
Volatile Semiochemical Mediated Plant Defense in Cereals: A Novel Strategy for Crop Protection


International Centre of Insect Physiology and Ecology (ICIPE), P.O. Box 30772-00100 Nairobi, Kenya
*
Author to whom correspondence should be addressed.
Agronomy 2017, 7(3), 58; https://doi.org/10.3390/agronomy7030058
Submission received: 31 July 2017
/
Revised: 27 August 2017
/
Accepted: 29 August 2017
/
Published: 1 September 2017
(This article belongs to the Special Issue Plant Resistance for the Protection of Cereal Crops from Insect Pests)
{kind=link}
{kind=link}
Abstract
:Plants have evolved highly intriguing ways of defending themselves against insect attacks, including through emission of defense volatiles. These volatiles serve the plant’s defense by directly repelling phytophagous insects and/or indirectly through attracting natural enemies antagonistic to the herbivores. Several laboratory studies established the potential of improving plant resistance against insect attacks by manipulating the plant-derived volatile semiochemicals emissions. Yet, more efforts need to be conducted to translate the promising laboratory studies to fight economically-important crop pests under real field conditions. This is needed to address an increasing demand for alternative pest control options driven by ecological and environmental costs associated with the use of broad-spectrum insecticides. The practical examples discussed in this review paper demonstrate the real prospect of exploiting an inducible and constitutive plant volatile semiochemicals for developing novel and ecologically-sustainable pest management strategies to protect cereal crops from damaging insect pests.
1. Introduction
Semiochemicals (Greek semeon, a mark or signal), also known as behaviour-modifying chemicals, are chemical substances or mixtures that convey a message from one organism to another so as to modify the behaviour of the recipient [1]. Semiochemicals can broadly be divided into two classes: pheromones mediating intraspecific communication between individuals of the same species, and allelochemicals mediate interspecific interactions among individuals of different species [1]. Volatile semiochemicals are perceived by olfaction while non-volatile semiochemicals are perceived by contact chemoreception. The terms attractant and repellent are two categories of semiochemicals referring to the behaviour-modifying olfactory active compounds which do not require contact with the source. The terms “stimulant” and “deterrent” are categories of behaviour modifying chemicals that are active at close range [2].
Plants produce a wide range of volatile semiochemicals which serve various physiological and ecological functions [3]. When attacked by herbivorous insects, plants respond by emitting defense volatiles often referred as herbivore-induced plant volatiles (HIPVs) as a ‘scream’ for help from herbivores’ natural enemies and alert neighbouring plants of impeding attack [4,5]. Parasitic and predatory arthropods recognize these plant ‘screams’ as ‘dinner bells’ and move towards the plants to find the signalled food (herbivore). These plant derived ‘screams’ or semiochemical compounds could play important role in host plant resistance to insect pest damage by virtue of attracting herbivores’ natural enemies [6,7] and/or repelling attacking herbivores [8,9].
A potential to improve plant resistance against insect pests by exploiting constitutive and inducible plant volatile semiochemicals has been demonstrated in agricultural settings [10,11]. Moreover, there is an increasing interest for semiochemicals based pest management in recent years, due to ecological and environmental costs associated with use of broad-spectrum insecticides. Unlike chemical insecticides, volatile semiochemicals are difficult for insects to develop resistance against, and work by a non-toxic mode of action through modifying the behaviour of the pest or its natural enemies to the detriment of the crop pest [12,13]. Furthermore, they are non-persistent, species-specific, cost-effective, and environmentally safe [11]. This review presents concise summary about plant derived volatile semiochemicals and their applications in improving plant resistance against insect pests drawing practical examples from two crop protections strategies, namely genetic engineering (GM) and a stimulo-deterrent diversionary (push-pull) strategy.
2. Plant Volatile Semiochemicals
Plant volatiles are classes of lipophilic secondary metabolites with high vapour pressures and low molecular weights at ambient temperatures [3]. They are formed as by-products of plant processes while their precursors originate from primary metabolism [3,14]. The non-conjugated plant volatiles are released into the surrounding environment across plant membrane, from their intracellular sites of synthesis, when there are no barriers to diffusion [14]. Plants synthesize and emit a large variety of volatiles mainly from vegetative and flower parts [3], but some are emitted from roots as well [6]. Some volatiles are commonly released by all plants species, while others are specific to only one or a few related taxa [14]. So far over 1700 plant volatile compounds have been identified from different plant families belonging to both angio- and gymnosperms [15]. This figure is expected to grow as more plants are examined and advanced and sensitive volatile detection and analysis methods are developed. Despite their enormous diversity, most plant volatile compounds could be categorized into a few major classes based on their biosynthetic origin namely, terpenoids (isoprenoids), phenylpropanoids, or benzenoids, fatty acid derivatives, and amino acid derivatives [5,16]. Species-/genus-specific compounds are not represented in those major classes.
3. Herbivore Induced Plant Volatile Compounds (HIPVs)
An attack on plants by herbivorous arthropods considerably modifies the plants’ volatile emission resulting in large amounts and varieties of the so-called herbivore-induced plant volatiles (HIPVs) [3,5]. Studies in more than 15 different tritrophic systems revealed induction of broad spectra of plant volatile compounds in response to herbivory attack, suggesting that production of HIPVs is a common phenomenon [5,6]. Although the volatile profiles display an enormous range of diversity among plant species, different plant taxa exhibited a considerable overlap in production of HIPVs [5,17]. Most HIPVs could fall into the following major categories (in order of decreasing size): terpenoids (e.g., (E)-β-ciemene, (E)-4,8-dimethyl-1,3,7-nonatriene (DMNT), (E,E)-α-farnesene), fatty acid derivatives including lipoxygenase pathway products (e.g., hexanal, (Z)-3-hexen-l-ol, (Z)-3-hexenyl acetate), benzenoids/phenylpropanoids (e.g., methyl salicylate, methyl eugenol, indole), C5-branched compounds, and various nitrogen- and sulphur-containing compounds (e.g., isothiocyanates or nitriles, respectively) [3,17].
