Efficient Callus Induction and Regeneration in Selected Indica Rice



Faculty of Biosciences and Medical Engineering, Universiti Teknologi Malaysia, 81310 UTM, Johor, Malaysia
*
Author to whom correspondence should be addressed.
Agronomy 2018, 8(5), 77; https://doi.org/10.3390/agronomy8050077
Submission received: 2 April 2018
/
Revised: 17 May 2018
/
Accepted: 18 May 2018
/
Published: 21 May 2018
Abstract
:An efficient callus induction and in vitro regeneration were developed using plant growth regulators, carbon sources, and basal media for three selected Malaysian wetland rice varieties (MR220, MR220-CL2, and MR232) and one upland variety (Bario). Effect of plant growth regulator (PGR) was carried out using four different concentrations (1–4 mg/L) of 2,4-D (2, 4-dichlorophenoxyacetic acid), and NAA (1-naphthalene acetic acid) (2.5, 5.0, 7.5, and 10 mg/L) with optimized 2,4-D. Effects of carbon sources (maltose and sorbitol), and basal media (MS, N6, and LS) were also studied with optimized PGR to maximize the induction of regenerable calli. This study found that all four varieties exhibited high frequency of callus induction on MS (Murashige and Skoog) medium that was supplemented with 3 mg/L 2,4-D and 30 g/L maltose. Callus induction frequencies in the cases of MR220, MR220-CL2, MR232, and Bario were found to be 76%, 94%, 85%, and 42% respectively. Morphological analysis through scanning electron microscopy (SEM) and histological analysis revealed the embryogenicity of the induced callus. In the regeneration study, it was observed that combination of 2 mg/L BAP (6-benzylaminopurine), 2 mg/L Kin (Kinetin) and 0.5 mg/L NAA supplemented MS medium has the potential to promote regeneration of selected indica rice varieties with higher regeneration percentage, i.e., 82% (MR220-CL2), 68% (both in MR220 and MR232), and 40% (Bario). The optimized conditions for callus formation and regeneration can be useful for biotechnological practices for the genetic improvement of Malaysian indica rice.
Keywords:
indica rice; plant growth regulator; carbon source; SEM; histology; callus induction; regeneration1. Introduction
Rice (Oryza sativa L.) is the second most widely cultivated crop in the world [1]. The major subspecies of rice are indica and japonica, while indica subspecies is the most widely cultivated crop in south and southeast Asian countries [2,3]. The National Agrofood Policy of Malaysia (2011–2012) highlighted that local rice production should be increased to ensure the country’s demand for the growing population. Malaysia sets to achieve food security up to 80% by 2020, which also increases the food demand. Several measures have been taken by the government to stabilize the rice supply, however, the food security only reached 72% as of 2017 [4]. Furthermore, the area of land available for cultivation of crop is decreasing rapidly because of urbanization as well as a decrease in the area of fertile land [5]. Previous reports showed that rice production is adversely affected by abiotic stress and is highly vulnerable to weather changes [6,7,8,9,10]. The increasing demand seriously requires an increase in yield to bridge the demand and production gap. Malaysian rice is grown both in wetland and upland areas of the country. Although the yield of wetland rice is comparatively higher than upland rice, upland rice is advantageous due to its low cost of production and low irrigation requirements [11]. The upland rice Bario is also beneficial for its sticky texture, fine elongated grains, mild pleasant aroma, and exquisite taste [12]. It could be promoted as a health food because of its low glycemic index [13]. In order to ensure food security, the country needs to develop new high yielding rice varieties [14] through available options such as molecular breeding and biotechnology [15].
To increase yield, the Malaysian Agricultural Research and Development Institute (MARDI) released several hybrid rice varieties including MR220, MR220CL2, and MR232. These hybrid rice varieties are prime in terms of their quality and comparatively higher yield. However, among the above varieties, only MR220-CL2 was superior to other varieties with regard to herbicide resistance [6,7,16]. Even though these varieties are said to be high yielding their plants are however susceptible to flooding, drought, and low resistance to diseases and pests (MR220, MR232, and Bario) [17,18,19]. To overcome these challenges, conventional breeding, genetic engineering, and in vitro tissue culture methods could be combined to develop new rice varieties [20]. Although conventional breeding methods improve rice varieties, their progress rate is slow [21]. In vitro method is known to be an efficient method for developing an improved rice variety [22,23]. However, the lack of efficient tissue culture protocols is one of the main barriers for breeding improvement and biotechnological studies. Previous research showed that the recalcitrance of indica rice was attributed to low callusing and regenerating abilities [24] compared to japonica subspecies [22] in particular to various conditions of in vitro tissue culture. Eventually, within indica subspecies, a significant variation was also found with in vitro culture response in different genotypes [3].
Somatic embryogenesis (SE) is an important plant propagation method, which can be utilized as a tool in various crop improvement programs [25]. The earliest stage of SE induction draws attention due to the situation of embryonic cells, which provides us with information of the mechanism on cell development as well as the regeneration potential that is used in plant biotechnology [26,27]. Besides, SE is a rapid and robust system for clonal propagation of many plant species receiving attention for its commercial value [28]. In this process, callus induction and subsequently plant regeneration are the prime steps and prerequisite before employing any genetic improvement program [20,29,30]. The success of SE depends on various factors including genotype, explant, plant growth regulator, basal salt, cultural conditions and developmental stages of the mother plants [31,32]. Among them, plant growth regulators play a key role in cell division and differentiation during somatic embryogenesis [33,34], thus embryogenic callus is prerequisite for successful regeneration. Success of callus induction and subsequent regeneration depends on exogenous supply of plant growth regulators (PGR) [35,36], as well as the type and its concentration of PGR [27,37,38]. In case of callus induction of rice, 2,4-D alone or in combination with 1-naphthalene acetic acid (NAA) have been successfully induced from callus-derived from seed [39,40]. Somatic embryogenesis of Malaysian rice has been studied on MRQ 74, MR80, MRQ50, MR219, MR123, MR127, MR232 by using different media combination [3,32,41,42,43]. Panjaitan et al. [31] observed that 2,4-D alone can induce calli up to 80% in MR219, whereas the combination of 2,4-D and kinetin induce lower calli percentage (61%) in the same variety [41]. However; the response of MR232 towards 2,4-D and NAA supplemented MS medium were the highest at 95% as reported by Rahman et al. [3]. These reports showed that Malaysian indica rice were genotype dependent. During the callus induction, cell necrosis was also a barrier for successful regeneration, but the reason is still unclear [44]. A previous report showed that inclusion of maltose in cultured media caused less browning compared to sucrose [40]. Although various studies have been conducted using different types and concentrations of carbon supplementation into basal media [24,45,46,47], the types and concentration of carbon sources for potential callus induction and plantlet regeneration has not been established yet [32]. During SE, different types of calli (such as embryogenic, non-embryogenic, and rhizogenic) can be induced while only embryogenic calli have potential for regeneration [48,49]. The difference between embryogenic calli and non-embryogenic calli can be revealed by scanning electron microscopy and histology [50] rather than visual observation alone. In this study, we present an efficient, robust, and rapid protocol for Malaysian rice that can be utilized as an efficient tool for further molecular studies and as a biotechnological approach.
2. Materials and Methods
2.1. Plant Materials
To improve callus induction and regeneration, three wetland Malaysian indica rice varieties namely MR220, MR232, and MR220-CL2 and one upland indica rice Bario were selected for this experiment. The wetland rice varieties were collected from MARDI, Seberang Prai and Penang, Malaysia and the upland rice variety, Bario, was collected from Sarawak, Malaysia.
2.2. Seed Surface Sterilization
Manually dehusked mature rice seeds were sterilized using 100% ethanol for 1 min. Afterwards, the ethanol was discarded and a mixture of 100% Clorox (sodium hypochlorite 5.99%) with 2–3 drops of Tween-20 (Sigma-Aldrich, St. Louis, MO, USA) were used. The seeds were then shaken on an orbital shaker at 120 rpm for 30 min. The sterilized seeds were rinsed 5 times with sterile double-distilled water and then blot dried on sterilized filter paper (90 mm).
2.3. Callus Induction Experiment
The seeds of four rice varieties were cultured on MS media [33] fortified with different concentrations of plant growth regulators. The plant growth regulators used in this study were 2,4-D and NAA. At first, 2,4-D (1–4 mg/L) was tested and then the optimized 2,4-D concentration were added with different concentrations of NAA (0.0, 2.5, 5.0, 7.5, and 10 mg/L). Subsequently, MS (Murashige and Skoog) [33], N6 [51], and LS (Linsmaier and Skoog) ([52] basal media were tested with above optimized auxin (from 2,4-D and NAA experiment) treatment. Different carbon sources (i.e., maltose and sorbitol) and sucrose with three different concentrations (10 g/L, 20 g/L, and 30 g/L) were selected as treatments on optimized basal media with optimal auxins for all four varieties. The pH of the medium was adjusted to 5.7 before autoclaving began. The culture medium was sterilized in an autoclave at 121 °C and 15 psi (103.4 kPa) for 20 min. Finally, 20–25 mL medium was poured into Petri dishes under laminar air flow to maintain a sterile condition. Seeds were cultured horizontally on to the media with different treatments under aseptic conditions and then incubated at 25 ± 2 °C in dark conditions. The callus induction frequency was recorded after 3 weeks of culturing. The following Equation (1) was used for determining the callus induction frequency as given below [53]:
2.4. Callus Morphology
Rice calli were characterized based on morphology and structure through a light microscope (Nikon Eclipse E200, Tokyo, Japan), scanning electron microscopy (SEM), and histology. The embryogenic calli were characterized and determined by these morphological studies.
