Trade-Offs in Arbuscular Mycorrhizal Symbiosis: Disease Resistance, Growth Responses and Perspectives for Crop Breeding


1
John Innes Centre, Norwich Research Park, Norwich NR4 7UH, UK
2
Centre of Excellence for Plant and Microbial Science, Institute of Plant Physiology and Ecology, Institutes for Biological Sciences, Chinese Academy of Sciences, Shanghai 200032, China
*
Author to whom correspondence should be addressed.
Agronomy 2017, 7(4), 75; https://doi.org/10.3390/agronomy7040075
Submission received: 5 October 2017
/
Revised: 30 October 2017
/
Accepted: 30 October 2017
/
Published: 16 November 2017
(This article belongs to the Special Issue Disease Resistance Trade-offs in Crop Breeding for Disease Resistance)
Abstract
:There is an increasing need to develop high-yielding, disease-resistant crops and reduce fertilizer usage. Combining disease resistance with efficient nutrient assimilation through improved associations with symbiotic microorganisms would help to address this. Arbuscular mycorrhizal fungi (AMF) form symbiotic relationships with most terrestrial plants, resulting in nutritional benefits and the enhancement of stress tolerance and disease resistance. Despite these advantages, arbuscular mycorrhizal (AM) interactions are not normally directly considered in plant breeding. Much of our understanding of the mechanisms of AM symbiosis comes from model plants, which typically exhibit positive growth responses. However, applying this knowledge to crops has not been straightforward. In many crop plants, phosphate uptake and growth responses in AM-colonized plants are variable, with AM plants exhibiting sometimes zero or negative growth responses and lower levels of phosphate acquisition. Host plants must also balance the ability to host AMF with the ability to resist pathogens. Advances in understanding the plant immune system have revealed similarities between pathogen infection and AM colonization that may lead to trade-offs between symbiosis and disease resistance. This review considers the potential trade-offs between AM colonization, agronomic traits and disease resistance and highlights the need for translational research to apply fundamental knowledge to crop improvement.

1. Introduction
Arbuscular mycorrhizal (AM) symbioses are formed between the majority of land plants and members of the ancient and diverse phylum of fungi, the Glomeromycota [1,2]. This relationship supplies water and nutrients, particularly phosphate [3] but also nitrogen (ammonium) [4] and other metals and salts including zinc [5,6], to the host plant. Arbuscular mycorrhizal fungi (AMF) are obligate biotrophs, relying on living root tissue for carbohydrate supply and to complete their asexual life cycle [7]. Species diversity and variation of AMF differs depending on season [8], biogeographical history and environmental conditions [9]. Nevertheless, the AM condition is normal for plants growing in most field situations [10], with the exception of plant species that are unable to form AM symbioses, such as the Brassicaceae family [1]. AMF can affect plant growth, development and disease resistance, but plant breeding often focuses on direct phenotypic traits rather than the contribution that associated microbes could make to them [11]. The ubiquitous nature of AM interactions and their potential to contribute to crop productivity presents an opportunity for plant breeding.
Phosphate is an important growth-limiting macronutrient for all plants [12]. Plants acquire phosphate via the direct pathway (DP) by transfer of ions from the rhizosphere, or via the AM symbiotic pathway (MP) [13]. In the majority of soils, phosphate supply is limited due to rapid immobilisation by free cations [14]. Consequently, phosphate uptake via the DP exceeds the rate of phosphate diffusion through the soil. This generates a depletion zone around plant roots and limits the supply of phosphate to the plant [12,13,15]. In developed countries, large volumes of phosphate fertilizer are added to agricultural soils [16]. However, phosphate fertilizer is both a limited resource [17] and polluting [18] so reducing its use would benefit the environment. The hyphal network of AMF extends beyond the depletion zone of the DP, bypassing the diffusion limitation and accessing a greater area of soil for phosphate uptake [14] (Figure 1). The symbiosis with AMF could enable more efficient use of phosphate, and breeders are considering their potential for sustainable agriculture [19].
To establish AM colonization, plant roots exude strigolactone hormones which stimulate AMF spore germination, metabolism and hyphal branching [20,21], and the release of lipochitooligosaccharide signalling molecules [22], known as mycorrhizal (Myc) factors, which are recognised by host plant receptors [23]. Signals from host receptors are transmitted through a transcriptional activation pathway common to both AM colonization and colonization by nitrogen-fixing rhizobia [24]. Following chemical and mechanical stimulation from the AMF hyphopodia, plant epidermal cells produce a sub-cellular structure called a pre-penetration apparatus (PPA) [25] which predicts the path of entry by fungal hypha. The hypha travels through root epidermal cells, then passes between the underlying cells to enter inner cortical cells. Once inside cortical cells, the hypha branches repeatedly to form arbuscules, the structures of nutrient exchange [13,26].
AMF provide water and nutrients to their host plants, which would be expected to have a beneficial effect on plant growth. However, many studies have shown varied agronomic responses (biomass and phosphate uptake) of plants colonized by AMF. In many cases, AM-colonized plants exhibit zero or negative growth responses and lower levels of phosphate acquisition [2,27], suggesting a trade-off between AM symbiosis and agronomic traits. Besides nutrient acquisition, benefits of AM symbiosis include improved tolerances to many abiotic (mainly salt and drought) [28,29] and biotic stresses (mainly soil-borne pathogens) [30,31] (Table 1; Figure 1). There are many resemblances between the infection processes of AMF and biotrophic pathogens and the plant immune system initially responds similarly to both [7,32]. It has been proposed that pathogens can recruit the symbiont-response pathway [33,34,35] and therefore it is conceivable that trade-offs between AM symbiosis and disease resistance may also exist. We consider the impact of the AM symbiosis on agronomic traits (plant growth and nutrition responses) and disease resistance, and highlight potential compromises that must be considered for plant breeding. AMF may also have potential for phytoremediation [36,37,38]; however, this review focusses on approaches that could be taken by crop breeding rather that adding AMF to soil.
2. Arbuscular Mycorrhizal Symbiosis and Agronomic Traits
2.1. Positive Effects
Studies in diverse plant species have shown AM symbiosis to increase host growth, for example, in Medicago truncatula Gaertn. [39,40]; tomato (Lycopersicon esculentum Mill.) [41]; flax (Linum usitatissimum L.) [2]; onion (Allium cepa L.) [42]; wheat (Triticum aestivum L.) [43]; barley (Hordeum vulgare L.) [44]; and subterranean clover (Trifolium subterraneum L.) [45]. The arbuscule, which consists of highly branched hyphae surrounded by plant membrane, forms the nutrient exchange interface. The associated plant membrane, called the periarbuscular membrane, is decorated with phosphate transporters, which are typically specific to AM roots, for example StPT3 in potato (Solanum tuberosum L.) [53] and MtPT4 in M. truncatula [54]. These transporters take up phosphate released into the periarbuscular space by the AMF. Increased phosphate uptake during AM symbiosis has been observed in a number of species by quantification of total shoot phosphate [45,46] and by using a split-compartment system and 33P to quantify AM phosphate uptake [2,55,56]. Yang et al. (2012) reported that AM-colonized rice (Oryza sativa L.) receives more than 70% of its phosphate via the MP [57].
Positive growth responses are thought to be largely due to increased phosphate uptake via the MP, but can also arise from increased uptake of other growth-limiting nutrients, for example nitrogen (ammonium) [10,13]. AM symbiosis has been shown to have a positive effect on legume nitrogen-fixation, with higher nitrogen concentrations observed in nodulating AM plants compared to their non-AM counterparts [60,61]. Experiments using split-compartments have also indicated nitrogen uptake via AM hyphae [62,63,64,65], including examples in agronomically important species maize (Zea mays L.) [63] and wheat [65]. An AM-specific ammonium transporter LjAMT2; 2 has been identified in Lotus japonicus (Regel) K. Larsen [66].
AM symbiosis can have a positive effect on abiotic stress tolerance. Increased drought tolerance in AM-colonized plants has been explained by effects on plant water relations [67,68] and improved phosphate nutrition [28]. Suggested mechanisms underlying improved tolerance of AM plants to salt stress include accumulation of soluble sugars in roots [29], maintenance of K+:N+ ratios [69] and enhanced nutrient acquisition [70,71]. AMF have also been shown to reduce symptoms of arsenic toxicity by improved phosphate nutrition and increased activity of compounds, such as glutathione-S-transferase, which transform arsenic into non-toxic forms [72,73]. AM symbiosis has been shown to reduce assimilation of other adverse heavy metals, such as copper, chromium, cadmium and cobalt [19,36,77]. It has been suggested that altered phosphate transport during AM colonization alleviates symptoms of phosphate toxicity in highly-fertilized soils [76]. AMF have also been suggested to reduce host sulphur starvation responses via AM-transport of sulphur-containing compounds [74,75].
2.2. Potential Trade-Offs
Under phosphate limited conditions, the MP generally results in higher phosphate uptake rates than in non-mycorrhizal plants [100,101]. Despite this, AM colonization does not always result in increased plant growth and many studies have shown negative growth effects. Graham and Abbott (2000) grew wheat with ten different AMF species in the glasshouse experiments using field soils and all caused growth depressions [46]. Growth depressions with AM colonization have also been observed in other pot experiments with wheat [43,47,48] and barley [49], and field experiments with tobacco (Nicotiana tabacum L.) [50,51,52]. The reduced growth in these experiments was attributed to a range of factors including “aggressive” AMF species [46], post-transcriptional or post-translational control of the phosphate transport pathway in AM-colonized plants [49], and competition with the host for photosynthates [52].
Bethlenfalvay (1982) showed that AM-colonized soybean (Glycine max (L.) Merr.) displayed early growth depressions that were later overcome [102]. In experiments by Li et al. (2005), AM plants exhibited early growth depressions but were still able to reach their reproductive goals [103]. It could be argued that early growth depressions are not trade-offs because AM plants reached the reproductive stage with lower investment in resources than non-AM plants [58]. However, in some cases, growth depressions are very detrimental to the host plant. “Tobacco stunt disease” is thought to be due to colonization by AMF, Glomus macrocarpum Tul. & C. Tul., although the authors note that experiments do not eliminate the possibility that G. macrocarpum is a vector for another pathogen. In these experiments, AM-colonized tobacco plants exhibited growth depressions of between 62–74% compared to their non-AM counterparts [52]. These cases suggest a trade-off between AM colonization and host growth response [51,52].
The key drivers of the symbiosis are generally accepted to be the exchange of plant photosynthate for fungal phosphate, so it seems likely that the ratio of carbon provision to phosphate acquisition is central to host success. Indeed, Tinker (1975) suggested that growth depressions arise if the photosynthetic carbon cost to the plant exceeds the benefit in growth acquired from increased phosphate uptake [104]. An important aspect of this is the relationship between phosphate acquisition by the DP vs the MP. It was previously assumed that the MP contributed extra phosphate to AM plants and the DP was unchanged by colonization [10,58]. However, phosphate uptake via the DP and MP is not completely additive [2,27]. During AM colonization, the contribution of the DP to phosphate uptake is reduced and, depending on levels of available phosphate and the genotypes of the host and AMF, the MP can fail to compensate for this, resulting in overall reduced phosphate uptake [2,47,58,59]. Phosphate transporters acting in the DP were shown to be downregulated during AM colonization in some studies [41,105,106] but not others [49,107]. The negative relationship between MP and DP may also explain AM-induced depressions in plant growth which have often been attributed to fungal “cheats”, i.e., AMF species that do not provide adequate phosphate in exchange for host plant carbon [108,109]. It seems possible that these depressions are not due to a lack of phosphate from the MP, but are in fact because the MP and DP are partially exclusive. Smith, Grace and Smith (2009) suggest that utilizing AM symbiosis in sustainable agriculture will necessitate making the DP and MP more additive rather than partly complimentary [58].
3. Arbuscular Mycorrhizal Symbiosis and Disease Resistance
3.1. Positive Effects
AM symbiosis has been shown to increase host resistance to a wide range of fungal and bacterial pathogens [78,79,80,81,82,83,84,85,110,111,112], particularly root pathogens. The AMF species Funneliformis mosseae (T.H. Nicholson @ Gerd.) C. Walker & A. Schüβler and Rhizophagus irregularis (Blaszk, Wubert, Renker & Buscot) C. Walker & A. Schüβler (formerly Glomus intraradices N.C. Schenk & G.S. Sm.) have been shown to alleviate pea root rot caused by Aphanomyces euteiches Drechsler [30,78]. Damping-off in pea (Pisum sativum L.) seedlings caused by Pythium ultimum Trow was also reduced with AM colonization [81]. Borowicz (2001) performed a meta-analysis and showed that AM-colonization reduced growth of root pathogens (fungal pathogens and nematodes) in 65 out of 125 experiments [112].