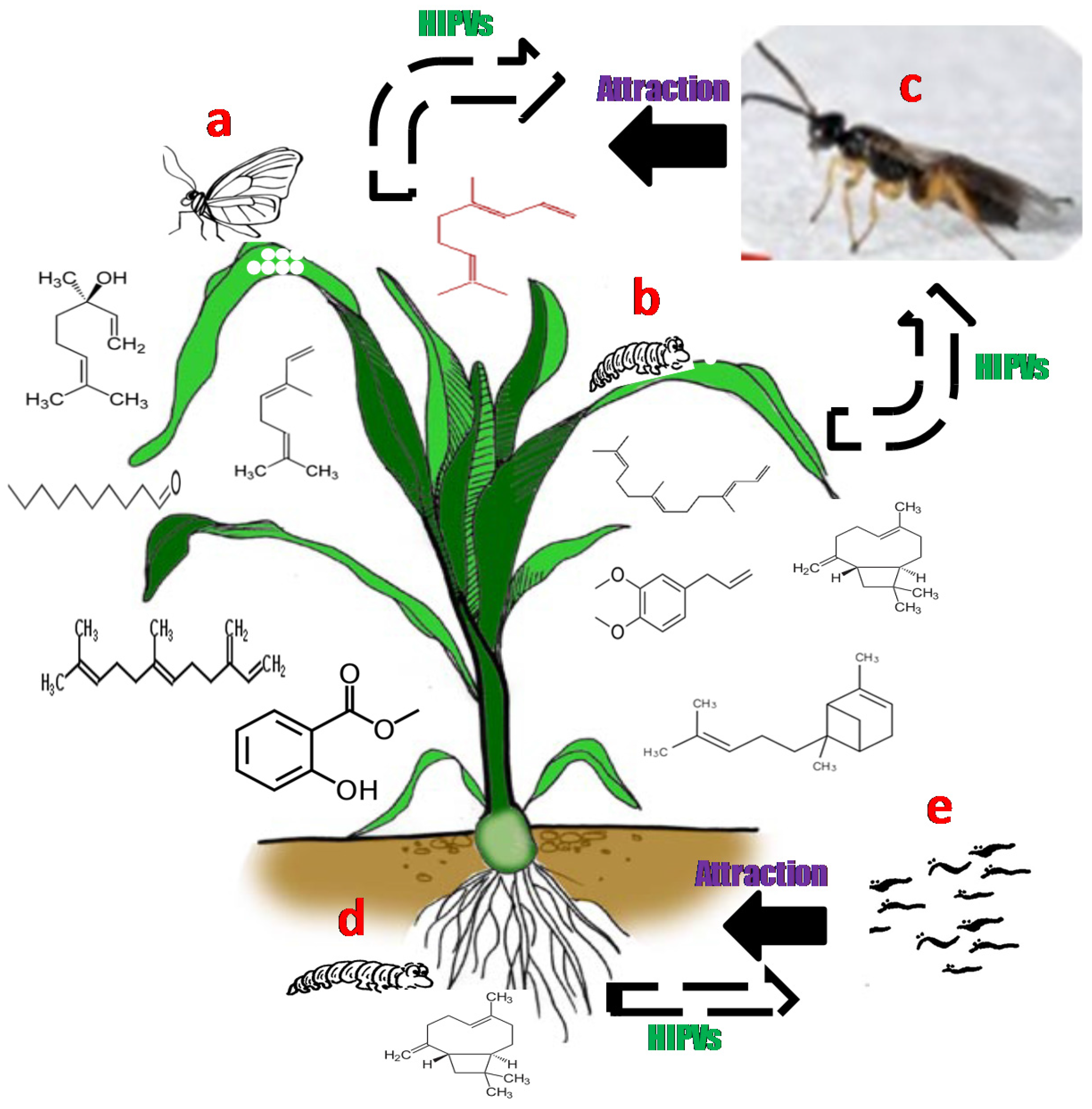
Intact plants usually release small quantities volatile compounds. However, the volatile profiles dramatically change, both in quantities and ratios, after feeding damage or egg deposition by herbivore [5,13]. The bouquets of volatile semiochemicals produced by plants in response to insect infestation provide parasitoids and predators with reliable cues to locate the attacking herbivores as they can differentially perceive the induced volatile signals in the presence of their hosts or prey [18]. For instance, the maize plant, Z. mays, has been shown to emit large quantities of different HIPVs, above and below ground, in response to insect feeding and egg deposition [6,13]. Moreover, the HIPVs were produced locally at the site of insect attack and systemically throughout the plant [13]. These HIPVs attract natural enemies of the herbivores, the parasitic wasp Cotesia sesamiae and entomopathogenic nematodes [19,20]. The identities of bioactive HIPVs emitted in response to herbivore feeding and oviposition include (E)-ocimene, (R)-linalool, methyl salicylate, decanal, methyl eugenol, (E)-caryophyllene, (E)-β-farnesene, α-bergamotene, DMNT, and (E,E)-4,8,12-trimethyl-1,3,7,11-tridecatetraene (TMTT) (Figure 1).