2.4.1. Scanning Electron Microscopy (SEM) Observation
The 3-week old calli were selected in order to observe the calli epidermal surface of callus under the scanning electron microscope (Hitachi, Tm 30000, Tokyo, Japan). The calli were fixed with formaldehyde acetic acid (FAA; 3.7% v/v formaldehyde, 70% v/v ethanol, 5% v/v acetic acid) solution for 48 h. (The function of the fixative stage is to preserve the callus tissue. Then the fixed callus was dehydrated in a different graded ethyl alcohol series i.e., 75% (v/v), 85% (v/v), 95% (v/v) and 100% (v/v) for one hour per concentration and then dried. Then the calli were dried using critical point drying at 35 °C for one hour (CPD: Leica). The dried calli were then coated with gold and after that the callus was observed.
2.4.2. Histological Analysis
The histological study was conducted to observe the microscopic anatomy of embryogenic (E) and non-embryogenic (NE) calli from all varieties. Samples from four different varieties were fixed in FAA to preserve the cells. The calli were then dehydrated through different grades of ethanol in a series for 48 h and embedded in paraffin wax using Tissue Embedding System 2900 (TEC) (Histo Line Laboratories, Milan, Italy). Finally, the specimens were sectioned into 5 µm sections and stained with hematoxylin. The sections were then mounted on slides and observed under a light microscope (Nikon Eclipse E200, Japan). The above protocol was followed according to Liu et al. and Vega et al. [50,54].
2.5. Plant Regeneration and Acclimatization
Calli were taken from four different rice varieties (MR220, MR220-CL2, MR232, and Bario) and placed on regeneration media (RM) (MS basal media) supplemented with a combination of BAP (1–3 mg/L), Kin (1–2 mg/L) and NAA (0.5 mg/L), and 30 g/L maltose. The media were solidified with 4 g/L gelrite [42]. Every treatment contained three replicates with 3 g of calli each. All cultures were then incubated under a 16/8 (light/dark) photoperiod at 27 °C. The in vitro regenerated plantlets were transferred to culture tubes containing the same MS basal medium for shoot elongation. The in vitro regeneration response based on green spot initiation in different varieties was observed every week. After the plantlet initiation, the regenerated plantlets were transferred to culture tubes containing the same MS basal medium for shoot elongation. The number of regenerated shoots was calculated every two weeks after transferring. To facilitate root proliferation, the regenerated in vitro plantlets were transferred to a half strength basal MS medium for 7 days without any PGR supplementation. After that, the plantlet was gently washed with sterile water to completely remove the adhering medium so as to minimize fungal attacks. Then the in vitro plantlets with healthy roots were transplanted into soil within pots for acclimatization. Regeneration percentage was counted as Equation (2) below [48].
Numbers of plantlet regeneration/calli sample were also counted. Survivability percentage of in vitro raised plantlets at natural environmental conditions (27 °C ± 2) was recorded at 2-week age after transplanting the plants into pots of best regeneration media. The experiments were set-up in a completely randomized block design.
2.6. Statistical Analysis
The callus induction frequency and plantlet regeneration were recorded for three replicates. The data analyses were conducted using SPSS software version 22.0 (SPSS, Chicago, IL, USA) where mean differences were evaluated using Tukey’s test. The differences were considered statistically significant when p ≤ 0.05. Different means were marked with different letters (a, b, c, d, e) in order to represent the significant value of the differences.
3. Results and Discussion
3.1. Callus Induction
High percentage of re-generable embryogenic calli is a prerequisite for genetic manipulation towards varietal improvement [55]. This study aimed to test and improve the embryogenic potential of calli for better plant regeneration efficiency.
3.1.1. Effect of 2,4-D
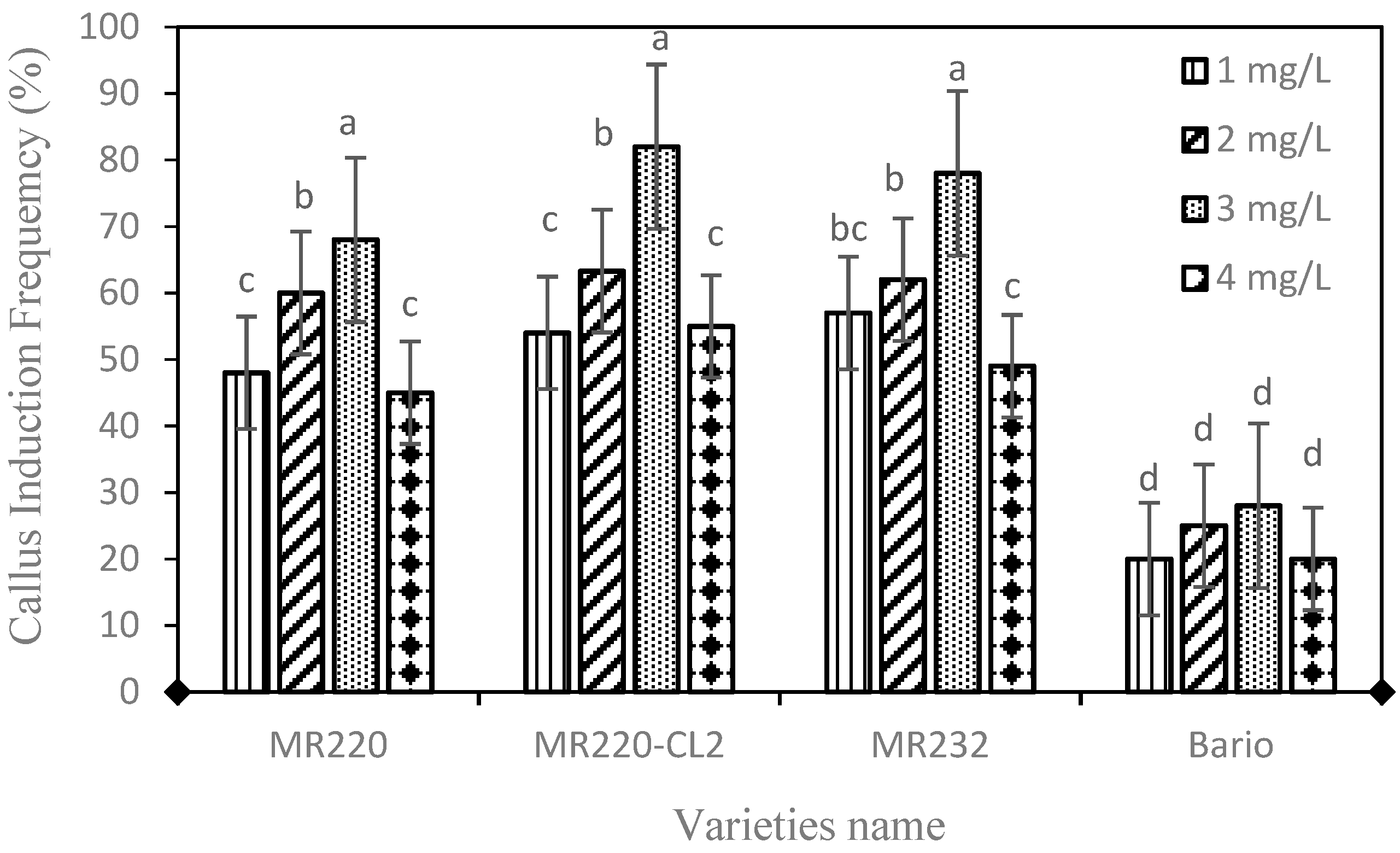
In vitro callus induction was studied from the mature seeds of four indica rice (Oryza sativa L.) varieties by using MS medium supplemented with different concentration of 2,4-D i.e., 1.0–4.0 mg/L, and without 2,4-D used as a control. The callus induction frequency (CIF) (%) in different media showed significant differences (p = 0.001). In general, all 2,4-D treatments were found to produce calli ranging from 20–82% in all four tested varieties (Figure 1). Increment of 2,4-D up to 3 mg/L increased the callus percentage but it then decreased. Within the four varieties, MR220 (68%), MR220-CL2 (82%), MR232 (78%), and Bario (28%), MS media containing 3 mg/L 2,4-D exhibit the highest frequency of calli induction. No callus formation was found on MS medium without 2,4-D which was tested as control.
The present study showed higher CIF on MS medium containing 3 mg/L 2,4-D for all wetland rice varieties, however, for Bario rice, the CIF exhibited very low compared to other studied rice varieties. Various plant growth regulators (PGR) play a key role in cell division and differentiation during somatic embryogenesis [33,34]. Within different PGR, auxin is responsible for shoot formation [44]. Previous research showed that calogenesis mostly depends on exogenous supply of auxin and the type and concentration of auxin [56]. Hence, in this study, no callus induction was found in control treatment (0.0 mg 2,4-D). Rice seeds of indica variety (CR-5272) were induced by 2,4-D at a different concentration [44]. This research also found that higher concentrations of 2,4-D decreased callus induction and turned to browning of the callus. Subsequently, the MS medium containing 2 mg/L 2,4-D was used to induce 61% callus induction frequency in MR219 [41]. This research also found that the lower percentage of callus induction was formed when using other auxin treatments namely, Picloram (16%), Dicamba (19%), and 2,4,5-T (14%) using the same rice variety. The 2 mg/L 2,4-D exhibits highest callus induction in another indica rice MR219 variety which was 75% [41]. In our study, 3 mg/L 2,4-D showed higher CIF compared to other concentrations. A variation between 2 mg and 3 mg/L 2,4-D, could be due to a different variety of indica rice as the ability to form callus is dependent on genotype, besides PGR treatment [57].
