Developmental Morphology and Biomass Yield of Upland and Lowland Switchgrass Ecotypes Grown in Iowa

 ,
,  , ,
, ,
Abstract
:1. Introduction
2. Material and Methods
2.1. Experimental Site
2.2. Biomass Harvest
2.3. Morphology and Development
2.4. Dry Matter Yield
2.5. Statistical Analysis
3. Results
3.1. Temperature and Precipitation
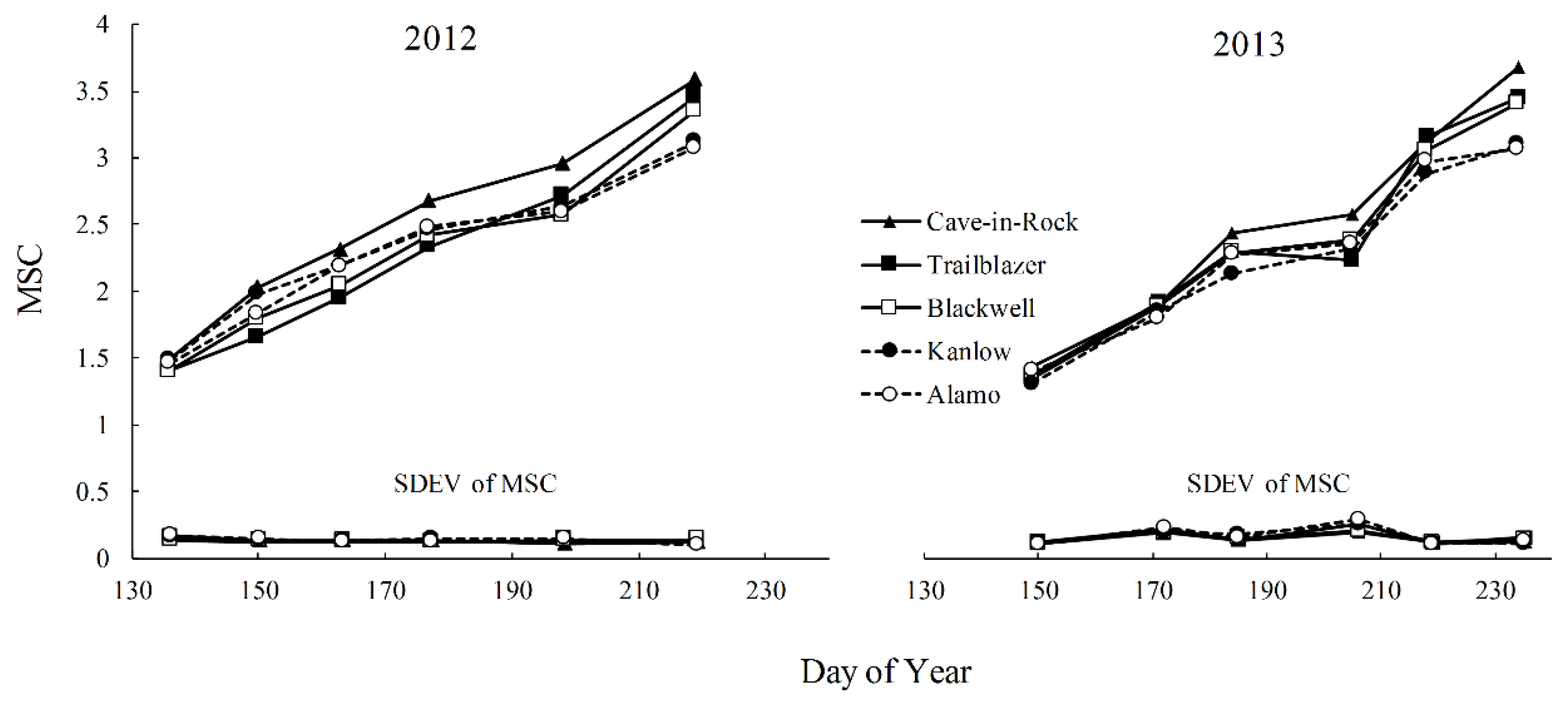
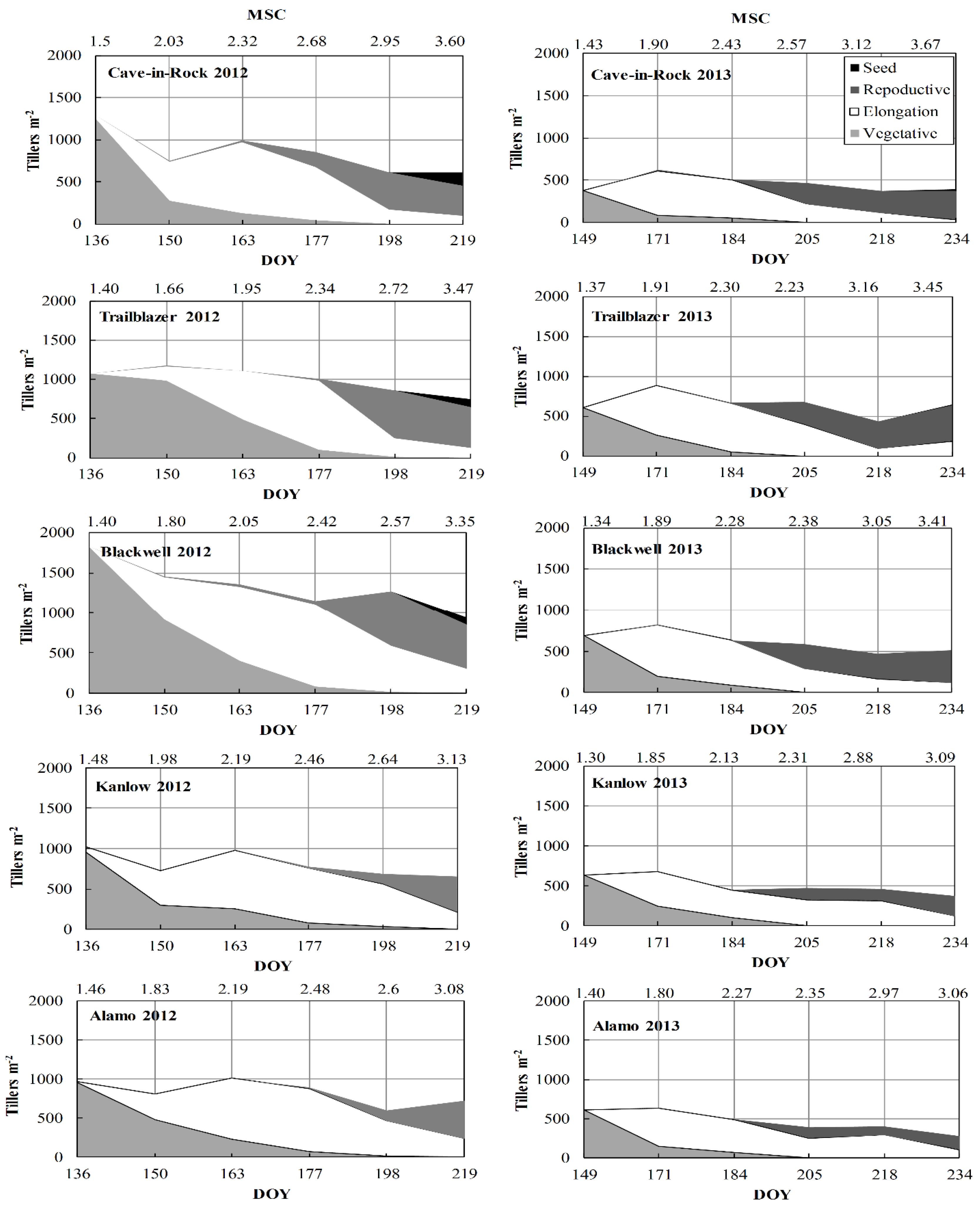
3.2. Morphology and Development
