Alleviation of Drought Stress by Nitrogen Application in Brassica campestris ssp. Chinensis L.

School of Agriculture and Biology, Shanghai Jiao Tong University, Shanghai 200240, China
*
Author to whom correspondence should be addressed.
Agronomy 2018, 8(5), 66; https://doi.org/10.3390/agronomy8050066
Submission received: 2 April 2018
/
Revised: 26 April 2018
/
Accepted: 2 May 2018
/
Published: 4 May 2018
(This article belongs to the Special Issue Biotic and Abiotic Stress Responses in Crop Plants)
Abstract
:To assess the influence of drought stress on the growth and nitrogen nutrition status of pakchoi (Brassica campestris ssp. Chinensis L.) at different nitrogen (N) levels, the changes in N accumulation and enzyme activities involved in N assimilation were investigated. The drought was induced by adding polyethylene glycol (PEG) under hydroponic culture conditions. Pakchoi seedlings were exposed to a modified nutrient solution with different nitrogen concentration (N1, N2, and N3 represent 2, 9 and 18 mM NaNO3, respectively) and osmotic potential (W1, W2 and W3 represent 0, 60 and 120 g·L−1 PEG 6000) in a full factorial, replicated randomized block design. A short time (seven days) of drought stress caused a significant decline in plant water content, transpiration rate, shoot biomass and shoot nitrogen concentration. Increasing N availability considerably alleviate drought stress by increasing the content of total free amino acids in the roots, promoting the acceleration of root biomass accumulation, and improving the activities of nitrate reductase (NR; EC 1.7.1.1) and glutamine synthetase (GS; EC 6.3.1.2) which would reduce moisture limitations. The results suggested that pakchoi supplied with relative higher N had better growth performance under drought stress.
1. Introduction
Water and nitrogen (N) are the two main limiting factors for crop growth and productivity, especially those grown under greenhouse conditions. However, tighter water restrictions and competition with other sectors of society are increasing pressure to diminish the share of fresh water for irrigation, thus resulting in the decrease in water diverted for agriculture in China [1,2]. In general, water deficiency depresses N uptake by the root and transport to the aboveground part due to a restricted transpiration rate affecting active transport and membrane permeability [3,4]. N is a critical mineral nutrient for crop growth and involves almost all physiological processes like adenosine triphosphate (ATP) synthesis, photosynthesis and active defense mechanisms in plants [5]. To ensure the high productivity of crops, it is of particular importance to better understand N metabolism, and the utilization of crops in response to water deficit.
Nitrate () is the main N source for the crop in the soil. Once taken up by plants, the is reduced to nitrite () by nitrate reductase (NR; EC 1.7.1.1) then to ammonium () by nitrite reductase (NiR; EC 1.7.2.2). The is incorporated into glutamic acid (Glu) by glutamine synthetase (GS; EC 6.3.1.2) and glutamate synthase (GOGAT; EC 1.4.7.1). Then amino transferases like glutamic oxalacetic transaminase (GOT; EC 2.6.1.1) and glutamate pyruvate transaminase (GPT; EC 2.6.1.2) work on transformation and synthesis among amino acids [6]. It has been well documented that N uptake, allocation, and assimilation in plants were regulated by adverse conditions including water stress [7]. For example, polyethylene glycol (PEG)-induced drought stress reduced the total N content in tall fescue, Italian ryegrass, and cocksfoot [4]. Drought stress has been reported to affect the rates of absorption in almonds, with harmful effects at Ψw below −0.18 MPa [8]. The drought has been reported to restrict the plants growth through enzymes inhibition implicated in N metabolism as well [9,10,11]. For instance, Pawar et al. [10] have demonstrated that drought stress leads to decreased NR, GS, and GOGAT enzyme activities. It has also been reported that drought can alter the expression level of genes such as NRT1;2 and NRT2;5, AMT1;3, AMT1;1b, NR, GS2, and GS1;2, which are involved in N uptake and assimilation [12,13].
Previous studies also have demonstrated that higher N metabolism enhanced the drought tolerance of crops [14]. Zhong et al. [15] reported that higher levels of nitrate transport and assimilation contributed to enhancing the drought resistance of AtTGA4 transgenic Arabidopsis thaliana (L.). Chang et al. [16] found that proper N level supports regular plant growth and helps plants to defense stress. Tran et al. [17] demonstrated that higher nitrogen application rate (>75 kg N·hm−2) enhances the plasticity expression of the root development in rice under mild drought stress conditions. Shabbir et al. [18] found that combined foliar spray of supplemental nitrogen, phosphorous and potassium influenced the accumulation of osmoprotectants and activity of both N assimilation and antioxidant enzymes for improved yield and drought tolerance potential in wheat. Conversely, Liu et al. [19] suggested that elevated N deposition reduces the tolerance and ability to recover from drought in two moss species. Therefore, the regulatory function of N in drought tolerance of plant depends upon the intensity of stress, N level, and plant species.
Pakchoi (Brassica campestris ssp. Chinensis L.) is one of the most popular vegetables, cultivated over a large northern to southern range in China. Excessive N fertilizer is often employed to ensure the adequate yield and profitability from vegetable cultivation [20]; leading to most of the N accumulates in soils or is lost by runoff or lixiviation [21,22,23]. It has been reported the residual soil N in Chinese cabbage production is even high enough to avoid the requirement for additional N in the subsequent season [24]. Many studies have investigated the response of pakchoi to different N levels and N forms at the physiological and molecular levels [5,25,26,27]. However, there is less information available about the influence of drought stress on the N assimilation process and N nutrition status. In the current study, in order to understand the growth and physiological responses of pakchoi to drought stress at different N levels, we examined morphological (leaf area and root characteristics) and physiological changes (N assimilation enzymes activity, accumulation of -N, -N and total free amino acids) associated with N metabolism. We hypothesized that increasing N availability might contribute to the enhancement of nitrogen assimilation ability and the accumulation of an osmotic substance in pakchoi suffering from water deficiency, which alleviates the growth restriction.
2. Materials and Methods
2.1. Plant Cultivation and Treatments
The experiment was carried out in a glass greenhouse at School of Agriculture and Biology, Shanghai Jiao Tong University, China. The pakchoi (cv. Hua wang) seeds were sown in the vegetable seedling substrate (Agriculture Technology Co., Ltd., Huaian, China) on 27 February 2017. After twenty-one days of germination, the seedlings were transferred to black containers (54 cm × 28.5 cm × 6 cm) and cultivated hydroponically. The seedlings received distilled water, half-strength nutrient solution and total nutrient solution (pH 5.6–5.8) for 3, 3, and 6 days, respectively (acclimating hydroponics environment). The total nutrient solution contained 2.5 mM MgSO4, 2.0 mM KH2PO4, 5.0 mM CaCl2, 5.0 mM KCl, 20 μM EDTA-Fe, 46 μM H3BO3, 9.1 μM MnCl2, 1.0 μM ZnSO4, 0.32 μM CuSO4 and 0.1 μM Na2MoO4 (all the reagents were purchased from Sinopharm Chemical Reagent Co., Ltd, Shanghai, China), and the pH was 5.6~5.8. On 2 April 2017, similar seedlings were selected and transferred to a modified nutrient solution with different N concentration and osmotic potential. For N treatment, the plants were supplied with nutrient solution containing 2 mM NaNO3 (N1, low nitrogen), 9 mM NaNO3 (N2, moderate nitrogen) and 18 mM NaNO3 (N3, high nitrogen). The osmotic potential of the solution was manipulated by adding PEG 6000. The plants received a third of the dose of PEG 6000 every 6 h until it was up to the set amount. The doses of PEG 6000 applied were 0 (W1, control), 60 (W2, mild drought stress) and 120 (W3, moderate drought stress) g·L−1, respectively. Totally there were nine treatments (W1N1, W1N2, W1N3, W2N1, W2N2, W2N3, W3N1, W3N2, and W3N3), three blocks, 21 repetitions per block for 567 plants. Drought stress lasted for one week; the old leaves of pakchoi were turning yellow and even fell off. The concentration in the nutrient solution was measured using the optical method described by García-Robledo et al. [28], and NaNO3 was added on day 4 to restore the mineral concentration to its initial value. The moisture loss was evaluated by a weighing method and supplemented every evening throughout the period.
The greenhouse environmental data were monitored every 5 min by using an automatic data logging system (PM-11 Phyton monitor, Bio Instruments S.R.L., Chisinau, Moldova), and it was showed in Table S1. During the whole experimental period, the air temperature ranged from 2.88 °C to 42.95 °C with an average of 15.45 °C. The average relative humidity was 73.9%, with the highest at 92.8% and lowest at 21.4%. Mean solar radiation in the greenhouse was 2.18 MJ·m−2·day−1.
2.2. Growth Parameter Assays
The growth parameter (dry weight of the shoots and roots, total leaf area, and root morphology) of four plants in each container was determined, and the mean was considered as one repetition. The transpiration of newest leaves was measured between 9:00 am and 11:00 am on the seventh day of drought. The transpiration rate (Tr) was measured using a portable photosynthetic measurement system (Li-3000) coupled with a light-emitting diode (LED) light source (400 μmol photon m−2 s−1). The air flow was 750 μmol·s−1.