Enhanced disease resistance in AM plants was also previously attributed to increased nutrient status. However, although this may contribute, it seems unlikely to be the sole factor involved because AM colonization is not necessarily correlated with nutrient uptake [2,47,58,59]. It has also been suggested that AM-induced reduction in root pathogens is due to direct competition for root space and resources [79]. However, in split-compartment experiments, AM-colonized tomato plants exhibited lower root infection by Phytophthora parasitica Dastur both locally and in non-AM compartments [80], contradicting the suggestion that root resistance is due to direct competition. In addition, AM plants can have increased resistance to foliar pathogens, for example Magnaporthe oryzae B.C. Couch in rice [82], Alternaria solani (Ellis & G. Martin) L.R. Jones & Grout in tomato [83] and Botrytis cinerea Pers. in rose (Rosa rugosa L.) and tomato [84,85], further suggesting that systemic mechanisms are involved in AM-induced immunity. Indeed, the induction of resistance mechanisms by AMF is now thought to generate long-lasting systemic acquired resistance (SAR)-like priming of SA-dependent genes [87,93], and preconditioning of JA-dependent defences [87,88,89,90,94,95] and cell wall defences [80,96,97]. During challenge of tomato by P. parasitica, host cell wall thickenings containing non-esterified pectins and pathogenesis-related protein 1 (PR-1) were observed in non-AM compartments of AM-colonized plants but not in non-AM control plants. AM-colonized plants show enhanced production of defence-compounds such as phenolics [98], β-1,3-glucanase [96] and chitinolytic enzymes [99]. Furthermore, Pozo et al. (1999) showed evidence of systemic priming where particular isoforms of β-1,3-glucanase were upregulated during pathogen challenge only in AM plants [97].
The dynamics and mechanisms of AM-induced resistance have been studied. It is generally accepted that AMF trigger transient induction of plant defences, followed by local suppression at the later stages of interaction. Pieterse et al. (2012) suggest that host plants initially treat AMF as potential invaders, activating defence programs that are later downregulated to allow colonization [32].
3.2. Potential Trade-Offs
The commonalities between pathogen infection and AMF colonisation are likely to be central to any potential trade-offs (Figure 2). There are many similarities between the infection processes of AMF and biotrophic pathogens. Biotrophic pathogens establish a long-term relationship with their host, resulting in nutrient assimilation via specialised intracellular infection structures called haustoria [113] while AMF form analogous structures, called arbuscules. Both structures are bound by host plasma membrane. Such intimate associations between fungus and plant can be expected to engage the host-immune system. Plants respond to pathogen infection using a two-part innate immune system [114]. The first line of plant defence involves recognition of pathogen- (or microbe-) associated molecular patterns (PAMPs/MAMPs), for example chitin, which trigger general plant defence responses, including rapid ion fluxes across the plasma membrane; MAP kinase activation; production of reactive-oxygen species; rapid changes in gene expression; and cell wall reinforcement [115]. This response is known as PAMP-triggered immunity (PTI). Pathogens can supress components of PTI by delivering effector proteins into the plant. The second element of plant immunity involves the recognition of these effectors by resistance (R-) genes. This generates a stronger resistance response, known as effector-triggered immunity (ETI) [116]. R-genes and their products control a broad set of disease resistance responses and usually lead to a hypersensitive cell death response (HR) at the infection site [114]. Before the establishment of symbiosis, elements of this plant immune response are used to differentiate between symbiont and pathogen. It is possible that this could be exploited by pathogens, resulting in trade-offs between symbiosis and disease resistance.
Transcriptomic studies suggest a large overlap in host responses to AMF vs fungal pathogens. Güimil et al. (2005) suggest that up to 40% of the symbiotic machinery recruited for AM symbiosis may be shared with pathogenic fungal interactions [34]. Their comparative genomic study of rice examined the expression of 224 AM-related genes during interactions with AMF and pathogens Magnaporthe grisea (T.T. Herbert) M.E. Barr and Fusarium moniliforme J. Sheld. Thirty out of 224 AM-related genes were shown to be differentially regulated in a similar way and 95 out of 224 responded similarly during both pathogenic and symbiotic interactions. Furthermore, similar transcriptome profiles have been observed in cells containing pathogen haustoria and AM arbuscules [117]. The observed similarity at the level of gene expression reflects the many parallels between fungal symbionts and pathogens. The similarities between biotrophic pathogen infection and AMF colonization are presumably based on similar requirements for host accommodation. It can be predicted that these commonalities will be strongest in the early stages of the interaction before the onset of disease, when the AMF and pathogen have shared requirements for accommodation such as, the attenuation of host defences, establishment of intracellular feeding structures and nutrient acquisition. These shared host responses need to be considered when breeding for AM interactions or against biotrophic pathogens. Although mechanistic details of the overlap in host requirements between pathogenic and AMF are mostly unknown, some genes with potential roles in both types of interactions have been identified.
Like all fungi, AMF have cell walls containing chitin which is readily recognized by the plant immune system [118]. Chitin oligosaccharides act as PAMPs that promote plant immunity via Lysine Motif Receptor-like Kinases (LysM-RLK). Some LysM-RLKs, such as Chitin Elicitor Receptor Kinase 1 (CERK1) have a dual role in pathogenic and symbiotic interactions [119] (Figure 2). CERK1 plays a central role in PTI against fungal pathogens and has been shown to be required for the recognition of chitin in both Arabidopsis thaliana (L.) Heynh and rice [120,121]. Carotenuto et al. (2017) demonstrated that the Oscerk1 mutant is unable to host AMF, therefore OsCERK1 is required for perception of chitin or related molecules to promote AM symbiosis [122]. In rice, CERK1 interacts with Chitin Elicitor Binding Protein (CEBiP), which lacks the kinase signalling domain, to perceive long-chain chitin oligomers during the host defence signalling pathway. However, mutants in OsCEBiP are only defective in the host immune response and can still be colonized by AMF [122]. These studies highlight the necessity to understand the overlaps and differences between AMF and pathogen signalling pathways if breeders are to improve AM interactions without compromising PTI.
Effector proteins play a key role in pathogen colonization by controlling the plant immune system. AMF secrete effectors, similar to those produced by pathogens during infection [123,124]. One of these, SP7, an effector from R. irregularis, downregulates host PTI via interaction with ERF19, a host ethylene-responsive transcription factor that regulates the expression of several defence-related genes in M. truncatula [123]. Sędzielewska Toro and Brachmann (2016) used a bioinformatics pipeline to predict the repertoire of effector proteins in two AMF species, R. irregularis and Rhizophagus clarus (T.H. Nicolson & N.C. Schenck) C. Walker & A. Schüβler [125]. They found candidate effectors with potential roles in signal transduction (eight candidate effectors); cell wall modification (four candidate effectors); and transcriptional regulation (one candidate effector); suggesting that the use of effectors to modulate host responses is of general importance in AM interactions.
One apparent overlap between the host-requirements of AMF and biotrophic fungi is the acquisition of lipids. AMF are lipid auxotrophs, and accordingly, lipids are transferred from the host plant to AMF to sustain AM colonization [126,127,128], which requires action of the glycerol-3-phosphate acyltransferase enzyme Required for Arbuscular Mycorrhization 2 (RAM2) [129]. Jiang et al. (2017) showed that plant fatty acids are also transferred to the powdery mildew pathogen, Golovinomyces cichoracerum (DC.) V.P. Heluta, suggesting that AMF and G. cichoracerum similarly recruit the fatty acid biosynthesis program to facilitate host colonization/invasion (Figure 2) [91]. Mutations in RAM2 result in decreased levels of AM colonization associated with reduced hyphopodium formation [35,130]. RAM2 has also been shown to be required for appressorium formation by the pathogen, Phytophthora palmivora [35]. Wang et al. (2012) suggest that P. palmivora has hijacked the preexisting AM-symbiotic signaling pathway to manipulate their host metabolism, via RAM2 and the production of cutin to promote appressorium formation [35]. However, these findings have been contested [131] and the biological relevance is uncertain since the pathogen/host combination studied (P. palmivora-M. truncatula) does not occur naturally. Gobbato et al. (2013) have shown that RAM2 is required during infection by root pathogen, A. euteiches [92]. However, in this case, there does not seem to be a trade-off with AM-colonization and A. eutiches infection; previous studies have shown AM-colonized plants to be more resistant to A. euteiches, or no correlation between AM colonization and A. euteiches infection [132].
MILDEW RESISTANCE LOCUS O is a gene conferring susceptibility to the biotrophic pathogen, powdery mildew [133]. Barley mlo mutants are resistant to powdery mildew infection by Blumeria graminis f.sp. hordei (DC.) Speer. The role of MLO in powdery mildew resistance was first described in 1942 and has since been mutagen-induced for use in barley breeding [133]. MLO is a modulator of defence and cell death [134]. The expression of the MLO gene is induced by powdery mildew [134], suggesting that the fungi may target MLO to subvert defence pathways and facilitate infection. There are many documented fitness costs and trade-offs associated with disease-resistant plants, including trade-offs linked to mlo mutants [135]. Ruiz-Lozano et al. (1999) examined AM colonization and found a reduction in arbuscule frequency in mlo mutants [86]. There are various similarities between the infection mechanisms of powdery mildew and AMF. Phylogenomic studies have shown an MLO family member to be conserved for AM symbiosis [136]. These findings suggest that MLO is shared by the symbiont-response and powdery mildew infection pathways (Figure 2). This potential trade-off between AM symbiosis and powdery mildew infection should be researched further when considering the utilisation of AMF in agriculture.
Other apparent overlaps in host accommodation requirements of AMF and pathogens have been identified. Examination of the transcriptome of AMF R. irregularis showed that it has lost the thiamine biosynthetic pathway [117], as have most haustorium-forming pathogenic fungi [137]. This implies that in both cases thiamine is sourced from the host, and raises the possibility that common mechanisms could be involved in provision of thiamine to the fungi. Compared to non-biotrophic fungi, several obligate biotrophic pathogens and AM symbionts also have a decreased repertoire of carbohydrates involved in degradation of plant cell wall polysaccharides [117,138,139]. This suggests that the cell wall remodelling required for formation of intracellular structures (i.e., haustoria and arbuscules) may be host-determined [117], opening the possibility that common mechanisms could be involved. Both bacterial and fungal root symbioses also share signal-transduction components with root-knot nematodes, suggesting recruitment of part of the symbiont-response pathway to enhance their parasitic ability [33]. These studies indicate that more research is required regarding the conserved and divergent mechanisms between the recruitment of symbionts and infection by pathogens to determine whether these shared genes could lead to trade-offs between symbiosis and disease resistance. Further identification of genes shared in pathogenic and symbiotic interactions would be valuable for crop research in order to benefit the formation of AM symbiosis without compromising disease resistance mechanisms.
4. Perspectives for Research and Crop Breeding
AMF interactions have benefits for assimilating nutrients, improving disease resistance and increasing abiotic stress tolerance, and so have potential for crop improvement. We consider here how AMF interactions could be relevant to plant breeding, and how this could be developed through translational research.
4.1. Growth Responses
It is clear that AM-induced growth responses are not straightforward and there are possible trade-offs between the phosphate acquisition via the MP and the DP. Smith and Smith (2011) suggest that, if molecular mechanisms underlying lower DP contributions in AM plants can be understood, it may be possible to eliminate them thereby making MP and DP contributions additive to sustainably increase phosphate uptake efficiency [10].
Hetrick, Wilson and Cox (1992) compared varieties of wheat developed before and after 1990 and suggested that varieties developed before 1990 were more responsive to AM colonization than those developed later [43,48]. They proposed that plant breeding under high nutrient conditions has selected against the ability to form AM symbiosis. However, Sawers et al. (2008) suggest that the observed differences in this study rather reflected increased DP in modern wheat lines [140]. When phosphate acquisition via the DP is sufficiently high, plants suppress AM symbiosis, a consequence of DP being less costly than MP. They argue that breeding has selected for a reduction in dependence on AMF, not a loss of compatibility and therefore modern cultivars have retained the ability to form AM symbioses.
Some plant and AMF combinations are more productive than others [109], further complicating the perspectives for crop breeding. The nutrient status of soils also affects the species composition of AMF and success for the symbiotic interaction. Johnson (1993) examined fungal structures formed by AMF from fertilized soil and unfertilized soil and found that AMF from fertilized soil exerted a higher net carbon cost on their host than the AMF from unfertilized soil [141]. This suggests that high nutrient status in soils selects for AMF that could be inferior symbionts. Positive agronomic responses and possible trade-offs must be considered when assessing the potential benefits of AMF for sustainable agriculture. Furthermore, it has been suggested that breeders need to think beyond AM-influenced agronomic traits and towards systemic traits, for example AM-induced disease resistance [11].