4. Role of Induced and Constitutive Volatiles in Plant Defence
Several studies have demonstrated that HIPVs released by plants after herbivore damage or egg deposition play important roles in plants defence against the attacking organisms [21,22,23]. The HIPVs could be involved in direct resistance by acting as feeding and/or oviposition deterrents to insect pests [8,24] and provide protection against pathogens [25]. Indirectly, they mediate above- and below-ground interactions that attract herbivores' natural enemies [6,9] and serve as signals to neighbouring plants [26]. Our recent study on a wide range of maize germplasm revealed some maize genotypes emit HIPVs attractive to egg and larval parasitic wasps at the egg-laying stage of insect attack [27,28]. This represents a sophisticated defence response which provides timely and effective biological control early on, before much damage is caused by the hatching larvae [11,28]. Some plant species constitutively release volatile semiochemicals that repel insect pests and attract natural enemies of the herbivore [29,30]. More examples on ecological functions of plant-derived volatile semiochemicals have been well documented [9,23,31,32].
Plant volatile mediated direct and indirect defense can be considered as an integral part of plant resistance as it enables recruitment of natural enemies (parasitoids and predators) that parasitizes and kill attacking insect pests and deter infestation by the herbivores [28,33]. Here, we discuss applications of plant-derived volatile semiochemicals in fighting cereal insect pests using examples from two practical crop protections strategies: (1) engineering the plants’ volatiles for their defensive functions, and (2) stimulo-deterrent diversionary (push-pull) strategies.
5. Genetic Engineering
Recent advances in biotechnology enabled experts working on the genetics of plant defence to manipulate plant volatile emissions for their defensive functions [34,35]. However, most tests have been conducted under laboratory conditions with model plants. For example, the (E)-β-farnesene synthase gene cloned from Mentha piperita was successfully expressed in Arabidopsis thaliana to cause the emission of (E)-β-farnesene [36]. The transformed A. thaliana plants emitted (E)-β-farnesene, which was sufficiently high to elicit a potent repellent response on the aphid, Myzus persicae and an arrestant response on its parasitoid, Diaeretiella rapae [36]. Transgenic maize plants with overexpression of the maize TPS10 gene produced (E)-β-farnesene, (E)-α-bergamotene and other volatile defense signals which attracted the natural enemies of maize herbivores, Cotesia marginiventris [37]. It has been proposed that similar advances in genetic engineering can be applied to enhance crop resistance to economically important insect pest under real field condition [38].
Degenhardt et al. [10] demonstrated the potential of this strategy by restoring the emission of a specific below-ground volatile signal, (E)-β-caryophyllene, to maize line that normally does not produce the signal in response to pest’s damage on the root. (E)-β-caryophyllene is released by some maize lines in response to feeding damage by larvae of western corn rootworm, Diabrotica virgifera virgifera. The semiochemical signal attracts the entomopathogenic nematodes Heterorhabditis megidis, which parasitizes and kills the pest larvae within a few days [19,39]. Most North American maize cultivars have reportedly lost the ability to produce the volatile signal and hence, receive little protection from entomopathogenic nematodes [10]. In this study, maize line that normally does not emit (E)-β-caryophyllene was transformed with a (E)-β-caryophyllene synthase gene from oregano (Origanum vulgare), resulting in constitutive emissions of the signal. In corn rootworm-infested field plots where entomopathogenic nematodes were released, the (E)-β-caryophyllene-emitting plants suffered significantly less root damage. Moreover, there was significantly lower survival of the pest (60% fewer emergence of adult beetles) on transformed plants when compared to untransformed, non-emitting lines [10].
More recently, a hexaploid wheat, Triticum aestivum cv. Cadenza was genetically engineered to emit (E)-β-farnesene using a synthetic gene based on a sequence from peppermint with a plastid targeting amino acid sequence, with or without a gene for biosynthesis of the precursor farnesyl diphosphate [40]. In the laboratory bioassay, volatiles from transformed plants were repellent to three species of cereal aphids and significantly enhanced the foraging behaviour of the parasitic wasp Aphidius ervi. Although the laboratory studies demonstrated considerable potential for aphid control, field experiment during 2012/2013 cropping seasons did not show evidence of aphid repellence or increased parasitism under field conditions [40]. Modification of the experimental approach has been recommended to further test the promising laboratory results under realistic field conditions [40]. The proposed modifications include considering aphid feeding-associated induced effects and targeting more reliable natural populations of aphids and parasitoids [9,40].
The above examples demonstrate the potential of manipulating plant volatile semiochemicals to provide better crop resistance against insect attack through improved biological control of the pest. Genetic engineering is faster, efficient, and more precise than conventional breeding in introgressing desirable defense volatile signals [41,42]. Nevertheless, conventional breeding could also be deployed to optimize the production of important bioactive volatiles by plants to balance the influence of pressure groups and regulatory restrictions to transgenic approach. The speed of conventional breeding can be improved with marker-assisted selection [28]. Moreover, the modified plant volatile semiochemicals could preferably be improved by making the emissions inducible by the target pest to selectively guide the natural enemies to those plants under insect attack and minimize metabolic costs and potential pest adaptation associated with constitutive HIPVs emissions.