Experimental plants were harvested and divided into shoots and roots after seven days of treatment. Fresh and dry weights were determined on an electronic scale (AUY220; SHIMADZU, Kyoto, Japan) and plant water content was calculated. Leaves were scanned by an EPSON scanner (EPSON PERFECTION V700 PH07, Nagano, Japan) and total leaf area was calculated by Image J freeware (National Institutes of Health, Bethesda, MD, USA). Whereas the roots were scanned and analyzed by a WinRHIZO root analyzer system (WinRHIZO version 2007d, RegentInstruments Canada, Montreal, QC, Canada).
2.3. Nitrogen Concentration Determination
The fresh shoot and root samples were heated at 105 °C for 20 min and dried at 75 °C for 72 h. The dried samples were ground and sifted (60 mesh), and nitrogen concentration was measured using an Elementar (ELIII, Hanau, Germany).
2.4. -N, -N, Total Free Amino Acids Measurements
The -N content in the roots and leaves was spectrophotometrically analyzed as described earlier [29]. Briefly, a total of 1.00 g fresh sample was added to 5 mL of deionized water and incubated at 95 °C for 30 min. After filtration, the filtrate was diluted to 25 mL with water. And an appropriate solution (0.1 mL) was thoroughly mixed with 0.4 mL of 5% (w/v) salicylic acid in concentrated H2SO4 followed by incubation at room temperature for 20 min. The concentration was measured at 410 nm.
For ammonium quantitation, 1.00 g of the fresh sample was ground with 8 mL of distilled water. The homogenates were then centrifuged (12,000× g for 10 min) at 4 °C. The resulting supernatant was diluted 1:30 (v/v) in deionized water, and 3 mL of the solution was mixed with 0.5 mL of 1% (w/v) phenol-0.01% (w/v) sodium nitroprusside solution and 0.5 mL of 1% (v/v) sodium hypochlorite-0.5% (w/v) sodium hydroxide solution (in water). After incubating the mixture at 37 °C for 30 min, 5.9 mL of the deionized water and 0.1 mL of 8% (w/v) potassium sodium tartrate, 2% (w/v) disodium ethylenediamine tetraacetate, and 0.08% (w/v) sodium hydroxide solution (in water) were added, and absorbance was measured at 625 nm [9].
Total free amino acids were determined according to the method described by Shabbir R. N. et al. [18] with slight modifications. For this, sample mixture containing 0.3 mL of the extract, 1 mL of pyridine (10%, w/v) and 1 mL of ninhydrin solution (3%, w/v), was heated in boiling water bath for 15 min. After cooling to room temperature, 5 mL of ethyl alcohol (95%) was added and optical density was recorded at 570 nm. Total free amino acids were estimated by a standard curve constructed with alanine.
2.5. Nitrogen Metabolic Enzyme Activity Assays
2.6. Data Processing and Statistical Analysis
All the statistical analyses were carried out using SPSS software (version 20.0, SPSS Inc., Chicago, IL, USA). A two-way analysis of variance (ANOVA) was employed to examine the effect of drought and nitrogen treatments on the experimental variables. Differences between means were calculated on the basis of least significant differences (p < 0.05).
3. Results
3.1. Plant Water Content
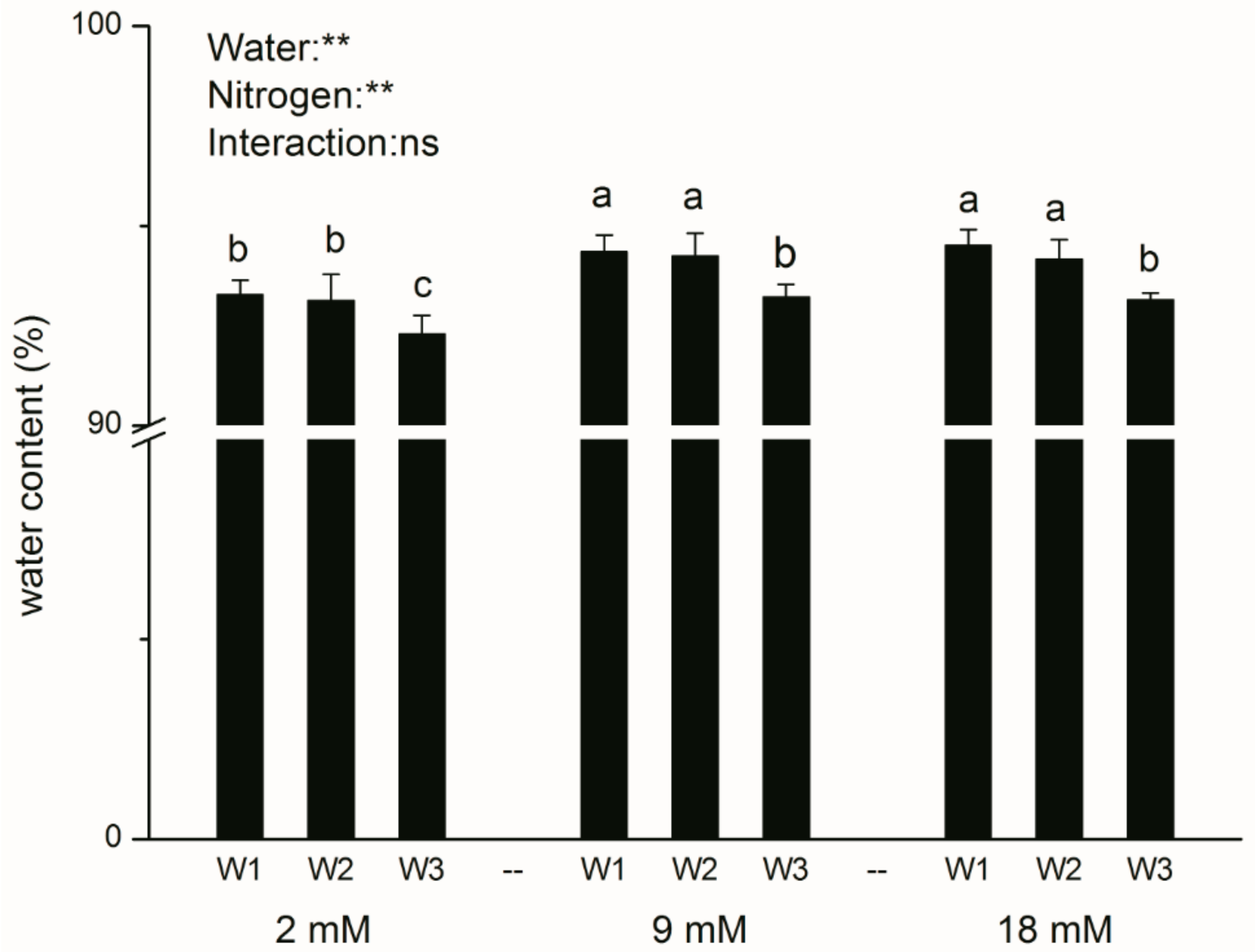
After seven days of exposure to drought stress, plant water content significantly decreased in the moderate drought group, but there were no differences between mild drought group and blank at three N levels (Figure 1). Simultaneously, plant water content was regulated by N levels, and the values in the groups of moderate and high N were dramatically higher than that in the low N treatment.