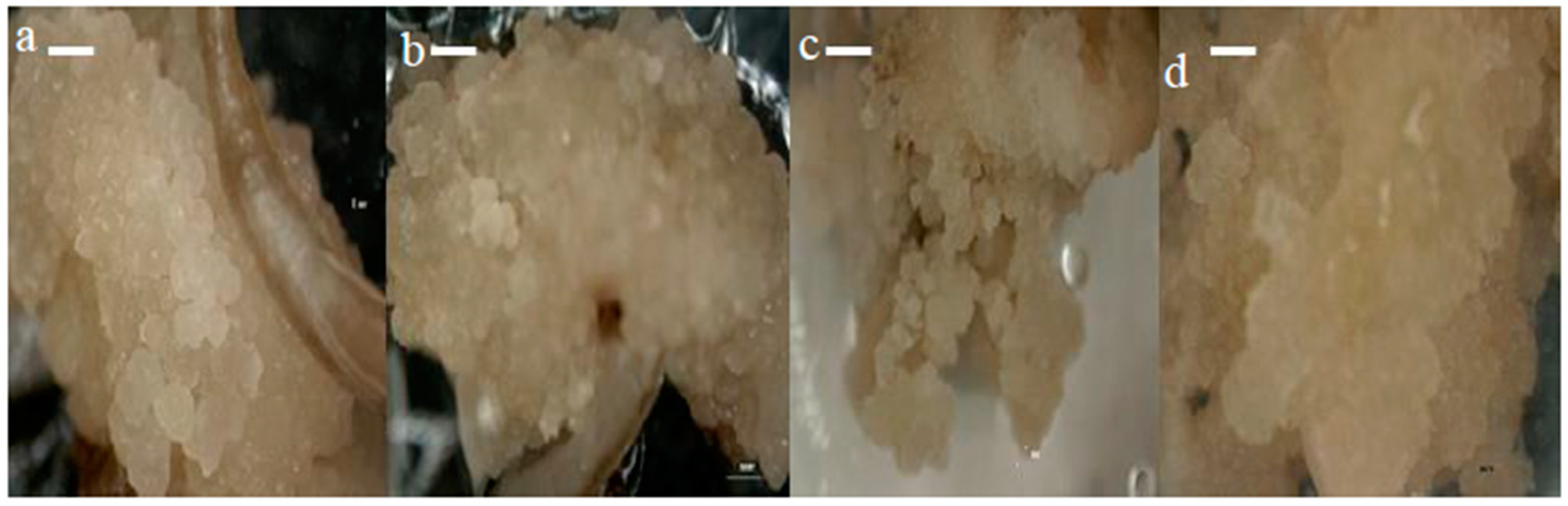
The callus morphology of four different rice varieties (MR220, MR220-CL2, MR232, and Bario) are presented in Figure 2a–d. The seeds cultured onto MS media supplemented with 3 mg/L 2,4-D, produced calli that were compact, nodular, dry, iso-diametric, and had a relatively whitish to light yellowish color with a smooth surface (Figure S1). In contrast, non-embryogenic calli had loosely held cells, were watery and yellowish to brownish in color. Our finding was in agreement with the previous rice calli induction works [41,49]. Based on color and structural morphology, the calli obtained from the present experiment were mostly embryogenic on MS media containing 3 mg/L 2,4-D. Previous literature illustrated that 2,4-D was effective for callusing in several other Malaysian indica rice [58], which corroborates the present study.
3.1.2. Effect of 2,4-D and NAA
The present study was conducted to see if combination of optimal 2,4-D (3 mg/L) and different concentrations of NAA (2.5 mg/L, 5.0 mg/L, 7.5 mg/L, 10 mg/L) would increase the callus induction percentage compared to 2,4-D only. The effect of 2,4-D and NAA with different combinations is presented in Table 1. The range of CIF was between 59.4% and 82% in varieties MR232, MR220, and MR220-CL2 for concentrations of 2,4-D and NAA. The effect of 2,4-D with combined NAA did not show any significant difference of CIF in the varieties of MR220, MR220-Cl2, and MR232. The highest callus induction frequency was 32.8% for Bario rice when explant was cultured on MS media with 3 mg/L 2,4-D and 2.5mg/L NAA. Increasing NAA concentrations for 5 mg/L NAA onwards decreased the callus percentage for Bario. In the case of Bario, the lowest callus induction (22.5%) was found on MS medium supplemented with 1 mg/L 2,4-D and 10 mg/L NAA. The results further demonstrated that combination of auxin (i.e., 3 mg 2,4-D + 2.5–10 mg/L NAA) did not show any significant difference in CIF for wetland rice varieties.
According to a previous study, auxin was known to play an important role in callus induction and proliferation [38]. Moreover, the combination of different types of auxins was a better alternative to the use of single auxin for callus induction of rice [59]. In this study, all varieties showed non-significant results when treated with the combination of NAA and 2,4-D. Moreover, a lower calli induction of all four varieties was recorded when using a higher concentration of auxin treatment. This result is in support of Verma et al. [60], lower calli induction were detected on three indica rice cultivars (Govind, Jaya, Basmati1) when using higher concentration of auxin. Previous research showed that combination of 2,4-D and NAA caused a reduction in callus induction frequency [42]. Both the above statements support the upland cultivar Bario, however, they differed in the cases of wetland cultivar i.e., MR232, MR220, MR220-CL2 [61]. Therefore, MS medium supplemented with 3 mg/L 2,4-D was selected to be the optimal concentration for callus induction in all varieties.
3.1.3. Effect of Different Basal Media
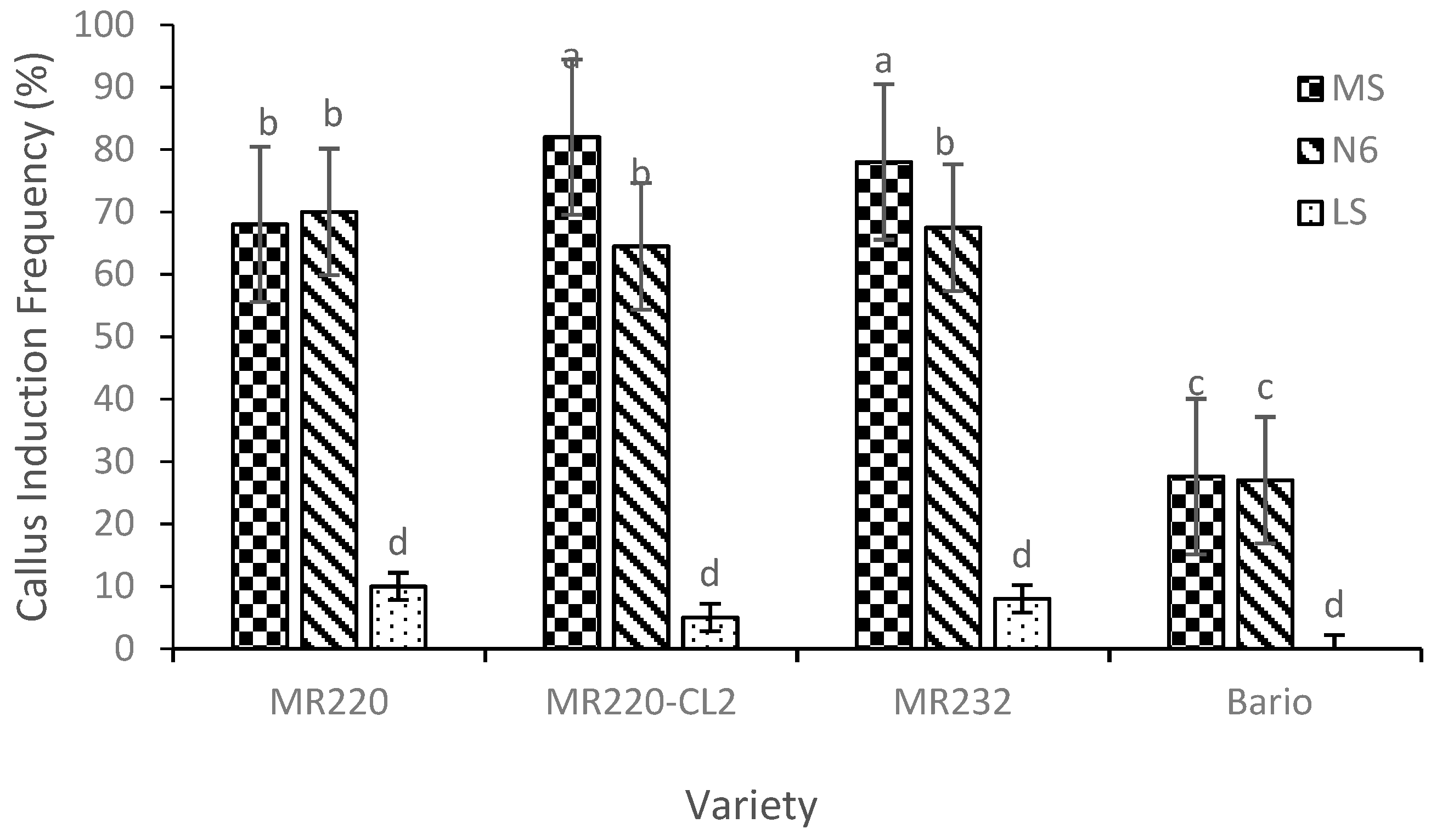
Following the optimization of callus induction percentage by 2,4-D, 3 mg/L of 2,4-D was used in subsequent experiment by using different basal media of MS, N6, and LS (Figure 3). The CIF (%) of different rice varieties from three different basal media showed significant differences (p = 0.001) (Figure 3).
