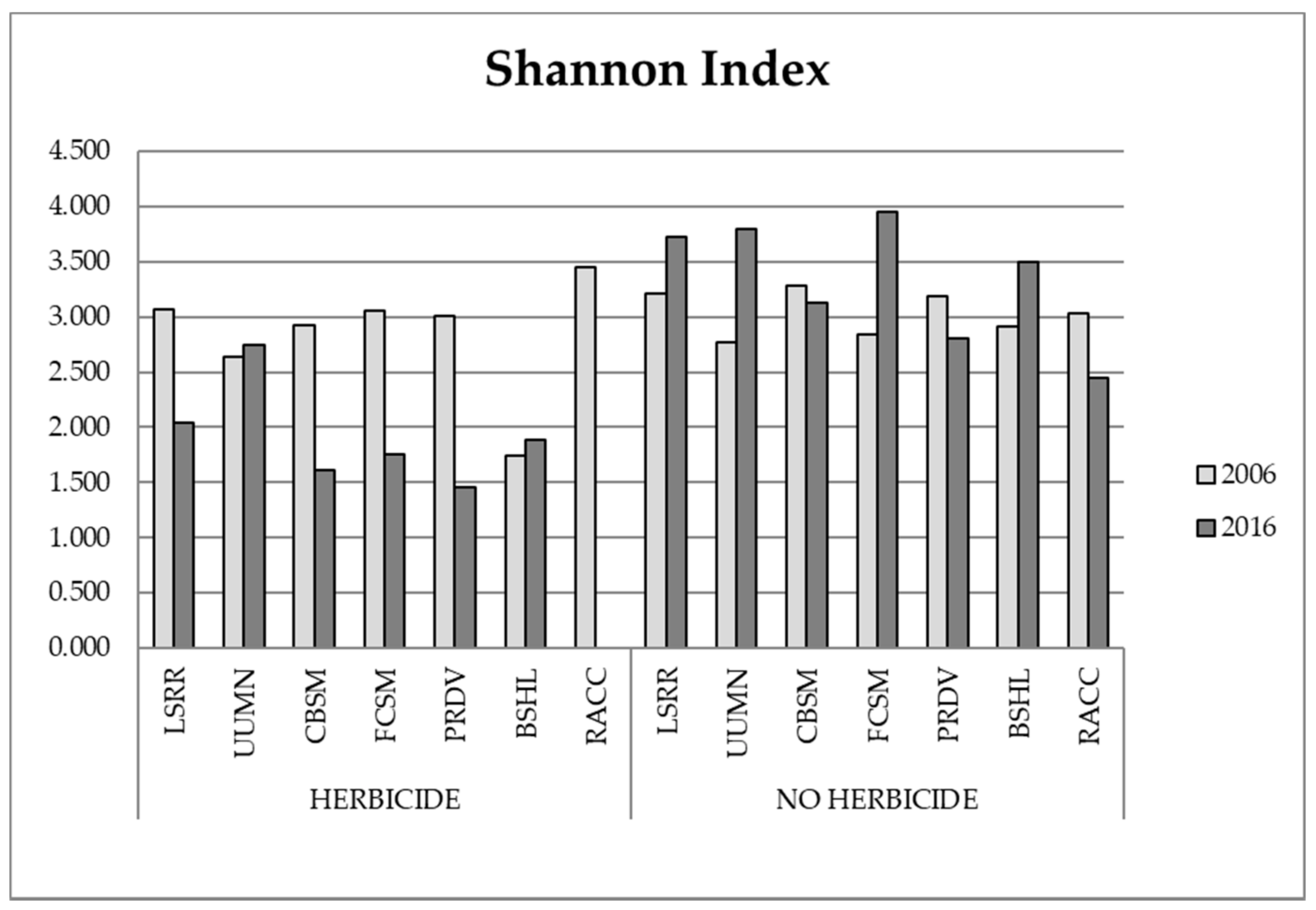
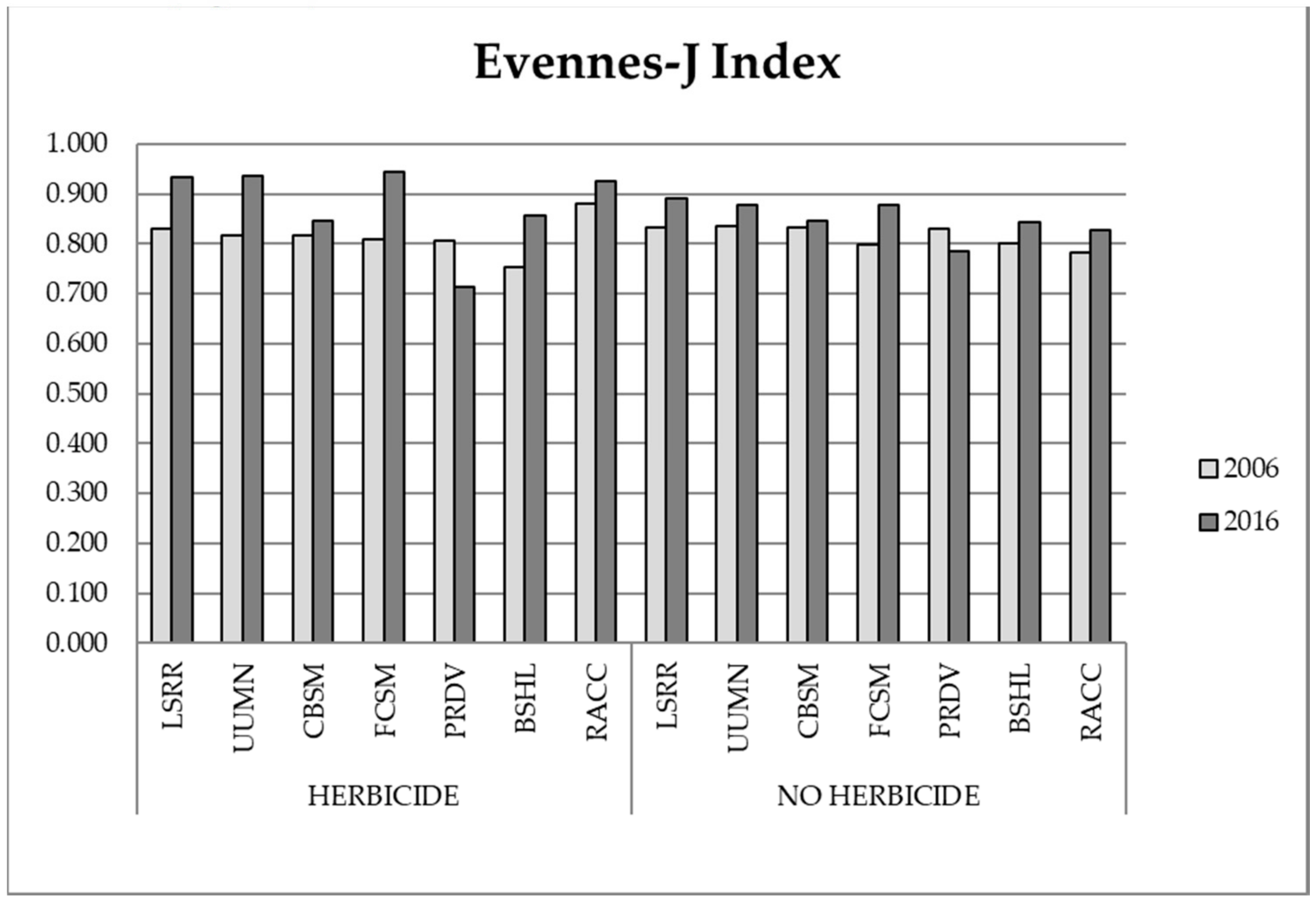
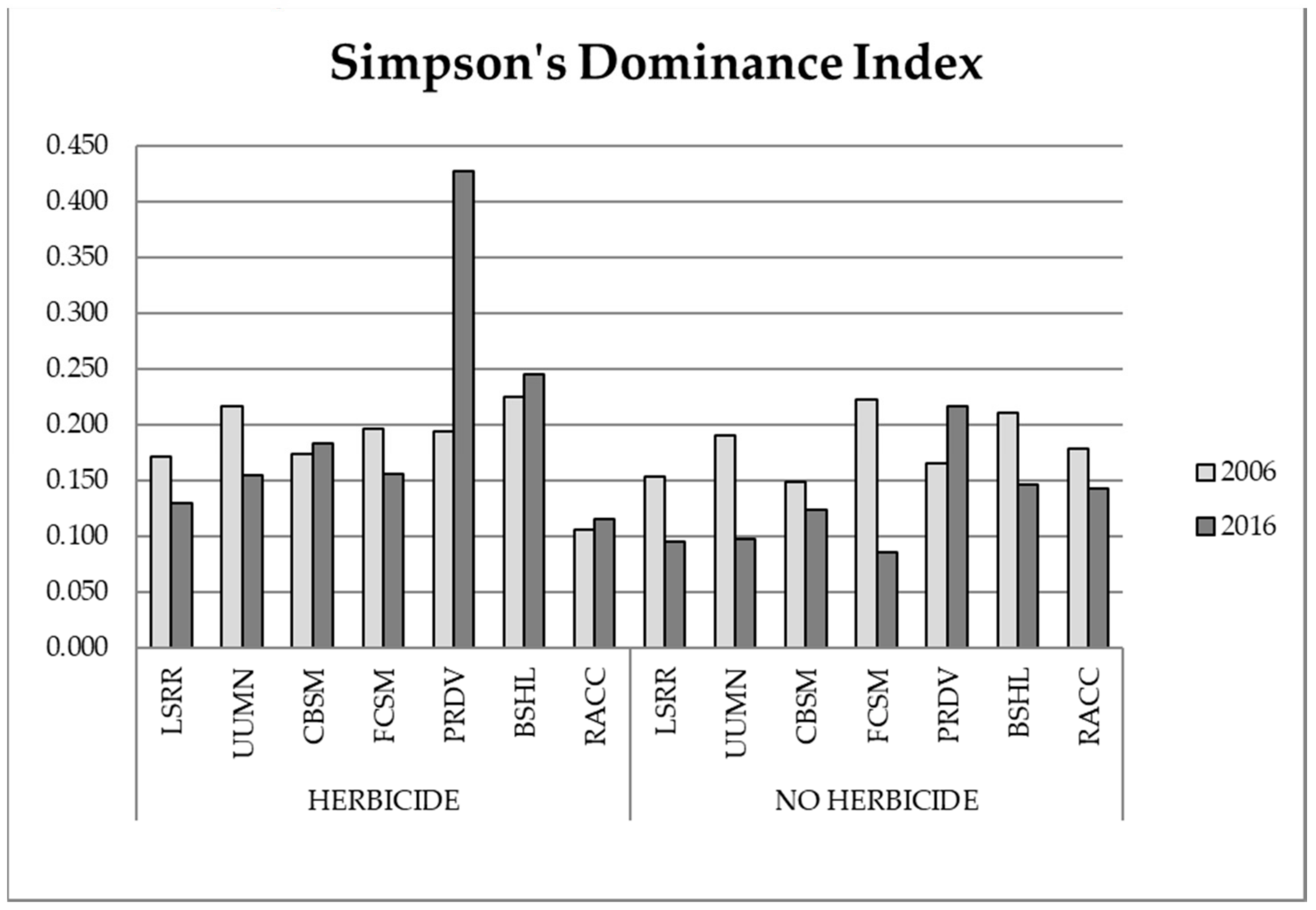
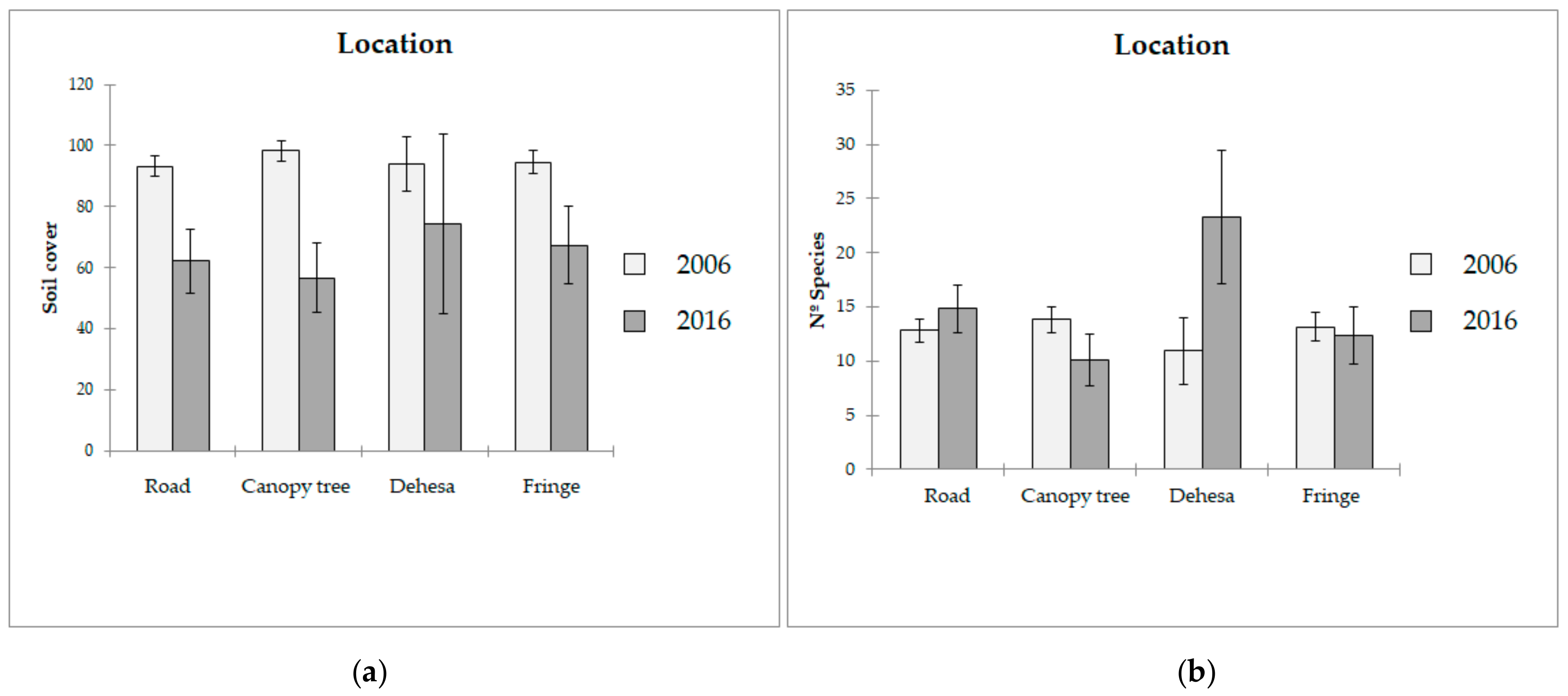
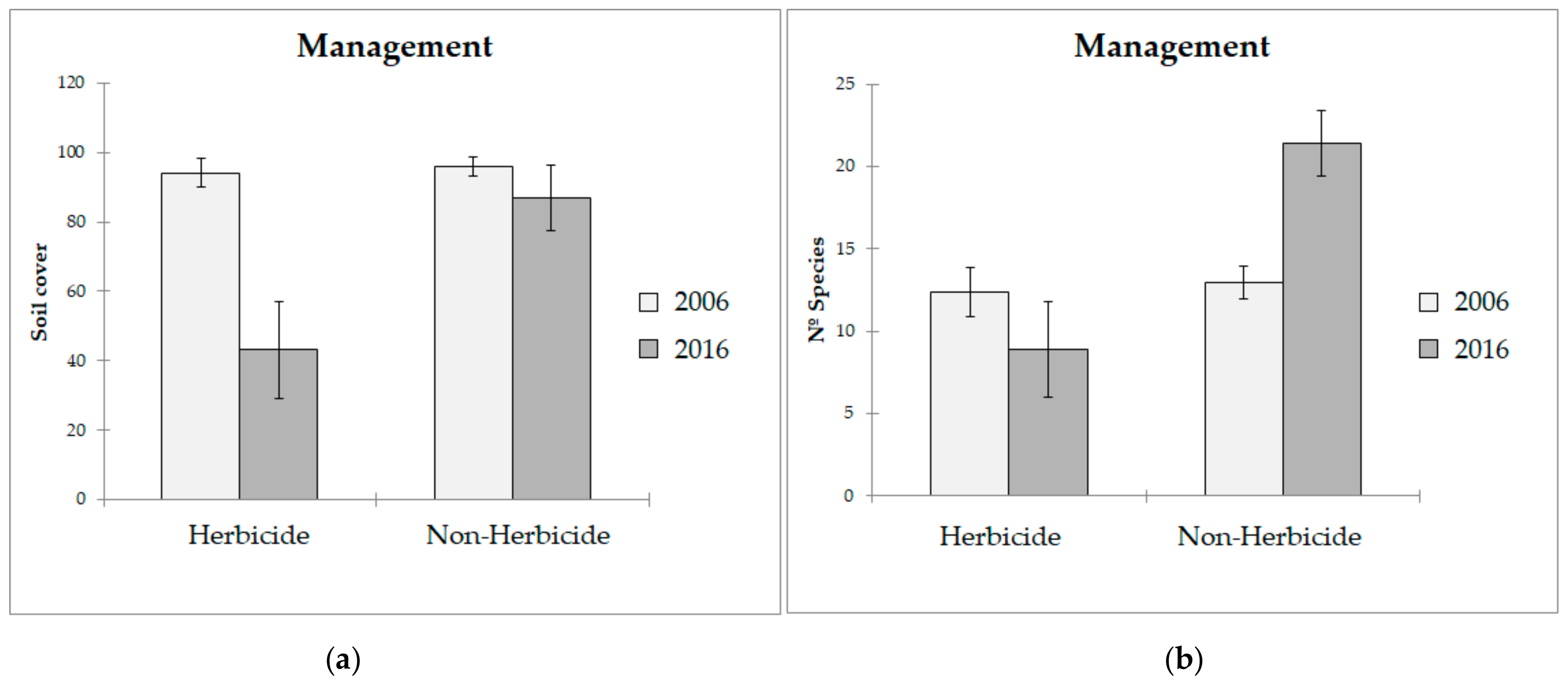
3.1. Biodiversity Analysis
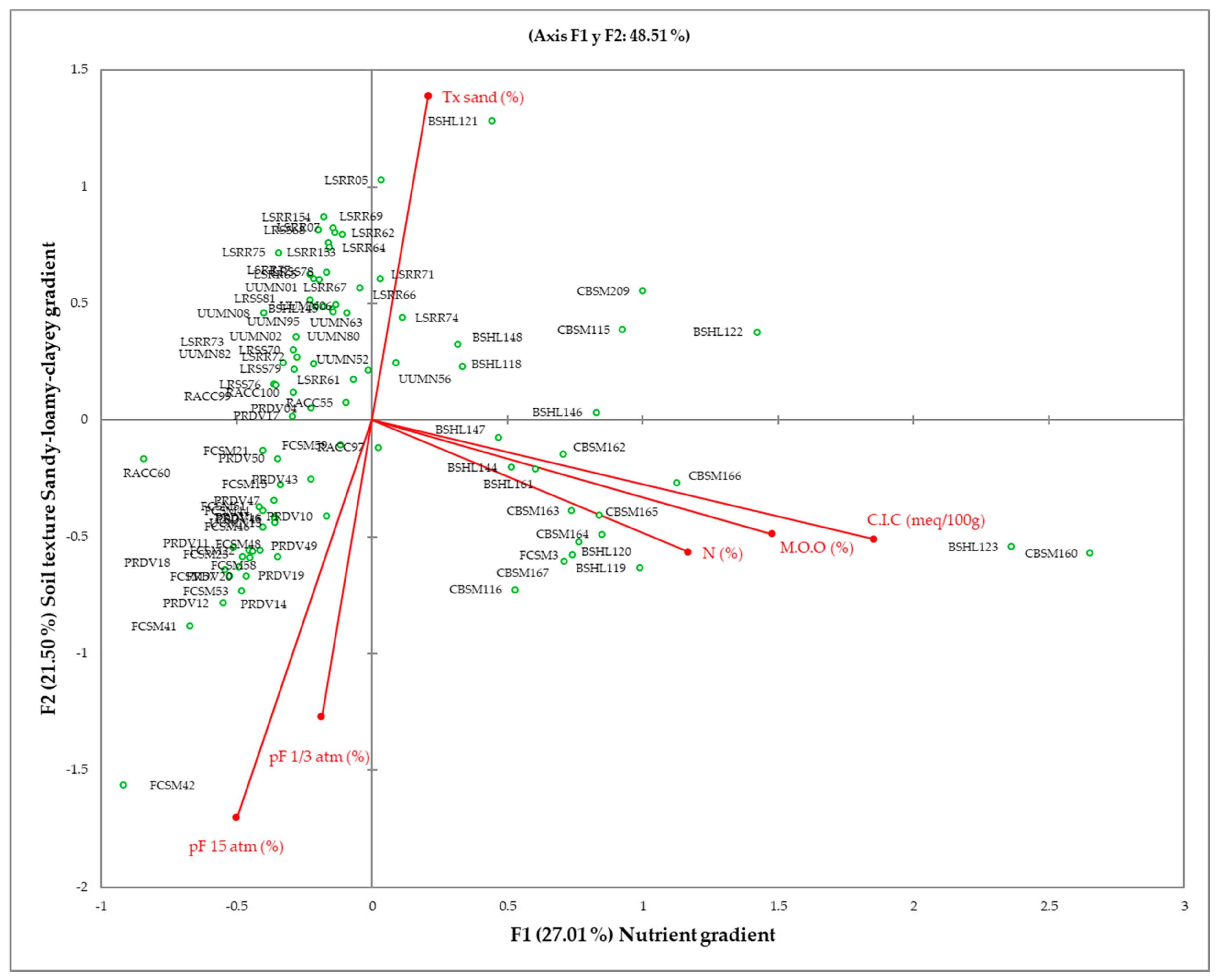
3.2. Soil and Bioclimatic Characterization of Plant Communities
3.3. Soil and Bioclimatic Characterization of Plant Communities
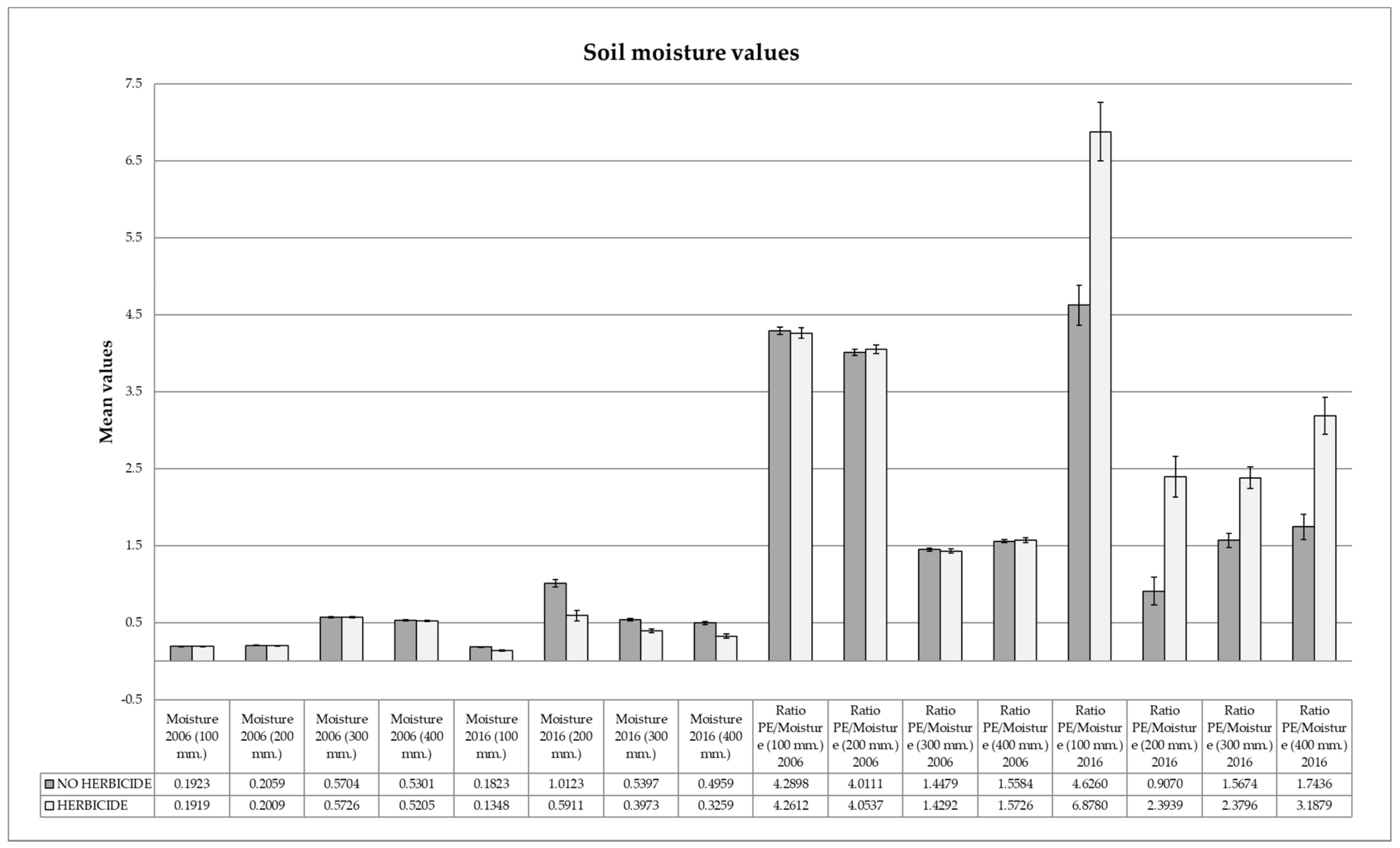
3.4. Soil Moisture Analysis
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Giorgi, F.; Lionello, P. Climate change projections for the Mediterranean region. Glob. Planet. Chang. 2008, 63, 90–104. [Google Scholar] [CrossRef]
- Tullot, I.F. Climatología de España y Portugal; Universidad de Salamanca: Salamanca, Spain, 2000; pp. 1–428. [Google Scholar]