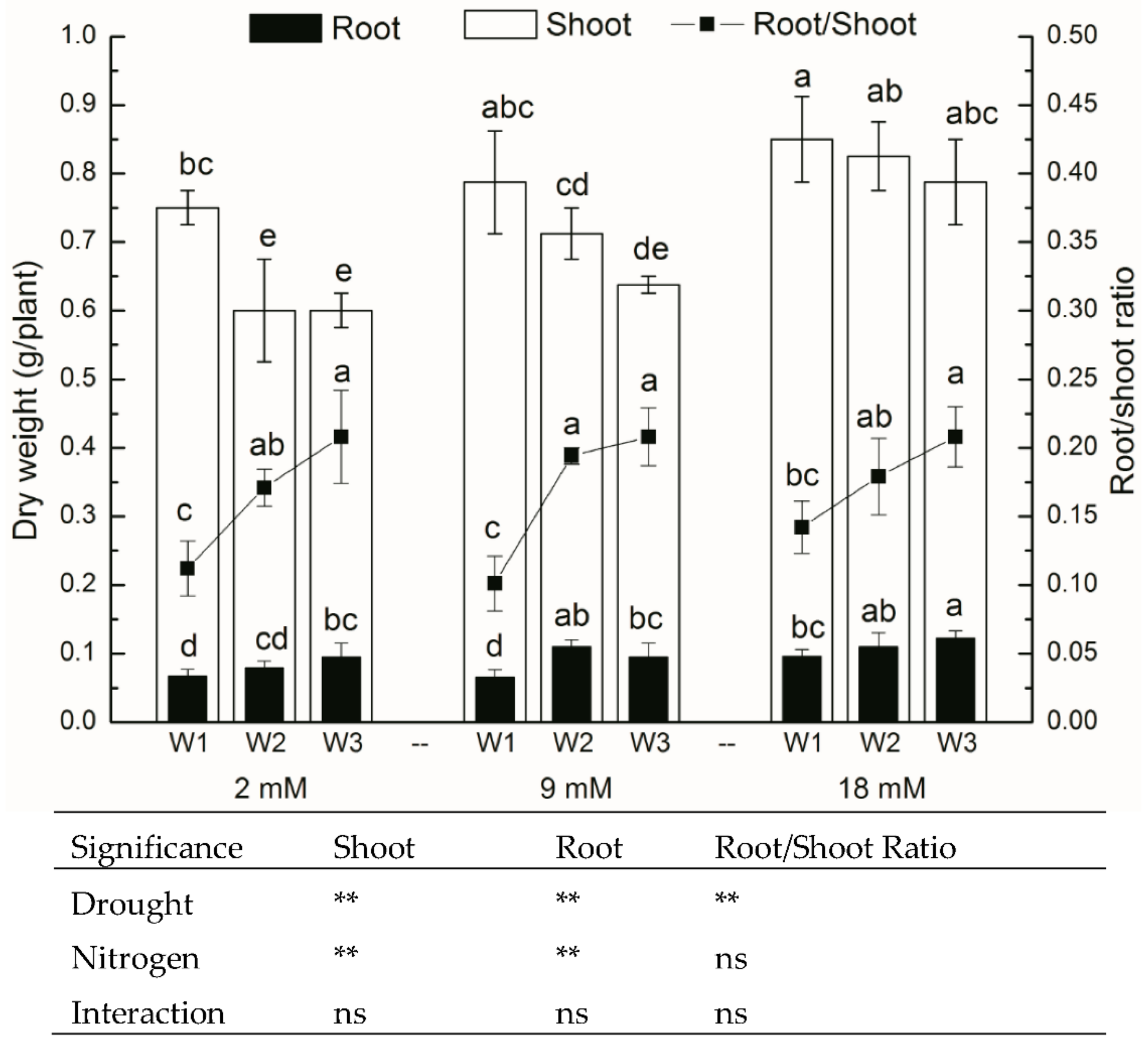
3.2. Growth Parameters
After seven days of exposure to drought stress, the aerial parts of pakchoi exhibited marked growth inhibition, resulting in a diminished leaf area and shoot biomass (Figure 2 and Table 1). Drought stress also significantly decreased (p < 0.05) the transpiration rate as well. The availability of N had a prominent effect on shoot growth under drought stress and blank groups. Under drought stress, the pakchoi supplied with higher N had higher leaf area and shoot biomass compared with the pakchoi supplied with low N.
However, root growth was promoted by the drought-induced by PEG 6000. Drought stress increased both the root biomass and root/shoot ratio (Figure 2). Total root length and root surface areas were evidently increased under drought stress conditions, and the effect was only remarkable under low nitrogen rate (Table 1). Drought stress also significantly increased the root volume and mean root diameter, and the degree of elevation positively correlated to the nitrogen rate (Table 1).
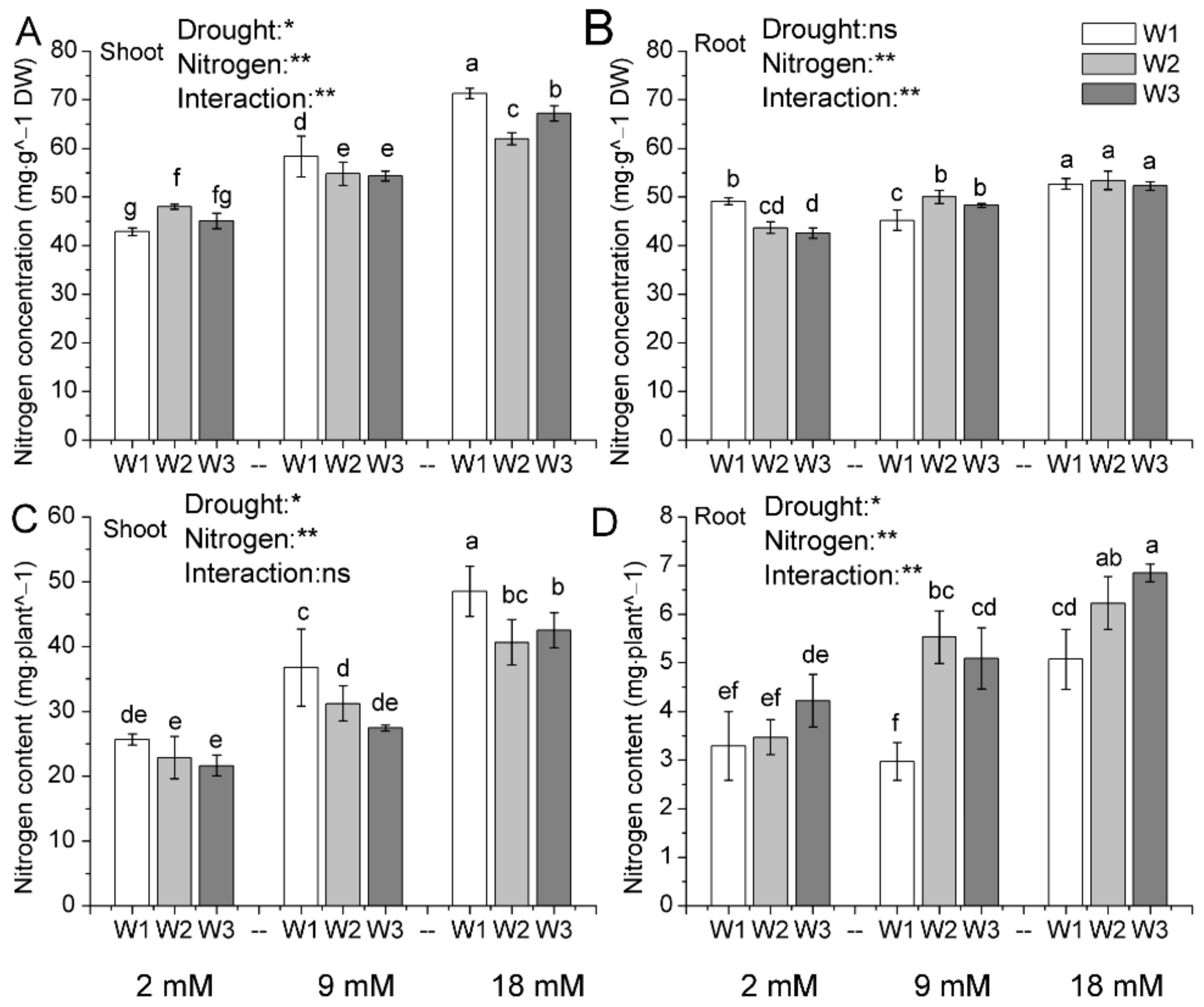
3.3. Nitrogen Content
Nitrogen accumulation responses to drought stress were measured, and the results were displayed in Figure 3 and Table S2. The N concentration was decreased in shoots with increasing drought level under normal and high nitrogen supply (Figure 3A). Mild and moderate drought stresses resulted in shoot N concentration decreased by 6.00% and 6.87% under normal nitrogen application rate. However, high nitrogen application rate could alleviate the drought stress. Under high nitrogen rate (18 mM), shoot N concentration increased by 11.61% and 23.75%, respectively, in mild and moderate drought stresses compared with the normal nitrogen application rate (9 mM). In addition, root nitrogen concentration was regulated by exogenous nitrogen application rate under the drought stress (Figure 3B). Drought stress decreased root nitrogen concentration under low nitrogen supply (2 mM) but increased under normal nitrogen supply (9 mM). On the other hand, no significant difference was observed among the high nitrogen supply (18 mM).
The total N content was decreased in shoots with increasing drought level, but it increased in roots (Figure 3C,D). Mild and moderate drought stresses resulted in total N content in shoots decreasing by 15.09% and 25.32% under normal nitrogen application rate. However, total N content in shoots would go up with increased nitrogen application rate under the drought stress. Under high nitrogen rate (18 mM), total N content in shoots increased by 30.24% and 54.92%, respectively, in mild and moderate drought stresses compared with the normal nitrogen application rate (9 mM). In addition, nitrogen accumulation in roots was regulated by both exogenous nitrogen application rate and drought stress (Figure 3D). Drought stress significantly increased root nitrogen content under normal and high nitrogen supply, but no significant change was found under low nitrogen supply. In addition, nitrogen accumulation in roots increased with the increased nitrogen supply under the drought stress.
