
A Combined Field/Laboratory Method for Assessment of Frost Tolerance with Freezing Tests and Chlorophyll Fluorescence

Abstract
:1. Introduction
- that leaf damage assessed with chlorophyll fluorescence measurements after freezing tests does not differ between leaves left attached to the living plant and cut leaves; and
- that the onset of leaf senescence in detached leaves is sufficiently late as to not interfere with measurements of recovery after freezing tests.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Experiment | Species | Growth Conditions | Treatments | Stress Temperature | Number of Genotypes |
|---|---|---|---|---|---|
| E1 | H. vulgare | GC | E1i, E1c, E1i, E1c | −13 and no stress test | 31 |
| E2 | H. vulgare | GC | E2i, E2c, E2i, E2c | −13 and no stress test | 31 |
| E3 | H. vulgare | field | E3 | −14 | 30 |
| E4 | A. sativa | field | E4 | No stress test | 34 |
| E5 | A. sativa | field | E5 | No stress test | 46 |
| E6 | T. aestivum | field | E6 FACE, ambient & elevated CO2 | No stress test | 1 |
| E7 | T. durum | field | E7a, E7b ambient CO2 | No stress test | 1 |
| E8 | T. durum | field | E8 FACE, ambient & elevated CO2 | −14 | 12 |
| E9 | H. vulgare | field | E9 | −14 | 55 |
2. Results and Discussion
2.1. Effects of Test Conditions
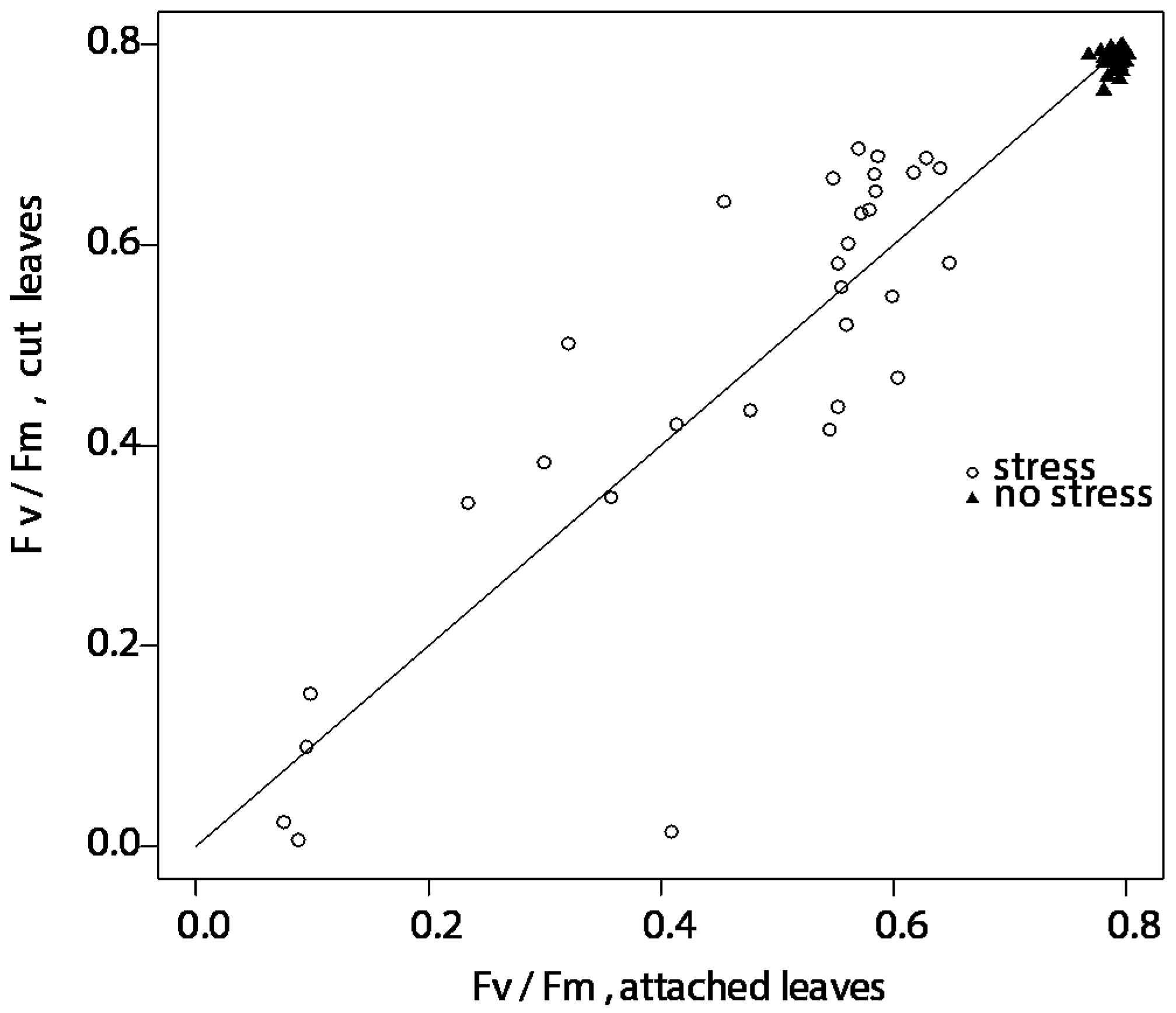
2.1.1. Effect of Test Conditions in Absence of Freezing Stress: Attached vs. Cut Leaves (Effect of Cutting) in Growth Chamber Experiments