- Navarro-Serrano, F.; López-Moreno, J.I.; Azorin-Molina, C.; Alonso-González, E.; Aznarez-Balta, M.; Buisán, S.T.; Revuelto, J. Elevation Effects on Air Temperature in a Topographically Complex Mountain Valley in the Spanish Pyrenees. Atmosphere 2020, 11, 656. [Google Scholar] [CrossRef]
- Cano, E.; Cano-Ortiz, A.; Musarella, C.M.; Fuentes, J.P.; Ighbareyeh, J.M.; Gea, F.L.; del Río, S. Mitigating Climate Change through Bioclimatic Applications and Cultivation Techniques in Agriculture (An-dalusia, Spain). In Sustainable Agriculture, Forest and Environmental Management; Jhariya, M., Banerjee, A., Meena, R., Yadav, D., Eds.; Springer: Singapore, 2019. [Google Scholar]
- Piñar Fuentes, J.C. Recent rainfalls trends in Andalusia (southern Spain). In Conservation Studies on Mediterranean Threatened Flora and Vegetation, Proceedings of the X International Meeting Biodiversity Conservation and Management, Sardinia, Italy, 13–17 June 2016; Bacchetta, G., Ed.; Sardinia University of Cagliari: Cagliari, Italy, 2016. [Google Scholar]