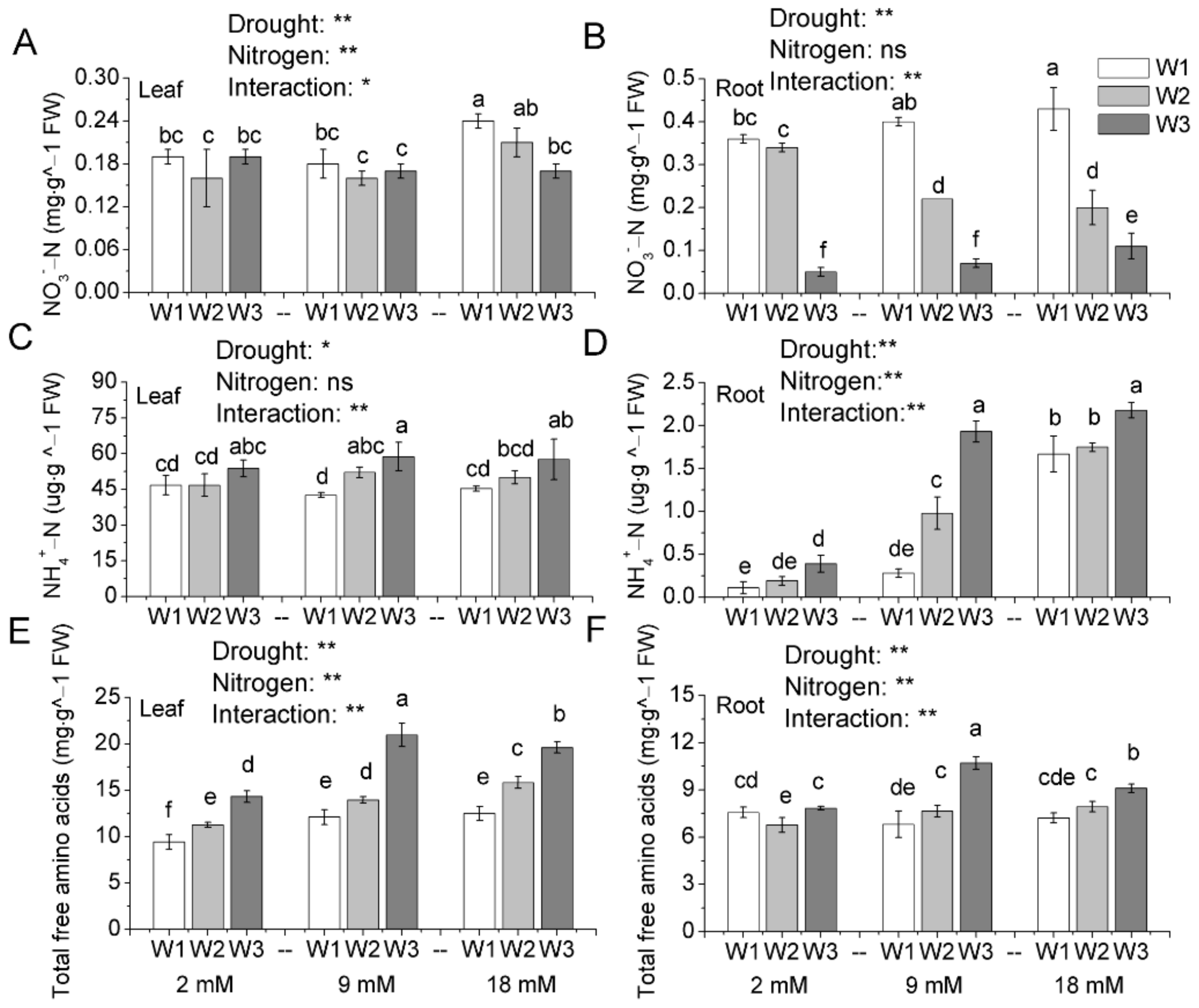
3.4. Accumulation of -N, -N, and Total Free Amino Acids
The -N, -N, and free amino acid content in the roots and leaves of pakchoi were determined and the data were showed in Table S2. Results showed that there was a significant interactive effect of drought stress and N on -N content in leaves (Figure 4A). Especially in the high N treatments, -N content significantly reduced with drought stress. Simultaneously, -N content in roots also decreased with the drought, and mild and moderate drought stress resulted in roots -N content decreased by 5.56%–53.49% and 74.42%–86.11%, respectively (Figure 4B). Similarly, -N content in roots displayed interactive effect of drought and nitrogen as well. Figure 4C indicated that drought stress increased the accumulation of -N in leaves, especially the moderate drought stress. The increment was more pronounced in the higher nitrogen supply (9 and 18 mM), displaying interaction effect of water and nitrogen. The accumulation of -N in roots was also affected by exogenous nitrogen and water supply (Figure 4D). Under the same level of nitrogen condition, drought significantly increased the -N accumulation in roots. Simultaneously, the -N accumulation increased with the increasing nitrogen rate under both control and drought stress condition. The drought stress increased the accumulation of free amino acids in shoots and roots, respectively (Figure 4E,F). Free amino acids were regulated by nitrogen application rate as well. Regardless of mild or moderate drought stress, normal and high nitrogen supply increased the accumulation of free amino acids.
3.5. Nitrogen Metabolic Enzyme Activity
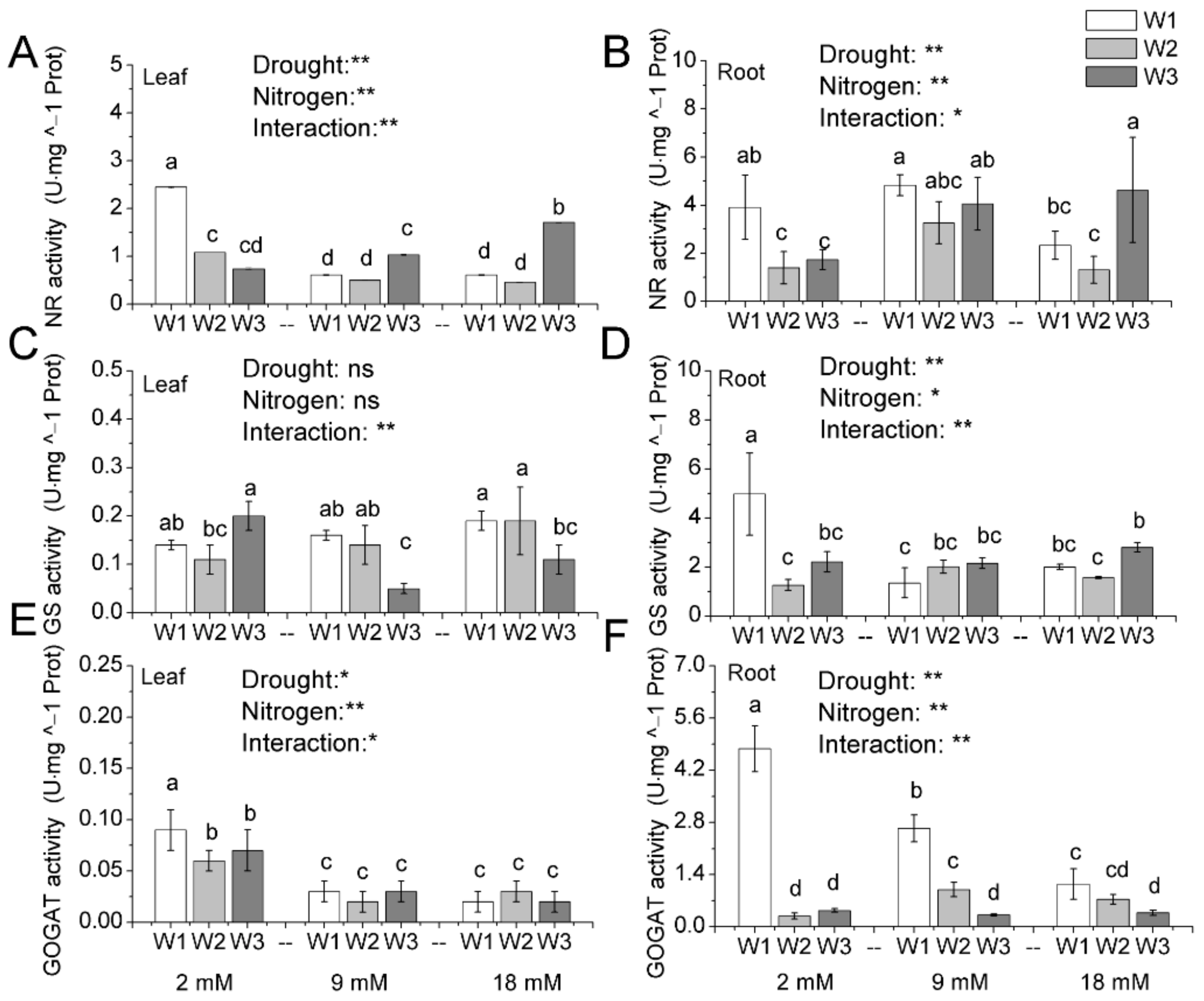
N accumulation was disturbed by drought stress, so we measured the enzyme activity involved in N assimilation (Figure 5 and Table S3). Results evidenced that there existed a significant effect of both drought and N on NR activity (Figure 5A,B). Under low N condition, drought decreased the NR activity in leaves and in roots. However, under normal and high N condition, the effect of mild drought stress on NR activity was not significant, moderate drought stress significantly enhanced NR activity in leaf and root.