4.2. Disease Resistance
The majority of studies regarding AMF and disease resistance have suggested that AMF have a positive effect, for example the SAR-like priming of defence responses [80,90,94,95,96,97]. Genotype-dependent host phenotypes for AM-mediated growth responses have been observed [43,48,109,142]. Yet, less is known about genotypic variation in AM-induced disease resistance. Some studies have shown cultivar-specific defence effects of AM colonization [143,144]. However, the mechanisms responsible for genotypic variation in AM-mediated disease resistance are not fully understood. Borowicz (2001) found that plants exhibited a greater increase in AM-mediated growth response whilst under challenge by fungal pathogens and nematodes [112]. This further highlights the need to consider AM symbiosis in the context of disease resistance to exploit potential benefits for crop breeding.
Despite a general consensus that AMF have potential to improve disease resistance, it is clear that before this can be factored into breeding programmes, more research is needed into the underlying mechanisms. For example, although AM-induced immunity is beneficial during pathogen challenge, could it be costly in terms of resources in the absence of pathogens? The similarity of intracellular structures formed during interaction with AMF and pathogens, plus similarity of host genetic responses led to the prediction that plants may use overlapping signal-transduction pathways to respond to AMF, rhizobia and pathogens. Transcriptomic studies [34] and identification of CERK1 [119,145] and RAM2 [35] have supported this. However, further research is needed to identify shared components because these could be targets for exploitation by pathogens, for example research into the role of MLO during AM symbiosis.
4.3. Translational Research for Crop Breeding
This review emphasizes the considerable progress made in mechanistic understanding of symbiosis between AMF and plants. Many of the host genes involved in AM symbiosis have been identified. We have also cited examples of how AMF can affect plant growth and stress tolerances. However, there are few examples of where knowledge from fundamental mechanistic research can be used practically in plant breeding programs. This gap between basic and applied research needs to be addressed if AM symbiosis can be effectively used in breeding for crop improvement.
Crop improvement through breeding is based on the selection of preferred traits from very large numbers of individual plants in field trials which are performed in different locations over several years. Gene markers, chips and genomic selection techniques can be used to accelerate the selection of key traits such as yield, disease resistance or quality. Clearly, AM symbiosis can affect crop traits, either directly, through phosphate uptake and growth responses, or indirectly through enhanced stress tolerance and disease resistance. However, the significance of this for variety improvement has not yet been realized. Further translational research is needed before these approaches can be used for selection of positive AM interactions, as described here.
AM colonization is often quantified using microscopy which is time-consuming and sometimes subjective. At present, phenotyping for positive AM-mediated traits is not sufficiently high-throughput for application in crop breeding. However, research on this topic is rapidly developing and could soon be applied to crop improvement. For example, AM colonization phenotyping has been used to identify Quantitative Trait Loci (QTL) for AM symbiosis in wheat using a 90k iSelect gene chip [146]. Such research could enable development of marker-assisted selection techniques for AM symbiosis in crops. In addition, important genes for AM colonization identified from fundamental studies on model plants could also be used in marker-assisted selection. However, these approaches assume that the markers used for selective breeding should be those associated with maximizing AM colonization. We have pointed out that it should not be assumed that colonization has only a beneficial effect on crop traits. In this review, we cited several examples where negative impacts of AM colonization have been reported. Breeders need to look beyond maximizing AM colonization for crop improvement and towards identifying genes for positive AM interactions, for example, genes associated with AM-induced disease resistance and stress tolerance. Translational research to characterize the positive and negative impacts of AM colonization should enable identification of the most significant genes and QTL for crop improvement.
Much of our understanding of genetic factors involved in AM symbiosis has been developed through studies using model plants. M. truncatula is a good model for researching the molecular basis of AM symbiosis because of its high levels of AM colonization, diploid genetics, small genome, ease of transformation and short lifecycle [147]. However, unlike many crop species, M. truncatula often shows a positive growth response during AM colonization [39,40]. This highlights that caution must be taken when translating research from model species into crops. Dreher et al. (2017) also point out that genetic diversity in AM colonization of M. truncatula is low, making it difficult to use for identification of loci conditioning this trait [132]. Furthermore, there are differences between the AM colonization in dicots and monocots. Gutjahr et al. (2009) showed that, in rice, AMF R. irregularis preferentially colonize and initiate growth of large lateral roots, with fine lateral roots appearing to be free of AM colonization [148]. Additionally, Mukherjee and Ané (2011) demonstrated significant differences between rice and M. truncatula in the genetic control of plant responses to AMF [149]. The differences between AM colonization of dicots and monocots highlights the importance of relevant monocot crop models. AMF are thought to generate SAR responses in their hosts. However, Cameron et al. (2013) comment that most SAR studies have been carried out in the model plant, but AM-incompatible, A. thaliana, making it more difficult to draw comparisons and research AM-induced resistance [150]. Furthermore, basic research often focusses on one or few AMF species, under controlled conditions which are not necessarily representative of environmental conditions relevant for agriculture. This review highlights a need for realistic crop models to bridge the gap between basic and applied research.
AM symbiosis is the natural state for most plants [10]. These symbiotic interactions are part of the normal ecosystem, and plants co-exist with a wide diversity of AMF species in uncultivated soil [8,9,151]. Monocultures and crop cultivation appear to alter AMF species composition and reduce species diversity [152]. The impact of these changes on crop production has not been thoroughly evaluated [153]. However, it is clear that there will be considerable differences between locations, seasons and soil types [8,9]. Verzeaux et al. (2017) review the effects of AM-colonization on crop nitrogen use efficiency and suggest that it is necessary to investigate which agricultural practices could be favorable to maximize the benefits of AMF [154]. Rotations to manage soil health also have a major impact on AMF species diversity [155]. Genomic technologies, such as small subunit (SSU) ribosomal RNA (rRNA) sequencing [8,9,152], now provide a much greater understanding of species diversity in the crop root microbiome and how this could be affected by soil types and rotations. Further research on the crop root microbiome could help establish how robust the crop–AMF interaction is in different soil types. Such research could be particularly relevant for low-input systems and developing countries.
5. Conclusions
Beneficial AM interactions have the potential to contribute to improved crop productivity through enhanced nutrient uptake, disease resistance and stress tolerance. There are also several examples of where AMF can also have negative impacts on crop performance, and further research is needed to understand the basis for this. Current research on the molecular basis of plant immunity and the conserved and divergent mechanisms between the recruitment of symbionts and infection by pathogens could help to define gene targets for plant breeding. In addition, improved understanding of the root microbiome should indicate how this research will help to advise future breeding approaches, targeting positive AM interactions and reducing trade-offs.
Acknowledgments
Catherine Jacott is supported by the Biotechnology and Biological Sciences Research Council (BBSRC) Norwich Research Park Bioscience Doctoral Training Grants BB/J014524/1 and BB/M011216/1. Jeremy Murray is supported by the BBSRC Grants BB/G023832/1 and BB/L010305/1. Christopher Ridout is supported by BBSRC grant BB/P012574/1 for this work. The authors would like to thank Henk-jan Schoonbeek for helpful discussions in the preparation of this manuscript. The authors would like to thank Timothy Hion for help with creating the figures.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Wang, B.; Qiu, Y.-L. Phylogenetic distribution and evolution of mycorrhizas in land plants. Mycorrhiza 2006, 16, 299–363. [Google Scholar] [CrossRef] [PubMed]