- Del Río, S.; Herrero, L.; Fraile, R.; Penas, A. Spatial distribution of recent rainfall trends in Spain (1961–2006). Int. J. Clim. 2011, 31, 656–667. [Google Scholar] [CrossRef]
- Aceite de Olive—Aceite de Oliva y Aceituna de Mesa-Producciones Agrícolas—Agricultura—Magrama.es. Available online: www.magrama.gob.esesagriculturatemasprodtucciones-agricolasaceite-oliva-y-aceituna-mesaaceite.aspx (accessed on 15 April 2019).
- Papadopoulou, M.P.; Charchousi, D.; Spanoudaki, K.; Karali, A.; Varotsos, K.V.; Giannakopoulos, C.; Markou, M.; Loizidou, M. Agricultural Water Vulnerability under Climate Change in Cyprus. Atmosphere 2020, 11, 648. [Google Scholar] [CrossRef]
- Perrino, E.V.; Musarella, C.M.; Magazzini, P. Management of grazing Italian river buffalo to preserve habitats defined by Directive 92/43/EEC in a protected wetland area on the Mediterranean coast: Palude Frattarolo, Apulia, Italy. Euro-Mediterr. J. Environ. Integr. 2021, 6, 1–18. [Google Scholar] [CrossRef]
- Vanwalleghem, T.; Amate, J.I.; De Molina, M.G.; Fernández, D.S.; Gómez, J.A. Quantifying the effect of historical soil management on soil erosion rates in Mediterranean olive orchards. Agric. Ecosyst. Environ. 2011, 142, 341–351. [Google Scholar] [CrossRef]
- Pimentel, D. Soil Erosion: A Food and Environmental Threat. Environ. Dev. Sustain. 2006, 8, 119–137. [Google Scholar] [CrossRef]
- Mielniczuk, J.; Schneider, P. Aspectos Sócio-Econômicos do Manejo de Solos no sul do Brasil. In Anais do I Simpósio de Manejo de Solo e Plantio Direto no Sul do Brasil e II Simpósio de Conservaçao de Solo do Planalto; Universidade de Passo Fundo: Passo Fundo, Brazil, 1984; pp. 3–19. [Google Scholar]
- Chang, F.-J.; Guo, S. Advances in Hydrologic Forecasts and Water Resources Management. Water 2020, 12, 1819. [Google Scholar] [CrossRef]
- Lv, J.; Xie, Y.; Luo, H. Erosion Process and Temporal Variations in the Soil Surface Roughness of Spoil Heaps under Mul-ti-Day Rainfall Simulation. Remote Sens. 2020, 12, 2192. [Google Scholar] [CrossRef]
- Pastor Muñoz-Cobo, M. Estudio de los Diversos Métodos de Manejo de Suelo Alternativos al Laboreo en el Cultivo del Olivo; Universidad de La Rioja: La Rioja, Spain, 1991; Volume 1, p. 302. [Google Scholar]
- Pimentel, D.; Harvey, C.; Resosudarmo, P.; Sinclair, K.; Kurz, D.; McNair, M.; Crist, S.; Shpritz, L.; Fitton, L.; Saffouri, R.; et al. Environmental and Economic Costs of Soil Erosion and Conservation Benefits. Science 1995, 267, 1117–1123. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- United Nations. Transforming Our World: The 2030 Agenda for Sustainable Development. 2015. A/RES/70/1. Available online: https://sustainabledevelopment.un.org/content/documents/21252030%20Agenda%20for%20Sustainable%20Development%20web.pdf (accessed on 15 January 2021).