Figure 5C exhibits that the effect of drought and N on the GS activity in leaves. Mild drought stress had no significant effect on GS activity in leaves. However, under moderate drought stress, normal and high N application rates significantly decreased the GS activity, but not at low N application rate. Similarly, drought and N played critical roles in the regulation of the GS activity in roots (Figure 5D). Under low N condition, drought significantly decreased the GS activity in roots. However, under normal and high N condition, the negative effect of drought stress on GS activity was not significant. The results in Figure 5E revealed that mild and moderate drought stress significantly decreased the GOGAT activity in leaves under low N application rate. However, drought stress has no significant effect on the GOGAT activity under normal and high N application rate. In roots, GOGAT activity declined with the drought stress, and high N application rate decreased the drop degree (Figure 5F). Generally, the interactive effect of drought and N on GOGAT activity was significant both in leaves and roots.
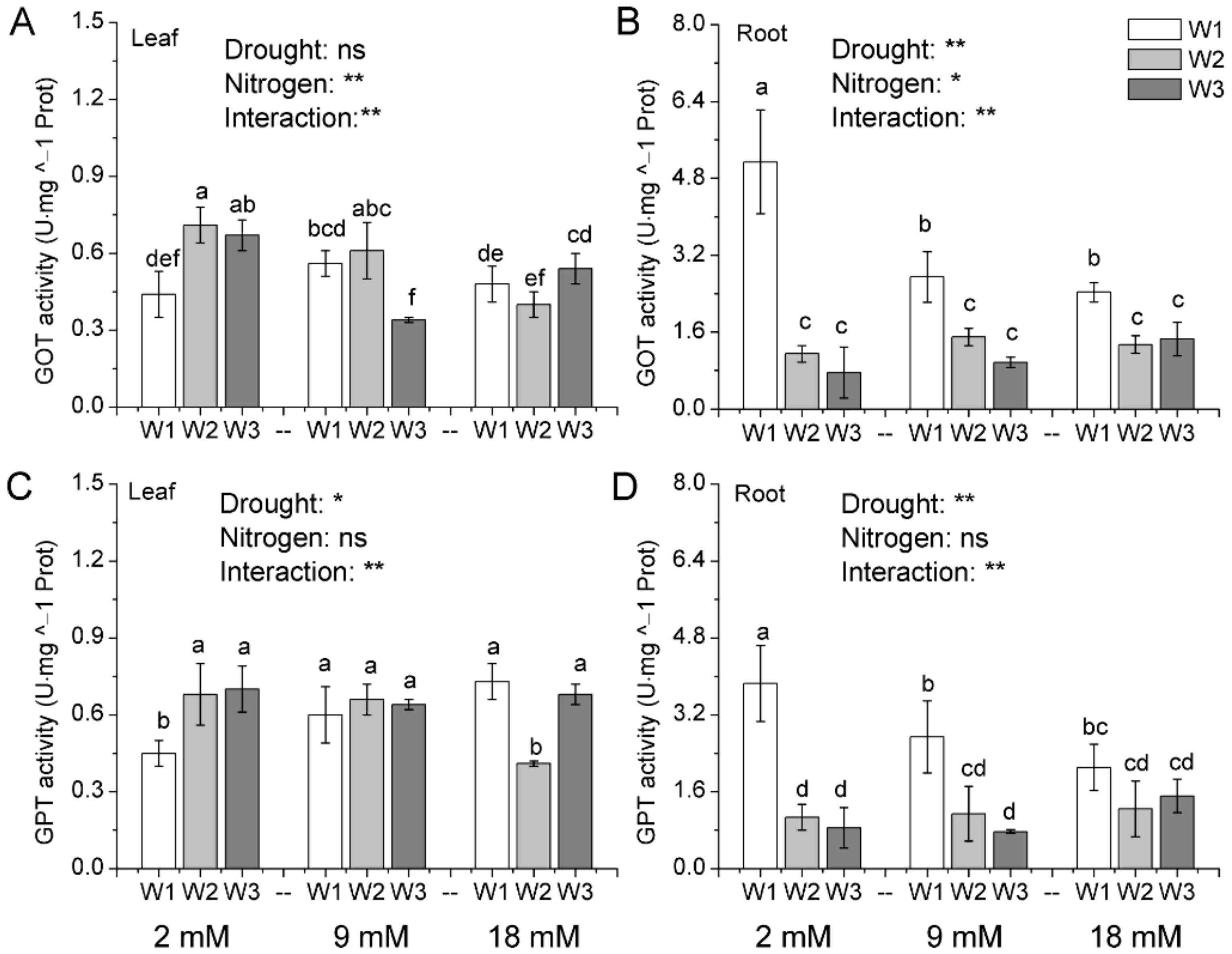
GOT and GPT are two key enzymes for transamination among amino acids in the plant. Results in Figure 6A showed that drought stress significantly increased the GOT activity in leaves under low nitrogen condition, but no significant effect was observed at normal and high N application rate. However, drought stress significantly decreased the root GOT activity, and the results were observed in all of the three N treatments (Figure 6B). The result also indicates that the normal and high nitrogen application rate decreased GOT activity in roots. Similarly, drought was found to be the critical factors in the regulation of GPT activity in leaves (Figure 6C). Under low N condition, drought significantly increased the GPT activity. However, under normal and high N condition, the positive effect of drought stress on GPT activity was not significant. Differently, the GPT activity of root was just controlled by drought stress, and the N effect was not significant (Figure 6D). For example, moderate drought stress decreased GPT activity under whatever the N application rate. Moreover, the interaction effect of drought and N on GOT and GPT activity was significant in leaves and roots, respectively.
4. Discussion
4.1. Drought Restricted the Transport of Nitrogen from Root to Aboveground
The size and configuration of roots are directly related to the absorption of moisture and nutrients because they are in direct contact with the nutrients in growth medium [33,34,35]. Generally, larger root systems with greater fine root surface area have a higher uptake rate of moisture and nutrients [36,37]. The results indicated that moderate drought stress considerably promoted the increase of total root volume and dry weight in pakchoi (Table 1 and Figure 2). Nitrate is absorbed by the root, but only a fraction of it remains in roots and participate in the building of amino acids and proteins. Most of the N is transported aboveground through the xylem by transpiration pull, and is involved in various physiological and metabolic processes in the leaves [38,39]. The drought stress significantly decreased the leaf transpiration (Table 1) and shoot N concentration (Figure 3), and this finding was consistent with Delhon et al. [40], who observed that experimental decrease in transpiration rate resulted in a lower N uptake, accompanied by an increase in root N concentration. On wheat, Nicolas et al. [41] showed that water deficit limits N translocation from roots to shoots by drastically reducing water xylem flux. Gonzalez-Dugo [4] reported that restricted root-to-shoot translocation and decreased sink size are responsible for limited nitrogen uptake in three grass species under water deficit. Therefore, drought might restrict the translocation of N from roots to shoots, and the restriction may further contribute to growth limitation. In addition, root-shoot ratios increased with drought, largely due to a stronger negative effect of water deficit on shoot growth than on root growth, which was in agreement with earlier reports [42,43].
4.2. Drought Disturbed the Nitrogen Metabolism Process
Drought might alter N metabolism and N allocation through the regulation of enzymes involved in N assimilation. In this study, water deficit alters the activities of NR, GS, and GOGAT (Figure 5), and this result agrees with Wang et al. [13] who observed that water deficit alters N uptake of maize. Since drought decreases crop growth, due to high sensitivity to leaf area expansion to plant water status, water deficit therefore also affects N demand [1]. Leaf area could act as an indicator of N demand [4]. In this study, we found that total leaf area and plant water content were decreased with drought, indicating that water limitation leads to a reduced shoot N demand, and thus a decrease in enzyme activity.
Drought also leads to an accumulation of N assimilation pathway intermediates. Ammonium from nitrate reduction is incorporated into amino acids by glutamine synthetase (GS) and glutamate synthase (GOGAT). In this study, moderate drought caused an increased accumulation of (Figure 4), an increment in NR activity, and a reduction in GS activity (Figure 5) under moderate and high N conditions, revealing that an increased accumulation of under drought might be attributed to the enhanced nitrate reduction. In addition to nitrate reduction, ammonia may also be released for re-assimilation by sink tissue from nitrogen transport compounds (e.g., asparagine, arginine and the uredine) and through breakdown of other nitrogenous compounds [3,44].