2.1.2. Freezing Tests on Attached and Cut Leaves Lead to Consistently Similar Results in Growth Chamber Experiments
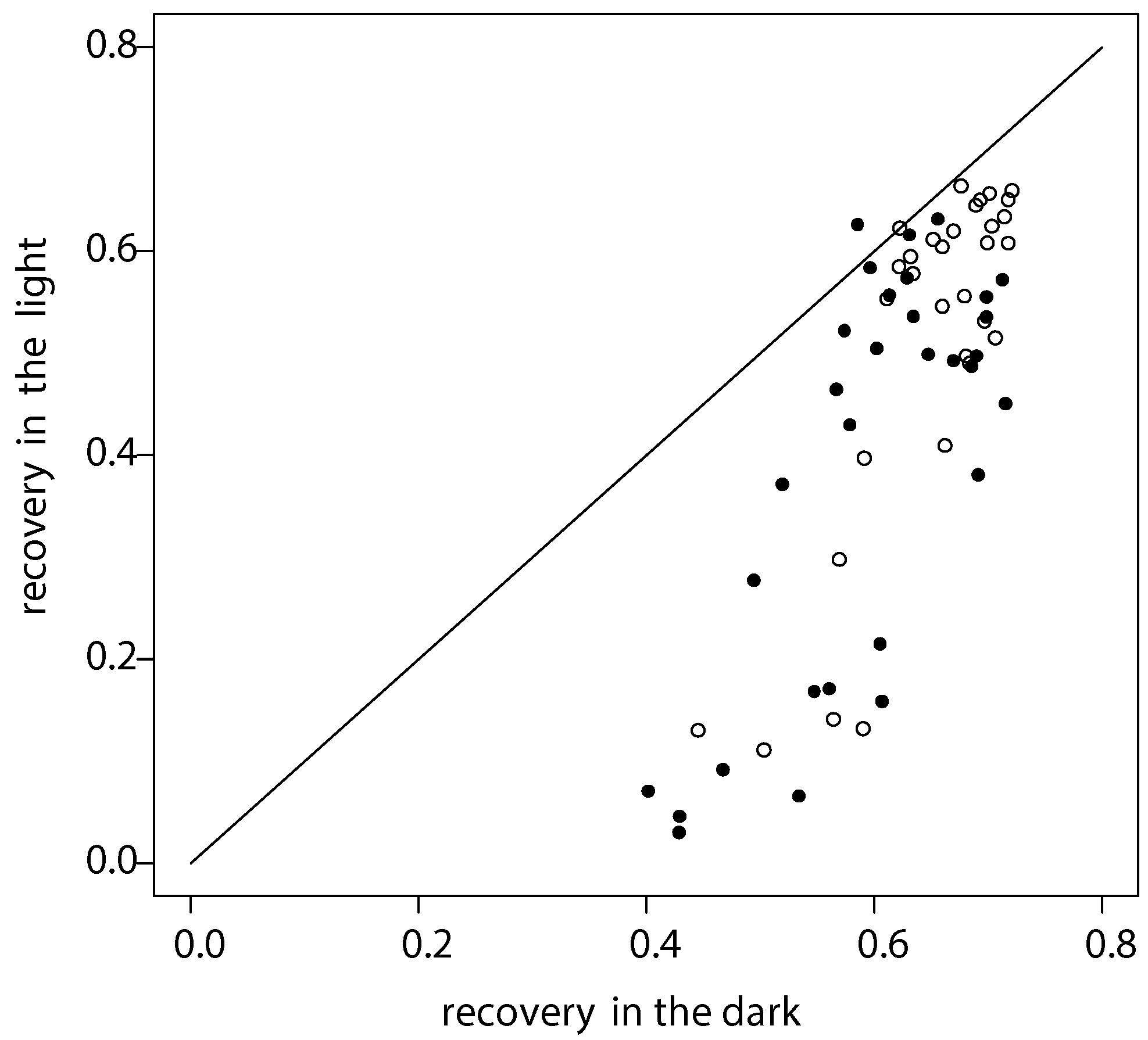
2.1.3. Daylight during Recovery Accelerates the Development of Damage Symptoms E1, E2

2.1.4. Effect of Cutting in Absence of Artificial Freezing Stress in Leaves Sampled in the Field
| Time after Sampling | Fv/Fm | Sd | Differences |
|---|---|---|---|
| 2 h | 0.68 (0.58, 0.72) | 0.061 | c |
| 4 h | 0.68 (0.57, 0.72) | 0.063 | c |
| 1 day | 0.70 (0.58, 0.74) | 0.062 | b |
| 2 days | 0.68 (0.59, 0.74) | 0.069 | c |
| 3 days | 0.72 (0.55, 0.76) | 0.077 | a |
| Treatment | Time after Sampling | Fv/Fm | Sd | Differences |
|---|---|---|---|---|
| A | 0.75 h | 0.76 (0.73–0.79) | 0.020 | c |
| A | 4 h | 0.79 (0.77–0.81) | 0.017 | b |
| A | 15.25 h | 0.81 (0.80–0.82) | 0.006 | a |
| E | 0.75 h | 0.76 (0.70–0.81) | 0.040 | c |
| E | 4 h | 0.79 (0.75–0.84) | 0.028 | bc |
| E | 15.25 h | 0.81 (0.79–0.83) | 0.014 | ab |
| Treatment | Time after Sampling | Fv/Fm | Sd | Differences |
|---|---|---|---|---|
| S1 | 0.75 h | 0.73 (0.68–0.78) | 0.036 | c |
| S1 | 4 h | 0.77 (0.75–0.80) | 0.015 | b |
| S1 | 15.25 h | 0.81 (0.80–0.83) | 0.010 | a |
| S2 | 0.75 h | 0.75 (0.70–0.80) | 0.034 | c |
| S2 | 4 h | 0.77 (0.75–0.80) | 0.019 | b |
| S2 | 15.25 h | 0.82 (0.81–0.83) | 0.010 | a |


2.2. Applications of the Field Sampling/Laboratory Test Methods
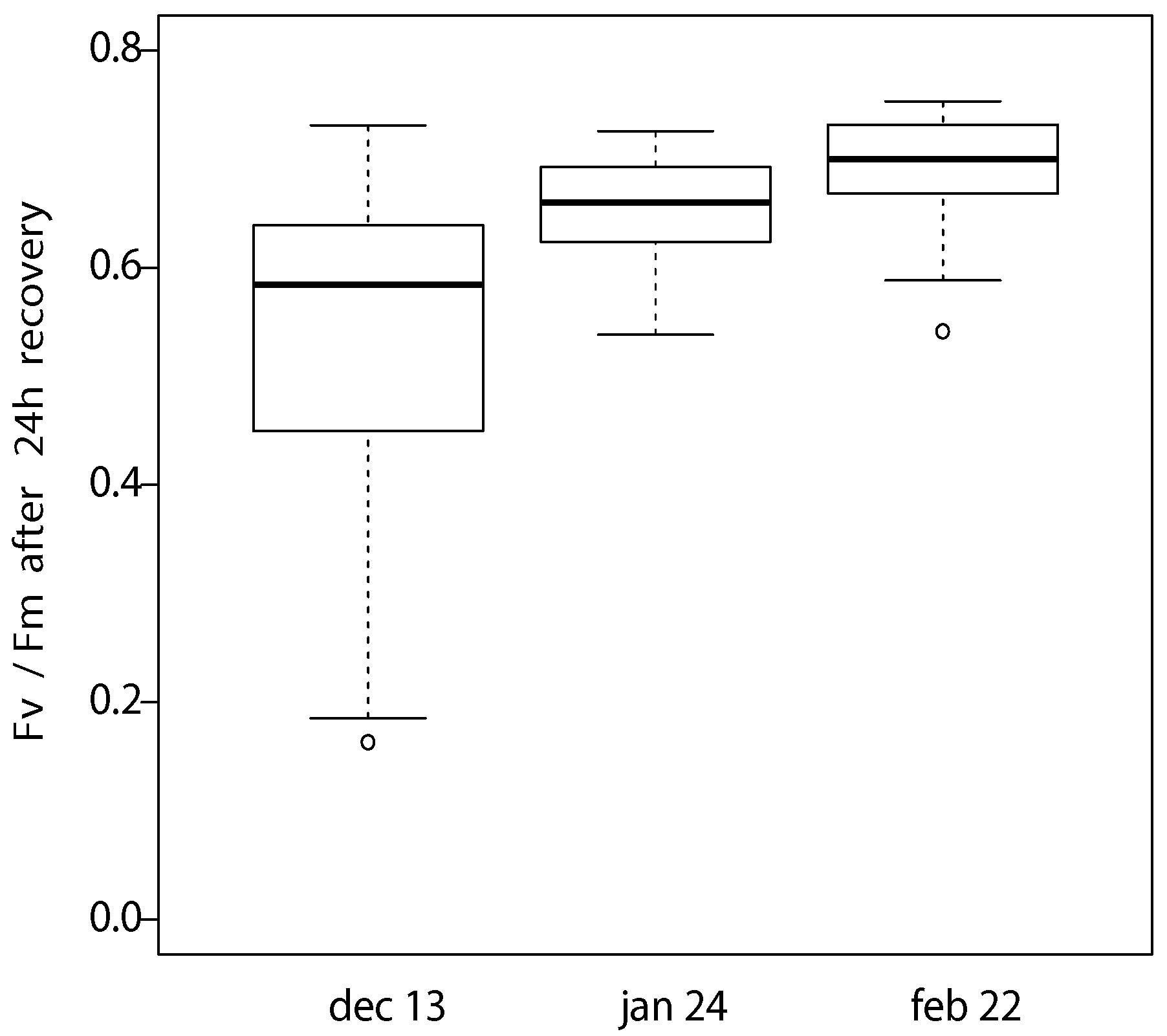
2.2.1. Seasonal Monitoring of Frost Hardiness