- López-Felices, B.; Aznar-Sánchez, J.A.; Velasco-Muñoz, J.F.; Piquer-Rodríguez, M. Contribution of Irrigation Ponds to the Sustainability of Agriculture. A Review of Worldwide Research. Sustainability 2020, 12, 5425. [Google Scholar] [CrossRef]
- Caparrós-Martínez, J.L.; Milán-García, J.; Rueda-López, N.; de Pablo-Valenciano, J. Green Infrastructure and Water: An Analysis of Global Research. Water 2020, 12, 1760. [Google Scholar] [CrossRef]
- Scheres, B.; Schüttrumpf, H. Investigating the Erosion Resistance of Different Vegetated Surfaces for Ecological Enhancement of Sea Dikes. J. Mar. Sci. Eng. 2020, 8, 519. [Google Scholar] [CrossRef]
- Leiva, F.; Cano-Ortiz, A.; Musarella, C.; Piñar Fuentes, J.C.; Pinto-Gomes, C.; Cano, E. New method for increasing sustainable agricultural yield. Transylv. Rev. 2017, 24, 3638–3648. [Google Scholar]
- Perrino, E.V.; Wagensommer, R.P. Crop Wild Relatives (CWR) Priority in Italy: Distribution, Ecology, In Situ and Ex Situ Conservation and Expected Actions. Sustainability 2021, 13, 1682. [Google Scholar] [CrossRef]
- Naveh, Z.; Whittaker, R.H. Structural and floristic diversity of shrublands and woodlands in Northern Israel and other Mediterranean areas. Vegetatio 1980, 41, 171–190. [Google Scholar] [CrossRef]
- Marañón, T. Diversidad florística y heterogeneidad ambiental en una dehesa de Sierra Morena. Anal. Edaf Agrobiol. 1985, 44, 1183–1197. [Google Scholar]
- Marañón, T. Diversidad en comunidades de pasto mediterráneo: Modelos y mecanismos de coexistencia. Ecología 1991, 5, 149–157. [Google Scholar]
- Fuentes, J.C.P.; Cano-Ortiz, A.; Musarella, C.M.; Quinto-Canas, R.; Gomes, C.J.P.; Spampinato, G.; González, S.D.R.; Cano, E. Bioclimatology, Structure, and Conservation Perspectives of Quercus pyrenaica, Acer opalus subsp. granatensis, and Corylus avellana Deciduous Forests on Mediterranean Bioclimate in the South-Central Part of the Iberian Peninsula. Sustainability 2019, 11, 6500. [Google Scholar] [CrossRef] [Green Version]
- Cano-Ortiz, A.; Musarella, C.M.; Fuentes, J.C.P.; Gomes, C.J.P.; Quinto-Canas, R.; Del Río, S.; Cano, E. Indicative Value of the Dominant Plant Species for a Rapid Evaluation of the Nutritional Value of Soils. Agronomy 2020, 11, 1. [Google Scholar] [CrossRef]
- Powles, S.B.; Yu, Q. Evolution in Action: Plants Resistant to Herbicides. Annu. Rev. Plant Biol. 2010, 61, 317–347. [Google Scholar] [CrossRef] [Green Version]
- Rodríguez, J.C. Cubiertas vegetales en el olivar: Funciones, tipos y manejo. Vida Rural 2000, 113, 38–40. [Google Scholar]
- Hermosín, M.C.; Rodríguez-Linaza, L.C.; Cornejo, J.; Ordóñez-Fernández, R. Efecto de uso de agroquímicos en el olivar sobre la calidad de las aguas. Sostenibilidad Prod. Oliv. Andal. 2009, 127–160. [Google Scholar]
- Cano-Ortiz, A. Bioindicadores Ecológicos y Manejo de Cubiertas Vegetales Como Herramienta Para la Implantación de una Agricultura Sostenible. Ph.D. Thesis, Universidad de Jaén, Jaén, Spain, 2007. [Google Scholar]
- Rivas-Martínez, S.; Rivas-Saenz, S.; Penas, A. Worldwide bioclimatic classification system. Glob. Geobot. 2002, 1, 1–638. [Google Scholar]
- Thornthwaite, C.W. An Approach toward a Rational Classification of Climate. Geogr. Rev. 1948, 38, 55–94. [Google Scholar] [CrossRef]
- Castroviejo, S. Flora Ibérica. Real Jardín Botánico; CSIC: Madrid, Spain, 2005. [Google Scholar]
- Cabezudo, B.; Cueto, M.; Salazar, C.; Morales Torres, C. Flora Vascular de Andalucía Oriental; Junta de Andalucia, Blanca, G., Eds.; Consejeria de Medio Ambiente: Sevilla, Spain, 2009; Volume 4. [Google Scholar]
- Valdés, B.; Talavera, S.; Galiano, E.F. Flora Vascular de Andalucía Occidental; Ketres: Barcelona, Spain, 1987. [Google Scholar]
- Vázquez, F.M. Aproximación al conocimiento del género Taraxacum FH Wigg (Asteraceae) en Extremadura (España). Folia Bot. Extrem. 2014, 8, 5–35. [Google Scholar]
- León, E.; Nieto, E.L.; Martínez, M.L.; Salvá, A.J.P. El agregado de Hordeum murinum (Poaceae) en “Flora Iberica”. Acta Bot. Malacit. 2014, 39, 311–319. [Google Scholar] [CrossRef]
- Devesa, J.A.; Téllez, T.; Benítez, M.; Olivencia, A.; Molina, R.; Claver, J. Las Gramíneas de Extremadura; Universidad de Extremadura, Servicio de Publicaciones: Badajoz, Spain, 1991. [Google Scholar]
- Braun-Blanquet, J.; Lalucat Jo, J. Fitosociología: Bases para el Estudio de las Comunidades Vegetales. Edición en Español de Pflanzensoziologie; Blume: Madrid, Spain, 1979. [Google Scholar]
- Rivas-Martínez, S.; Fernández-González, F.; Loidi, J.; Lousã, M.; Penas, A. Syntaxonomical checklist of vascular plant communities of Spain and Portugal to association level. Itinera Geobot. 2001, 14, 5–341. [Google Scholar]
- Piñar Fuentes, J.C.; Cano-Ortiz, A.; Musarella, C.M.; Pinto Gomes, C.J.; Spampinato, G.; Cano, E. Rupicolous habitats of interest for conservation in the central-southern Iberian Peninsula. Plant Sociol. 2017, 54 (Suppl. 1), 29–42. [Google Scholar] [CrossRef]