- Smith, S.E.; Smith, F.A.; Jakobsen, I. Functional diversity in arbuscular mycorrhizal (AM) symbioses: The contribution of the mycorrhizal P uptake pathway is not correlated with mycorrhizal responses in growth or total P uptake. New Phytol. 2004, 162, 511–524. [Google Scholar] [CrossRef]
- Cooper, K.M.; Tinker, P.B. Translocation and transfer of nutrients in vesicular arbuscular mycorrhizas: II. Uptake and translocation of phosphorus, zinc and sulphur. New Phytol. 1978, 81, 43–52. [Google Scholar] [CrossRef]
- Govindarajulu, M.; Pfeffer, P.E.; Jin, H.; Abubaker, J.; Douds, D.D.; Allen, J.W.; Bücking, H.; Lammers, P.J.; Shachar-Hill, Y. Nitrogen transfer in the arbuscular mycorrhizal symbiosis. Nature 2005, 435, 819–823. [Google Scholar] [CrossRef] [PubMed]
- Lambert, D.H.; Baker, D.E.; Cole, H. The role of mycorrhizae in the interactions of phosphorus with zinc, copper and other elements. Soil Sci. Soc. Am. J. 1979, 43, 976–980. [Google Scholar] [CrossRef]
- Watts-Williams, S.J.; Cavagnaro, T.R. Arbuscular mycorrhizas modify tomato responses to soil zinc and phosphorus addition. Biol. Fertil. Soils 2012, 48, 285–294. [Google Scholar] [CrossRef]
- Paszkowski, U. A journey through signaling in arbuscular mycorrhizal symbioses. New Phytol. 2006, 172, 35–46. [Google Scholar] [CrossRef] [PubMed]
- Dumbrell, A.J.; Ashton, P.D.; Aziz, N.; Feng, G.; Nelson, M.; Dytham, C.; Fitter, A.H.; Helgason, T. Distinct seasonal assemblages of arbuscular mycorrhizal fungi revealed by massively parallel pyrosequencing. New Phytol. 2011, 190, 794–804. [Google Scholar] [CrossRef] [PubMed]
- Öpik, M.; Zobel, M.; Cantero, J.J.; Davison, J.; Facelli, J.M.; Hiiesalu, I.; Jairus, T.; Kalwij, J.M.; Koorem, K.; Leal, M.E.; et al. Global sampling of plant roots expands the described molecular diversity of arbuscular mycorrhizal fungi. Mycorrhiza 2013, 23, 411–430. [Google Scholar] [CrossRef] [PubMed]
- Smith, S.E.; Smith, F.A. Roles of arbuscular mycorrhizas in plant nutrition and growth: New paradigms from cellular to ecosystem scales. Annu. Rev. Plant Biol. 2011, 62, 227–250. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hohmann, P.; Messmer, M.M. Breeding for mycorrhizal symbiosis: Focus on disease resistance. Euphytica 2017, 213, 1–11. [Google Scholar] [CrossRef]
- Marschner, H. Mineral Nutrition of Higher Plants; Academic Press: Cambridge, MA, USA, 1995; ISBN 0123849063. [Google Scholar]
- Smith, S.E.; Read, D.J. Mycorrhizal Symbiosis; Elsevier: Amsterdam, The Netherlands, 1997; ISBN 9780126528404. [Google Scholar]
- Tinker, P.B.; Nye, P.H. Solute Movement in the Rhizosphere; Oxford University Press: Oxford, UK, 2000; ISBN 0195352319. [Google Scholar]
- Karandashov, V.; Bucher, M. Symbiotic phosphate transport in arbuscular mycorrhizas. Trends Plant Sci. 2005, 10, 22–29. [Google Scholar] [CrossRef] [PubMed]
- FAO. Use of phosphate rocks for sustainable agriculture. Fertil. Plant Nutr. Bull. 2009, 13. [Google Scholar] [CrossRef]
- Khasawneh, F.E. The Role of Phosphorus in Agriculture, 2nd ed.; American Society of Agronomy: Madison, WI, USA, 1980. [Google Scholar]
- Glendining, M.J.; Dailey, A.G.; Williams, A.G.; Evert, F.K.V.; Goulding, K.W.T.; Whitmore, A.P. Is it possible to increase the sustainability of arable and ruminant agriculture by reducing inputs? Agric. Syst. 2009, 99, 117–125. [Google Scholar] [CrossRef] [Green Version]
- Thirkell, T.J.; Charters, M.D.; Elliott, A.J.; Sait, S.M.; Field, K.J. Are mycorrhizal fungi our sustainable saviours? Considerations for achieving food security. J. Ecol. 2017, 105, 921–929. [Google Scholar] [CrossRef]
- Akiyama, K.; Matsuzaki, K.-I.; Hayashi, H. Plant sesquiterpenes induce hyphal branching in arbuscular mycorrhizal fungi. Nature 2005, 453, 824–827. [Google Scholar] [CrossRef] [PubMed]
- Besserer, A.; Puech-Pagès, V.; Kiefer, P.; Gomez-Roldan, V.; Jauneau, A.; Roy, S.; Portais, J.C.; Roux, C.; Bécard, G.; Séjalon-Delmas, N. Strigolactones stimulate arbuscular mycorrhizal fungi by activating mitochondria. PLoS Biol. 2006, 4, 1239–1247. [Google Scholar] [CrossRef] [PubMed]
- Genre, A.; Chabaud, M.; Balzergue, C.; Puech-Pagès, V.; Novero, M.; Rey, T.; Fournier, J.; Rochange, S.; Bécard, G.; Bonfante, P.; et al. Short-chain chitin oligomers from arbuscular mycorrhizal fungi trigger nuclear Ca2+ spiking in Medicago truncatula roots and their production is enhanced by strigolactone. New Phytol. 2013, 198, 190–202. [Google Scholar] [CrossRef] [PubMed]
- Kosuta, S.; Chabaud, M.; Lougnon, G. A diffusible factor from arbuscular mycorrhizal fungi induces symbiosis-specific MtENOD11 expression in roots of Medicago truncatula. Plant 2003, 131, 952–962. [Google Scholar] [CrossRef] [PubMed]
- Stracke, S.; Kistner, C.; Yoshida, S.; Mulder, L.; Sato, S.; Kaneko, T.; Tabata, S.; Sandal, N.; Stougaard, J.; Szczyglowski, K.; et al. A plant receptor-like kinase required for both bacterial and fungal symbiosis. Nature 2002, 417, 959–962. [Google Scholar] [CrossRef] [PubMed]
- Genre, A.; Chabaud, M.; Timmers, T.; Bonfante, P.; Barker, D.G. Arbuscular mycorrhizal fungi elicit a novel intracellular apparatus in Medicago truncatula root epidermal cells before infection. Plant Cell 2005, 17, 3489–3499. [Google Scholar] [CrossRef] [PubMed]
- Parniske, M. Arbuscular mycorrhiza: The mother of plant root endosymbioses. Nat. Rev. Microbiol. 2008, 6, 763–775. [Google Scholar] [CrossRef] [PubMed]
- Smith, S.E.; Smith, F.A.; Jakobsen, I. Mycorrhizal fungi can dominate phosphate supply to plants irrespective of growth responses. Plant Physiol. 2003, 133, 16–20. [Google Scholar] [CrossRef] [PubMed]
- Nelsen, C.E.; Safir, G.R. Increased drought tolerance of mycorrhizal onion plants caused by improved phosphorus nutrition. Planta 1982, 154, 407–413. [Google Scholar] [CrossRef] [PubMed]
- Feng, G.; Zhang, F.S.; Li, X.L.; Tian, C.Y.; Tang, C.; Rengel, Z. Improved tolerance of maize plants to salt stress by arbuscular mycorrhiza is related to higher accumulation of soluble sugars in roots. Mycorrhiza 2002, 12, 185–190. [Google Scholar] [CrossRef] [PubMed]
- Bødker, L.; Kjøller, R.; Rosendahl, S. Effect of phosphate and the arbuscular mycorrhizal fungus Glomus intraradices on disease severity of root rot of peas (Pisum sativum) caused by Aphanomyces euteiches. Mycorrhiza 1998, 8, 169–174. [Google Scholar] [CrossRef]
- Sikes, B.A. When do arbuscular mycorrhizal fungi protect plant roots from pathogens? Plant Signal. Behav. 2010, 5, 763–765. [Google Scholar] [CrossRef] [PubMed]
- Pieterse, C.M.J.; Van der Does, D.; Zamioudis, C.; Leon-Reyes, A.; Van Wees, S.C.M. Hormonal Modulation of Plant Immunity. Annu. Rev. Cell Dev. Biol. 2012, 28, 489–521. [Google Scholar] [CrossRef] [PubMed]
- Weerasinghe, R.R.; Bird, D.M.; Allen, N.S. Root-knot nematodes and bacterial Nod factors elicit common signal transduction events in Lotus japonicus. Proc. Natl. Acad. Sci. USA 2005, 102, 3147–3152. [Google Scholar] [CrossRef] [PubMed]
- Güimil, S.; Chang, H.-S.; Zhu, T.; Sesma, A.; Osbourn, A.; Roux, C.; Ioannidis, V.; Oakeley, E.J.; Docquier, M.; Descombes, P.; et al. Comparative transcriptomics of rice reveals an ancient pattern of response to microbial colonization. Proc. Natl. Acad. Sci. USA 2005, 102, 8066–8070. [Google Scholar] [CrossRef] [PubMed]
- Wang, E.; Schornack, S.; Marsh, J.F.; Gobbato, E.; Schwessinger, B.; Eastmond, P.; Schultze, M.; Kamoun, S.; Oldroyd, G.E.D. A common signaling process that promotes mycorrhizal and oomycete colonization of plants. Curr. Biol. 2012, 22, 2242–2246. [Google Scholar] [CrossRef] [PubMed]
- Ferrol, N.; Tamayo, E.; Vargas, P. The heavy metal paradox in arbuscular mycorrhizas: From mechanisms to biotechnological applications. J. Exp. Bot. 2016, 67, 6253–6265. [Google Scholar] [CrossRef] [PubMed]
- Coninx, L.; Martinova, V.; Rineau, F. Mycorrhiza-assisted phytoremediation. Adv. Bot. Res. 2017, 83, 127–188. [Google Scholar] [CrossRef]
- Rajtor, M.; Piotrowska-Seget, Z. Prospects for arbuscular mycorrhizal fungi (AMF) to assist in phytoremediation of soil hydrocarbon contaminants. Chemosphere 2016, 162, 105–116. [Google Scholar] [CrossRef] [PubMed]
- Feddermann, N.; Boller, T.; Salzer, P.; Elfstrand, S.; Wiemken, A.; Elfstrand, M. Medicago truncatula shows distinct patterns of mycorrhiza-related gene expression after inoculation with three different arbuscular mycorrhizal fungi. Planta 2008, 227, 671–680. [Google Scholar] [CrossRef] [PubMed]
- Monzon, A.; Azcón, R. Relevance of mycorrhizal fungal origin and host plant genotype to inducing growth and nutrient uptake in Medicago species. Agric. Ecosyst. Environ. 1996, 60, 9–15. [Google Scholar] [CrossRef]
- Burleigh, S.H.; Cavagnaro, T.; Jakobsen, I. Functional diversity of arbuscular mycorrhizas extends to the expression of plant genes involved in P nutrition. J. Exp. Bot. 2002, 53, 1593–1601. [Google Scholar] [CrossRef] [PubMed]
- Hayman, D.S.; Mosse, B. Plant growth responses to vesicular-arbuscular mycorrhiza III. Increased uptake of labile P from soil. New Phytol. 1972, 71, 41–47. [Google Scholar] [CrossRef]
- Hetrick, B.A.D.; Wilson, G.W.T.; Cox, T.S. Mycorrhizal dependence of modern wheat-varieties, landraces, and ancestors. Can. J. Bot. Can. Bot. 1992, 70, 2032–2040. [Google Scholar] [CrossRef]