3.3. Dry Matter Yield
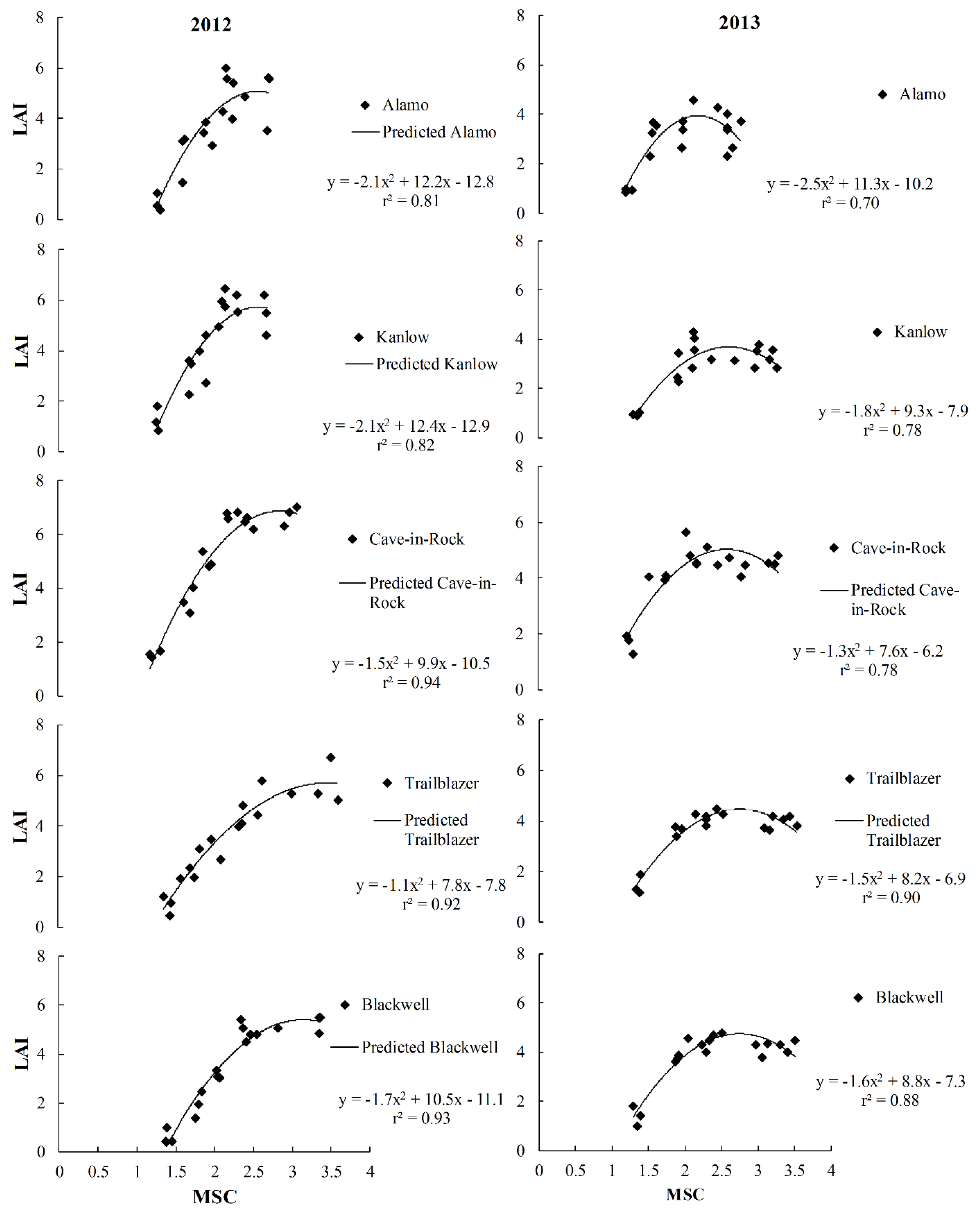
3.4. Predictive Relationships
4. Discussion
5. Conclusions
Author Contributions
Acknowledgments
Conflicts of Interest
References
- Ragauskas, A.J.; Williams, C.K.; Davison, B.H.; Britovsek, G.; Cairney, C.A.; Frederick, W.J., Jr.; Hallett, J.P.; Leak, D.J.; Liotta, C.L.; Mielenz, J.R.; et al. The path forward for biofuels and biomaterials. Science 2006, 311, 484–489. [Google Scholar] [CrossRef] [PubMed]
- Farrell, A.J.; Richard, J.P.; Plevin, T.T.; Brian, D.J.; Andrew, O.M.; Daniel, M.K. Ethanol can contribute to energy and environmental goals. Science 2006, 311, 506–508. [Google Scholar] [CrossRef] [PubMed]
- Council for Agricultural Science and Technology (CAST). Convergence of Agriculture and Energy: II. Producing Cellulosic Biomass for Biofuels; CAST Commentary QTA 2007-2; USDA-ARS/UNL Faculty: Lincoln, NE, USA, 2007.