Plants antagonize drought stress mainly depend on osmotic regulation, and the ability of osmotic adjustment directly reflects drought resistance [45]. Amino acids, as an important osmotic substance, play a vital role in maintaining the cell turgor pressure and in the reduction of tissue dehydration [46,47]. In this paper, moderate drought stress significantly increased the accumulation of total free amino acids (Figure 4E,F), which contributed to reduce the cell osmotic potential and maintain cell turgor pressure. Additionally, GOT and GPT are two key enzymes for transamination among amino acids in the plant. The different activities of GOT and GPT (Figure 6) suggested that drought affect the transformation of amino acids. However, Liu et al. reported that amino N content was not affected by drought and drought did not alter the composition of amino N in the xylem and phloem in Robiniapseudoacacia L. [48]. So the composition of amino compounds responded to drought depend upon plant species, therefore the drought-mediated changes in amino N in pakchoi need to be elucidated further.
4.3. Higher Nitrogen Application Could Alleviate the Adverse Effect of Drought Stress on Pakchoi
Our data showed that drought stress decreased the leaf area, biomass and nitrogen concentration in the shoot, while higher nitrogen application mitigated the negative effects of drought stress (Table 1, Figure 1 and Figure 2). Simultaneously, the accelerated accumulation of total free amino acids in the roots and dry weight was observed in the high N supply under moderate drought stress (Figure 2 and Figure 4). Thus, increasing N availability may help alleviate drought stress by increasing osmotic regulators in the roots and promoting the acceleration of root biomass accumulation which would reduce moisture limitations. We also found that NR activity in leaves and roots were increased with the increasing nitrogen application rate under moderate drought stress, our results are in a good agreement with those reported earlier [13]. Zhong et al. reported that higher levels of nitrate transport and assimilation contributed to enhancing the drought resistance of AtTGA4 transgenic plants [15]. So we assumed that the enhancement of nitrogen assimilation ability also alleviates the growth limitation caused by water deficiency in pakchoi. However, the expressions of genes related to nitrogen transport and assimilation should be determined to reveal whether nitrogen transport and assimilation contribute to enhanced drought resistance of pakchoi from the molecular level.
5. Conclusions
PEG 6000-induced drought stress limited pakchoi development and nitrogen nutrition. Increasing N availability may alleviate drought stress by increasing osmotic regulators in the roots, promoting the acceleration of root biomass accumulation, and improving nitrogen assimilation ability which would reduce moisture limitations. Thus, our results proposed that pakchoi supplied with relative higher N had better growth performance under drought stress.
Supplementary Materials
The following are available online at https://www.mdpi.com/2073-4395/8/5/66/s1, Table S1: The greenhouse environmental data, including solar radiation (w/m2), temperature (°F), relative humidity (RH, %), photosynthetically active radiation (PAR, μE); Table S2: Nitrate-N (-N) content, Ammonium-N (-N) content, free amino acid-N content, plant nitrogen concentration and plant nitrogen content in leaves and roots of pakchoi at the end of the experiment. Table S3: The activity of nitrate reductase (NR), glutamine synthetase (GS), glutamate synthetase (GOGAT), glutamic oxalacetic transaminase (GOT) and glutamic pyruvic transaminase (GPT) in leaves and roots of pakchoi by different drought and nitrogen supply.
Author Contributions
X.X. and D.H. conceived and designed the experiments; X.X. performed the experiments; X.X. and L.C. analyzed the data and produced the figures. X.X. and M.K. wrote the manuscript. L.C., J.Z. and D.H. supervised and coordinated work. All authors reviewed the manuscript.
Acknowledgments
This study was supported in part by the Science and Technology Commission of Shanghai Municipality (Project No. 16391901700), the Science and Technology Commission of Shanghai Municipality (Project No. 17DZ2252300), the Science and Technology Commission of Shanghai Municipality (Project No. 16391904100), the earmarked fund for Shanghai Modern Leaf-vegetable Industry Technology Research System (Grant No. 201702). Any opinions, findings, conclusions, or recommendations expressed in this publication are those of the authors and do not necessarily reflect the view of the Science and Technology Commission of Shanghai Municipality or Shanghai Jiao Tong University. The authors are grateful to Muhammad Bilal from School of Life Sciences and Biotechnology, Shanghai Jiao Tong University providing assistance in language editing.
Conflicts of Interest
The authors have declared that no competing interests exist.
Abbreviations
| N | nitrogen |
| PEG | polyethylene glycol |
| nitrite | |
| Nitrate | |
| ammonium | |
| NiR | nitrite reductase |
| NR | reductase |
| GS | glutamine synthetase |
| Glu | glutamic acid |
| GOGAT | glutamate synthase |
| GOT | goxalacetic transaminase |
| GPT | glutamate pyruvate transaminase |
| W1 | control |
| W2 | mild drought stress |
| W3 | moderate drought stress |
| Tr | transpiration rate |
References
- Gonzalez-Dugo, V.; Durand, J.L. Water deficit and nitrogen nutrition of crops. Agron. Sustain. Dev. 2010, 30, 529–544. [Google Scholar] [CrossRef]
- Chai, Q.; Gan, Y. Regulated deficit irrigation for crop production under drought stress. Agron. Sustain. Dev. 2015, 36. [Google Scholar] [CrossRef]
- Sánchez-Rodríguez, E.; Romero, L. Role of Grafting in Resistance to Water Stress in Tomato Plants: Ammonia Production and Assimilation. J. Plant Growth Regul. 2013, 32, 831–842. [Google Scholar] [CrossRef]
- Gonzalez-Dugo, V.; Durand, J.L. Restricted root-to-shoot translocation and decreased sink size are responsible for limited nitrogen uptake in three grass species under water deficit. Environ. Exp. Bot. 2012, 75, 258–267. [Google Scholar] [CrossRef]
- Wang, X.L.; Tang, D.Q. Proteomic analysis of pakchoi leaves and roots under glycine-nitrogen conditions. Plant Physiol. Bioch. 2014, 75, 96–104. [Google Scholar] [CrossRef] [PubMed]
- Shi, W.M.; Xu, W.F. Responses of two rice cultivars differing in seedling-stage nitrogen use efficiency to growth under low-nitrogen conditions. Plant Soil 2009, 326, 291–302. [Google Scholar] [CrossRef]
- Chen, C.Z.; Lv, X.F. Arabidopsis NRT1.5 is another essential component in the regulation of nitrate reallocation and stress tolerance. Plant Physiol. 2012, 159, 1582–1590. [Google Scholar] [CrossRef] [PubMed]
- Martinez, H.E.P.; Olivos, A. Short-term water stress affecting absorption by almond plants. Sci. Hortic. 2015, 197, 50–56. [Google Scholar] [CrossRef]
- Meng, S.; Zhang, C. Nitrogen uptake and metabolism of Populus simonii in response to PEG-induced drought stress. Environ. Exp. Bot. 2016, 123, 78–87. [Google Scholar] [CrossRef]
- Pawar, V.V.; Lokhande, P.K. Effect of osmotic stress on osmolyte accumulation and ammonia assimilating enzymes in chickpea cultivars. Indian J. Plant Physiol. 2015, 20, 276–280. [Google Scholar] [CrossRef]
- Zahoor, R.; Zhao, W. Potassium application regulates nitrogen metabolism and osmotic adjustment in cotton (Gossypium hirsutum L.) functional leaf under drought stress. J. Plant Physiol. 2017, 215, 30–38. [Google Scholar] [CrossRef] [PubMed]
- Goel, P.; Singh, A.K. Abiotic stresses downregulate key genes involved in nitrogen uptake and assimilation in Brassica juncea L. PLoS ONE 2015, 10, e0143645. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Yang, Z. Drought Enhances Nitrogen Uptake and Assimilation in Maize Roots. Agron. J. 2017, 109, 39. [Google Scholar] [CrossRef]
- Zhong, C.; Cao, X. Nitrogen Metabolism in Adaptation of Photosynthesis to Water Stress in Rice Grown under Different Nitrogen Levels. Front. Plant Sci. 2017, 8, 1079. [Google Scholar] [CrossRef] [PubMed]
- Zhong, L.; Chen, D. AtTGA4, a bZIP transcription factor, confers drought resistance by enhancing nitrate transport and assimilation in Arabidopsis thaliana. Biochem. Biophys. Res. Commun. 2015, 457, 433–439. [Google Scholar] [CrossRef] [PubMed]
- Chang, Z.; Liu, Y. Effects of Cytokinin and Nitrogen on Drought Tolerance of Creeping Bentgrass. PLoS ONE 2016, 11, e0154005. [Google Scholar] [CrossRef] [PubMed]
- Tran, T.T.; Kano-Nakata, M. Nitrogen application enhanced the expression of developmental plasticity of root systems triggered by mild drought stress in rice. Plant Soil 2014, 378, 139–152. [Google Scholar] [CrossRef]