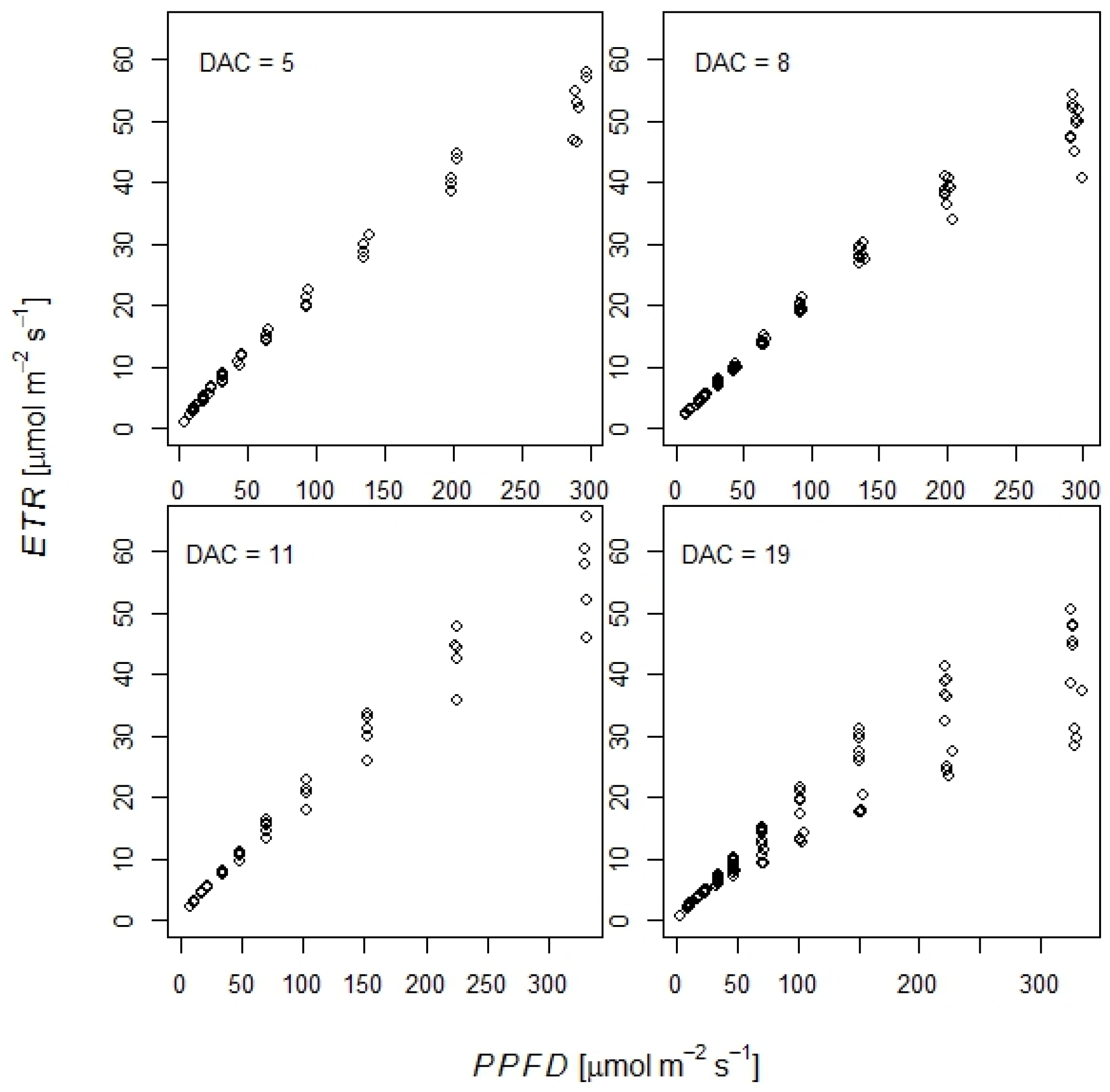
2.2.2. Effect of Elevated CO2 on Frost Tolerance of Durum Wheat
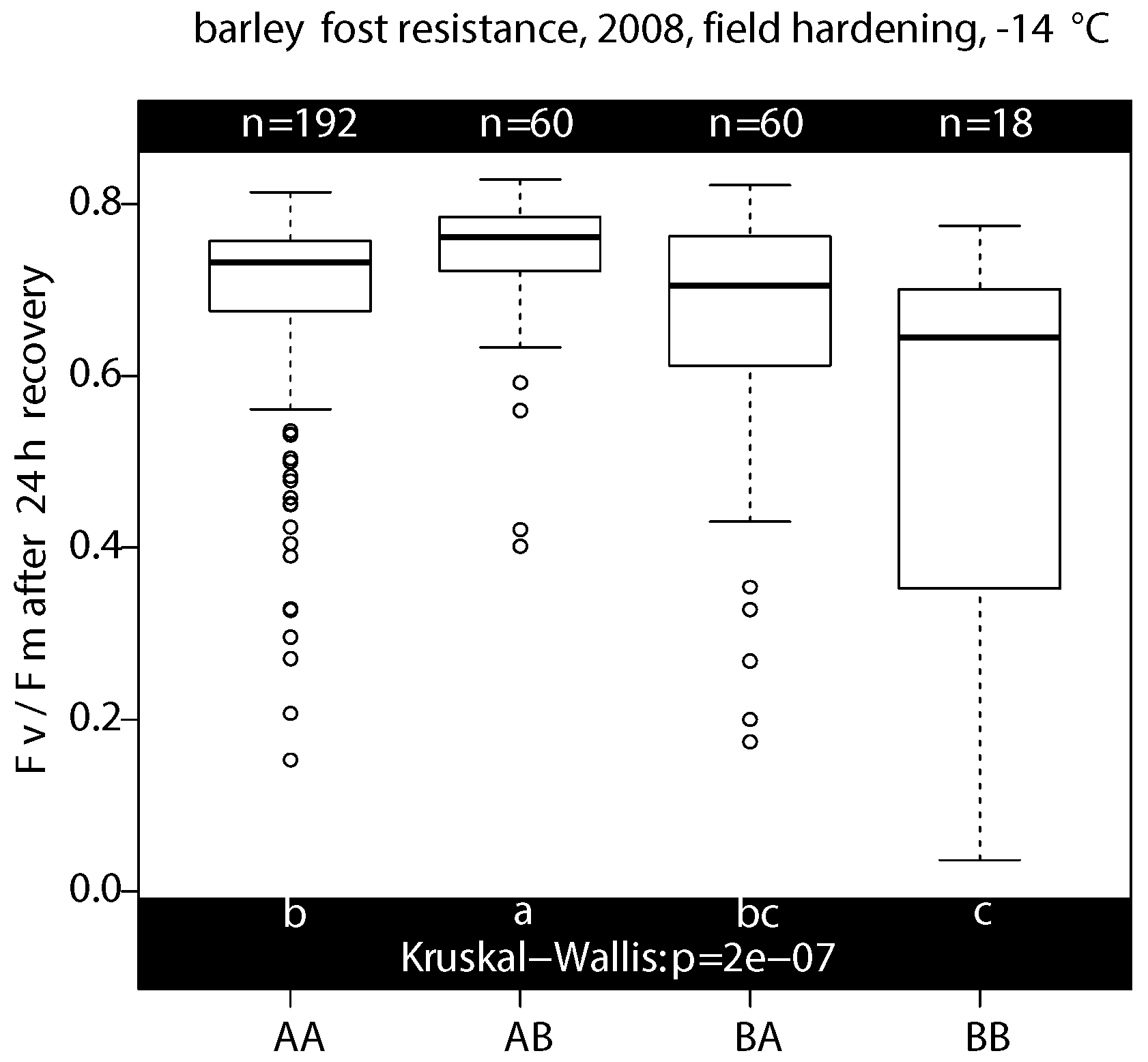
2.2.3. Phenotyping/Genotyping Study under Field Hardening Conditions

3. Experimental Section
4. Conclusions
Acknowledgments
Conflict of Interest
References
- Saulescu, N.N.; Braun, H.J. Cold tolerance. In Application of Physiology in Wheat Breeding; Reynolds, M.P., Ortiz-Monasterio, J.I., McNab, A., Eds.; CIMMYT: Mexico, DF, Mexico City, 2001; pp. 111–123. [Google Scholar]
- Levitt, J. Response of Plants to Environmental Stresses: Chilling, Freezing, and High Temperature Stresses, 2nd ed.; Academic Press: New York, NY, USA, 1980. [Google Scholar]
- Rizza, F.; Pagani, D.; Gut, M.; Prasil, I.T.; Lago, C.; Tondelli, A.; Orru, L.; Mazzucotelli, E.; Francia, E.; Badeck, F.W.; et al. Diversity in the Response to Low Temperature in Representative Barley Genotypes Cultivated in Europe. Crop Sci. 2011, 51, 2759–2779. [Google Scholar] [CrossRef]
- Thomashow, M.F. Plant cold acclimation: Freezing tolerance genes and regulatory mechanisms. Annu. Rev. Plant Physiol. Plant Mol. Biol. 1999, 50, 571–599. [Google Scholar] [CrossRef] [PubMed]
- Fowler, D.B.; Limin, A.E. Interactions among factors regulating phenological development and acclimation rate determine low-temperature tolerance in wheat. Ann. Bot. 2004, 94, 717–724. [Google Scholar] [CrossRef] [PubMed]