The CIF on MS was higher than that of N6 and LS media for MR220-CL2 and MR232. However, MS and N6 showed differences between MR220 and Bario variety, with a high CIF found in MR220-CL2 (82%) for MS medium. However, callus were not produced for Bario when cultured on LS medium. Hence, the use of LS media was also not preferable in these three wetland varieties which showed the lowest percentage of callus induction (less than 10%). The MR220 callus percentage was almost similar to that when cultured on MS and N6 medium.
Previous work showed that most Malaysian rice callus were induced from MS medium but with different auxin treatment such as reported by Panjaitan et al. [31] and Rahman et al. [3]. From the present study, it was shown that auxin influenced the callus induction frequency but differed among varieties which concurs with a previous reported study [53]. The findings of the present study also showed that responses of different rice varieties were varied and depended on the auxin and basal media used. This statement was supported by Bevitori et al. [62] where they reported that somatic embryogenesis of rice depends upon various factors, such as explant age, donor plant, genotype, nutrient media, auxin concentration, and environmental conditions.
3.1.4. Effect of Carbon Sources
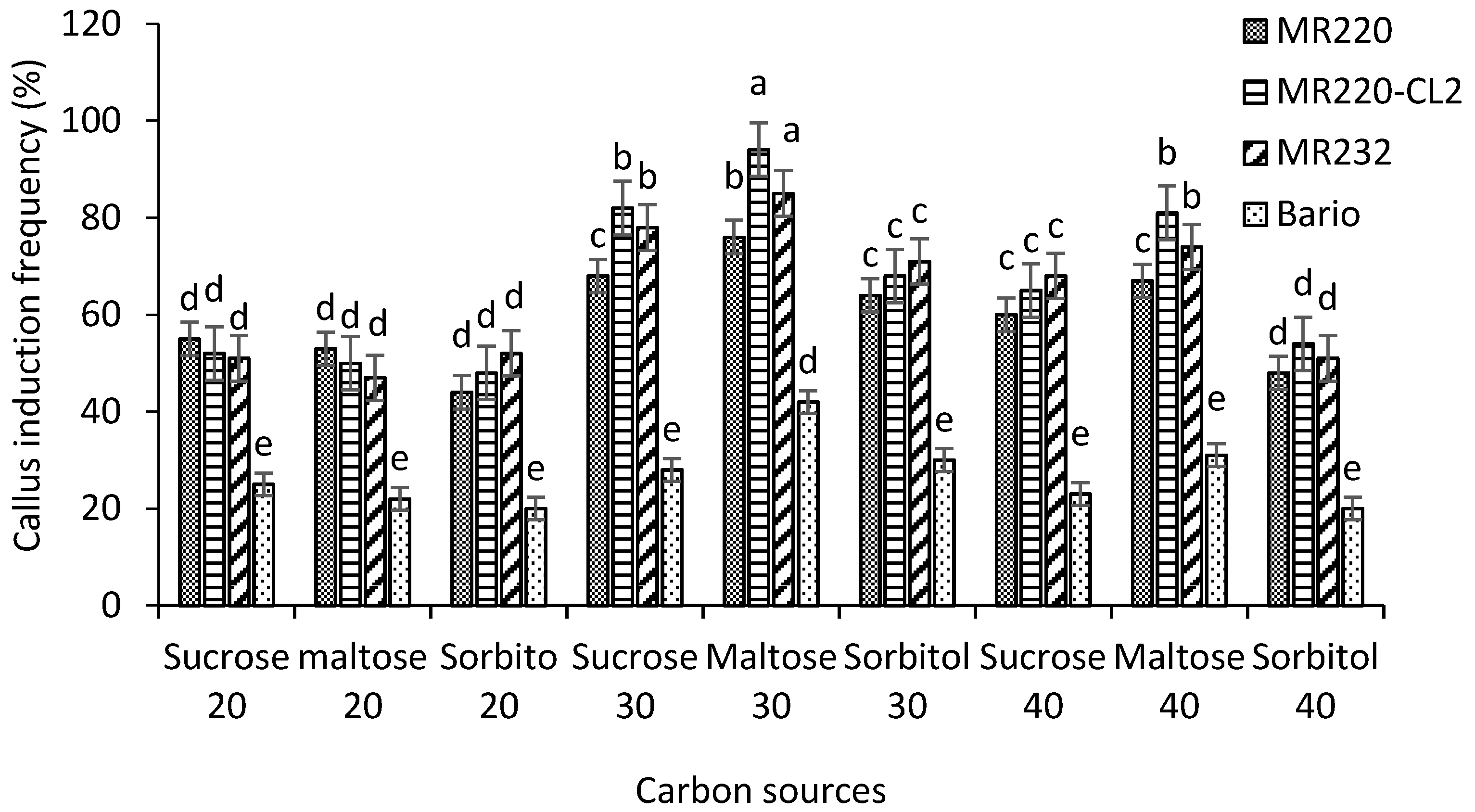
The present study evaluated the CIF on MS media supplemented with 3 mg/L 2,4-D (optimized medium). In this work, sucrose, maltose, and sorbitol were used with different concentration (20, 30, 40 g/L) treatments for all varieties tested. The CIF differed significantly (p = 0.002) between the media (Figure 4). Within different treatments, 30 g/L maltose showed a higher frequency of callus induction compared to other concentrations of sucrose, sorbitol, and maltose in all four varieties of MR220, MR220-CL2, MR232, and Bario (76%, 94%, 85%, and 42% respectively). On the other hand, low CIF was found in 20 g/L sucrose, maltose, sorbitol, and 40 g/L sorbitol in all varieties. The results also showed that decreasing the amount of sucrose and maltose caused a significant reduction in CIF except for sorbitol. Significantly lower CIF was found in sorbitol media for all varieties.
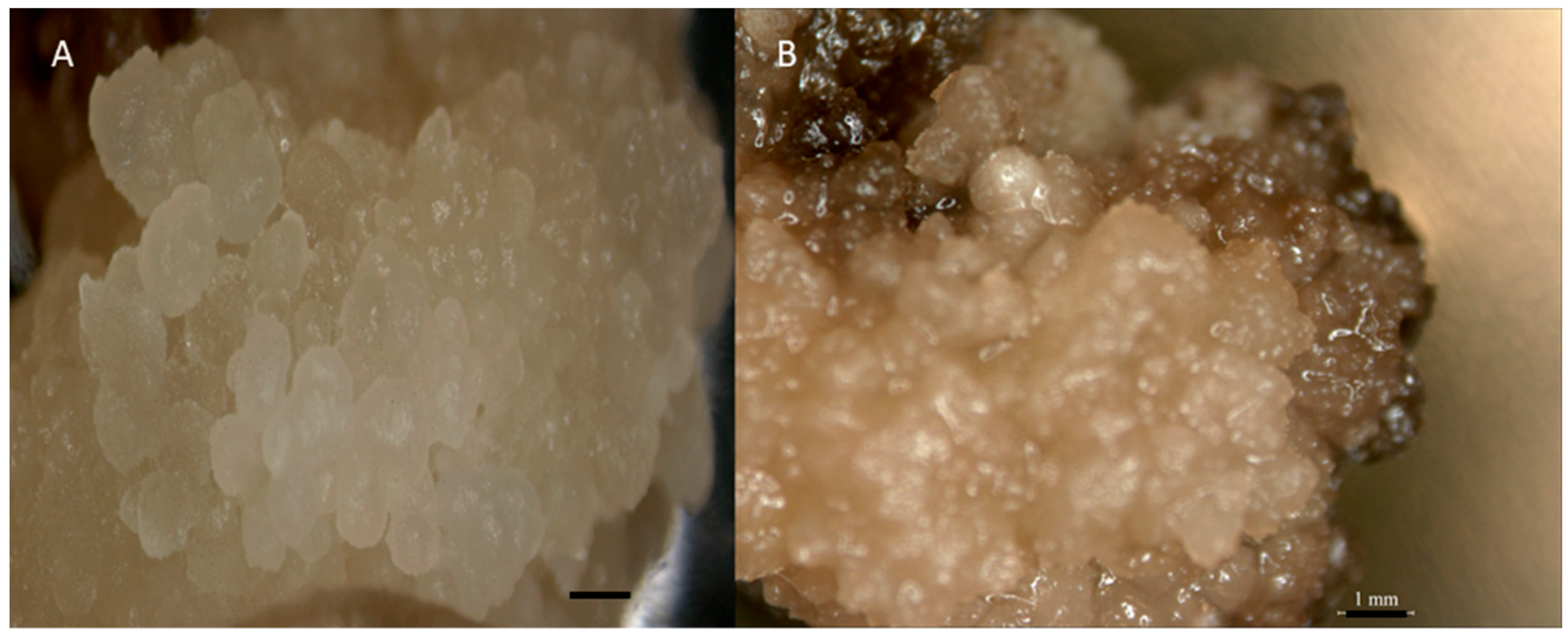
We evaluated the role of carbon sources in controlling excessive production of phenolic compounds from callus on the callusing medium in the case of indica rice. Maltose was found to control phenolic secretion when it was used as a sole carbon source. Carbon source maltose (30 g/L) gave the highest CIF compared to sucrose (30 g/L). The highest CIF was found in MR220-CL2 (94%) in 30 g/L maltose. However, for maltose showed no browning effect after 3 weeks compared to sucrose (Figure 5).