- Bransby, D.I.; Bouton, J.; Conger, B.; McLaughlin, S.B.; Ocumpaugh, W.R.; Parrish, D.; Taliaferro, K.; Vogel, K.; Wallschleger, S. Perspectives on New Crops and New Uses; Janick, J., Ed.; ASHS Press: Alexandria, VA, USA, 1999; pp. 282–299. [Google Scholar]
- Missaoui, A.M.; Bouton, J.H.; Paterson, A.H. Molecular markers for the classification of switchgrass (Panicum virgatum L.) germplasm and to assess genetic diversity in three synthetic switchgrass populations. Gen. Res. Crop Evol. 2006, 53, 1291–1302. [Google Scholar] [CrossRef]
- Douglas, J.L.; Edwards, S.D.; Grabowski, J.M. Evaluation of Warm Season Grass Species and Management Practices to Improve Biomass Production Potential in the Mid-South; USDA Natural Resource Conservation Services: Washington, DC, USA, 2004.
- Cassida, K.A.; Muir, J.P.; Hussey, M.A.; Read, J.C.; Venuto, B.C.; Ocumpaugh, W.R. Biomass yield and stand characteristics of switchgrass in south central U.S. environments. Crop Sci. 2005, 44, 673–681. [Google Scholar] [CrossRef]
- Bouton, J.H. Molecular breeding of switchgrass for use as a biofuel crop. Curr. Opin. Genet. Dev. 2007, 17, 553–558. [Google Scholar] [CrossRef] [PubMed]
- Moser, L.E.; Vogel, K.P. Switchgrass, big bluestem, and indiangrass. In Forages, Volume 1: An Introduction to Grassland Agriculture; Barnes, R.F., Miller, D.A., Nelson, C.J., Eds.; Iowa State University Press: Ames, IN, USA, 1995; Volume 5, pp. 9–420. [Google Scholar]
- Paine, L.K. Some ecological and socio-economic considerations for biomass energy crop production. Biomass Bioenergy 1996, 10, 231–242. [Google Scholar] [CrossRef]
- Sanderson, M.A. Switchgrass as a sustainable bioenergy crop. Biores. Technol. 1996, 56, 83–93. [Google Scholar] [CrossRef]
- McLaughing, S.B.; Kszos, L.A. Development of switchgrass (Panicum virgatum) as a bioenergy feedstock in the United States. Biomass Bioenergy 2005, 28, 515–535. [Google Scholar] [CrossRef]
- Gunter, L.E.; Tuskan, G.A.; Wullschleger, S.D. Diversity among populations of switchgrass based on RAPD markers. Crop Sci. 1996, 36, 1017–1022. [Google Scholar] [CrossRef]
- Stroup, J.A.; Sanderson, M.A.; Muir, J.P.; McFarland, M.J.; Reed, R.L. Comparison of growth and performance in upland and lowland switchgrass types to water and nitrogen stress. Bioresour. Technol. 2003, 86, 65–72. [Google Scholar] [CrossRef]
- Casler, M.D.; Vogel, K.P.; Taliaferro, C.M.; Wynia, R.L. Latitudinal adaptation of switchgrass populations. Crop Sci. 2004, 44, 293–303. [Google Scholar] [CrossRef]
- Das, M.K.; Fuentes, R.G.; Taliaferro, C.M. Genetic variability and trait relationships in switchgrass. Crop Sci. 2004, 44, 443–448. [Google Scholar] [CrossRef]
- Yang, J.; Worley, E.; Wang, M.; Lahner, B.; Salt, D.E.; Saha, M.; Udvardi, M. Natural variation for nutrient use and remobilization efficiencies in switchgrass. Bioenergy Res. 2009, 2, 257–266. [Google Scholar] [CrossRef]
- Cortese, L.M.; Honig, J.; Miller, C.; Bonos, S.A. Genetic diversity of twelve switchgrass populations using molecular and morphological markers. Bioenergy Res. 2010, 3, 262–271. [Google Scholar] [CrossRef]