- Francia, E.; Rizza, F.; Cattivelli, L.; Stanca, A.M.; Galiba, G.; Toth, B.; Hayes, P.M.; Skinner, J.S.; Pecchioni, N. Two loci on chromosome 5H determine low-temperature tolerance in a “Nure” (winter) × “Tremois” (spring) barley map. Theor. Appl. Genet. 2004, 108, 670–680. [Google Scholar] [CrossRef] [PubMed]
- Akar, T.; Francia, E.; Tondelli, A.; Rizza, F.; Stanca, A.M.; Pecchioni, N. Marker-assisted characterization of frost tolerance in barley (Hordeum vulgare L.). Plant Breed. 2009, 128, 381–386. [Google Scholar] [CrossRef]
- Fisk, S.P.; Cuesta-Marcos, A.; Cistue, L.; Russell, J.; Smith, K.P.; Baenziger, S.; Bedo, Z.; Corey, A.; Filichkin, T.; Karsai, I.; et al. FR-H3: A new QTL to assist in the development of fall-sown barley with superior low temperature tolerance. Theor. Appl. Genet. 2013, 126, 335–347. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clement, J.M.A.M.; van Hasselt, P.R. Chlorophyll fluorescence as a parameter for frost hardiness in winter wheat. A comparison with other hardiness parameters. Phyton 1996, 36, 29–41. [Google Scholar]
- Herzog, H.; Olszewski, A. A rapid method for measuring freezing resistance in crop plants. J. Agron. Crop Sci. (Z. Acker Pflanzenbau) 1998, 181, 71–79. [Google Scholar] [CrossRef]
- Rizza, F.; Pagani, D.; Stanca, A.M.; Cattivelli, L. Use of chlorophyll fluorescence to evaluate the cold acclimation and freezing tolerance of winter and spring oats. Plant Breed. 2001, 120, 389–396. [Google Scholar] [CrossRef]