Exogenous carbon sources are the main energy sources for explants because the explant is unable to manufacture its own food, which is called heterotrophism nature [34]. It was also proven that carbohydrate supplementation enhances enzyme activities for callus formation in Oryza sativa indica [32]. Carbon sources are needed in certain genotypes for potential callus induction and shoot organogenesis. Higher mass occurred more from maltose during regeneration then sucrose in the indica rice variety [32]. Previous reports showed that maltose had been used in another culture with profound response but only in certain rice genotypes [63]. Maltose promotes signals to trigger carbohydrate metabolism for higher callus mass formation in indica rice variety MR232 and MR219 [40]. The above study also showed that maltose provides comparatively better development of the green spot of somatic embryos and regeneration plantlet than sucrose in Malaysian aromatic indica rice. There was also a browning effect found in sucrose media. The statement of the previous research supports our studies, where callus browning was observed in sucrose media, but there was no browning effect found in maltose treated media [32].
Maltose (3%) was found to be efficient in controlling phenolic secretion and so it was used for further studies. Extracellular hydrolysis of maltose takes place at a slower rate when compared to sucrose and it is taken up and hydrolyzed more slowly. This activity can be the major reason for the ability of maltose in controlling phenolic secretion in culture medium. The accumulation of toxic products such as phenolic compounds and depletion of nutrients might have led to cell death and eventually a decline in the bio-mass growth [58]. By changing the composition of the media, such as plant growth regulators, the carbon source has a possible effect of improving tissue dissociation [32]. There are some color differences between brown calli and whitish calli: observation revealed that brown callus is unable to regeneration [24,64,65], this is similar to our observation.
The findings of our study were in agreement with the previous works [17,22,41,44,60,66] which stated that indica rice varieties produced the highest callus percentages when cultured on an MS medium supplemented with 2–3 mg/L 2,4-D. Nonetheless, the CIF were between 60% and 87% compared to the present study where they reached up to 94%. Callus induction of rice occurs with the presence of different plant growth regulators in certain nutrient media. However, high callus induction and regeneration may differ in different varieties and media types. In this study, whitish and globular calli were found in maltose treated media and brownish calli were found in sucrose treated media. In parallel to the above observations, Visarada et al. and Abiri et al. [41,49] reported that nodular, compact and whitish calli were highly embryogenic while watery, yellowish, rhizogenic and necrotic calli were stated to be non-embryogenic. Non-embryogenic calli are not competent to regenerate [35]. Based on color and structural morphology, the calli of the present study were mostly embryogenic in maltose treated media. Previous studies illustrated that maltose gave high embryo formation in indica rice [3]. Only embryogenic calli of rice exhibited high frequency regeneration for use in genetic transformation experiments [40,67,68].
3.2. Morphological Characterization
3.2.1. Scanning Electron Microscopy (SEM)
The 3-week old Calli were collected from MS media supplemented with 3 mg/L 2,4-D with 30 g/L maltose for SEM analysis. A scanning electron microscopy (SEM) study was conducted to observe the appearance of the callus epidermal surface with embryogenic calli. From the SEM photo obtained, embryogenic calli confirmed a globular shape and compact structure, formed between cells which were clustered together (Figure 6) for all four varieties i.e., MR220, MR220-CL2, MR232, and Bario.
A similar appearance of embryogenic callus was reported in a previous study conducted by Rahman et al. [40] with different Malaysian rice varieties (MR219) but the developmental ages were older (8 weeks) compare to our study (3 weeks) [69]. In this study, embryogenic structure could be observed after 3 weeks as visualized by SEM. Morphology of embryogenic callus may vary in different varieties. However, in general, most of the structures of the embryogenic calli of different varieties plants and crops examined were of globular form. Similar results have been reported in different plants and crops including O. sativa var. Japonica, Stylosanthes spp. and Cenchrus ciliaris) [62,70,71,72,73]. Bevitori et al. [62] also reported that embryogenic callus of IR 54 rice cultivar was globular shape with tightly packed cells, our observations corroborate with these findings. The result of SEM observation gave evidence of the embryogenic potential of calli obtained in the present study which is in agreement with the findings of Shen et al. [74].
3.2.2. Histology
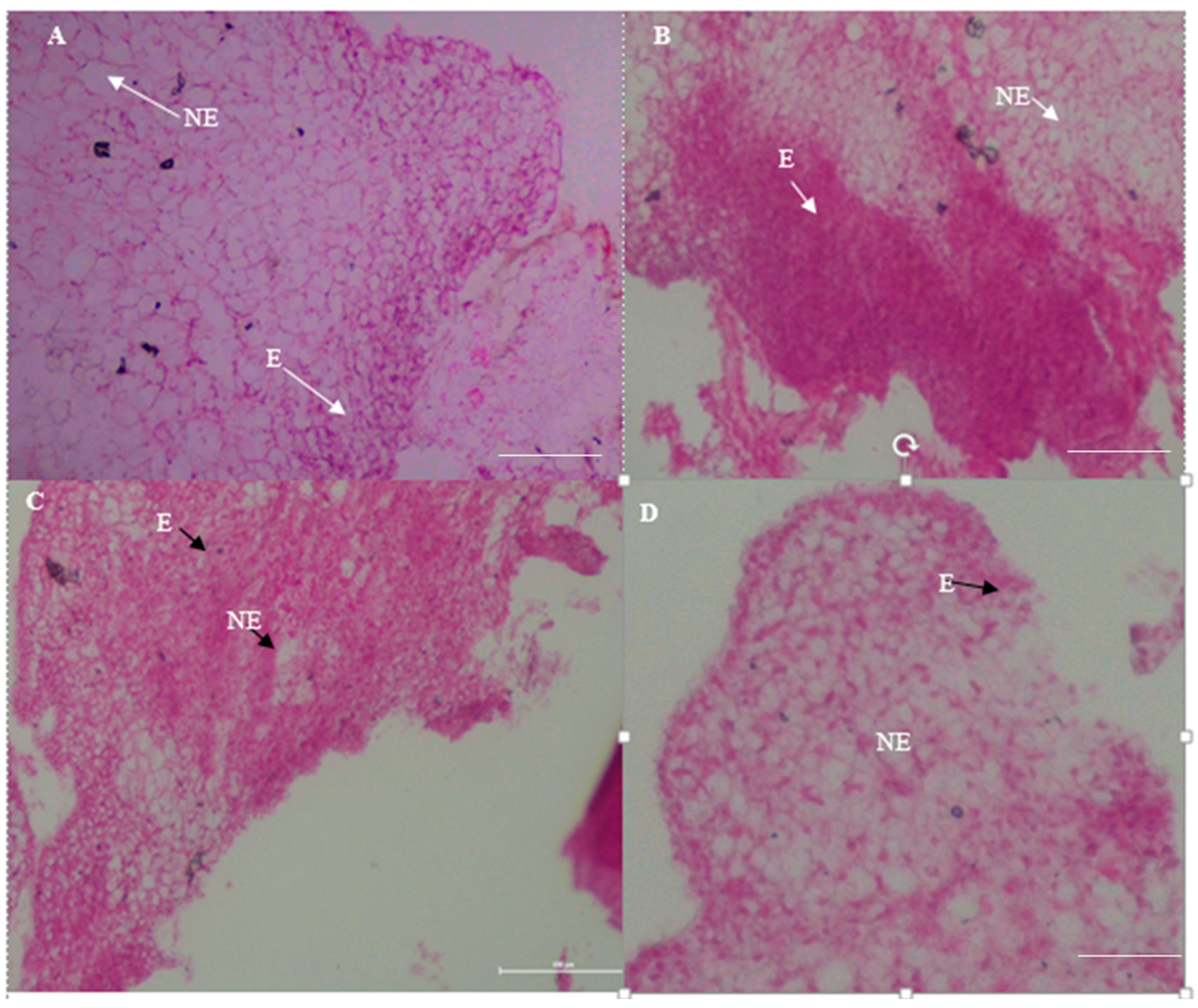
The 3-week old calli from MR220, MR220-CL2, MR232 and Bario varieties cultured on MS media (supplemented by 3 mg/L 2,4-D) were subjected to histological analysis to determine the embryogenic or non-embryogenic nature of the calli. This was accomplished by analyzing the characteristics of the cells’ ultrastructure. This present study showed that embryogenic calli were exhibited as small, stained and compact isodiametric cells while the non-embryogenic calli were shown to be large and highly vacuolated cells (indicated by the arrows) in all varieties (Figure 7). These embryogenic cells have dense cytoplasm, nuclei undergoing uninterrupted cell division with small vacuoles. Moreover, the presence of numerous starch granules intensifying was observed in all varieties which indicated high metabolic activity. However, both non-embryogenic and embryogenic regions were detected in the same callus.
The observations are in agreement with the findings of previous studies where embryogenic calli exposed meristematic cells with high division rates and isodiametric cells with small, and dense cytoplasm [50,75]. Non-embryogenic cells have large vacuoles, a small starch grain and an abundance of intercellular space [62]. A dense cytoplasm with several starch grains found in all embryogenic cells indicates high metabolic activity and the capability to dedifferentiate in order to rise up to embryogenic calli [50,76]. Histological analysis provides information for the ultrastructure of calli for the purpose of selecting embryogenic calli for further regeneration processes [77]. The production of the highest rate of embryogenic calli is very important for developing an efficient regeneration protocol. That, in turn, can be used as a substantial tool for crop improvement and in functional studies of genes [20]. The maximum number of plantlets of Malaysian rice varieties was regenerated from the highest rate of embryogenic calli [78].