- Casler, M.D.; Boe, A.R. Cultivar x environment interactions in switchgrass. Crop Sci. 2003, 43, 2226–2233. [Google Scholar] [CrossRef]
- Casler, M.D. Ecotypic variation among switchgrass populations from northern USA. Crop Sci. 2005, 45, 388–398. [Google Scholar] [CrossRef]
- Parrish, D.J.; Fike, J.H.; Bransby, D.I.; Samson, R. Establishing and managing switchgrass as an energy crop. Forage Grazinglands 2008. [Google Scholar] [CrossRef]
- Lewandowski, I.; Scurlock, J.M.O.; Lindvall, E.; Christou, M. The development and current status of perennial rhizomatous grasses as energy crops in the US and Europe. Biomass Bioenerg. 2003, 25, 335–361. [Google Scholar] [CrossRef]
- Casler, M.D. Switchgrass breeding, genetics, and genomics. In Switchgrass: A Valuable Biomass Crop for Energy; Monti, A., Ed.; Springer: London, UK, 2012; Volume 43, pp. 29–53. [Google Scholar]
- Aspinwall, M.J.; Lowry, D.B.; Taylor, S.H.; Juenger, T.E.; Hawkes, C.V.; Johnson, M.V.; Kiniry, J.R.; Fay, P.A. Genotypic variation in traits linked to climate and aboveground productivity in a widespread C4 grass: Evidence for a functional trait syndrome. New Phytol. 2013, 199, 966–980. [Google Scholar] [CrossRef] [PubMed]
- Moore, K.J.; Moser, L.E.; Vogel, K.P.; Waller, S.S.; Johnson, B.E.; Pedersen, J.F. Describing and quantifying growth stages of perennial forage grasses. Agron. J. 1991, 83, 1073–1077. [Google Scholar] [CrossRef]
- Sanderson, M.A.; Wolf, D.D. Morphological development of switchgrass in diverse environments. Agron. J. 1995, 87, 908–915. [Google Scholar] [CrossRef]
- Sanderson, M.A.; Wolf, D.D. Switchgrass biomass composition during morphological development in diverse environments. Crop Sci. 1995, 35, 1432–1438. [Google Scholar] [CrossRef]
- Madakadze, I.; Coulman, B.E.; Stewart, K.; Peterson, P.; Samson, R.; Smith, D.L. Phenology and tiller characteristics of big bluestem and switchgrass cultivars in a short growing season area. Agron. J. 1998, 90, 489–495. [Google Scholar] [CrossRef]
- Cornelius, D.R.; Johnston, C.O. Differences in plant type and reaction to rust among several collections of Panicum virgatum L. J. Am. Soc. Agron. 1941, 33, 115–124. [Google Scholar] [CrossRef]
- Hopkins, A.A.; Vogel, K.P.; Moore, K.J.; Johnson, K.D.; Carlson, I.T. Genotype effects and genotype by environment interactions for traits of elite switchgrass populations. Crop Sci. 1995, 35, 125–132. [Google Scholar] [CrossRef]
- Sanderson, M.A.; Reed, R.L.; Ocumpaugh, W.R.; Hussey, M.A.; Van Esbroeck, G.; Read, J.C.; Tischler, C.; Hons, F.M. Switchgrass cultivars and germplasm for biomass feedstock production in Texas. Bioresour. Technol. 1999, 67, 209–219. [Google Scholar] [CrossRef]
- Hopkins, A.A.; Vogel, K.P.; Moore, K.J.; Johnson, K.D.; Carlson, I.T. Genotypic variability and genotype × environment interactions among switchgrass accessions from the midwestern USA. Crop Sci. 1995, 35, 565–571. [Google Scholar] [CrossRef]
- Welles, J.M.; Norman, J.M. Instrument for indirect measurement of canopy architecture. Agron. J. 1991, 83, 818–825. [Google Scholar] [CrossRef]
- Sanderson, M.A.; Wedin, W.F. Phenological stage and herbage quality relationships in temperate grasses and legumes. Agron. J. 1989, 81, 864–869. [Google Scholar] [CrossRef]