- Thalhammer, A.; Bryant, G.; Sulpice, R.; Hincha, D.K. Disordered Cold Regulated15 Proteins Protect Chloroplast Membranes during Freezing through Binding and Folding, But Do Not Stabilize Chloroplast Enzymes in Vivo. Plant Physiol. 2014, 166, 190–201. [Google Scholar] [CrossRef] [PubMed]
- Mishra, A.; Mishra, K.B.; Höermiller, I.I.; Heyer, A.G.; Nedbal, L. Chlorophyll fluorescence emission as a reporter on cold tolerance in Arabidopsis thaliana accessions. Plant Signal. Behav. 2011, 6, 301–310. [Google Scholar] [CrossRef] [PubMed]
- Dai, F.; Zhou, M.; Zhang, G. The change of chlorophyll fluorescence parameters in winter barley during recovery after freezing shock and as affected by cold acclimation and irradiance. Plant Physiol. Biochem. 2007, 45, 915–921. [Google Scholar] [CrossRef] [PubMed]
- Steponkus, P.L.; Webb, M.S. Freeze-induced dehydration and membrane destabilisation in plants. In Water and Life; Somero, G.N., Osmond, C.B., Bolis, C.L., Eds.; Springer: Berlin, Germany, 1992; pp. 338–362. [Google Scholar]
- Rapacz, M.; Tyrka, M.; Kaczmarek, W.; Gut, M.; Wolanin, B.; Mikulski, W. Photosynthetic acclimation to cold as a potential physiological marker of winter barley freezing tolerance assessed under variable winter environment. J. Agron. Crop Sci. 2008, 194, 61–71. [Google Scholar] [CrossRef]
- Prášil, I.T.; Prášilová, P.; Marík, P. Comparative study of direct and indirect evaluations of frost tolerance in barley. Field Crops Res. 2007, 102, 1–8. [Google Scholar] [CrossRef]