- Wani, S.P.; McGill, W.B.; Tewari, J.P. Mycorrhizal and common root-rot infection, and nutrient accumulation in barley grown on breton loam using N from biological fixation or fertilizer. Biol. Fertil. Soils 1991, 12, 46–54. [Google Scholar] [CrossRef]
- Abbott, L.K.; Robson, A.D. Growth of subterranean clover in relation to formation of endomycorrhizas by introduced and indigenous fungi in a field soil. New Phytol. 1978, 81, 575–585. [Google Scholar] [CrossRef]
- Graham, J.H.; Abbott, L.K. Wheat responses to aggressive and non-aggressive arbuscular mycorrhizal fungi. Plant Soil 2000, 220, 207–218. [Google Scholar] [CrossRef]
- Li, H.; Smith, S.E.; Holloway, R.E.; Zhu, Y.; Smith, F.A. Arbuscular mycorrhizal fungi contribute to phosphorus uptake by wheat grown in a phosphorus-fixing soil even in the absence of positive growth responses. New Phytol. 2006, 172, 536–543. [Google Scholar] [CrossRef] [PubMed]
- Hetrick, B.A.D.; Wilson, G.W.T.; Cox, T.S. Mycorrhizal dependence of modern wheat cultivars and ancestors: A synthesis. Can. J. Bot. 1993, 71, 512–518. [Google Scholar] [CrossRef]
- Grace, E.J.; Cotsaftis, O.; Tester, M.; Smith, F.A.; Smith, S.E. Arbuscular mycorrhizal inhibition of growth in barley cannot be attributed to extent of colonization, fungal phosphorus uptake or effects on expression of plant phosphate transporter genes. New Phytol. 2009, 181, 938–949. [Google Scholar] [CrossRef] [PubMed]
- Guo, B.Z.; An, Z.Q.; Hendrix, J.W. A mycorrhizal pathogen (Glomus macrocarpum Tul. & Tul.) of tobacco: Effects of long- and short-term cropping on the mycorrhizal fungal community and stunt disease. Appl. Soil Ecol. 1994, 1, 269–276. [Google Scholar] [CrossRef]
- Modjo, H.; Hendrix, J.W.; Nesmith, W. Mycorrhizal fungi in relation to control of tobacco stunt disease with soil fumigants. Soil Biol. Biochem. 1987, 19, 289–295. [Google Scholar] [CrossRef]
- Modjo, H.S.; Hendrix, J.W. The mycorrhizal fungus Glomus macrocarpum as a cause of tobacco stunt disease. Phytopathology 1986, 76, 688–691. [Google Scholar] [CrossRef]
- Rausch, C.; Daram, P.; Brunner, S.; Jansa, J.; Laloi, M.; Leggewie, G.; Amrhein, N.; Bucher, M. A phosphate transporter expressed in arbuscule-containing cells in potato. Nature 2001, 414, 462–470. [Google Scholar] [CrossRef] [PubMed]
- Harrison, M.J.; Dewbre, G.R.; Liu, J.Y. A phosphate transporter from Medicago truncatula involved in the acquisiton of phosphate released by arbuscular mycorrhizal fungi. Plant Cell 2002, 14, 2413–2429. [Google Scholar] [CrossRef] [PubMed]
- Sawers, R.J.H.; Svane, S.F.; Quan, C.; Grønlund, M.; Wozniak, B.; Gebreselassie, M.N.; González-Muñoz, E.; Chávez Montes, R.A.; Baxter, I.; Goudet, J.; et al. Phosphorus acquisition efficiency in arbuscular mycorrhizal maize is correlated with the abundance of root-external hyphae and the accumulation of transcripts encoding PHT1 phosphate transporters. New Phytol. 2017, 214, 632–643. [Google Scholar] [CrossRef] [PubMed]
- Smith, S.E.; Manjarrez, M.; Stonor, R.; McNeill, A.; Smith, F.A. Indigenous arbuscular mycorrhizal (AM) fungi contribute to wheat phosphate uptake in a semi-arid field environment, shown by tracking with radioactive phosphorus. Appl. Soil Ecol. 2015, 96, 68–74. [Google Scholar] [CrossRef]
- Yang, S.-Y.; Grønlund, M.; Jakobsen, I.; Grotemeyer, M.S.; Rentsch, D.; Miyao, A.; Hirochika, H.; Kumar, C.S.; Sundaresan, V.; Salamin, N.; et al. Nonredundant regulation of rice arbuscular mycorrhizal symbiosis by two members of the PHOSPHATE TRANSPORTER1 gene family. Plant Cell 2012, 24, 4236–4251. [Google Scholar] [CrossRef] [PubMed]
- Smith, F.A.; Grace, E.J.; Smith, S.E. More than a carbon economy: Nutrient trade and ecological sustainability in facultative arbuscular mycorrhizal symbioses. New Phytol. 2009, 182, 347–358. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Smith, F.A.; Dickson, S.; Holloway, R.E.; Smith, S.E. Plant growth depressions in arbuscular mycorrhizal symbioses: Not just caused by carbon drain? New Phytol. 2008, 178, 852–862. [Google Scholar] [CrossRef] [PubMed]
- Barea, J.M.; Azcon-Aguilar, C. Mycorrhizas and their significance in nodulating nitrogen-fixing plants. Adv. Agron. 1983, 36, 1–54. [Google Scholar] [CrossRef]
- Azcon, R.; Rubio, R.; Barea, J.M. Selective interactions between different species of mycorrhizal fungi and rhizobium-meliloti strains, and their effects on growth, N2-fixation (N-15) and nutrition of Medicago-sativa-L. New Phytol. 1991, 117, 399–404. [Google Scholar] [CrossRef]
- Ames, R.N.; Reid, C.P.P.; Porter, L.K.; Cambardella, C. Hyphal uptake and transport of nitrogen from two 15N-labelled sources by glomus mosseae, a vesicular-arbuscular mycorrhizal fungus. New Phytol. 1983, 95, 381–396. [Google Scholar] [CrossRef]
- Frey, B.; Schüepp, H. Acquisition of nitrogen by external hyphae of arbuscular mycorrhizal fungi associated with Zea mays L. New Phytol. 1993, 124, 221–230. [Google Scholar] [CrossRef]
- George, E.; Haussler, K.-W.; Vetterlein, D.; Gorgus, E.; Marschner, H. Water and nutrient translocation by hyphae of Glomus mosseae compartment. Can. J. Bot. 1992, 70, 2130–2137. [Google Scholar] [CrossRef]
- Hawkins, H. Reduced15N-nitrogen transport through arbuscular mycorrhizal hyphae to Triticum aestivum L. supplied with ammonium vs. nitrate nutrition. Ann. Bot. 2001, 87, 303–311. [Google Scholar] [CrossRef]
- Guether, M.; Neuhauser, B.; Balestrini, R.; Dynowski, M.; Ludewig, U.; Bonfante, P. A mycorrhizal-specific ammonium transporter from Lotus japonicus acquires nitrogen released by arbuscular mycorrhizal fungi. Plant Physiol. 2009, 150, 73–83. [Google Scholar] [CrossRef] [PubMed]
- Augé, R.M. Water relations, drought and vesicular-arbuscular mycorrhizal symbiosis. Mycorrhiza 2001, 11, 3–42. [Google Scholar] [CrossRef]
- Ruiz-Lozano, J.M.; Azcon, R.; Gomez, M. Effects of arbuscular-mycorrhizal glomus species on drought tolerance: Physiological and nutritional plant responses. Appl. Environ. Microbiol. 1995, 61, 456–460. [Google Scholar] [PubMed]
- Giri, B.; Kapoor, R.; Mukerji, K.G. Improved tolerance of Acacia nilotica to salt stress by arbuscular mycorrhiza, Glomus fasciculatum may be partly related to elevated K/Na ratios in root and shoot tissues. Microb. Ecol. 2007, 54, 753–760. [Google Scholar] [CrossRef] [PubMed]
- Al-Karaki, G.; Al-Raddad, A. Effects of arbuscular mycorrhizal fungi and drought stress on growth and nutrient uptake of two wheat genotypes differing in drought resistance. Mycorrhiza 1997, 7, 83–88. [Google Scholar] [CrossRef]
- Evelin, H.; Kapoor, R.; Giri, B. Arbuscular mycorrhizal fungi in alleviation of salt stress: A review. Ann. Bot. 2009, 104, 1263–1280. [Google Scholar] [CrossRef] [PubMed]
- Ultra, V.U.; Tanaka, S.; Sakurai, K.; Iwasaki, K. Effects of arbuscular mycorrhiza and phosphorus application on arsenic toxicity in sunflower (Helianthus annuus L.) and on the transformation of arsenic in the rhizosphere. Plant Soil 2007, 290, 29–41. [Google Scholar] [CrossRef]
- Sharma, S.; Anand, G.; Singh, N.; Kapoor, R. Arbuscular mycorrhiza augments arsenic tolerance in wheat (Triticum aestivum L.) by strengthening antioxidant defense sstem and thiol metabolism. Front. Plant Sci. 2017, 8, 1–21. [Google Scholar] [CrossRef] [PubMed]
- Allen, J.W.; Shachar-Hill, Y. Sulfur transfer through an arbuscular mycorrhiza. Plant Physiol. 2009, 149, 549–560. [Google Scholar] [CrossRef] [PubMed]
- Sieh, D.; Watanabe, M.; Devers, E.A.; Brueckner, F.; Hoefgen, R.; Krajinski, F. The arbuscular mycorrhizal symbiosis influences sulfur starvation responses of Medicago truncatula. New Phytol. 2013, 197, 606–616. [Google Scholar] [CrossRef] [PubMed]
- Kariman, K.; Barker, S.J.; Finnegan, P.M.; Tibbett, M. Ecto- and arbuscular mycorrhizal symbiosis can induce tolerance to toxic pulses of phosphorus in jarrah (Eucalyptus marginata) seedlings. Mycorrhiza 2014, 24. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rezvani, M.; Ardakani, M.R.; Rejali, F.; Zaefarian, F.; Teimouri, S.; Noormohammadi, G.; Miransari, M. Uptake of heavy metals by mycorrhizal barley (Hordeum vulgare L.). J. Plant Nutr. 2014, 38, 904–919. [Google Scholar] [CrossRef]
- Slezack, S.; Dumas-Gaudot, E.; Rosendahl, S.; Kjøller, R.; Paynot, M.; Negrel, J.; Gianinazzi, S. Endoproteolytic activities in pea roots inoculated with the arbuscular mycorrhizal fungus Glomus mosseae and/or Aphanomyces euteiches in relation to bioprotection. New Phytol. 1999, 142, 517–529. [Google Scholar] [CrossRef]
- Bødker, L.; Kjøller, R.; Kristensen, K.; Rosendahl, S. Interactions between indigenous arbuscular mycorrhizal fungi and Aphanomyces euteiches in field-grown pea. Mycorrhiza 2002, 12, 7–12. [Google Scholar] [CrossRef] [PubMed]
- Cordier, C.; Pozo, M.J.; Barea, J.M.; Gianinazzi, S.; Gianinazzi-Pearson, V. Cell defense responses associated with localized and systemic resistance to Phytophthora parasitica induced in tomato by an arbuscular mycorrhizal fungus. Mol. Plant-Microbe Interact. 1998, 11, 1017–1028. [Google Scholar] [CrossRef]
- Rosendahl, C.N.; Rosendahl, S. The role of vesicular-arbuscular mycorrhiza in controlling damping-off and growth reduction in cucumber caused by pythium-ultimum. Symbiosis 1990, 9, 363–366. [Google Scholar]
- Campos-Soriano, L.; García-Martínez, J.; Segundo, B.S. The arbuscular mycorrhizal symbiosis promotes the systemic induction of regulatory defence-related genes in rice leaves and confers resistance to pathogen infection. Mol. Plant Pathol. 2012, 13, 579–592. [Google Scholar] [CrossRef] [PubMed]