6. Stimulo-Deterrent Diversionary Strategy
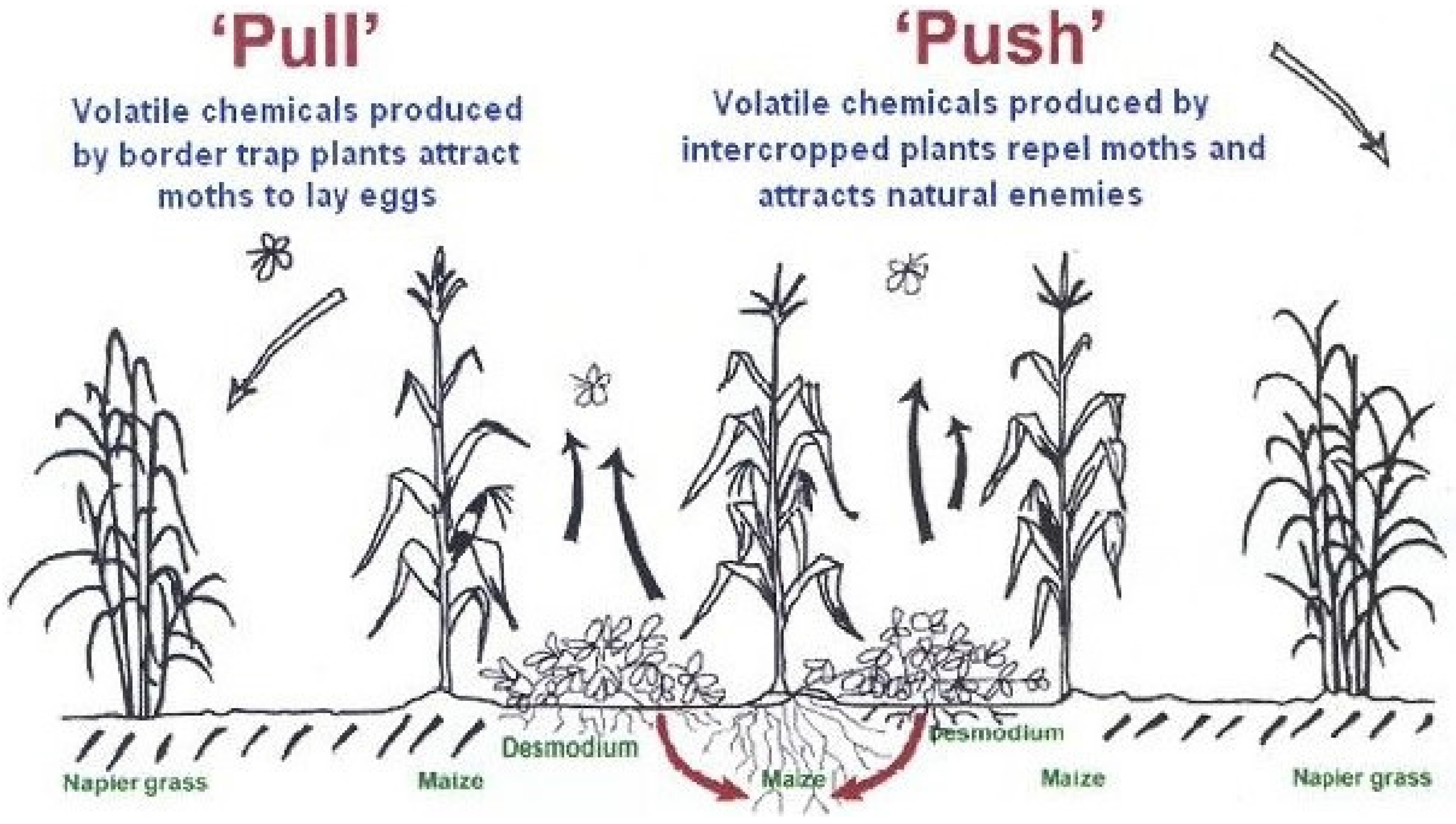
A stimulo-deterrent diversionary strategy, also called push-pull, is developed by exploiting plant volatile semiochemicals to repel insect pests from the main crop (push) and attract them into trap crops (pull) [2]. One of the most successful examples of a stimulo-deterrent diversionary strategy is a ‘push-pull’ cropping system developed to control cereal stemborers for resource-poor farmers in Africa [30]. The system involves intercropping cereal crops maize or sorghum, with the fodder legumes desmodium spp. (silverleaf Desmodium uncinatum or greenleaf D. intortum) and molasses grass (Melinis minutiflora) and planting Napier grass (Pennisetum purpureum) or Sudan grass (Sorghum vulgare sudanense) around this intercrop (Figure 2). Volatile semiochemicals such as (E)-ocimene and (E)-4,8-dimethyl-1,3,7-nonatriene produced by the desmodium and molasses repel (push) push away the stemborer pests while those produced by Napier and Sudan grasses attract (pull) stemborer moths to lay eggs in the grass instead of maize [30]. In addition to producing repellent volatile semiochemicals that push away the stemborer moths, the root exudates of Desmodium spp have been shown to effectively suppress the parasitic weed, Striga spp. through an allelopathic mechanism [11,30].