- Prášil, I.T.; Rogalewicz, V. Accuracy of wheat winterhardiness evaluation by a provocation method in natural conditions. Genetika Šlechtení (Genet. Plant Breed. Praha) 1989, 25, 223–230. [Google Scholar]
- Koch, H.-D.; Lehmann, C.O. Resistenzeigenschaften im Gersten- und Weizensortiment Gatersleben. 7. Prüfung der Frostresistenz von Wintergersten im künstlichen Gefrierversuch. Kulturpflanze 1966, 14, 263–282. [Google Scholar] [CrossRef]
- Rapacz, M. Chlorophyll a fluorescence transient during freezing and recovery in winter wheat. Photosynthetica 2007, 45, 409–418. [Google Scholar] [CrossRef]
- Rapacz, M.; Wozniczka, A. A selection tool for freezing tolerance in common wheat using the fast chlorophyll a fluorescence transient. Plant Breed. 2009, 128, 227–234. [Google Scholar] [CrossRef]
- Witkowski, E.; Waga, J.; Witkowska, K.; Rapacz, M.; Gut, M.; Bielawska, A.; Luber, H.; Lukaszewski, A.J. Association between frost tolerance and the alleles of high molecular weight glutenin subunits present in Polish winter wheats. Euphytica 2008, 159, 377–384. [Google Scholar] [CrossRef]
- Willits, D.H.; Peet, M.M. Measurement of chlorophyll fluorescence as a heat stress indicator in tomato: Laboratory and greenhouse comparisons. J. Am. Soc. Hortic. Sci. 2001, 126, 188–194. [Google Scholar]