3.3. Plant Regeneration
Good quality embryogenic calli were transferred to regeneration medium (RM) in order to investigate their potential for plant regeneration. Calli from the most responsive callus induction medium MS supplemented 3 mg/L 2,4-D with 30 g/L maltose were used for assessing their regeneration potential. The plantlet regeneration frequency and the number of plantlets regenerated are shown in Table 2. Plantlet regeneration (%) and number of plantlets regenerated for four different varieties varied significantly between the regeneration media except for the two varieties viz. MR232 and Bario. The highest plantlet regeneration was found in RM5 and RM6 in the case of all the varieties whereas th highest plantlet regeneration was found in MR220-CL2 (82%) and the lowest in Bario (40%). The number of plantlet was also high in MR220-CL2 and was 9.1 in RM5. A higher number of plantlets was also found in RM5 and RM6 for MR220 (7 and 5, respectively) and MR220-CL2 (9.1 and 6.1, respectively).
In the current investigation, greening of the calli on RM was observed after two weeks of transfer in the case of all varieties. Then the green spots further developed into multiple shoots after 3 weeks, in vitro plantlets, and then developed into healthy plants in the pot (Figure S2). All varieties showed flowering within 45 days except Bario, this is because Bario is upland rice and takes more time (53 days) to flower compared to wetland rice varieties. In the present study, plantlet regeneration percentage was found to increase significantly with increasing concentration of cytokinin (RM5) in the case of all the four rice varieties (Table 2).
NAA was constantly used in all media according to Rahman et al. [3]. These researchers reported that regenerated plantlet was weak without NAA. According to Rueb et al. [35], higher amounts of BAP and Kinetin may stimulate initial cell division and play an important role in somatic embryogenesis and plantlet regeneration. The present study revealed that 2 BAP + 2 Kinetin + 0.5 NAA contributed to the highest plantlet regeneration in all four Malaysian rice varieties. Previous report showed that the use of kinetin helps to improve somatic embryogenesis by influencing mitosis, cytokinesis, total protein synthesis, lignin biosynthesis, vascular differentiation, and differentiation of mature chloroplasts from protoplastids [70].
Morphological features of regenerating callus under the light microscope are presented in Figure 8. Greening of the callus was observed within 21 days on regeneration media and then regenerated shoot and plantlet regeneration were observed in all the four rice varieties.
3.4. Acclimatization
A large number of somatic embryos derived plantlets were successfully acclimatized to ex vitro conditions (Figure S1). The roots of plant were washed and placed in soil in a glass house at 27 ± 2 °C for 10 days. Maximum survival of plants was noticed in the case of MR220-CL2 followed by MR220 and MR232 varieties (Table 3). After 10 days, plants were exposed to natural environmental conditions. It was observed that after 30 days, plants showed a remarkable growth performance. The plants regenerated on RM5 medium showed significantly highest survivability percentage for all four varieties after 2 weeks in the natural environmental conditions.
4. Conclusions
In this study, in vitro callus induction was studied on four selected indica rice (Oryza sativa L.) varieties. Plant growth regulators (i.e., 2,4-D and NAA), carbon sources, and basal media in the cases of the four Malaysian rice varieties were tested for improving callus induction and for plant regeneration frequency. In callus induction studies, with different concentrations of 2,4-D, higher callus induction frequency was observed with 3 mg/L 2,4-D for all wetland rice varieties. The combination of 2,4-D with NAA did not have any significant difference on the callus induction frequency in the cases of MR220, MR220-Cl2, and MR232, whereas 30 g/L maltose showed a higher frequency of callus induction compared to other concentrations of sucrose, sorbitol, and maltose in all the four varieties. The callus induction frequency of MS was higher than that of N6 and LS for MR220-CL2 and MR232. In all the varieties tested, callus induction was poorly induced on LS media. The present study also showed morphological analysis through scanning electron microscopy (SEM) and histological analysis of callus by analyzing the structural appearance of cells and the embryogenic nature of the calli. Regeneration percentage was found to be the highest on RM5. Survival of the plants regenerated on RM5 under natural environmental conditions was the highest. This study showed that the PGR combination of 2 mg/L of BAP + 2 mg/L of Kinetin + 0.5 mg/L of NAA has the potential to promote regeneration of selected indica rice varieties.
Supplementary Materials
The following are available online at https://www.mdpi.com/2073-4395/8/5/77/s1, Figure S1: The morphology of calli on MS media of different concentrations of 2,4-D (1-4 mg/L) of (a) MR220 (b) MR220-CL2 (c) MR232 and (d) Bario, Figure S2: Establishment of MR220, MR220-CL2, MR232 and Bario plant through in vitro callus induction to ripening stage.
Author Contributions
S.B.M. carried out tissue culture, light microscopy, histology, data analysis and wrote the manuscript, A.W. conceived the project, partially wrote the manuscript, and supplied the explant.
Acknowledgments
This tissue culture, scanning electron microscopy, SEM and histology works were performed in the Plant Biotechnology Laboratory at Faculty of Biosciences and Medical Engineering, Universiti Teknologi Malaysia. This work was financially supported by the Tier 1 GUP Grant; Q.J 130000.2545.09H22 under the Ministry of Higher Education Malaysia and thanks to Universiti Teknologi Malaysia for experimental support.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Sandhu, N.; Kumar, A. Bridging the rice yield gaps under drought: QTLs, genes, and their use in breeding programs. Agronomy 2017, 7, 27. [Google Scholar] [CrossRef]
- Zhang, J.; Feng, Q.; Jin, C.; Qiu, D.; Zhang, L.; Xie, K.; Yuan, D.; Han, B.; Zhang, Q.; Wang, S. Features of the expressed sequences revealed by a large-scale analysis of ESTs from a normalized cDNA library of the elite indica rice cultivar Minghui 63. Plant J. 2005, 42, 772–780. [Google Scholar] [CrossRef] [PubMed]
- Rahman, Z.A.; Roowi, S.; Zaliha, W.W.S.; Subramaniam, S. Regeneration of Malaysian indica rice (Oryza sativa) variety MR232 via optimised somatic embryogenesis system. J. Phytol. 2010, 2, 30–38. [Google Scholar]
- Shabery, A. Rice Production at 72% of Self-Sufficiency Level: Ahmad Shabery. Available online: http://www.thesundaily.my/news/2017/09/26/rice-production-72-self-sufficiency-level-ahmad-shabery (accessed on 16 April 2018).
- Kumar, S. Phytochemistry and functional food: The needs of healthy life. J. Phytochem. Biochem. 2017, 1, 2. [Google Scholar]
- Bzour, M.I.; Zuki, F.M.; Mispan, M.S. Introduction of imidazolinone herbicide and Clearfield® rice between weedy rice—Control efficiency and environmental concerns. Environ. Rev. 2018, 1–18. [Google Scholar] [CrossRef]
- Azmi, M.; Azlan, S.; Yim, M.K.; George, V.T.; Chew, E.S. Control of weedy rice in direct-seeded rice using the clearfield production system in Malaysia. Pak. J. Weed Sci. Res. 2012, 18, 49–53. [Google Scholar]
- Zulkarnain, W.M.; Ismail, M.R.; Saud, H.M.; Othman, R.; Habib, S.H.; Kausar, H. Growth and yield response to water availability at different growth stages of rice. J. Food Agric. Environ. 2013, 11, 540–544. [Google Scholar]
- FAOSTAT. Food and Agriculture Organization (FAO) Statistics Division 2017. Available online: http://faostat.fao.org/site/567/default.aspx#ancor (accessed on 10 December 2017).
- Wahab, A.G. Grain and Feed Annual—Kuala Lumpur Malaysia; Global Agricultural Information Network: Kuala Lumpur, Malaysia, 2017. [Google Scholar]
- Fageria, N.K.; Baligar, V.C. Methodology for evaluation of lowland rice genotypes for nitrogen use efficiency. J. Plant Nutr. 2003, 26, 1315–1333. [Google Scholar] [CrossRef]
- Wong, S.C.; Yiu, P.H.; Bong, S.T.W.; Lee, H.H.; Neoh, P.N.P.; Rajan, A. Analysis of Sarawak Bario rice diversity using microsatellite markers. Am. J. Agric. Biol. Sci. 2009, 4, 298–304. [Google Scholar] [CrossRef]
- Nicholas, D.; Hazila, K.K.; Chua, H.P.; Rosniyana, A. Nutritional value and glycemic index of Bario rice varieties. J. Trop. Agric. Food Sci. 2014, 42, 1–8. [Google Scholar]
- FAO; FAOSTAT. Available online: http://www.fao.org/news/archive/news-by-date/2015/en/ (accessed on 5 May 2017).
- Thuy, T.L.; Saitoh, K. Responses of fourteen vietnamese rice (Oryza sativa L.) cultivars to high temperatures during grain filling period under field conditions. Agronomy 2017, 7, 57. [Google Scholar] [CrossRef]
- Terano, R.; Mohamed, Z.; Din, N.S.Z. Determinants of farmers’ adoption of clearfield production system in Malaysia. Agric. Agric. Sci. Procedia 2016, 9, 103–107. [Google Scholar] [CrossRef]
- Libin, A.; King, P.J.H.; Ong, K.H.; Chubo, J.K.; Sipen, P. Callus induction and plant regeneration of Sarawak rice (Oryza sativa L.) variety Biris. Afr. J. Agric. Res. 2012, 7, 4260–4265. [Google Scholar] [CrossRef]
- Kevin, M.T.S.; Ahmed, O.H.; Asrina, W.Y.W.; Rajan, A.; Ahzam, M. Towards growing bario rice on lowland soils: A preliminary nitrogen and potassium fertilization trial. Am. J. Agric. Biol. Sci. 2007, 2, 99–105. [Google Scholar] [CrossRef]
- Naeg, D. Bario Rice—A Rare Grain. Available online: http://www.theborneopost.com/2012/01/29/bario-rice-a-rare-grain/ (accessed on 9 May 2017).