- Hintz, R.W.; Albrecht, K.A. Prediction of alfalfa chemical composition from maturity and plant morphology. Crop Sci. 1991, 31, 1561–1565. [Google Scholar] [CrossRef]
- Dien, B.S.; Jung, H.G.; Vogel, K.P.; Casler, M.D.; Lamb, J.F.S.; Mitchell, R.B.; Sarath, G. Chemical composition and response to dilute-acid pretreatment and enzymatic saccharification of alfalfa, reed canarygrass and switchgrass. Biomass Bioenergy 2006, 30, 880–891. [Google Scholar] [CrossRef]
- Moore, K.J.; Moser, L.E. Quantifying developmental morphology of perennial grasses. Crop Sci. 1995, 35, 37–43. [Google Scholar] [CrossRef]
- Mitchell, R.B.; Fritz, J.; Moore, K.; Moser, L.E.; Vogel, K.P.; Redfearn, D.; Wester, D. Predicting forage quality in switchgrass and big bluestem. Agron. J. 2001, 93, 118–124. [Google Scholar] [CrossRef]
- Dwyer, D.S.; Elder, W.C.; Singh, G. Effects of height and frequency of clipping on pure stands of range grasses in North Central Oklahoma. Oklahoma Agricultural Experiment Station Bulletin; Oklahoma Agricultural Experiment Station: Stillwater, OK, USA, 1963; B-614. [Google Scholar]
- Balasko, J.A.; Burner, D.M.; Thayne, W.V. Yield and quality of switchgrass grown without soil amendments. Agron. J. 1984, 76, 204–208. [Google Scholar] [CrossRef]
- Trocsanyi, Z.K.; Fieldsend, A.F.; Wolf, D.D. Yield and canopy characteristics of switchgrass (Panicum virgatum L.) as influenced by cutting management. Biomass Bioenergy 2009, 33, 442–448. [Google Scholar] [CrossRef]
- Madakadze, I.C.; Stewart, K.; Peterson, P.R.; Coulman, B.E.; Smith, D.L. Switchgrass biomass and chemical composition for biofuel in eastern Canada. Agron. J. 1999, 91, 696–701. [Google Scholar] [CrossRef]
- Muir, J.P.; Sanderson, M.A.; Ocumpaugh, W.R.; Jones, R.M.; Reed, R.L. Biomass production of “Alamo” switchgrass in response to nitrogen, phosphorus, and row spacing. Agron. J. 2001, 93, 896–901. [Google Scholar] [CrossRef]
- Vogel, K.P.; Brejda, J.J.; Walters, D.T.; Buxton, D.R. Switchgrass biomass production in the Midwest USA: Harvest and nitrogen management. Agron. J. 2002, 94, 413–420. [Google Scholar] [CrossRef]
- Smart, A.J.; Moser, L.E.; Vogel, K.P. Morphological characteristics of big bluestem and switchgrass plants divergently selected for seedling tiller number. Crop Sci. 2004, 44, 607–613. [Google Scholar] [CrossRef]
- Jung, H.G.; Vogel, K.P. Lignification of switchgrass (Panicum virgatum) and big bluestem (Andropogon gerardii) plant parts during maturation and its effect on fiber degradability. J. Sci. Food Agric. 1992, 59, 169–176. [Google Scholar] [CrossRef]
- Mulkey, V.R.; Owens, V.N.; Lee, D.K. Management of switchgrass dominated Conservation Reserve Program lands for biomass production in South Dakota. Crop Sci. 2006, 46, 712–720. [Google Scholar] [CrossRef]
- Waramit, N.; Moore, K.J.; Heggenstaller, A.H. Composition of native warm-season grasses for bioenergy production, in response to nitrogen fertilization rate and harvest date. Agron. J. 2011, 103, 655–662. [Google Scholar] [CrossRef]