- Wu, T.T.; Weaver, D.B.; Locy, R.D.; McElroy, S.; van Santen, E. Identification of vegetative heat-tolerant upland cotton (Gossypium hirsutum L.) germplasm utilizing chlorophyll fluorescence measurement during heat stress. Plant Breed. 2013, 133, 250–255. [Google Scholar] [CrossRef]
- Sharma, D.K.; Fernández, J.O.; Rosenqvist, E.; Ottosen, C.O.; Andersen, S.B. Genotypic response of detached leaves vs. intact plants for chlorophyll fluorescence parameters under high temperature stress in wheat. J. Plant Physiol. 2014, 171, 576–586. [Google Scholar] [CrossRef] [PubMed]
- Sthapit, B.R.; Witcombe, J.R.; Wilson, J.M. Methods of selection for chilling tolerance in Nepalese rice by chlorophyll fluorescence analysis. Crop Sci. 1995, 35, 90–94. [Google Scholar] [CrossRef]
- Strauss, A.J.; Krüger, G.H.J.; Strasser, R.J.; Heerden, P.D.R.V. Ranking of dark chilling tolerance in soybean genotypes probed by the chlorophyll a fluorescence transient O-J-I-P. Environ. Exp. Bot. 2006, 56, 147–157. [Google Scholar] [CrossRef]
- Housley, T.L.; Pollock, C.J. Photosynthesis and Carbohydrate-Metabolism in Detached Leaves of Lolium-Temulentum L. New Phytol. 1985, 99, 499–507. [Google Scholar] [CrossRef]
- Fischer, A.M. The Complex Regulation of Senescence. Crit. Rev. Plant Sci. 2012, 31, 124–147. [Google Scholar] [CrossRef]
- Min, K.; Chen, K.; Arora, R. Effect of short-term vs. prolonged freezing on freeze-thaw injury and post-thaw recovery in spinach: Importance in laboratory freeze-thaw protocols. Environ. Exp. Bot. 2014, 106, 124–131. [Google Scholar] [CrossRef]