Napier grass has been shown to produce higher levels of attractive volatile semiochemicals, such as the green leaf volatiles hexanal, (E)-2-hexenal, (Z)-3-hexen-1-ol, and (Z)-3-hexen-1-yl acetate, cues used by gravid female stemborer moths in the host plant location, compared to maize or sorghum [30]. Moreover, Napier grass tissues produce a sticky sap in response to feeding by the stemborer larvae which traps them causing significant mortality [11,30]. In this way, it is possible to divert stemborers from the main crop and significantly reduce damage caused by the pests. Planting intercrops, such as molasses grass between rows of maize not only reduces stemborer infestation but also increase parasitism of stemborer larvae by the parasitic wasp C. sesamiae [43]. Behavioural and analytical study results revealed female C. sesamiae was significantly attracted to (E)-4,8-dimethyl-1,3,7-nonatriene (DMNT), volatile signals emitted by intact molasses grass [44]. DMNT is also released in response to herbivore egg deposition and feeding on maize plants and has been implicated in recruiting parasitoids and predators [13,45]. The ‘push-pull’ system [46] provide an excellent example of exploiting plants with inherent ability to produce appropriate semiochemicals to develop innovative and ecologically-sound crop protection strategies. In this strategy, the combined effects of volatile semiochemicals from the attractive trap and repellent intercrops has resulted in a significant reduction of stemborer pest damage on cereal crops leading to major improvements in crop yield gains [11].
7. Conclusions
There is an intricate inter-trophic chemical interaction between plants, herbivores, and herbivores’ natural enemies as they have been coevolving for over 400 million years [47,48]. As a result, plants have evolved highly sophisticated ways of defending themselves from herbivorous insects such as, through emission of volatile semiochemicals. Some of these volatile semiochemicals are constitutively emitted while others are induced after insect attack [9]. Both constitutive and induced defenses play an important role in protecting plants from insect damage [9,24]. Several studies have shown the potential of manipulating plant volatile semiochemicals to reduce herbivory and enhance attraction of herbivore’ natural enemies, though mainly under laboratory conditions [23,31]. The practical examples in this paper further demonstrate the real prospect of improving direct and indirect crop resistance against economically-important insect pests by exploiting plant-derived volatile semiochemicals. Proper understanding of biochemical, molecular and behavioural ecology of plant-insect interactions is essential to develop similar novel and sustainable insect pest management strategies [10,11]. Additionally, recent advances in the biotechnology could provide new tools to improve crop genetics for better resistance against insect pest attack [28,49].
Acknowledgments
We gratefully acknowledge the financial support by the following organizations and agencies: the European Union, Biovision Foundation, UK’s Department for International Development (DFID); Swedish International Development Cooperation Agency; the Swiss Agency for Development and Cooperation (SDC); and the Kenyan Government. The views expressed herein do not necessarily reflect the official opinion of these donors.
Conflicts of Interest
The authors do not have any conflict of interest.
References
- Law, J.H.; Regnier, F.E. Pheronomes. Annu. Rev. Biochem. 1971, 40, 533–548. [Google Scholar] [CrossRef] [PubMed]
- Cook, S.M.; Khan, Z.R.; Pickett, J.A. The use of Push-Pull strategies in integrated pest management. Annu. Rev. Entomol. 2007, 52, 375–400. [Google Scholar] [CrossRef] [PubMed]
- Dudareva, N.; Klempien, A.; Muhlemann, J.K.; Kaplan, I. Biosynthesis, function and metabolic engineering of plant volatile organic compounds. New Phytol. 2013, 198, 16–32. [Google Scholar] [CrossRef] [PubMed]
- Kost, C.; Heil, M. Herbivore-induced plant volatiles induce an indirect defence in neighbouring plants. J. Ecol. 2006, 94, 619–628. [Google Scholar] [CrossRef]
- Dicke, M.; van Loon, J.J.A.; Soler, R. Chemical complexity of volatiles from plants induced by multiple attack. Nat. Chem. Biol. 2009, 5, 317–324. [Google Scholar] [CrossRef] [PubMed]
- Degenhardt, J. Indirect defence responses to herbivory in grasses. Plant Physiol. 2009, 149, 96–102. [Google Scholar] [CrossRef] [PubMed]
- Büchel, K.; Malskies, S.; Mayer, M.; Fenning, T.M.; Gershenzon, J.; Hilker, M.; Meiners, T. How plants give early herbivore alert: Volatile terpenoids attract parasitoids to egg-infested elms. Basic Appl. Ecol. 2011, 12, 403–412. [Google Scholar] [CrossRef]
- De Moraes, C.M.; Mescher, M.C.; Tumlinson, J.H. Caterpillar-induced nocturnal plant volatiles repel nonspecific females. Nature 2001, 410, 577–580. [Google Scholar] [CrossRef] [PubMed]