- Gosal, S.S.; Kang, M.S. Plant tissue culture and genetic transformation for crop improvement. In Improving Crop Resistance to Abiotic Stress; Tuteja, N., Gill, S.S., Tiburcio, A.F., Tuteja, R., Eds.; Wiley-VCH Verlag GmbH & Co. KGaA: Weinheim, Germany, 2012; pp. 357–397. ISBN 978-3-527-63293-0. [Google Scholar]
- Wang, L.; Lin, G.; Zhao, D.; Wang, F.; Chen, J. Tissue culture system for different hybrid of indica rice. J. Northeast Agric. Univ. 2011, 18, 13–17. [Google Scholar] [CrossRef]
- Kalhori, N.; Nulit, R.; Go, R.; Zulkifly, S.; Azizi, P.; Abiri, R. Selection, characterizations and somatic embryogenesis of Malaysian salt-tolerant rice (Oryza sativa cv. MR219) through callogenesis. Int. J. Agric. Biol. 2017, 19, 157–163. [Google Scholar] [CrossRef]
- Kumar, S.; Arul, L.; Talwar, D. Generation of marker-free Bt transgenic indica rice and evaluation of its yellow stem borer resistance. J. Appl. Genet. 2010, 51, 243–257. [Google Scholar] [CrossRef] [PubMed]
- Silva, T.D. Indica rice anther culture: Can the impasse be surpassed? Plant Cell Tissue Organ Cult. 2010, 100, 1–11. [Google Scholar] [CrossRef]
- Talapatra, S.; Ghoshal, N.; Raychaudhuri, S.S. Molecular characterization, modeling and expression analysis of a somatic embryogenesis receptor kinase (SERK) gene in Momordica charantia L. during somatic embryogenesis. Plant Cell Tissue Organ Cult. 2013, 116, 271–283. [Google Scholar] [CrossRef]
- Zimmerman, J. Somatic embryogenesis: A model for early development in higher plants. Plant Cell 1993, 5, 1411–1423. [Google Scholar] [CrossRef] [PubMed]
- Wójcikowska, B.; Gaj, M.D. Expression profiling of AUXIN RESPONSE FACTOR genes during somatic embryogenesis induction in Arabidopsis. Plant Cell Rep. 2017, 36, 843–858. [Google Scholar] [CrossRef] [PubMed]
- Karami, O.; Aghavaisi, B.; Pour, A.M. Molecular aspects of somatic-to-embryogenic transition in plants. J. Chem. Biol. 2009, 2, 177–190. [Google Scholar] [CrossRef] [PubMed]
- Datta, S.K. Rice Biotechnology: A Need for Developing Countries. Available online: http://www.agbioforum.org/v7n12/v7n12a06-datta.htm (accessed on 25 September 2017).
- Dievart, A.; Perin, C.; Hirsch, J.; Bettembourg, M.; Lanau, N.; Artus, F.; Bureau, C.; Noel, N.; Droc, G.; Peyramard, M.; et al. The phenome analysis of mutant alleles in Leucine-Rich Repeat Receptor-Like Kinase genes in rice reveals new potential targets for stress tolerant cereals. Plant Sci. 2016, 242, 240–249. [Google Scholar] [CrossRef] [PubMed]
- Panjaitan, S.B.; Abdullah, S.N.A.; Aziz, M.A.; Meon, S.; Omar, O. Somatic embryogenesis from scutellar embryo of Oryza sativa L. var. MR219. J. Trop. Agric. Sci. 2009, 32, 185–194. [Google Scholar]
- Rahman, Z.A.; Ramli, A.; Hosni, H.; Kamaruzaman, R.; Seman, Z.A.; Othman, A.N.; Zainal, Z.; Uddain, J.; Subramaniam, S. Efficient plant regeneration of Malaysian aromatic rice (Oryza sativa L.) through somatic embryogenesis. Emir. J. Food Agric. 2015, 27, 857–863. [Google Scholar] [CrossRef]
- Murashige, T.; Skoog, F. A revised medium for rapid growth and bio assays with tobacco tissue cultures. Physiol. Plant. 1962, 15, 473–497. [Google Scholar] [CrossRef]
- Lee, S.-T.; Huang, W.-L. Cytokinin, auxin, and abscisic acid affects sucrose metabolism conduce to de novo shoot organogenesis in rice (Oryza sativa L.) callus. Bot. Stud. 2013, 54, 5. [Google Scholar] [CrossRef] [PubMed]
- Rueb, S.; Leneman, M.; Schilperoort, R.A.; Hensgens, L.A.M. Efficient plant regeneration through somatic embryogenesis from callus induced on mature rice embryos (Oryza sativa L.). Plant Cell Tissue Organ Cult. 1994, 36, 259–264. [Google Scholar] [CrossRef]
- Kumar, S.; Singh, R.; Kalia, S.; Sharma, S.K.; Kalia, A.K. Recent advances in understanding the role of growth regulators in plant growth and development in vitro-I. conventional growth regulators. Indian For. 2016, 142, 459–470. [Google Scholar]
- Seldimirova, O.A.; Titova, G.E.; Kruglova, N.N. A Complex morpho-histological approach to the in vitro study of morphogenic structures in a wheat anther culture. Biol. Bull. Russ. Acad. Sci. 2016, 43, 121–126. [Google Scholar] [CrossRef]
- Nic-Can, G.I.; Loyola-Vargas, V.M. The role of the auxins during somatic embryogenesis. In Somatic Embryogenesis: Fundamental Aspects and Applications; Springer: Cham, Switzerland, 2016; pp. 171–182. ISBN 978-3-319-33704-3. [Google Scholar]
- Hiei, Y.; Komari, T. Agrobacterium-mediated transformation of rice using immature embryos or calli induced from mature seed. Nat. Protoc. 2008, 3, 824–834. [Google Scholar] [CrossRef] [PubMed]
- Zuraida, A.R.; Naziah, B.; Zamri, Z.; Sreeramanan, S. Efficient plant regeneration of Malaysian indica rice MR 219 and 232 via somatic embryogenesis system. Acta Physiol. Plant. 2011, 33, 1913–1921. [Google Scholar] [CrossRef]
- Abiri, R.; Maziah, M.; Shaharuddin, N.A.; Yusof, Z.N.B.; Atabaki, N.; Hanafi, M.M.; Sahebi, M.; Azizi, P.; Kalhori, N.; Valdiani, A. Enhancing somatic embryogenesis of Malaysian rice cultivar MR219 using adjuvant materials in a high-efficiency protocol. Int. J. Environ. Sci. Technol. 2017, 14, 1091–1108. [Google Scholar] [CrossRef]
- Mohd Din, A.R.J.; Iliyas Ahmad, F.; Wagiran, A.; Abd Samad, A.; Rahmat, Z.; Sarmidi, M.R. Improvement of efficient in vitro regeneration potential of mature callus induced from Malaysian upland rice seed (Oryza sativa cv. Panderas). Saudi J. Biol. Sci. 2016, 23, S69–S77. [Google Scholar] [CrossRef] [PubMed]
- Sahoo, K.K.; Tripathi, A.K.; Pareek, A.; Sopory, S.K.; Singla-Pareek, S.L. An improved protocol for efficient transformation and regeneration of diverse indica rice cultivars. Plant Methods 2011, 7, 49. [Google Scholar] [CrossRef] [PubMed]
- Meneses, A.; Flores, D.; Munoz, M.; Arrieta, G.; Espinoza, A.M. Effect of 2,4-D, hydric stress and light on indica rice (Oryza sativa) somatic embryogenesis. Rev. Biol. Trop. 2005, 53, 361–368. [Google Scholar] [CrossRef] [PubMed]
- Fu, X.; Yang, S.; Bao, M. Factors affecting somatic embryogenesis in anther cultures of Chinese pink (Dianthus chinensis). In Vitro Cell. Dev. Biol.-Plant 2008, 44, 194–202. [Google Scholar] [CrossRef]
- Huang, S.; Hill, R.D.; Wally, O.S.D.; Dionisio, G.; Ayele, B.T.; Jami, S.K.; Stasolla, C. Hemoglobin control of cell survival/death decision regulates in vitro plant embryogenesis. Plant Physiol. 2014, 165, 810–825. [Google Scholar] [CrossRef] [PubMed]
- Fang, C.; Xu, T.; Ye, C.; Huang, L.; Wang, Q.; Lin, W. Method for RNA extraction and cDNA library construction from microbes in crop rhizosphere soil. World J. Microbiol. Biotechnol. 2014, 30, 783–789. [Google Scholar] [CrossRef] [PubMed]
- Alagarsamy, K.; Pandian, S.; Ramesh, M. High frequency plant regeneration from embryogenic callus of popular indica rice (Oryza sativa L.). Physiol. Mol. Biol. Plants 2009, 15, 371–375. [Google Scholar] [CrossRef]
- Visarada, K.B.R.S.; Sailaja, M.; Sarma, N.P. Effect of callus induction media on morphology of embryogenic calli in rice genotypes. Biol. Plant. 2002, 45, 495–502. [Google Scholar] [CrossRef]
- Vega, R.; Vásquez, N.; Espinoza, A.M.; Gatica, A.M.; Valdez-Melara, M. Histology of somatic embryogenesis in rice (Oryza sativa cv. 5272). Rev. Biol. Trop. 2009, 57, 141–150. [Google Scholar]
- Chu, C.C.; Wang, C.C.; Sun, C.S.; Hsu, C.; Yin, K.C.; Chu, C.Y.; Pi, F.Y. Establishment of an efficient medium for anther culture of rice through comparative experiments on the nitrogen sources. Sci. Sin. 1975, 18, 659–668. [Google Scholar]
- Linsmaier, E.M.; Skoog, F. Organic Growth Factor Requirements of Tobacco Tissue Cultures. Physiol. Plant. 1965, 18, 100–127. [Google Scholar] [CrossRef]