- Mahfoozi, S.; Limin, A.E.; Fowler, D.B. Influence of vernalization and photoperiod responses on cold hardiness in winter cereals. Crop Sci. 2001, 41, 1006–1011. [Google Scholar] [CrossRef]
- Badeck, F.-W.; Rizza, F.; Maré, C.; Cattivelli, L.; Zaldei, A.; Miglietta, F. Durum wheat growth under elevated CO2: First results of a FACE experiment. In Atti del XV Convegno Nazionale di Agrometeorologia. Nuovi Scenari Agro Ambientali: Fenologia, Produzioni Agrarie ed Avversita. Palermo, 5-6-7 Giugno 2012; Ventura, F., Pieri, L., Eds.; PA.TRON EDITORE: Bologna, Italy, 2012; pp. 15–16. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing. Vienna: R Foundation for Statistical Computing. 2014. Available online: http://www.R-project.org/ (accessed on 21 November 2014).
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Badeck, F.-W.; Rizza, F. A Combined Field/Laboratory Method for Assessment of Frost Tolerance with Freezing Tests and Chlorophyll Fluorescence. Agronomy 2015, 5, 71-88. https://doi.org/10.3390/agronomy5010071
Badeck F-W, Rizza F. A Combined Field/Laboratory Method for Assessment of Frost Tolerance with Freezing Tests and Chlorophyll Fluorescence. Agronomy. 2015; 5(1):71-88. https://doi.org/10.3390/agronomy5010071
Chicago/Turabian StyleBadeck, Franz-W., and Fulvia Rizza. 2015. "A Combined Field/Laboratory Method for Assessment of Frost Tolerance with Freezing Tests and Chlorophyll Fluorescence" Agronomy 5, no. 1: 71-88. https://doi.org/10.3390/agronomy5010071