- Pickett, J.A.; Khan, Z.R. Plant volatile-mediated signalling and its application in agriculture: Successes and challenges. New Phytol. 2016, 212, 856–870. [Google Scholar] [CrossRef] [PubMed]
- Degenhardt, J.; Hiltpold, I.; Köllner, T.G.; Frey, M.; Gierl, A.; Gershenzon, J.; Hibbard, B.E.; Ellersieck, M.R.; Turlings, T.C.J. Restoring a maize root signal that attracts insect-killing nematodes to control a major pest. Proc. Natl. Acad. Sci. USA. 2009, 32, 13213–13218. [Google Scholar] [CrossRef] [PubMed]
- Khan, Z.R.; Midega, C.A.O.; Hooper, A.M.; Pickett, J.A. Push-Pull: Chemical Ecology-Based Integrated Pest Management Technology. J. Chem. Ecol. 2016, 42, 689–697. [Google Scholar] [CrossRef] [PubMed]
- Bruce, T.J.A. Tackling the threat to food security caused by crop pests in the new millennium. Food Secur. 2010, 2, 133–141. [Google Scholar] [CrossRef]
- Tamiru, A.; Bruce, T.J.A.; Woodcock, C.M.; Caulfield, J.C.; Midega, C.A.O.; Ogol, C.K.P.O.; Mayon, P.; Birkett, M.A.; Pickett, J.A.; Khan, Z.R. Maize landraces recruit egg and larval parasitoids in response to egg deposition by a herbivore. Ecol. Lett. 2011, 14, 1075–1083. [Google Scholar] [CrossRef] [PubMed]
- Pichersky, E.; Noel, J.P.; Dudareva, N. Biosynthesis of plant volatile: Nature’s diversity and ingenuity. Science 2006, 311, 808–818. [Google Scholar] [CrossRef] [PubMed]
- Knudsen, J.T.; Eriksson, R.; Gershenzon, J.; Stahl, B. Diversity and distribution of floral scent. Bot. Rev. 2006, 72, 1–120. [Google Scholar] [CrossRef]
- Arimura, G.; Matsui, K.; Takabayashi, J. Chemical and molecular ecology of herbivore-induced plant volatiles: Proximate factors and their ultimate functions. Plant Cell Physiol. 2009, 50, 911–923. [Google Scholar] [CrossRef] [PubMed]
- Tamiru, A.; Bruce, T.J.A.; Woodcock, C.M.; Birkett, M.A.; Midega, C.A.O.; Pickett, J.A.; Khan, Z.R. Chemical cues modulating electrophysiological and behavioral responses in the parasitic wasp Cotesia sesamiae. Can. J. Zool. 2015, 93, 281–287. [Google Scholar] [CrossRef]
- Heil, M. Herbivore-induced plant volatiles: Targets, perception and unanswered questions. New Phytol. 2014, 204, 297–306. [Google Scholar] [CrossRef]
- Rasmann, S.; Köllner, T.G.; Degenhardt, J.; Hiltpold, I.; Toepfer, S.; Kuhlmann, U.; Gershenzon, J.; Turlings, T.C.J. Recruitment of entomopathogenic nematodes by insect-damaged maize roots. Nature 2005, 434, 732–737. [Google Scholar] [CrossRef] [PubMed]
- Tamiru, A.; Bruce, T.J.A.; Richter, A.; Woodcock, C.M.; Midega, C.A.O.; Degenhardt, J.; Kelemu, S.; Pickett, J.A.; Khan, Z.R. A maize landrace that emits defense volatiles in response to herbivore eggs possesses a strongly inducible terpene synthase gene. Ecol. Evol. 2017, 7, 2835–2845. [Google Scholar] [CrossRef] [PubMed]
- Hilker, M.; Meiners, T. Early herbivore alert: Insect eggs induce plant defense. J. Chem. Ecol. 2006, 32, 1379–1397. [Google Scholar] [CrossRef] [PubMed]
- Heil, M. Indirect defence via tritrophic interactions. New Phytol. 2008, 178, 41–61. [Google Scholar] [CrossRef] [PubMed]
- Hare, J.D. Ecological role of volatiles produced by plants in response to damage by herbivorous insects. Ann. Rev. Entomol. 2011, 56, 161–180. [Google Scholar] [CrossRef] [PubMed]
- Kessler, A.; Baldwin, I.T. Defensive function of herbivore-induced plant volatile emissions in nature. Science 2001, 291, 2141–2144. [Google Scholar] [CrossRef] [PubMed]
- Huang, M.; Sanchez-Moreiras, A.M.; Abel, C.; Sohrabi, R.; Lee, S.; Gershenzon, J.; Tholl, D. The major volatile organic compound emitted from Arabidopsis thaliana flowers, the sesquiterpene (E)-β-caryophyllene, is a defense against a bacterial pathogen. New Phytol. 2012, 193, 997–1008. [Google Scholar] [CrossRef] [PubMed]
- Baldwin, I.T.; Halitschke, R.; Paschold, A.; von Dahl, C.C.; Catherine, A.; Preston, C.A. Volatile Signaling in Plant-Plant Interactions: “Talking Trees” in the Genomics Era. Science 2006, 311, 812–815. [Google Scholar] [CrossRef] [PubMed]
- Tamiru, A.; Bruce, T.J.A.; Midega, C.A.O.; Woodcock, C.M.; Birkett, M.A.; Pickett, J.A.; Khan, Z.R. Oviposition induced volatile emissions from African smallholder farmers’ maize varieties. J. Chem. Ecol. 2012, 38, 231–234. [Google Scholar] [CrossRef] [PubMed]
- Tamiru, A.; Khan, Z.R.; Bruce, T.J.A. New directions for improving crop resistance to insects by breeding for egg induced defence. Curr. Opin. Insect Sci. 2015, 9, 51–55. [Google Scholar] [CrossRef]
- Glinwood, R.; Ahmed, E.; Qvarfordt, E.; Ninkovic, V.; Pettersson, J. Airborne interactions between undamaged plants of different cultivars affect insect herbivores and natural enemies. Arth. Plant Int. 2009, 3, 215–224. [Google Scholar] [CrossRef]
- Khan, Z.R.; Midega, C.A.O.; Bruce, T.J.A.; Hooper, A.M.; Pickett, J.A. Exploiting phytochemicals for developing a ‘push-pull’ crop protection strategy for cereal farmers in Africa. J. Exp. Bot. 2010, 61, 4185–4196. [Google Scholar] [CrossRef] [PubMed]