- Shabbir, R.N.; Waraich, E.A. Supplemental exogenous NPK application alters biochemical processes to improve yield and drought tolerance in wheat (Triticum aestivum L.). Environ. Sci. Pollut. Res. Int. 2016, 23, 2651–2662. [Google Scholar] [CrossRef] [PubMed]
- Liu, B.Y.; Lei, C.Y. Physiological responses of two moss species to the combined stress of water deficit and elevated N deposition (II): Carbon and nitrogen metabolism. Ecol. Evol. 2016, 6, 7596–7609. [Google Scholar] [CrossRef]
- Chen, Q.; Zhang, X.S. Evaluation of current fertilizer practice and soil fertility in vegetable production in the Beijing region. Nutr. Cycl. Agroecosyst. 2004, 69, 51–58. [Google Scholar] [CrossRef] [Green Version]
- Jia, X.; Shao, L. Effect of different nitrogen and irrigation treatments on yield and nitrate leaching of summer maize (Zea mays L.) under lysimeter conditions. Agric. Water Manag. 2014, 137, 92–103. [Google Scholar] [CrossRef]
- Nachmansohn, J. Minimized Nutrient Leaching through Fertilizer Management. Master’s Thesis, Swedish University of Agricultural Sciences, Uppsala, Sweden, 2016. [Google Scholar]
- Yan, X.; Ti, C. Fertilizer nitrogen recovery efficiencies in crop production systems of China with and without consideration of the residual effect of nitrogen. Environ. Res. Lett. 2014, 9, 095002. [Google Scholar] [CrossRef]
- Gao, N.; Liu, Y. Interactive effects of irrigation and nitrogen fertilizer on yield, nitrogen uptake, and recovery of two successive Chinese cabbage crops as assessed using 15N isotope. Sci. Hortic. 2017, 215, 117–125. [Google Scholar] [CrossRef]
- Tang, Y.; Sun, X. Genotypic differences in nitrate uptake, translocation and assimilation of two Chinese cabbage cultivars [Brassica campestris L. ssp. Chinensis (L.)]. Plant Physiol. Biochem. 2013, 70, 14–20. [Google Scholar] [CrossRef] [PubMed]
- Luo, J.; Jia, L. The mechanism of nitrate accumulation in pakchoi [Brassica campestris L.ssp. Chinensis (L.)]. Plant Soil 2006, 282, 10. [Google Scholar] [CrossRef]
- Han, R.F.; Yang, D. Evaluation of the yield, nitrogen use efficiency and adaptation in pakchoi cultivars under organic and conventional management systems during two successive seasons. Acta Agric. Scand. Sect. B Soil Plant Sci. 2016, 67, 85–93. [Google Scholar] [CrossRef]
- García-Robledo, E.; Corzo, A. A fast and direct spectrophotometric method for the sequential determination of nitrate and nitrite at low concentrations in small volumes. Mar. Chem. 2014, 162, 30–36. [Google Scholar] [CrossRef]
- Patterson, K.; Cakmak, T. Distinct signalling pathways and transcriptome response signatures differentiate ammonium- and nitrate-supplied plants. Plant Cell Environ. 2010, 33, 1486–1501. [Google Scholar] [CrossRef] [PubMed]
- Foyer, C.H.; Valadier, M.H. Drought induced effects on nitrate reductase activity and mRNA and on the coordination of nitrogen and carbon metabolism in maize leaves. Plant Physiol. 1998, 117, 283–292. [Google Scholar] [CrossRef] [PubMed]
- Lin, C.C.; Kao, C.H. Disturbed ammonium assimilation is associated with growth inhibition of roots in rice seedlings caused by NaCl. Plant Growth Regul. 1996, 18, 233–238. [Google Scholar] [CrossRef]
- Wu, L.H.; Tang, Q.N. The application of colormetric method on the determination of transaminase activity. Chin. J. Soil Sci. 1998, 03, 41–43. [Google Scholar]
- Kiba, T.; Krapp, A. Plant Nitrogen Acquisition Under Low Availability: Regulation of Uptake and Root Architecture. Plant Cell Physiol. 2016, 57, 707–714. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Zhang, J. Changes in root size and distribution in relation to nitrogen accumulation during maize breeding in China. Plant Soil 2013, 374, 121–130. [Google Scholar] [CrossRef]
- Aziz, M.M.; Palta, J.A. Five decades of selection for yield reduced root length density and increased nitrogen uptake per unit root length in Australian wheat varieties. Plant Soil 2016, 413, 181–192. [Google Scholar] [CrossRef]
- Feng, H.; Li, B. Overexpression of the nitrate transporter, OsNRT2.3b, improves rice phosphorus uptake and translocation. Plant Cell Rep. 2017, 36, 1287–1296. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Zheng, H. Efficacies of biochar and biochar-based amendment on vegetable yield and nitrogen utilization in four consecutive planting seasons. Sci. Total Environ. 2017, 593–594, 124–133. [Google Scholar] [CrossRef] [PubMed]
- Luo, J.; Sun, S. The Mechanism of Nitrate Accumulation in Pakchoi [Brassica Campestris L.ssp. Chinensis (L.)]. Plant Soil 2006, 282, 291–300. [Google Scholar] [CrossRef]
- Oh, K.; Kato, T. Transport of Nitrogen Assimilation in Xylem Vessels of Green Tea Plants Fed with N and N. Pedosphere 2008, 18, 222–226. [Google Scholar] [CrossRef]
- Delhon, P.; Gojon, A. Diurnal regulations of uptake in soybean plants. I. Changes in influx, efflux, and N utilization in the plant during the day/night cycle. J. Exp. Bot. 1995, 46, 1585–1594. [Google Scholar] [CrossRef]
- Nicolas, M.E.; Simpson, R.J. Effects of drought on partitioning of nitrogen in two wheat varieties differing in drought tolerance. Ann. Bot. 1985, 55, 743–754. [Google Scholar] [CrossRef]
- Xu, W.; Cui, K. Drought stress condition increases root to shoot ratio via alteration of carbohydrate partitioning and enzymatic activity in rice seedlings. Acta Physiol. Plant. 2015, 37. [Google Scholar] [CrossRef]
- Gargallo-Garriga, A.; Sardans, J. Opposite metabolic responses of shoots and roots to drought. Sci. Rep. 2014, 4, 6829. [Google Scholar] [CrossRef] [PubMed]
- Hirel, B.; Lea, P.J. Ammonia assimilation. In Plant Nitrogen; Springer: Berlin/Heidelberg, Germany, 2001; pp. 79–99. [Google Scholar]
- Serraj, R.; Sinclair, T.R. Osmolyte accumulation: Can it really help increase crop yield under drought conditions? Plant Cell Environ. 2002, 25, 333–341. [Google Scholar] [CrossRef] [PubMed]
- Kiani, S.P.; Talia, P. Genetic analysis of plant water status and osmotic adjustment in recombinant inbred lines of sunflower under two water treatments. Plant Sci. 2007, 172, 773–787. [Google Scholar] [CrossRef]
- Hussain, M.; Malik, M.A. Exogenous glycinebetaine and salicylic acid application improves water relations, allometry and quality of hybrid sunflower under water deficit conditions. J. Agron. Crop Sci. 2009, 195, 98–109. [Google Scholar] [CrossRef]
- Liu, X.P.; Gong, C.M. Response pattern of amino compounds in phloem and xylem of trees to soil drought depends on drought intensity and root symbiosis. Plant Biol. 2013, 15 (Suppl. 1), 101–108. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Plant water content of pakchoi at the end of the experiment. Bars with the different letters indicate significant difference (p < 0.05). W1: 0 g·L−1 polyethylene glycol (PEG) 6000, blank; W2: 60 g·L−1 PEG 6000, mild drought stress; W3: 120 g·L−1 PEG 6000, moderate drought. Error bars represent the standard deviation. p-values of the ANOVAs of drought, nitrogen supply, and their interaction are indicated. ns, not significant. ** p < 0.01.
Figure 1.
Plant water content of pakchoi at the end of the experiment. Bars with the different letters indicate significant difference (p < 0.05). W1: 0 g·L−1 polyethylene glycol (PEG) 6000, blank; W2: 60 g·L−1 PEG 6000, mild drought stress; W3: 120 g·L−1 PEG 6000, moderate drought. Error bars represent the standard deviation. p-values of the ANOVAs of drought, nitrogen supply, and their interaction are indicated. ns, not significant. ** p < 0.01.

Figure 2.
Shoot biomass and root biomass (bars) of pakchoi at the end of the experiment. Root/Shoot ratio is also represented (continuous line). W1: 0 g·L−1 PEG 6000, blank; W2: 60 g·L−1 PEG 6000, mild drought stress; W3: 120 g·L−1 PEG 6000, moderate drought. For the dry weight of shoot and root, bars with the different letters indicate significant difference (p < 0.05). For Root/Shoot ratio, lines with the different letters indicate significant difference (p < 0.05). Error bars represent the standard deviation. p-values of the ANOVAs of drought, nitrogen supply, and their interaction are indicated. ns, not significant. ** p < 0.01.
Figure 2.
Shoot biomass and root biomass (bars) of pakchoi at the end of the experiment. Root/Shoot ratio is also represented (continuous line). W1: 0 g·L−1 PEG 6000, blank; W2: 60 g·L−1 PEG 6000, mild drought stress; W3: 120 g·L−1 PEG 6000, moderate drought. For the dry weight of shoot and root, bars with the different letters indicate significant difference (p < 0.05). For Root/Shoot ratio, lines with the different letters indicate significant difference (p < 0.05). Error bars represent the standard deviation. p-values of the ANOVAs of drought, nitrogen supply, and their interaction are indicated. ns, not significant. ** p < 0.01.