- Fritz, M.; Jakobsen, I.; Lyngkjær, M.F.; Thordal-Christensen, H.; Pons-Kühnemann, J. Arbuscular mycorrhiza reduces susceptibility of tomato to Alternaria solani. Mycorrhiza 2006, 16, 413–419. [Google Scholar] [CrossRef] [PubMed]
- Møller, K.; Kristensen, K.; Yohalem, D.; Larsen, J. Biological management of gray mold in pot roses by co-inoculation of the biocontrol agent Ulocladium atrum and the mycorrhizal fungus Glomus mosseae. Biol. Control 2009, 49, 120–125. [Google Scholar] [CrossRef]
- Pozo, M.J.; Jung, S.C.; Lopez-Raez, J.A.; Azcón-Aguilar, C. Impact of arbuscular mycorrhizal symbiosis on plant response to biotic stress: The role of plant defence mechanisms. In Arbuscular Mycorrhizas: Physiology and Function; Springer: Dordrecht, The Netherlands, 2010; pp. 193–2007. ISBN 978-90-481-9488-9. [Google Scholar]
- Ruiz-Lozano, J.M.; Gianinazzi, S.; Gianinazzi-Pearson, V. Genes involved in resistance to powdery mildew in barley differentially modulate root colonization by the mycorrhizal fungus Glomus mosseae. Mycorrhiza 1999, 9, 237–240. [Google Scholar] [CrossRef]
- Pozo, M.J.; Azcón-Aguilar, C. Unraveling mycorrhiza-induced resistance. Curr. Opin. Plant Biol. 2007, 10, 393–398. [Google Scholar] [CrossRef] [PubMed]
- Van der Ent, S.; Van Wees, S.C.M.; Pieterse, C.M.J. Jasmonate signaling in plant interactions with resistance-inducing beneficial microbes. Phytochemistry 2009, 70, 1581–1588. [Google Scholar] [CrossRef] [PubMed]
- Pozo, M.J.; Van Loon, L.C.; Pieterse, C.M.J. Jasmonates—Signals in plant-microbe interactions. J. Plant Growth Regul. 2004, 23, 211–222. [Google Scholar]
- Hao, Z.; Fayolle, L.; Van Tuinen, D.; Chatagnier, O.; Li, X.; Gianinazzi, S.; Gianinazzi-Pearson, V. Local and systemic mycorrhiza-induced protection against the ectoparasitic nematode Xiphinema index involves priming of defence gene responses in grapevine. J. Exp. Bot. 2012, 63, 3657–3672. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Y.; Wang, W.; Xie, Q.; Liu, N.; Liu, L.; Wang, D.; Zhang, X.; Yang, C.; Chen, X.; Tang, D.; et al. Plants transfer lipids to sustain colonization by mutualistic mycorrhizal and parasitic fungi. Science 2017, 356, 1172–1175. [Google Scholar] [CrossRef] [PubMed]
- Gobbato, E.; Wang, E.; Higgins, G.; Bano, S.A.; Henry, C.; Schultze, M.; Oldroyd, G.E.D. RAM1 and RAM2 function and expression during arbuscular mycorrhizal symbiosis and Aphanomyces euteiches colonization. Plant Signal. Behav. 2013, 8, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Glazebrook, J. Contrasting Mechanisms of Defense Against Biotrophic and Necrotrophic Pathogens. Annu. Rev. Phytopathol. 2005, 43, 205–227. [Google Scholar] [CrossRef] [PubMed]
- Song, Y.Y.; Cao, M.; Xie, L.J.; Liang, X.T.; Zeng, R.S.; Su, Y.J.; Huang, J.H.; Wang, R.L.; Luo, S.M. Induction of DIMBOA accumulation and systemic defense responses as a mechanism of enhanced resistance of mycorrhizal corn (Zea mays L.) to sheath blight. Mycorrhiza 2011, 21, 721–731. [Google Scholar] [CrossRef] [PubMed]
- Jung, S.C.; Martinez-Medina, A.; Lopez-Raez, J.A.; Pozo, M.J. Mycorrhiza-induced resistance and priming of plant defenses. J. Chem. Ecol. 2012, 38, 651–664. [Google Scholar] [CrossRef] [PubMed]
- Pozo, M.J.; Azcón-Aguilar, C.; Dumas-Gaudot, E.; Barea, J.M. B-1,3-glucanase activities in tomato roots inoculated with arbuscular mycorrhizal fungi and/or Phytophthora parasitica and their possible involvement in bioprotection. Plant Sci. 1999, 141, 149–157. [Google Scholar] [CrossRef]
- Pozo, M.J.; Cordier, C.; Dumas-Gaudot, E.; Gianinazzi, S.; Barea, J.M.; Azcón-Aguilar, C. Localized versus systemic effect of arbuscular mycorrhizal fungi on defence responses to Phytophthora infection in tomato plants. J. Exp. Bot. 2002, 53, 525–534. [Google Scholar] [CrossRef] [PubMed]
- López-Ráez, J.A.; Flors, V.; García, J.M.; Pozo, M.J. AM symbiosis alters phenolic acid content in tomato roots. Plant Signal. Behav. 2010, 5, 1138–1140. [Google Scholar] [CrossRef]
- Benhamou, N.; Fortin, J.A.; Hamel, C.; St-Arnaud, M.; Shatilla, A. Resistance responses of mycorrhizal Ri T-DNA-transformed carrot roots to infection by Fusarium oxysporum f.sp. chrysanthemi. Phytopathology 1994, 85, 958–968. [Google Scholar] [CrossRef]
- Haynes, R.J. Effects of liming on phosphate availability in acid soils—A critical review. Plant Soil 1982, 68, 289–308. [Google Scholar] [CrossRef]
- Kumar, M.; Yadav, V.; Kumar, H.; Sharma, R.; Singh, A.; Tuteja, N.; Johri, A.K. Piriformospora indica enhances plant growth by transferring phosphate. Plant Signal. Behav. 2011, 6, 723–725. [Google Scholar] [CrossRef] [PubMed]
- Bethlenfalvay, G.J. Parasitic and mutualistic associations between a mycorrhizal fungus and soybean: Development of the host plant. Phytopathology 1982, 72, 889. [Google Scholar] [CrossRef]
- Li, H.Y.; Zhu, Y.G.; Marschner, P.; Smith, F.A.; Smith, S.E. Wheat responses to arbuscular mycorrhizal fungi in a highly calcareous soil differ from those of clover, and change with plant development and P supply. Plant Soil 2005, 277, 221–232. [Google Scholar] [CrossRef]
- Tinker, P.B. Soil chemistry of phosphorus and mycorrhizal effects on plant growth. In Endomycorrhizas; Sanders, F.E., Mosse, B., Tinker, P.B., Eds.; Academic Press: London, UK, 1975; pp. 353–371. [Google Scholar]
- Paszkowski, U.; Kroken, S.; Roux, C.; Briggs, S.P. Rice phosphate transporters include an evolutionarily divergent gene specifically activated in arbuscular mycorrhizal symbiosis. Proc. Natl. Acad. Sci. USA 2002, 99, 13324–13329. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Trieu, A.T.; Blaylock, L.A.; Harrison, M.J. Cloning and characterization of two phosphate transporters from Medicago truncatula roots: Regulation in response to phosphate and to colonization by arbuscular mycorrhizal (AM) fungi. Mol. Plant Microbe Interact. 1998, 11, 14–22. [Google Scholar] [CrossRef] [PubMed]
- Karandashov, V.; Nagy, R.; Wegmüller, S.; Amrhein, N.; Bucher, M. Evolutionary conservation of a phosphate transporter in the arbuscular mycorrhizal symbiosis. Proc. Natl. Acad. Sci. USA 2004, 101, 6285–6290. [Google Scholar] [CrossRef] [PubMed]
- Fitter, A.H. What is the link between carbon and phosphorus fluxes in arbuscular mycorrhizas? A null hypothesis for symbiotic function. New Phytol. 2006, 172, 3–6. [Google Scholar] [CrossRef] [PubMed]
- Klironomos, J.N. Variation in plant response to native and exotic arbuscular mycorrhizal fungi. Ecology 2003, 84, 2292–2301. [Google Scholar] [CrossRef]
- Azcón-Aguilar, C.; Barea, J. Arbuscular mycorrhizas and biological control of soil-borne plant pathogens—An overview of the mechanisms involved. Mycorrhiza 1996, 457–464. [Google Scholar] [CrossRef]
- Whipps, J.M. Prospects and limitations for mycorrhizas in biocontrol of root pathogens. Can. J. Bot. 2004, 82, 1198–1227. [Google Scholar] [CrossRef]
- Borowicz, V.A. Do arbuscular mycorrhizal fungi alter plant-pathogen interactions. Ecology 2001, 82, 3057–3068. [Google Scholar] [CrossRef]
- Perfect, S.E.; Green, J.R. Infection structures of biotrophic and hemibiotrophic fungal plant pathogens. Mol. Plant Pathol. 2001, 2, 101–108. [Google Scholar] [CrossRef] [PubMed]
- Jones, J.D.G.; Dangl, J.L. The plant immune system. Nature 2006, 444, 323–329. [Google Scholar] [CrossRef] [PubMed]
- Zipfel, C. Pattern-recognition receptors in plant innate immunity. Curr. Opin. Immunol. 2008, 20, 10–16. [Google Scholar] [CrossRef] [PubMed]
- Boyd, L.A.; Ridout, C.; O’Sullivan, D.M.; Leach, J.E.; Leung, H. Plant-pathogen interactions: Disease resistance in modern agriculture. Trends Genet. 2013, 29, 233–240. [Google Scholar] [CrossRef] [PubMed]
- Tisserant, E.; Kohler, A.; Dozolme-Seddas, P.; Balestrini, R.; Benabdellah, K.; Colard, A.; Croll, D.; da Silva, C.; Gomez, S.K.; Koul, R.; et al. The transcriptome of the arbuscular mycorrhizal fungus Glomus intraradices (DAOM 197198) reveals functional tradeoffs in an obligate symbiont. New Phytol. 2012, 193, 755–769. [Google Scholar] [CrossRef] [PubMed]
- Kaku, H.; Nishizawa, Y.; Ishii-Minami, N.; Akimoto-Tomiyama, C.; Dohmae, N.; Takio, K.; Minami, E.; Shibuya, N. Plant cells recognize chitin fragments for defense signaling through a plasma membrane receptor. Proc. Natl. Acad. Sci. USA 2006, 103, 11086–11091. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Dong, W.; Sun, J.; Feng, F.; Deng, Y.; He, Z.; Oldroyd, G.E.D.; Wang, E. The receptor kinase CERK1 has dual functions in symbiosis and immunity signalling. Plant J. 2015, 81, 258–267. [Google Scholar] [CrossRef] [PubMed]
- Miya, A.; Albert, P.; Shinya, T.; Desaki, Y.; Ichimura, K.; Shirasu, K.; Narusaka, Y.; Kawakami, N.; Kaku, H.; Shibuya, N. CERK1, a LysM receptor kinase, is essential for chitin elicitor signaling in Arabidopsis. Proc. Natl. Acad. Sci. USA 2007, 104, 19613–19618. [Google Scholar] [CrossRef] [PubMed]
- Shimizu, T.; Nakano, T.; Takamizawa, D.; Desaki, Y.; Ishii-Minami, N.; Nishizawa, Y.; Minami, E.; Okada, K.; Yamane, H.; Kaku, H.; et al. Two LysM receptor molecules, CEBiP and OsCERK1, cooperatively regulate chitin elicitor signaling in rice. Plant J. 2010, 64, 204–214. [Google Scholar] [CrossRef] [PubMed]