- Shannon, C.E. A mathematical theory of communication. Bell Syst. Tech. J. 1948, 27, 379–423. [Google Scholar] [CrossRef] [Green Version]
- Pielou, E.C. The measurement of diversity in different types of biological collections. J. Theor. Biol. 1966, 13, 131–144. [Google Scholar] [CrossRef]
- Simpson, E. Measurement of Diversity. Nature 1949, 163, 688. [Google Scholar] [CrossRef]
- Rivas-Martínez, S. Vascular plant communities of Spain and Portugal (addenda to the syntaxonomical checklist of 2001, part II). Itinera Geobot. 2002, 15, 433–922. [Google Scholar]
- Aranda, S.; Montes-Borrego, M.; Jiménez-Díaz, R.M.; Landa, B.B. Microbial communities associated with the root system of wild olives (Olea europaea L. subsp. europaea var. sylvestris) are good reservoirs of bacteria with antagonistic potential against Verticillium dahliae. Plant Soil 2011, 343, 329–345. [Google Scholar] [CrossRef] [Green Version]
- Marshall, E.J.P. Biodiversity, herbicides and non-target plants. Brighton Crop Prot. Conf. Weeds. 2001, 2, 855–862. [Google Scholar]
- Vyas, M.D.; Jain, A.K. Effect of pre-and post-emergence herbicides on weed control and productivity of soybean (Glycine max). Indian J. Agron. 2003, 48, 309–311. [Google Scholar]
- Marshall, J.; Brown, V.; Boatman, N.; Lutman, P.; Squire, G. The impact of herbicides on weed abundance and biodiversity PN0940. A Report for the UK Pesticides Safety Directorate. IACR Long Ashton Res. Stn. 2001, 1, 1–147. [Google Scholar]
- Tang, Y.C.; Mao, S.F.; Ma, X.Q.; Qiu, M.M.; Ma, K.; Zhu, M.X.; Wang, Z.J. The Influence of Three Different Types of Herbicides on Biodiversity. Adv. Mater. Res. 2013, 838, 2417–2426. [Google Scholar] [CrossRef]
- Kairis, O.; Karavitis, C.; Kounalaki, A.; Salvati, L.; Kosmas, C. The effect of land management practices on soil erosion and land desertification in an olive grove. Soil Use Manag. 2013, 29, 597–606. [Google Scholar] [CrossRef]
- Zhang, Y.K.; Schilling, K.E. Effects of land cover on water table, soil moisture, evapotranspiration, and groundwater re-charge: A field observation and analysis. J. Hydrol. 2006, 319, 328–338. [Google Scholar] [CrossRef]
- Gómez, J.A.; Romero, P.; Giráldez, J.V.; Fereres, E. Experimental assessment of runoff and soil erosion in an olive grove on a Vertic soil in southern Spain as affected by soil management. Soil Use Manag. 2004, 20, 426–431. [Google Scholar] [CrossRef]
- Dalley, C.D.; Bernards, M.L.; Kells, J.J. Effect of Weed Removal Timing and Row Spacing on Soil Moisture in Corn (Zea mays). Weed Technol. 2006, 20, 399–409. [Google Scholar] [CrossRef]
- Chamizo, S.; Meijide, A.; Serrano-Ortiz, P.; Sánchez-Cañete, E.P.; López-Ballesteros, A.; Kowalski, A.S. The influence of weeds on evapotranspiration and water use efficiency in an irrigated Mediterranean olive orchard. EGU Gen. Assem. Conf. Abstr. 2018, 14431. [Google Scholar]
- Wang, S.; Fu, B.J.; Gao, G.Y.; Yao, X.L.; Zhou, J. Soil moisture and evapotranspiration of different land cover types in the Loess Plateau, China. Hydrol. Earth Syst. Sci. 2012, 16, 2883–2892. [Google Scholar] [CrossRef] [Green Version]
- Fernández Luque, J.E.; Moreno Lucas, F.; Cabrera, F.; Martín Aranda, J. Determinación de las zonas radiculares activas en el olivar regado gota a gota. II Congreso Nacional de la Ciencia del Suelo. Comunicaciones 1988, 1, 40–45. [Google Scholar]
- Verhoef, A.; Diaz-Espejo, A.; Knight, J.R.; Villagarcía, L.; Fernández, J. Adsorption of Water Vapor by Bare Soil in an Olive Grove in Southern Spain. J. Hydrometeorol. 2006, 7, 1011–1027. [Google Scholar] [CrossRef] [Green Version]
- Zucco, G.; Brocca, L.; Moramarco, T.; Morbidelli, R. Influence of land use on soil moisture spatial–temporal variability and monitoring. J. Hydrol. 2014, 516, 193–199. [Google Scholar] [CrossRef]
- Lovelli, S.; Perniola, M.; Scalcione, E.; Troccoli, A.; Ziska, L.H. Future climate change in the Mediterranean area: Implica-tions for water use and weed management. Ital. J. Agron. 2012, 7, 44–49. [Google Scholar] [CrossRef] [Green Version]
- Klages, S.; Heidecke, C.; Osterburg, B. The Impact of Agricultural Production and Policy on Water Quality during the Dry Year 2018, a Case Study from Germany. Water 2020, 12, 1519. [Google Scholar] [CrossRef]
- Cano-Ortiz, A.; Fuentes, J.C.P.; Quinto-Canas, R.; Gomes, C.J.P.; Cano, E. Analysis of the Relationship between Bioclimatology and Sustainable Development. In Proceedings of the International Symposium: New Metropolitan Perspectives, Online, Italy, 26–28 May 2020; 2020; pp. 1291–1301. [Google Scholar]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ic Value | C Value |
|---|---|
| ≤8 | C = (Ic − 8) × 10 |
| 8 < Ic ≤ 18 | C = 0 |
| 18 < Ic ≤ 21 | C = (Ic − 18) × 5 |
| 21 < Ic ≤ 28 | C = (Ic − 21) × 15 |
| 28 < Ic ≤ 46 | C = (Ic − 28) × 25 |
| 46 < Ic ≤ 65 | C = (Ic − 46) × 60 |
| Phytosociological Index | % of Average Cover |
|---|---|
| 5 | 85 |
| 4 | 65 |
| 3 | 45 |
| 2 | 25 |
| 1 | 12 |
| + | 6 |
| r | 3 |