Figure 3.
Nitrogen concentration (A,B) in shoots and roots, nitrogen content (C,D) in shoots and roots of pakchoi at the end of the experiment. W1: 0 g·L−1 PEG 6000, blank; W2: 60 g·L−1 PEG 6000, mild drought stress; W3: 120 g·L−1 PEG 6000, moderate drought. Bars with the different letters indicate significant difference (p < 0.05). Error bars represent the standard deviation. p-values of the ANOVAs of drought, nitrogen supply, and their interaction are indicated. ns, not significant. * p < 0.05. ** p < 0.01.
Figure 3.
Nitrogen concentration (A,B) in shoots and roots, nitrogen content (C,D) in shoots and roots of pakchoi at the end of the experiment. W1: 0 g·L−1 PEG 6000, blank; W2: 60 g·L−1 PEG 6000, mild drought stress; W3: 120 g·L−1 PEG 6000, moderate drought. Bars with the different letters indicate significant difference (p < 0.05). Error bars represent the standard deviation. p-values of the ANOVAs of drought, nitrogen supply, and their interaction are indicated. ns, not significant. * p < 0.05. ** p < 0.01.

Figure 4.
-N content (A,B), -N content (C,D) and total free amino acids content (E,F) in leaves and roots of pakchoi at the end of the experiment. W1: 0 g·L−1 PEG 6000, blank; W2: 60 g·L−1 PEG 6000, mild drought stress; W3: 120 g·L−1 PEG 6000, moderate drought. Bars with the different letters indicate significant difference (p < 0.05). Error bars represent the standard deviation. p-values of the ANOVAs of drought, nitrogen supply, and their interaction are indicated. ns, not significant. * p < 0.05. ** p < 0.01.
Figure 4.
-N content (A,B), -N content (C,D) and total free amino acids content (E,F) in leaves and roots of pakchoi at the end of the experiment. W1: 0 g·L−1 PEG 6000, blank; W2: 60 g·L−1 PEG 6000, mild drought stress; W3: 120 g·L−1 PEG 6000, moderate drought. Bars with the different letters indicate significant difference (p < 0.05). Error bars represent the standard deviation. p-values of the ANOVAs of drought, nitrogen supply, and their interaction are indicated. ns, not significant. * p < 0.05. ** p < 0.01.

Figure 5.
The activity of nitrate reductase (A,B), glutamine synthetase (C,D) and glutamate synthetase (E,F) in leaves and roots of pakchoi by different drought and nitrogen supply. W1: 0 g·L−1 PEG 6000, blank; W2: 60 g·L−1 PEG 6000, mild drought stress; W3: 120 g·L−1 PEG 6000, moderate drought. Bars with the different letters indicate significant difference (p < 0.05). Error bars represent the standard deviation. p-values of the ANOVAs of drought, nitrogen supply, and their interaction are indicated. ns, not significant. * p < 0.05. ** p < 0.01.
Figure 5.
The activity of nitrate reductase (A,B), glutamine synthetase (C,D) and glutamate synthetase (E,F) in leaves and roots of pakchoi by different drought and nitrogen supply. W1: 0 g·L−1 PEG 6000, blank; W2: 60 g·L−1 PEG 6000, mild drought stress; W3: 120 g·L−1 PEG 6000, moderate drought. Bars with the different letters indicate significant difference (p < 0.05). Error bars represent the standard deviation. p-values of the ANOVAs of drought, nitrogen supply, and their interaction are indicated. ns, not significant. * p < 0.05. ** p < 0.01.

Figure 6.
The activity of glutamic oxalacetic transaminase (A,B) and glutamic pyruvic transaminase (C,D) in leaves and roots of pakchoi as affected by drought and nitrogen supply. W1: 0 g·L−1 PEG 6000, blank; W2: 60 g·L−1 PEG 6000, mild drought stress; W3: 120 g·L−1 PEG 6000, moderate drought. Bars with the different letters indicate significant difference (p < 0.05). Error bars represent the standard deviation. p-values of the ANOVAs of drought, nitrogen supply, and their interaction are indicated. ns, not significant. * p < 0.05. ** p < 0.01.
Figure 6.
The activity of glutamic oxalacetic transaminase (A,B) and glutamic pyruvic transaminase (C,D) in leaves and roots of pakchoi as affected by drought and nitrogen supply. W1: 0 g·L−1 PEG 6000, blank; W2: 60 g·L−1 PEG 6000, mild drought stress; W3: 120 g·L−1 PEG 6000, moderate drought. Bars with the different letters indicate significant difference (p < 0.05). Error bars represent the standard deviation. p-values of the ANOVAs of drought, nitrogen supply, and their interaction are indicated. ns, not significant. * p < 0.05. ** p < 0.01.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Growth parameters and transpiration of pakchoi under different drought and nitrogen levels.
Table 1.
Growth parameters and transpiration of pakchoi under different drought and nitrogen levels.
| Nitrogen Level | Drought | Total Leaf Area (cm2) | Transpiration Rate (mmol m−2 s−1) | Total Root Length (cm) | Total Root Surface Area (cm2) | Total Root Volume (cm3) | Mean Root Diameter (mm) |
|---|---|---|---|---|---|---|---|
| 2 mM | Blank | 118.5 ± 11.6 d | 2.6 ± 0.5 a | 265.2 ± 10.2 b | 28.6 ± 4.1 d | 0.29 ± 0.05 e | 0.35 ± 0.04 e |
| Mild drought | 134.1 ± 10.9 cd | 2.3 ± 0.8 a | 310.7 ± 24.0 a | 46.5 ± 2.4 a | 0.44 ± 0.04 bc | 0.44 ± 0.02 cd | |
| Moderate drought | 83.2 ± 9.3 e | 1.2 ± 0.2 c | 325.3 ± 6.4 a | 36.8 ± 1.3 bc | 0.39 ± 0.04 cd | 0.44 ± 0.01 cd | |
| 9 mM | Blank | 163.2 ± 8.8 b | 2.7 ± 0.3 a | 258.4 ± 7.3 bc | 35.0 ± 2.9 bcd | 0.30 ± 0.03 e | 0.35 ± 0.02 e |
| Mild drought | 156.1 ± 13.0 bc | 1.8 ± 0.9 abc | 261.0 ± 9.0 bc | 39.1 ± 2.8 bc | 0.48 ± 0.04 b | 0.49 ± 0.05 bc | |
| Moderate drought | 92.9 ± 16.7 e | 1.7 ± 0.5 bc | 261.6 ± 22.5 b | 33.6 ± 2.2 bcd | 0.44 ± 0.02 bc | 0.52 ± 0.02 b | |
| 18 mM | Blank | 197.4 ± 15.6 a | 2.2 ± 0.8 ab | 225.4 ± 34.3 c | 32.5 ± 2.4 cd | 0.34 ± 0.03 de | 0.41 ± 0.05 de |
| Mild drought | 173.8 ± 14.8 b | 1.9 ± 0.5 abc | 245.2 ± 19.6 bc | 40.1 ± 7.5 b | 0.57 ± 0.03 a | 0.53 ± 0.02 b | |
| Moderate drought | 139.7 ± 13.6 cd | 1.7 ± 0.5 bc | 245.2 ± 16.8 bc | 29.5 ± 3.9 d | 0.44 ± 0.05 bc | 0.60 ± 0.09 a | |
| Significance | |||||||
| Drought | ** | * | * | ** | ** | ** | |
| Nitrogen | ** | ns | ** | ns | ** | ** | |
| Interaction | * | ns | ns | * | ns | ns | |
(1) Blank: 0 PEG 6000, mild drought stress: 60 g·L−1 PEG 6000, moderate drought: 120 g·L−1 PEG 6000; (2) Mean values ± SD with different letters in the column are significantly different according to the Duncan test at p < 0.05; (3) ns: non-significant, * p < 0.05, ** p < 0.01, The statistics is based on two-way analysis of variance (ANOVA).
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Xiong, X.; Chang, L.; Khalid, M.; Zhang, J.; Huang, D. Alleviation of Drought Stress by Nitrogen Application in Brassica campestris ssp. Chinensis L. Agronomy 2018, 8, 66. https://doi.org/10.3390/agronomy8050066
AMA Style
Xiong X, Chang L, Khalid M, Zhang J, Huang D. Alleviation of Drought Stress by Nitrogen Application in Brassica campestris ssp. Chinensis L. Agronomy. 2018; 8(5):66. https://doi.org/10.3390/agronomy8050066
Chicago/Turabian StyleXiong, Xin, Liying Chang, Muhammad Khalid, Jingjin Zhang, and Danfeng Huang. 2018. "Alleviation of Drought Stress by Nitrogen Application in Brassica campestris ssp. Chinensis L." Agronomy 8, no. 5: 66. https://doi.org/10.3390/agronomy8050066
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.