- Nelson, C.J.; Moser, L.E. Plant factors affecting forage quality. In Forage Quality, Evaluation, and Utilization; Fahey, G.C., Jr., Collins, M., Mertens, D.R., Moser, L.E., Eds.; ASA/CSSA/SSSA: Madison, WI, USA, 1994; pp. 115–154. [Google Scholar]
- Hernandez, G.A.; Matthew, C.; Hodgson, J. Tiller size/density compensation in perennial ryegrass miniature swards subject to differing defoliation heights and a proposed productivity index. Grass Forage Sci. 1999, 54, 347–356. [Google Scholar]
- Kandel, T.P.; Wu, Y.; Kakani, V. Growth and yield responses of switchgrass ecotypes to temperature. Am. J. Plant Sci. 2013, 4, 1173–1180. [Google Scholar] [CrossRef]
- Van Esbroeck, G.A.; Hussey, M.A.; Sanderson, M.A. Leaf appearance and final leaf number of switchgrass cultivars. Agron. J. 1997, 37, 864–870. [Google Scholar] [CrossRef]
- Dahl, B.E.; Hyder, D.N. Developmental morphology and management implications. In Rangeland Plant Physiology; Sosebee, R.E., Ed.; Society for Range Management: Denver, CO, USA, 1977; pp. 257–290. [Google Scholar]
- Mitchell, R.B.; Moore, K.J.; Moser, L.E.; Fritz, J.O.; Redfearn, D.D. Predicting developmental morphology in switchgrass and big bluestem. Agron. J. 1997, 89, 827–832. [Google Scholar] [CrossRef]
- Kalu, B.A.; Fick, G.W. Quantifying morphological development for studies of herbage quality. Crop Sci. 1981, 21, 267–271. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Temperature (°C) | Precipitation (mm) | |||||
|---|---|---|---|---|---|---|
| Month | 2012 | 2013 | 20-Year Mean | 2012 | 2013 | 20-Year Mean |
| January | −2 | −5 | −7 | 7 | 15 | 20 |
| February | −1 | −3 | −4 | 44 | 20 | 25 |
| March | 12 | −1 | 3 | 60 | 38 | 50 |
| April | 12 | 8 | 10 | 122 | 148 | 98 |
| May | 19 | 16 | 16 | 62 | 180 | 125 |
| June | 23 | 21 | 21 | 75 | 76 | 132 |
| July | 27 | 23 | 23 | 37 | 26 | 107 |
| August | 22 | 23 | 22 | 74 | 55 | 124 |
| September | 18 | 19 | 18 | 47 | 50 | 81 |
| October | 10 | 11 | 12 | 59 | 97 | 61 |
| November | 6 | 1 | 4 | 23 | 40 | 51 |
| December | −2 | −8 | −4 | 26 | 8 | 27 |
| ANOVA | MSC | LAI | DMY |
|---|---|---|---|
| Year (Y) | 3.3 | 4.1 | 14.5 ** |
| Harvest date (H) | 992.1 ** | 406.3 ** | 233 ** |
| Y × H | 14.4 ** | 59.3 ** | 7.3 ** |
| Variety (V) | 23 ** | 54.7 ** | 12 ** |
| Y × V | 5.6 ** | 26.2 ** | 1.1 |
| H × V | 3.8 ** | 1.5 | 1.1 |
| Y × H × V | 0.7 | 0.8 | 0.8 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Aurangzaib, M.; Moore, K.J.; Lenssen, A.W.; Archontoulis, S.V.; Heaton, E.A.; Fei, S. Developmental Morphology and Biomass Yield of Upland and Lowland Switchgrass Ecotypes Grown in Iowa. Agronomy 2018, 8, 61. https://doi.org/10.3390/agronomy8050061
Aurangzaib M, Moore KJ, Lenssen AW, Archontoulis SV, Heaton EA, Fei S. Developmental Morphology and Biomass Yield of Upland and Lowland Switchgrass Ecotypes Grown in Iowa. Agronomy. 2018; 8(5):61. https://doi.org/10.3390/agronomy8050061
Chicago/Turabian StyleAurangzaib, Muhammad, Kenneth J. Moore, Andrew W. Lenssen, Sotirios V. Archontoulis, Emily A. Heaton, and Shuizhang Fei. 2018. "Developmental Morphology and Biomass Yield of Upland and Lowland Switchgrass Ecotypes Grown in Iowa" Agronomy 8, no. 5: 61. https://doi.org/10.3390/agronomy8050061