- Carotenuto, G.; Chabaud, M.; Miyata, K.; Capozzi, M.; Takeda, N.; Kaku, H.; Shibuya, N.; Nakagawa, T.; Barker, D.G.; Genre, A. The rice LysM receptor-like kinase OsCERK1 is required for the perception of short-chain chitin oligomers in arbuscular mycorrhizal signaling. New Phytol. 2017, 214, 1440–1446. [Google Scholar] [CrossRef] [PubMed]
- Kloppholz, S.; Kuhn, H.; Requena, N. A secreted fungal effector of glomus intraradices promotes symbiotic biotrophy. Curr. Biol. 2011, 21, 1204–1209. [Google Scholar] [CrossRef] [PubMed]
- Koeck, M.; Hardham, A.R.; Dodds, P.N. The role of effectors of biotrophic and hemibiotrophic fungi in infection. Cell. Microbiol. 2011, 13, 1849–1857. [Google Scholar] [CrossRef] [PubMed]
- Sędzielewska Toro, K.; Brachmann, A. The effector candidate repertoire of the arbuscular mycorrhizal fungus Rhizophagus clarus (supp. data). BMC Genom. 2016, 53, 1689–1699. [Google Scholar] [CrossRef]
- Tisserant, E.; Malbreil, M.; Kuo, A.; Kohler, A.; Symeonidi, A.; Balestrini, R.; Charron, P.; Duensing, N.; Frei dit Frey, N.; Gianinazzi-Pearson, V.; et al. Genome of an arbuscular mycorrhizal fungus provides insight into the oldest plant symbiosis. Proc. Natl. Acad. Sci. USA 2013, 110, 20117–20122. [Google Scholar] [CrossRef] [PubMed]
- Wewer, V.; Brands, M.; Dörmann, P. Fatty acid synthesis and lipid metabolism in the obligate biotrophic fungus Rhizophagus irregularis during mycorrhization of Lotus japonicus. Plant J. 2014, 79, 398–412. [Google Scholar] [CrossRef] [PubMed]
- Tang, N.; San Clemente, H.; Roy, S.; Bécard, G.; Zhao, B.; Roux, C. A survey of the gene repertoire of Gigaspora rosea unravels conserved features among Glomeromycota for obligate biotrophy. Front. Microbiol. 2016, 7. [Google Scholar] [CrossRef] [PubMed]
- Luginbuehl, L.H.; Menard, G.N.; Kurup, S.; Van Erp, H.; Radhakrishnan, G.V.; Breakspear, A.; Oldroyd, G.E.D.; Eastmond, P.J. Fatty acids in arbuscular mycorrhizal fungi are synthesized by the host plant. Science 2017, 356, 1175–1178. [Google Scholar] [CrossRef] [PubMed]
- Gobbato, E.; Marsh, J.F.; Vernié, T.; Wang, E.; Maillet, F.; Kim, J.; Miller, J.B.; Sun, J.; Bano, S.A.; Ratet, P.; et al. A GRAS-type transcription factor with a specific function in mycorrhizal signaling. Curr. Biol. 2012, 22, 2236–2241. [Google Scholar] [CrossRef] [PubMed]
- Huisman, R.; Bouwmeester, K.; Brattinga, M.; Govers, F.; Bisseling, T.; Limpens, E. Haustorium formation in Medicago truncatula roots infected by Phytophthora palmivora does not involve the common endosymbiotic program shared by arbuscular mycorrhizal fungi and rhizobia. Mol. Plant-Microbe Interact. 2015, 28, 1271–1280. [Google Scholar] [CrossRef] [PubMed]
- Dreher, D.; Yadav, H.; Zander, S.; Hause, B. Is there genetic variation in mycorrhization of Medicago truncatula? PeerJ 2017, 5, e3713. [Google Scholar] [CrossRef] [PubMed]
- Jorgensen, J.H. Discovery, characterization and exploitation of Mlo powdery mildew resistance in barley. Euphytica 1992, 66, 141–152. [Google Scholar] [CrossRef]
- Piffanelli, P. The barley MLO modulator of defense and cell death is responsive to biotic and abiotic stress stimuli. Plant Physiol. 2002, 129, 1076–1085. [Google Scholar] [CrossRef] [PubMed]
- Brown, J.K.M.; Andrivon, D.; Collinge, D.B.; Nicholson, P. Fitness costs and trade-offs in plant disease. Plant Pathol. 2013, 62, 1. [Google Scholar] [CrossRef]
- Bravo, A.; York, T.; Pumplin, N.; Mueller, L.A.; Harrison, M.J. Genes conserved for arbuscular mycorrhizal symbiosis identified through phylogenomics. Nat. Plants 2015, 1–23. [Google Scholar] [CrossRef] [PubMed]
- Kemen, E.; Gardiner, A.; Schultz-Larsen, T.; Kemen, A.C.; Balmuth, A.L.; Robert-Seilaniantz, A.; Bailey, K.; Holub, E.; Studholme, D.J.; MacLean, D.; et al. Gene gain and loss during evolution of obligate parasitism in the white rust pathogen of Arabidopsis thaliana. PLoS Biol. 2011, 9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martin, F.; Aerts, A.; Ahrén, D.; Brun, A.; Danchin, E.G.J.; Duchaussoy, F.; Gibon, J.; Kohler, A.; Lindquist, E.; Pereda, V.; et al. The genome of Laccaria bicolor provides insights into mycorrhizal symbiosis. Nature 2008, 452, 88–92. [Google Scholar] [CrossRef] [PubMed]
- Baxter, L.; Tripathy, S.; Ishaque, N.; Boot, N.; Cabral, A.; Kemen, E.; Thines, M.; Ah-Fong, A.; Anderson, R.; Badejoko, W.; et al. Signatures of adaptation to obligate biotrophy in the Hyaloperonospora arabidopsidis genome. Science 2010, 330, 1549–1551. [Google Scholar] [CrossRef] [PubMed]
- Sawers, R.J.H.; Gutjahr, C.; Paszkowski, U. Cereal mycorrhiza: An ancient symbiosis in modern agriculture. Trends Plant Sci. 2008, 13, 93–97. [Google Scholar] [CrossRef] [PubMed]
- Johnson, N.C. Can fertilization of soil select less mutualistic mycorrhizae? Ecol. Appl. 1993, 3, 749–757. [Google Scholar] [CrossRef] [PubMed]
- Kaeppler, S.M.; Parke, J.L.; Mueller, S.M.; Senior, L.; Stuber, C.; Tracy, W.F. Variation among maize inbred lines and detection of quantitative trait loci for growth at low phosphorus and responsiveness to arbuscular mycorrhizal fungi. Crop Sci. 2000, 40, 358–364. [Google Scholar] [CrossRef]
- Mark, G.L.; Cassells, A.C. Genotype-dependence in the interaction between Glomus fistulosum, Phytophthora fragariae and the wild strawberry (Fragaria vesca). Plant Soil 1996, 185, 233–239. [Google Scholar] [CrossRef]
- Steinkellner, S.; Hage-Ahmed, K.; García-Garrido, J.M.; Illana, A.; Ocampo, J.A.; Vierheilig, H. A comparison of wild-type, old and modern tomato cultivars in the interaction with the arbuscular mycorrhizal fungus Glomus mosseae and the tomato pathogen Fusarium oxysporum f. sp. lycopersici. Mycorrhiza 2012, 22, 189–194. [Google Scholar] [CrossRef] [PubMed]
- Miyata, K.; Kozaki, T.; Kouzai, Y.; Ozawa, K.; Ishii, K.; Asamizu, E.; Okabe, Y.; Umehara, Y.; Miyamoto, A.; Kobae, Y.; et al. The bifunctional plant receptor, OsCERK1, regulates both chitin-triggered immunity and arbuscular mycorrhizal symbiosis in rice. Plant Cell Physiol. 2014, 55, 1864–1872. [Google Scholar] [CrossRef] [PubMed]
- Lehnert, H.; Serfling, A.; Enders, M.; Friedt, W.; Ordon, F. Genetics of mycorrhizal symbiosis in winter wheat (Triticum aestivum). New Phytol. 2017, 215, 779–791. [Google Scholar] [CrossRef] [PubMed]
- Küster, H.; Becker, A.; Samac, D.; Tesfaye, M. The Medicago Truncatula Handbook; Samuel Roberts Noble Foundation: Ardmore, TN, USA, 2006. [Google Scholar]
- Gutjahr, C.; Banba, M.; Croset, V.; An, K.; Miyao, A.; An, G.; Hirochika, H.; Imaizumi-Anraku, H.; Paszkowski, U. Arbuscular mycorrhiza-specific signaling in rice transcends the common symbiosis signaling pathway. Plant Cell 2008, 20, 2989–3005. [Google Scholar] [CrossRef] [PubMed]
- Mukherjee, A.; Ané, J.-M. Germinating spore exudates from arbuscular mycorrhizal fungi: Molecular and developmental responses in plants and their regulation by ethylene. Mol. Plant-Microbe Interact. 2011, 24, 260–270. [Google Scholar] [CrossRef] [PubMed]
- Cameron, D.D.; Neal, A.L.; van Wees, S.C.M.; Ton, J. Mycorrhiza-induced resistance: More than the sum of its parts? Trends Plant Sci. 2013, 18, 539–545. [Google Scholar] [CrossRef] [PubMed]
- Öpik, M.; Vanatoa, A.; Vanatoa, E.; Moora, M.; Davison, J.; Kalwij, J.M.; Reier, Ü.; Zobel, M. The online database MaarjAM reveals global and ecosystemic distribution patterns in arbuscular mycorrhizal fungi (Glomeromycota). New Phytol. 2010, 188, 223–241. [Google Scholar] [CrossRef] [PubMed]
- Daniell, T.J.; Husband, R.; Fitter, A.H.; Young, J.P. Molecular diversity of arbuscular mycorrhizal fungi colonising arable crops. FEMS Microbiol. Ecol. 2001, 36, 203–209. [Google Scholar] [CrossRef] [PubMed]
- Abbott, L.K.; Lumley, S. Mycorrhizal Fungi: Use in Sustainable Agriculture and Land Restoration; Springer: Berlin, Germany, 2014; Volume 41, ISBN 978-3-662-45369-8. [Google Scholar]
- Verzeaux, J.; Hirel, B.; Dubois, F.; Lea, P.J.; Tétu, T. Agricultural practices to improve nitrogen use efficiency through the use of arbuscular mycorrhizae: Basic and agronomic aspects. Plant Sci. 2017, 264, 48–56. [Google Scholar] [CrossRef] [PubMed]
- An, Z.Q.; Hendrix, J.W.; Hershman, D.E.; Ferriss, R.S.; Henson, G.T. The influence of crop rotation and soil fumigation on a mycorrhizal fungal community associated with soybean. Mycorrhiza 1993, 3, 171–182. [Google Scholar] [CrossRef]
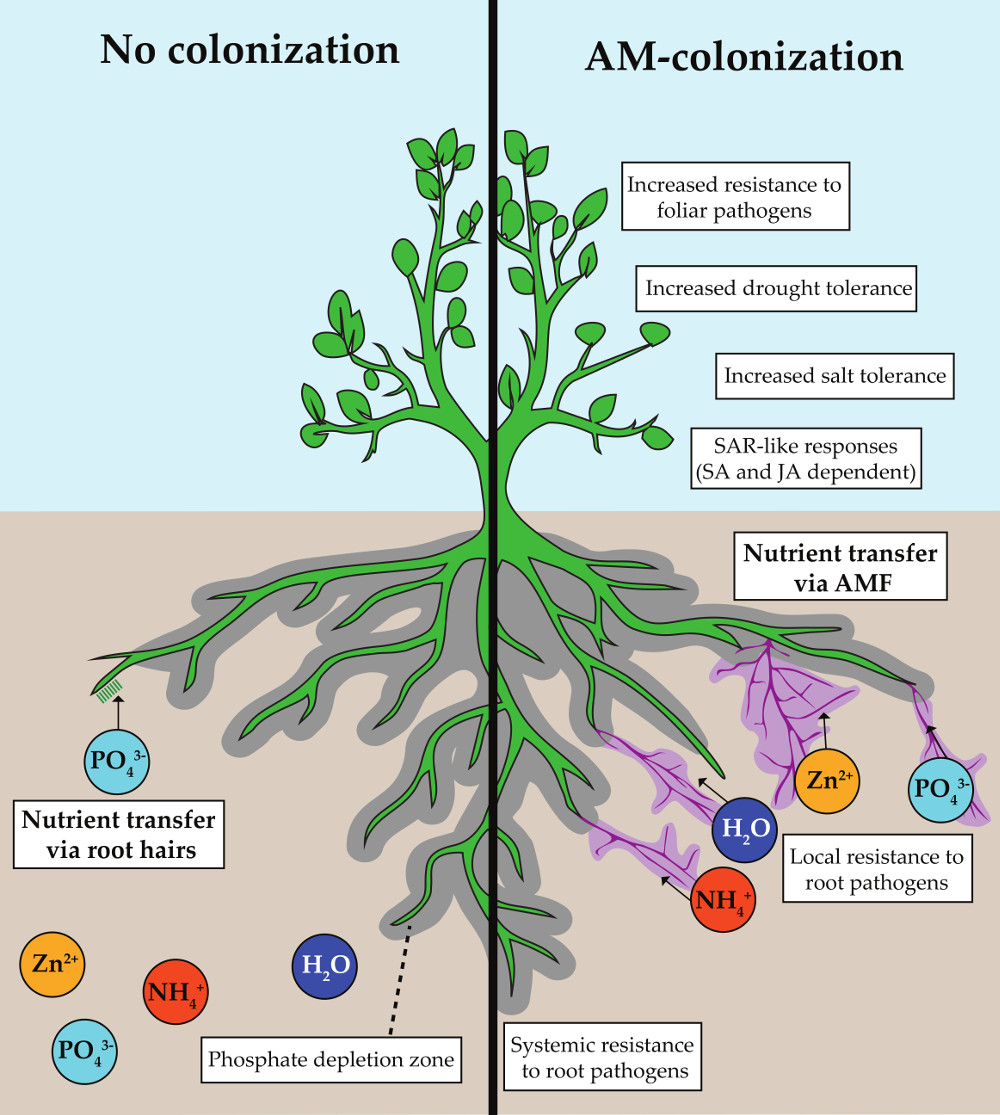
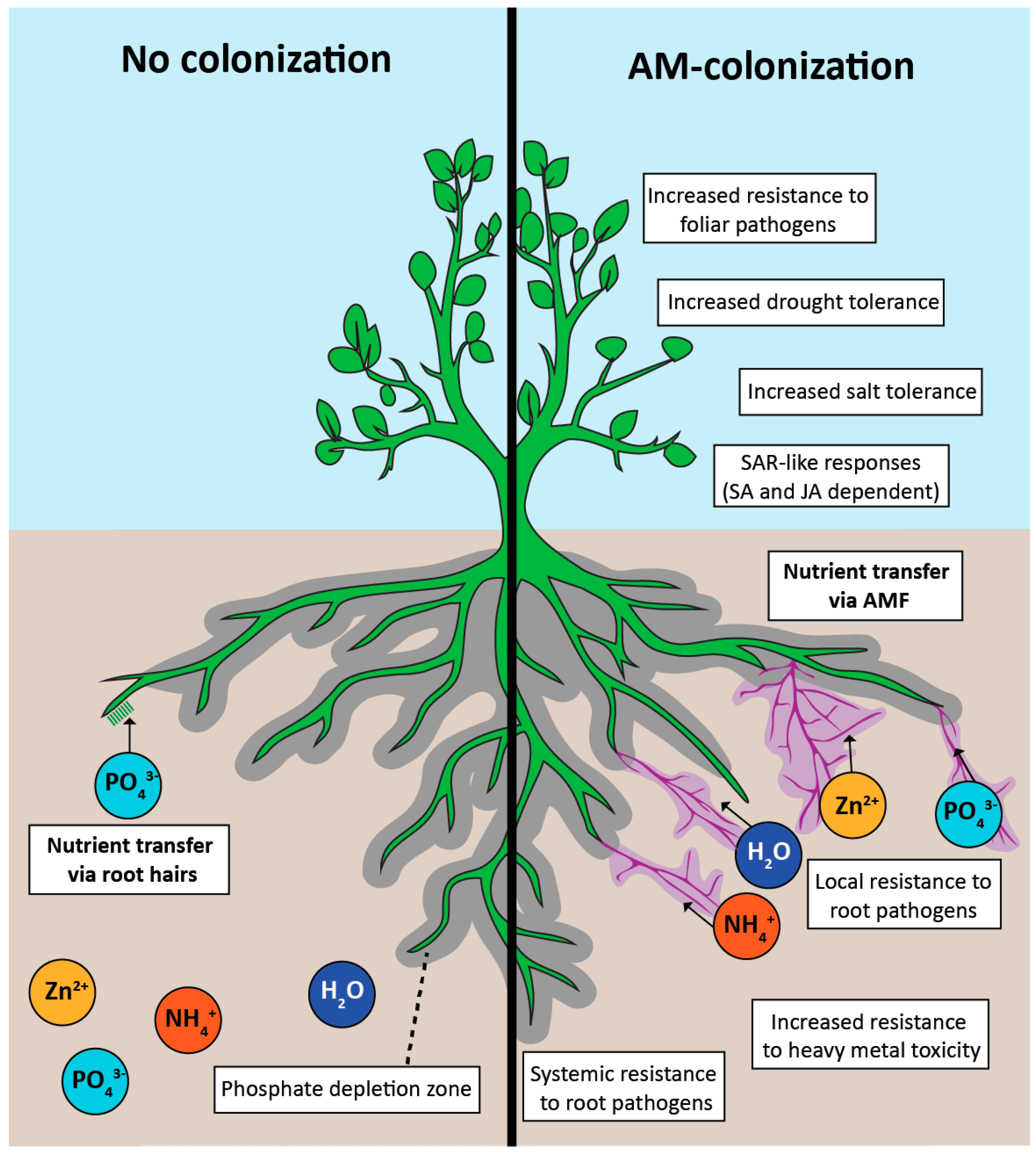
Figure 1.
Positive effects of arbuscular mycorrhizal (AM) colonization. The hyphal network of arbuscular mycorrhizal fungi (AMF) extends beyond the depletion zone (grey), accessing a greater area of soil for phosphate uptake. A mycorrhizal-phosphate depletion zone will also eventually form around AM hyphae (purple). Other nutrients that have enhanced assimilation in AM-roots include nitrogen (ammonium) and zinc. Benefits from colonization include tolerances to many abiotic and biotic stresses through induction of systemic acquired resistance (SAR).
Figure 1.
Positive effects of arbuscular mycorrhizal (AM) colonization. The hyphal network of arbuscular mycorrhizal fungi (AMF) extends beyond the depletion zone (grey), accessing a greater area of soil for phosphate uptake. A mycorrhizal-phosphate depletion zone will also eventually form around AM hyphae (purple). Other nutrients that have enhanced assimilation in AM-roots include nitrogen (ammonium) and zinc. Benefits from colonization include tolerances to many abiotic and biotic stresses through induction of systemic acquired resistance (SAR).