- Unsicker, S.B.; Kunert, G.; Gershenzon, J. Protective perfumes: The role of vegetative volatiles in plant defense against herbivores. Curr. Opin. Plant Biol. 2009, 12, 479–485. [Google Scholar] [CrossRef] [PubMed]
- Mumm, R.; Dicke, M. Variation in natural plant products and the attraction of bodyguards involved in indirect plant defense. Can. J. Zool. 2010, 88, 628–667. [Google Scholar] [CrossRef]
- Åhman, I.; Glinwood, R.; Ninkovic, V. The potential for modifying plant volatile composition to enhance resistance to arthropod pests. CAB Rev. 2010, 5, 1–10. [Google Scholar] [CrossRef]
- Kappers, I.F.; Aharoni, A.; van Herpen, T.W.J.M.; Luckerhoff, L.L.P.; Dicke, M.; Bouwmeester, H.J. Genetic engineering of terpenoid metabolism attracts, bodyguards to Arabidopsis. Science 2005, 309, 2070–2072. [Google Scholar] [CrossRef] [PubMed]
- Schnee, C.; Köllner, T.G.; Held, M.; Turlings, T.C.J.; Gershenzon, J.; Degenhardt, J. The products of a single maize sesquiterpene synthase form a volatile defence signal that attracts natural enemies of maize herbivores. Proc. Natl. Acad. Sci. USA 2006, 103, 1129–1134. [Google Scholar] [CrossRef] [PubMed]
- Beale, M.H.; Birkett, M.A.; Bruce, T.J.A.; Chamberlain, K.; Field, L.M.; Huttly, A.K.; Martin, J.L.; Parker, R.; Phillips, A.L.; Pickett, J.A.; et al. Aphid alarm pheromone produced by transgenic plants affects aphid and parasitoid behavior. Proc. Natl. Acad. Sci. USA 2006, 103, 10509–10513. [Google Scholar] [CrossRef] [PubMed]
- Kos, M.; van Loon, J.J.A.; Dicke, M.; Vet, L.E.M. Transgenic plants as vital components of integrated pest management. Trends Biotechnol. 2009, 27, 621–627. [Google Scholar] [CrossRef] [PubMed]
- Bruce, T.J.A. GM as a route for delivery of sustainable crop protection. J. Exp. Bot. 2012, 63, 537–541. [Google Scholar] [CrossRef] [PubMed]
- Kurtz, B.; Hiltpold, I.; Turlings, T.C.J.; Kuhlmann, U.; Toepfer, S. Comparative susceptibility of larval instars and pupae of the western corn rootworm to infection by three entomopathogenic nematodes. Biocontrol 2009, 54, 255–262. [Google Scholar] [CrossRef]
- Bruce, T.J.A.; Aradottir, G.I.; Smart, L.E.; Martin, J.L.; Caulfield, J.C.; Doherty, A.; Sparks, C.A.; Woodcock, C.M.; Birkett, M.A.; Naipier, J.A.; et al. The first crop plant genetically engineered to release an insect pheromone for defence. Sci. Rep. 2015, 5, 118–125. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Konermann, S.; Brigham, M.D.; Trevino, A.E.; Joung, J.; Abudayyeh, O.O.; Barcena, C.; Hsu, P.D.; Habib, N.; Gootenberg, J.S.; Nishimasu, H.; et al. Genome-scale transcriptional activation by an engineered CRISPR-Cas9 complex. Nature 2014, 517, 583–588. [Google Scholar] [CrossRef] [PubMed]
- Voytas, D.F.; Gao, C. Precision genome engineering and agriculture: Opportunities and regulatory challenges. PLoS Biol. 2014, 12, e1001877. [Google Scholar] [CrossRef] [PubMed]
- Khan, Z.R.; Ampong-Nyarko, K.; Chilishwa, P.; Hassanali, A.; Kimani, S.; Lwande, W.; Overholt, W.A.; Pickett, J.A.; Smart, L.E.; Wadhams, L.J.; et al. Intercropping increases parasitism of pests. Nature 1997, 388, 631–632. [Google Scholar] [CrossRef]
- Khan, Z.R.; Pickett, J.A.; Van Den Berg, J.; Wadhams, L.J.; Woodcock, C.M. Exploiting chemical ecology and species diversity: Stemborer and Striga control for maize and sorghum in Africa. Pest Manag. Sci. 2000, 56, 957–962. [Google Scholar] [CrossRef]
- Turlings, T.C.J.; Wäckers, F. Recruitment of predators and parasitoids by herbivore-injured plants. In Advances in Insect Chemical Ecology; Carde, R.T., Millar, J.G., Eds.; Cambridge University Press: Cambridge, UK, 2004; pp. 21–75. [Google Scholar]
- A novel farming system for ending hunger and poverty in sub-Sanharan Africa. Available online: http://www.push-pull.net/works.shtml (accessed on 20 July 2017).
- Labandeira, C.C. A paleobiologic perspective on plant–insect interactions. Curr. Opin. Plant Biol. 2013, 16, 414–421. [Google Scholar] [CrossRef] [PubMed]
- Moles, A.T.; Peco, B.; Wallis, I.R.; Foley, W.J.; Poore, A.G.; Seabloom, E.W.; Vesk, P.A.; Bisigato, A.J.; Cella-Pizarro, L.; Clark, C.J. Correlations between physical and chemical defences in plants: Tradeoffs, syndromes, or just many different ways to skin a herbivorous cat? New Phytol. 2013, 198, 252–263. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seifi, A.; Visser, R.G.F.; Bai, Y. How to effectively deploy plant resistances to pests and pathogens in crop breeding. Euphytica 2013, 190, 321–334. [Google Scholar] [CrossRef]
Figure 1.
Aboveground and belowground herbivore-induced plant volatiles (HIPVs) emitted by maize plant in response to spotted stemborer Chilo partellus oviposition (a) and feeding (b) and the corn rootworm (Diabrotica virgifera virgifera) feeding (d). The bioactive HIPVs produced in response to the herbivores infestation were attractive to natural enemies of the herbivores, i.e., the parasitic wasp Cotesia sesamia (c) and entomopathogenic nematode (e).