| Location | Management | Association | Location/Management | Location/Association | Management/Association | Location/Management/Association | |
|---|---|---|---|---|---|---|---|
| Lambda | 0.7227 | 0.9511 | 0.2358 | 0.8167 | 0.4691 | 0.5699 | 0.8921 |
| F (Observed values) | 0.9782 | 0.4367 | 2.0845 | 1.9077 | 0.8834 | 1.7684 | 1.0282 |
| GL1 | 18.0000 | 6.0000 | 42.0000 | 6.0000 | 48.0000 | 18.0000 | 6.0000 |
| GL2 | 144.7351 | 51.0000 | 242.6633 | 51.0000 | 255.0037 | 144.7351 | 51.0000 |
| F (Observed values) | 1.6755 | 2.2826 | 1.4350 | 2.2826 | 1.4080 | 1.6755 | 2.2826 |
| p-value | 0.4880 | 0.8509 | 0.0003 | 0.0974 | 0.6905 | 0.0345 | 0.4179 |
| Correlation F1 | Correlation F2 | Correlation F1 | Correlation F2 | ||||
|---|---|---|---|---|---|---|---|
| Edaphic variables | C.I.C (meq/100 g) | −0.0394 | 0.8076 | Bioclimatics variables | Iar | 0.9785 | 0.7685 |
| Carbonates (%) | 0.0011 | 0.4411 | Id | −0.9122 | −0.8940 | ||
| Ca (meq/100 g) | −0.0758 | 0.4859 | IH | −0.9779 | −0.7700 | ||
| P assimilable (p.p.m) | 0.0007 | 0.7312 | Ioe | −0.9779 | −0.7700 | ||
| Mg (meq/100 g) | 0.6487 | −0.1194 | Ic | −0.8990 | −0.9017 | ||
| M.O.O. (%) | 0.1588 | 0.8624 | o | −0.9631 | −0.7961 | ||
| N (%) | 0.1034 | 0.8563 | It | 0.9234 | 0.8834 | ||
| pH 1/2.5 | −0.0896 | 0.1262 | Itc | 0.9234 | 0.8834 | ||
| K (meq/100 g) | 0.3419 | 0.3974 | PEs | 0.9587 | 0.7198 | ||
| pF 1/3 atm (%) | 0.8825 | 0.0462 | PE4 | 0.9532 | 0.8381 | ||
| pF 15 atm (%) | 0.8966 | −0.0460 | PE5 | 0.9624 | 0.7354 | ||
| Tx clay (%) | 0.4742 | −0.0703 | PE6 | 0.9442 | 0.6733 | ||
| Tx sand (%) | −0.8644 | −0.0146 | PE7 | 0.9432 | 0.6571 | ||
| Tx slime (%) | 0.6321 | 0.1294 | PE8 | 0.9640 | 0.7986 | ||
| Sieve 2 mm (%) | 0.4172 | 0.1738 | PE9 | 0.9278 | 0.8751 | ||
| Salinity (mmhos/cm) | 0.2863 | 0.3869 | PE10 | 0.8816 | 0.9021 |
| Location | Grass Cover Management | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| Variables | Mean | Standard Deviation | Road vs. Tree Canopy | Road vs. Dehesa | Road vs. Fringe | Fringe vs. Tree Canopy | Fringe vs. Dehesa | Dehesa vs. Tree Canopy | Herbicide vs. No Herbicide |
| Soil Moisture 2006 (100 mm.) | 0.1917 | 0.0104 | 0.0054 | 0.3833 | 0.2411 | 0.1733 | 0.7999 | 0.6092 | 0.8754 |
| Soil Moisture 2006 (200 mm.) | 0.2058 | 0.0115 | 0.0677 | 0.3234 | 0.3909 | 0.0593 | 0.7493 | 0.0866 | 0.1109 |
| Soil Moisture 2006 (300 mm.) | 0.5685 | 0.0361 | 0.0026 | 0.2107 | 0.1220 | 0.2264 | 0.6593 | 0.8034 | 0.8180 |
| Soil Moisture 2006 (400 mm.) | 0.5281 | 0.0339 | 0.3125 | 0.8047 | 0.8168 | 0.1080 | 0.9926 | 0.1586 | 0.3030 |
| Soil Moisture 2016 (100 mm.) | 0.1632 | 0.0409 | 0.2310 | 0.2239 | 0.6483 | 0.3228 | 0.3979 | 0.5749 | <0.0001 |
| Soil Moisture 2016 (200 mm.) | 0.8483 | 0.3521 | 0.2689 | 0.2232 | 0.4733 | 0.4424 | 0.5420 | 0.6443 | <0.0001 |
| Soil Moisture 2016 (300 mm.) | 0.4809 | 0.1264 | 0.2095 | 0.2285 | 0.7952 | 0.2534 | 0.3121 | 0.5294 | <0.0001 |
| Soil Moisture 2016 (400 mm.) | 0.4309 | 0.1404 | 0.2941 | 0.2255 | 0.3928 | 0.5199 | 0.6326 | 0.6855 | <0.0001 |
| Ratio PE/Soil Moisture (100 mm.) 2006 | 4.3078 | 0.2899 | 0.0025 | 0.0486 | 0.7942 | 0.2116 | 0.3797 | 0.4429 | 0.7075 |
| Ratio PE/Soil Moisture (200 mm.) 2006 | 4.0114 | 0.2262 | 0.2761 | 0.0627 | 0.4085 | 0.4962 | 0.2779 | 0.9297 | 0.4907 |
| Ratio PE/Soil Moisture (300 mm.) 2006 | 1.4546 | 0.1126 | 0.0015 | 0.0711 | 0.9514 | 0.1261 | 0.3228 | 0.2719 | 0.5244 |
| Ratio PE/Soil Moisture (400 mm.) 2006 | 1.5663 | 0.1268 | 0.0558 | 0.0493 | 0.4942 | 0.6908 | 0.7791 | 0.8516 | 0.6781 |
| Ratio PE/Soil Moisture (100 mm.) 2016 | 5.5016 | 1.8894 | 0.3747 | 0.5445 | 0.5721 | 0.4961 | 0.9775 | 0.5668 | <0.0001 |
| Ratio PE/Soil Moisture (200 mm.) 2016 | 1.4497 | 1.2702 | 0.1238 | 0.3283 | 0.9542 | 0.4888 | 0.6248 | 0.6764 | <0.0001 |
| Ratio PE/Soil Moisture (300 mm.) 2016 | 1.8860 | 0.6901 | 0.3834 | 0.5230 | 0.5356 | 0.6030 | 0.9316 | 0.6197 | <0.0001 |
| Ratio PE/Soil Moisture (400 mm.) 2016 | 2.2807 | 1.1947 | 0.4246 | 0.6068 | 0.8782 | 0.1827 | 0.5271 | 0.5703 | <0.0001 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Piñar Fuentes, J.C.; Leiva, F.; Cano-Ortiz, A.; Musarella, C.M.; Quinto-Canas, R.; Pinto-Gomes, C.J.; Cano, E. Impact of Grass Cover Management with Herbicides on Biodiversity, Soil Cover and Humidity in Olive Groves in the Southern Iberian. Agronomy 2021, 11, 412. https://doi.org/10.3390/agronomy11030412
Piñar Fuentes JC, Leiva F, Cano-Ortiz A, Musarella CM, Quinto-Canas R, Pinto-Gomes CJ, Cano E. Impact of Grass Cover Management with Herbicides on Biodiversity, Soil Cover and Humidity in Olive Groves in the Southern Iberian. Agronomy. 2021; 11(3):412. https://doi.org/10.3390/agronomy11030412
Chicago/Turabian StylePiñar Fuentes, J.C., Felipe Leiva, Ana Cano-Ortiz, Carmelo M. Musarella, Ricardo Quinto-Canas, Carlos J. Pinto-Gomes, and Eusebio Cano. 2021. "Impact of Grass Cover Management with Herbicides on Biodiversity, Soil Cover and Humidity in Olive Groves in the Southern Iberian" Agronomy 11, no. 3: 412. https://doi.org/10.3390/agronomy11030412