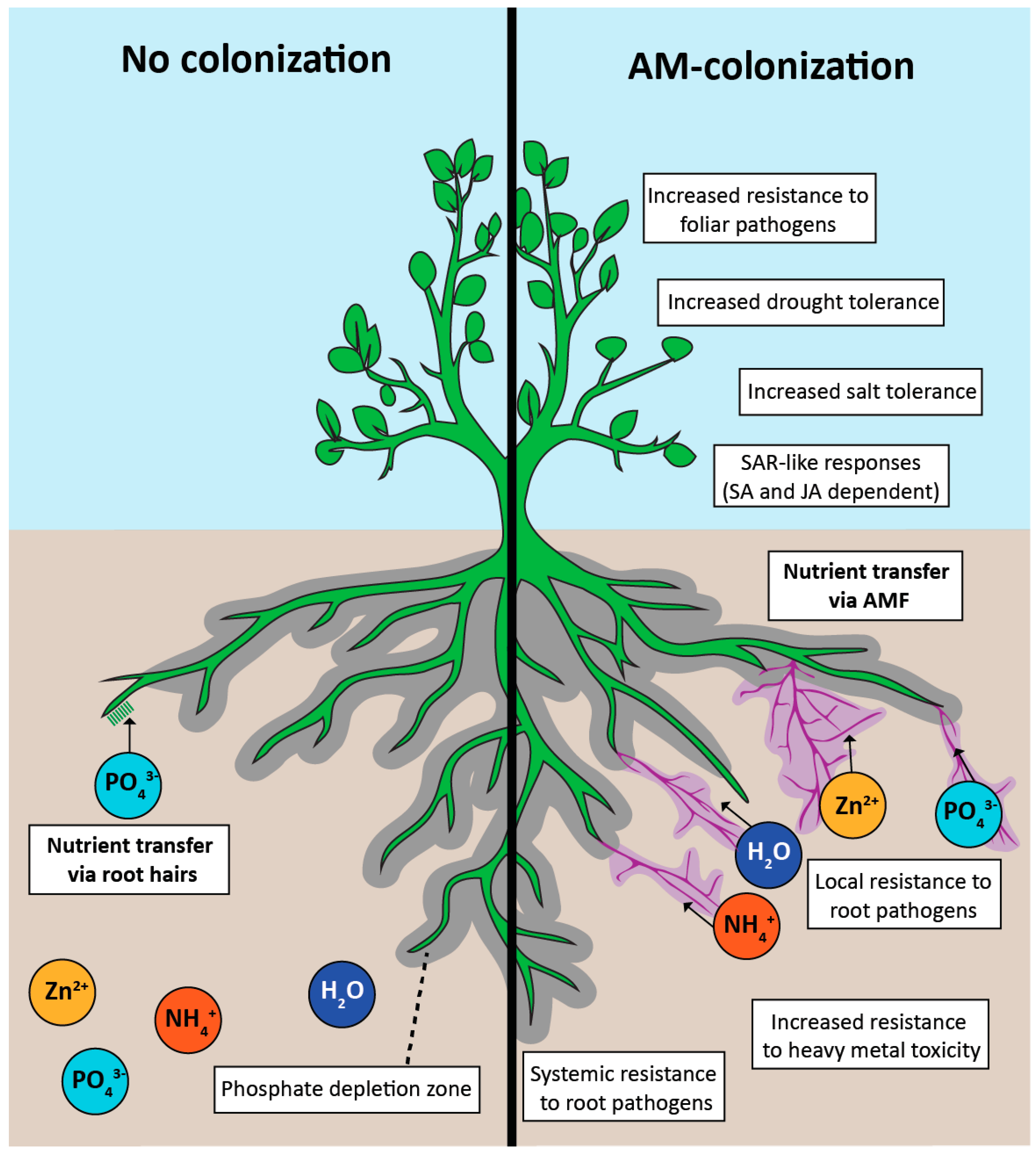
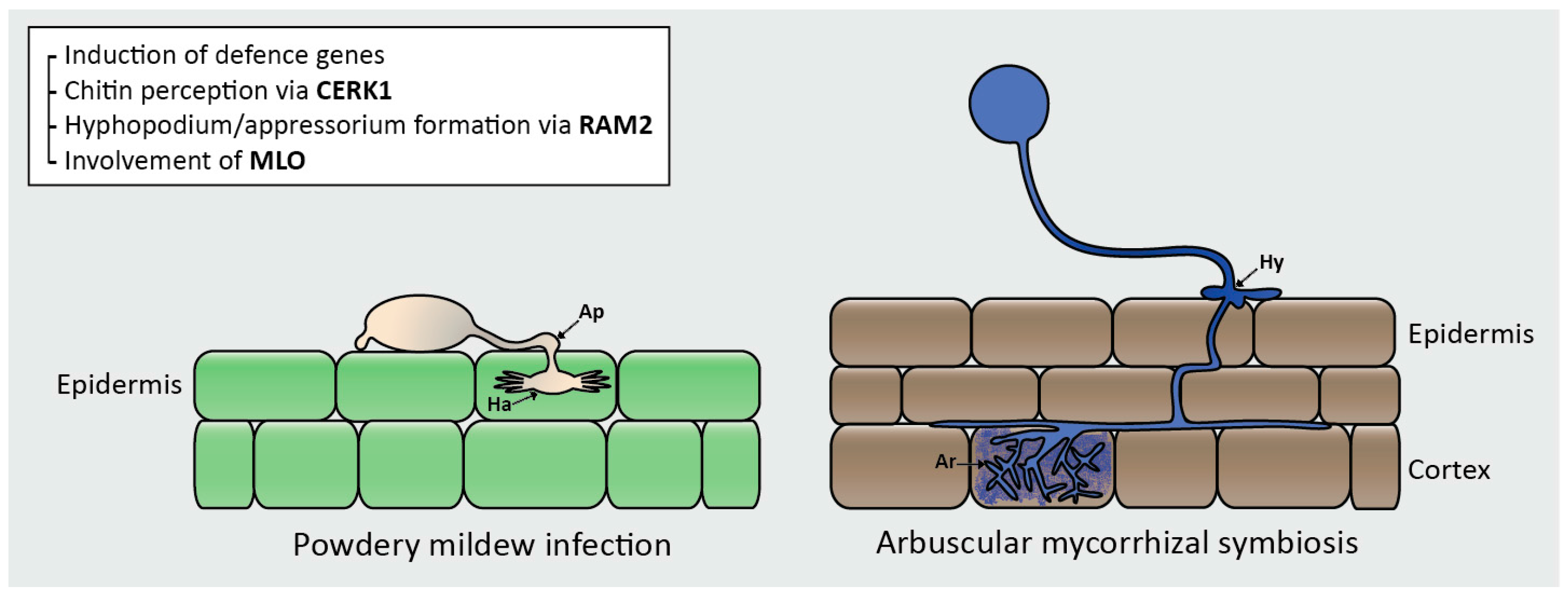
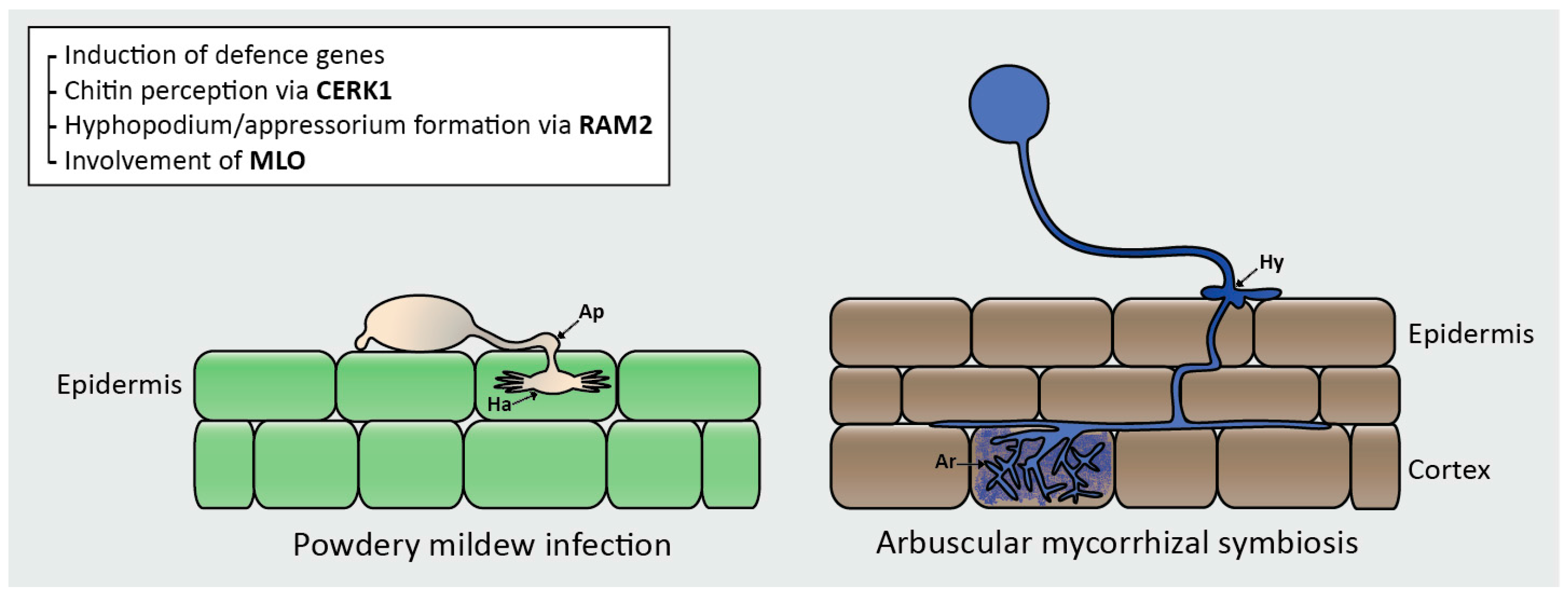
Figure 2.
An example of the physical and genetic similarities between infection by a biotrophic pathogen, powdery mildew, and colonisation by AMF. These similarities are predicted to be central to potential trade-offs. During powdery mildew infection, a spore lands on the leaf surface and germinates, forming a primary germ tube. An appressorium (Ap) is then formed which produces a penetration peg and permeates the leaf surface. Inside the epidermal cells, a haustorium (Ha) develops. During AM colonization, a spore germinates and forms an extracellular hypha which travels towards the root surface and a hyphopodium (Hy) is formed. The AM hypha travels through the epidermal cell layer to the mesodermis where it grows between the cells. Upon reaching the inner cortex, the hyphae spread laterally, entering cortical cells and branching repeatedly to form arbuscules (Ar). Common genes and defence programmes (shown in text box) are also involved in infection by pathogens and colonisation by AMF.
Figure 2.
An example of the physical and genetic similarities between infection by a biotrophic pathogen, powdery mildew, and colonisation by AMF. These similarities are predicted to be central to potential trade-offs. During powdery mildew infection, a spore lands on the leaf surface and germinates, forming a primary germ tube. An appressorium (Ap) is then formed which produces a penetration peg and permeates the leaf surface. Inside the epidermal cells, a haustorium (Ha) develops. During AM colonization, a spore germinates and forms an extracellular hypha which travels towards the root surface and a hyphopodium (Hy) is formed. The AM hypha travels through the epidermal cell layer to the mesodermis where it grows between the cells. Upon reaching the inner cortex, the hyphae spread laterally, entering cortical cells and branching repeatedly to form arbuscules (Ar). Common genes and defence programmes (shown in text box) are also involved in infection by pathogens and colonisation by AMF.

{kind=link}
{kind=link}
{kind=link}
Table 1.
Summary of the positive and negative effects reported to be associated with arbuscular mycorrhizal (AM) symbiosis.
Table 1.
Summary of the positive and negative effects reported to be associated with arbuscular mycorrhizal (AM) symbiosis.
| Responses | Positive | Negative |
|---|---|---|
| Plant growth and abiotic stresses | Increased biomass [2,39,40,41,42,43,44,45] | Reduced biomass [46,47,48,49,50,51,52] |
| Phosphate acquisition [45,46,53,54,55,56,57] | Reduced phosphate [2,27,47,58,59] | |
| Nitrogen (ammonium) acquisition [4,60,61,62,63,64 ,65,66] | ||
| Water acquisition [67,68] | ||
| Zinc acquisition [5,6] | ||
| Drought tolerance [67,68] | ||
| Salinity tolerance [29,69,70,71] | ||
| Reduced arsenic toxicity [72,73] Reduced sulphur starvation [74,75] | ||
| Reduced phosphate toxicity [76] | ||
| Reduced assimilation of adverse heavy metals [19,36,77] | ||
| Disease resistance | Increased resistance to root [31,78,79,80] and foliar [81,82,83,84,85] pathogens | Reduced arbuscular mycorrhizal colonization in mlo mutants [86] |
| Jasmonic acid production [87,88,89,90] | Components required for AM colonization used by pathogens, Golovinomyces cichoracerum (DC.) V.P. Heluta [91], Phytophthora palmivora E.J. Butler [35] and Aphanomyces euteiches Drechsler [92] | |
| Salicylic acid production [87,93] | ||
| Systemic acquired resistance [80,90,94,95,96,97] | ||
| Production of other defence compounds, e.g., phenolics [98], β-1,3-glucanase [96,97] and chitinolytic enzymes [99] |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Jacott, C.N.; Murray, J.D.; Ridout, C.J. Trade-Offs in Arbuscular Mycorrhizal Symbiosis: Disease Resistance, Growth Responses and Perspectives for Crop Breeding. Agronomy 2017, 7, 75. https://doi.org/10.3390/agronomy7040075
AMA Style
Jacott CN, Murray JD, Ridout CJ. Trade-Offs in Arbuscular Mycorrhizal Symbiosis: Disease Resistance, Growth Responses and Perspectives for Crop Breeding. Agronomy. 2017; 7(4):75. https://doi.org/10.3390/agronomy7040075
Chicago/Turabian StyleJacott, Catherine N., Jeremy D. Murray, and Christopher J. Ridout. 2017. "Trade-Offs in Arbuscular Mycorrhizal Symbiosis: Disease Resistance, Growth Responses and Perspectives for Crop Breeding" Agronomy 7, no. 4: 75. https://doi.org/10.3390/agronomy7040075
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.