Figure 1.
Aboveground and belowground herbivore-induced plant volatiles (HIPVs) emitted by maize plant in response to spotted stemborer Chilo partellus oviposition (a) and feeding (b) and the corn rootworm (Diabrotica virgifera virgifera) feeding (d). The bioactive HIPVs produced in response to the herbivores infestation were attractive to natural enemies of the herbivores, i.e., the parasitic wasp Cotesia sesamia (c) and entomopathogenic nematode (e).
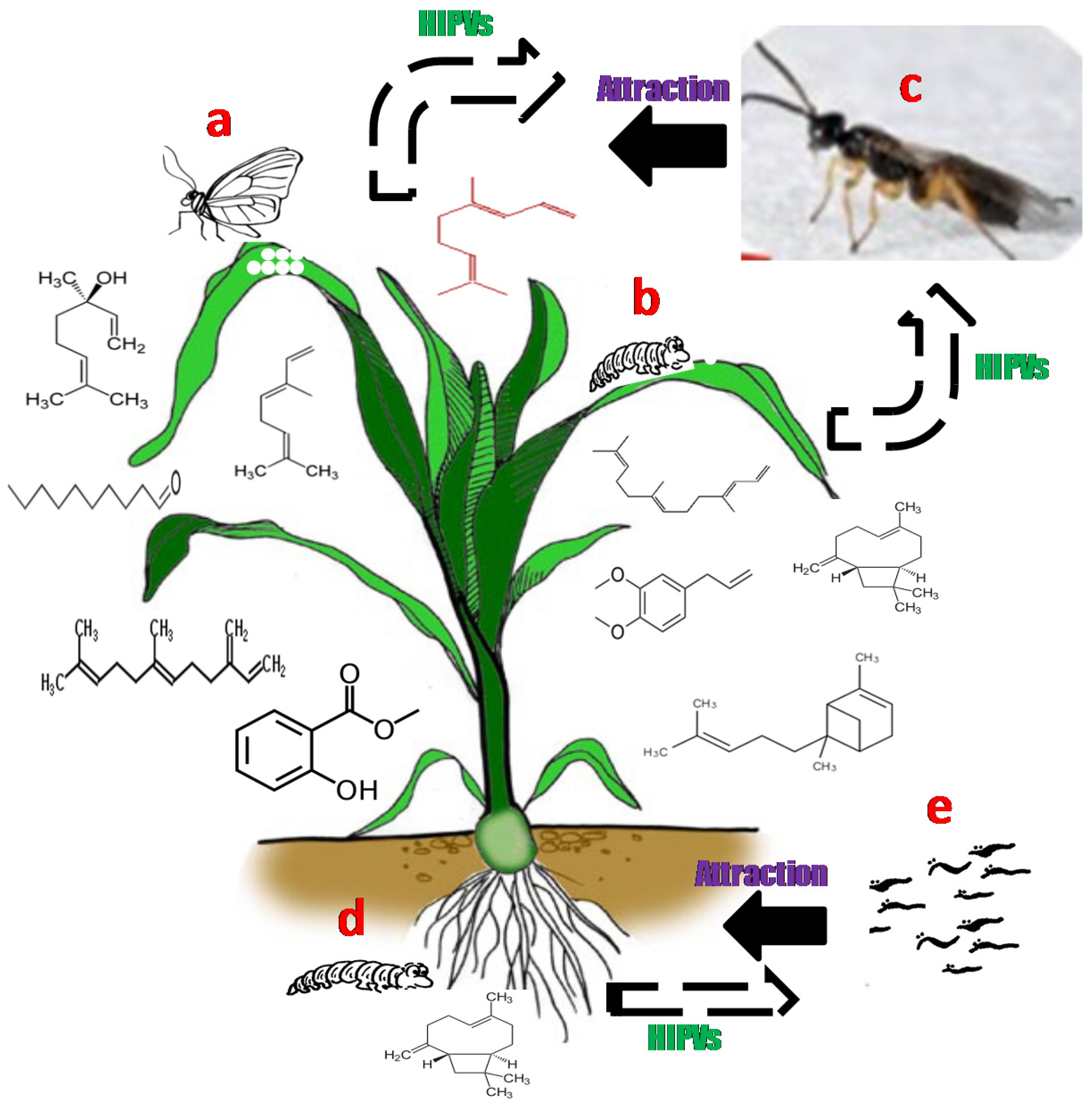
Figure 2.
Schematic representation of “push-pull” cropping system (source: www.push-pull.net) [46].
Figure 2.
Schematic representation of “push-pull” cropping system (source: www.push-pull.net) [46].
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Tamiru, A.; Khan, Z.R. Volatile Semiochemical Mediated Plant Defense in Cereals: A Novel Strategy for Crop Protection. Agronomy 2017, 7, 58. https://doi.org/10.3390/agronomy7030058
AMA Style
Tamiru A, Khan ZR. Volatile Semiochemical Mediated Plant Defense in Cereals: A Novel Strategy for Crop Protection. Agronomy. 2017; 7(3):58. https://doi.org/10.3390/agronomy7030058
Chicago/Turabian StyleTamiru, Amanuel, and Zeyaur R. Khan. 2017. "Volatile Semiochemical Mediated Plant Defense in Cereals: A Novel Strategy for Crop Protection" Agronomy 7, no. 3: 58. https://doi.org/10.3390/agronomy7030058
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.