- Shahsavari, E.; Maheran, A.A.; Akmar, A.S.N.; Hanafi, M.M. The effect of plant growth regulators on optimization of tissue culture system in Malaysian upland rice. Afr. J. Biotechnol. 2010, 9, 2089–2094. [Google Scholar]
- Liu, Y.M.; Diao, F.Q.; Zhang, L.; Huang, M.J.; Wu, N.H. Cloning and characterization of DcLEA1, a new member of carrot LEA gene family. Prog. Nat. Sci. 2005, 15, 220–228. [Google Scholar]
- Juturu, V.N.; Mekala, G.K.; Garladinne, M.; Reddy, P.C.O.; Sekhar, A.C. Optimization of in vitro regeneration protocol for a popular indica rice (Oryza sativa L. cv Swarna). Ann. Plant Sci. 2016, 5, 1395–1401. [Google Scholar] [CrossRef]
- Naik, N.; Rout, P.; Umakanta, N.; Verma, R.L.; Katara, J.L.; Sahoo, K.K.; Singh, O.N.; Samantaray, S. Development of doubled haploids from an elite indica rice hybrid (BS6444G) using anther culture. Plant Cell Tissue Organ Cult. 2017, 128, 679–689. [Google Scholar] [CrossRef]
- Ghobeishavi, H.; Uliaie, E.D.; Alavikia, S.S.; Valizadeh, M. The effect of plant growth regulators on embryogenic callus induction and regeneration from coleoptile in rice. Int. J. Biosci. 2014, 5, 144–150. [Google Scholar]
- Htwe, N.N.; Maziah, M.; Ling, H.C.; Zaman, F.Q.; Zain, A.M. Responses of some selected Malaysian rice genotypes to callus induction under in vitro salt stress. Afr. J. Biotechnol. 2011, 10, 350–362. [Google Scholar]
- Ge, X.J.; Chu, Z.H.; Lin, Y.J.; Wang, S.P. A tissue culture system for different germplasms of indica rice. Plant Cell Rep. 2006, 25, 392–402. [Google Scholar] [CrossRef] [PubMed]
- Verma, D.; Joshi, R.; Shukla, A.; Kumar, P. Protocol for in vitro somatic embryogenesis and regeneration of rice (Oryza sativa L.). Indian J. Exp. Biol. 2011, 49, 958–963. [Google Scholar] [PubMed]
- Hoang, T.M.L.; Tran, T.N.; Nguyen, T.K.T.; Williams, B.; Wurm, P.; Bellairs, S.; Mundree, S. Improvement of salinity stress tolerance in rice: Challenges and opportunities. Agronomy 2016, 6, 54. [Google Scholar] [CrossRef]
- Bevitori, R.; Popielarska-Konieczna, M.; dos Santos, E.M.; Grossi-de-Sa, M.F.; Petrofeza, S. Morpho-anatomical characterization of mature embryo-derived callus of rice (Oryza sativa L.) suitable for transformation. Protoplasma 2014, 251, 545–554. [Google Scholar] [CrossRef] [PubMed]
- Park, S.G.; Ubaidillah, M.; Kim, K.-M. Effect of maltose concentration on plant regeneration of anther culture with different genotypes in rice (Oryza sativa). Emir. J. Food Agric. 2013, 4, 2265–2270. [Google Scholar] [CrossRef]
- Tamura, S. Shoot formation in calli originated from rice embryo. Proc. Japan Acad. 1968, 44, 544–548. [Google Scholar] [CrossRef]
- Roly, Z.Y.; Islam, M.M.; Shaekh, M.P.E.; Arman, M.S.I.; Shahik, S.M.; Das, D.; Haamem, M.M.E.; Khalekuzzaman, D.M. In Vitro Callus Induction and Regeneration Potentiality of Aromatic Rice (Oryza sativa L.) Cultivars in differential growth regulators. Int. J. Appl. Sci. Biotechnol. 2014, 2, 160–167. [Google Scholar] [CrossRef]
- Kadhimi, A.A.; Zain, C.R.C.M.; Alhasnawi, A.N.; Isahak, A.; Ashraf, M.F.; Mohamad, A.; Doni, F.; Yusoff, W.M.W.; Ibrahim, K.; Badri, K.H.; et al. Impact of gamma rays exposure and growth regulators on Oryza sativa L. cv MR269 callus induction. AIP Conf. Proc. 2016, 1784, 020006. [Google Scholar] [CrossRef]
- Zuo, S.; Zhou, X.; Chen, M.; Zhang, S.; Schwessinger, B.; Ruan, D.; Yuan, C.; Wang, J.; Chen, X.; Ronald, P.C. OsSERK1 regulates rice development but not immunity to Xanthomonas oryzae pv. oryzae or Magnaporthe oryzae. J. Integr. Plant Biol. 2014, 56, 1179–1192. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.; Li, F.; Zhang, C.; Liu, C.; Zhang, X. Differential gene expression of cotton cultivar CCRI24 during somatic embryogenesis. J. Plant Physiol. 2009, 166, 1275–1283. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.-T.; Huang, W.-L. Osmotic stress stimulates shoot organogenesis in callus of rice (Oryza sativa L.) via auxin signaling and carbohydrate metabolism regulation. Plant Growth Regul. 2013, 73, 193–204. [Google Scholar] [CrossRef]
- Abe, T.; Futsuhara, Y. Genotypic variability for callus formation and plant regeneration in rice (Oryza sativa L.). Theor. Appl. Genet. 1986, 72, 3–10. [Google Scholar] [CrossRef] [PubMed]
- Andi Brisibe, E.; Miyake, H.; Taniguchi, T.; Maeda, E. Callus formation and scanning electron microscopy of plantlet regeneration in African rice (Oryza glaberrima steud). Plant Sci. 1992, 83, 217–224. [Google Scholar] [CrossRef]
- Kumar, S.; Chandra, A. In vitro plantlet regeneration in Stylosanthes spp. via callus induction from cotyledonary and hypocotyl explants. Natl. Acad. Sci. Lett.-India 2010, 33, 289–297. [Google Scholar]
- Kumar, S.; Sahu, N.; Singh, A. High-frequency in vitro plant regeneration via callus induction in a rare sexual plant of Cenchrus ciliaris L. In Vitro Cell. Dev. Biol.-Plant 2015, 51, 28–34. [Google Scholar] [CrossRef]
- Shen, H.S.; Sundstøl, F.; Eng, E.R.; Eik, L.O. Studies on untreated and urea-treated rice straw from three cultivation seasons: 3. Histological investigations by light and scanning electron microscopy. Anim. Feed Sci. Technol. 1999, 80, 151–159. [Google Scholar] [CrossRef]
- Bec, S.; Chen, L.L.; Ferriere, N.M.; Legavre, T.; Fauquet, C.; Guiderdoni, E. Comparative histology of microprojectile-mediated gene transfer to embryogenic calli in japonica rice (Oryza sativa L.): Influence of the structural organization of target tissues on genotype transformation ability. Plant Sci. 1998, 138, 177–190. [Google Scholar] [CrossRef]
- Maeda, E.; Radi, S.H. Ultrastructural aspects of rice scutellum as related to seminal root cultures. In Rice; Bajaj, Y.P.S., Ed.; Biotechnology in Agriculture and Forestry; Springer: Berlin/Heidelberg, Germany, 1991; pp. 78–91. ISBN 978-3-642-83988-7. [Google Scholar]
- Oliveira, E.J.; Koehler, A.D.; Rocha, D.I.; Vieira, L.M.; Marques Pinheiro, M.V.; de Matos, E.M.; Ferreira da Cruz, A.C.; Ribeiro da Silva, T.C.; Ossamu Tanaka, F.A.; Silveira Nogueira, F.T.; et al. Morpho-histological, histochemical, and molecular evidences related to cellular reprogramming during somatic embryogenesis of the model grass Brachypodium distachyon. Protoplasma 2017, 254, 2017–2034. [Google Scholar] [CrossRef] [PubMed]
- Azizi, P.; Rafii, M.Y.; Mahmood, M.; Hanafi, M.M.; Abdullah, S.N.A.; Abiri, R.; Sahebi, M. Highly efficient protocol for callogenesis, somagenesis and regeneration of indica rice plants. C. R. Biol. 2015, 338, 463–470. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Calli induction frequency in four indica rice varieties (MR220, Mr220-CL2, MR232, and Bario) supplemented with different concentration of 2,4-D (2, 4-dichlorophenoxyacetic acid) on MS (Murashige and Skoog) basal media after 3 weeks of culture. Mean values of calli induction frequency marked with the same letters do not differ significantly (P ≤ 0.05) in Tukey’s test. Vertical bars represent ± SD (n = 3).
Figure 1.
Calli induction frequency in four indica rice varieties (MR220, Mr220-CL2, MR232, and Bario) supplemented with different concentration of 2,4-D (2, 4-dichlorophenoxyacetic acid) on MS (Murashige and Skoog) basal media after 3 weeks of culture. Mean values of calli induction frequency marked with the same letters do not differ significantly (P ≤ 0.05) in Tukey’s test. Vertical bars represent ± SD (n = 3).

Figure 2.
The callus morphology of MR220 (a), MR220-CL2 (b), MR232 (c), and Bario (d) varieties on MS media supplemented with 3 mg/L of 2,4-D (Bar = 1 mm).
Figure 2.
The callus morphology of MR220 (a), MR220-CL2 (b), MR232 (c), and Bario (d) varieties on MS media supplemented with 3 mg/L of 2,4-D (Bar = 1 mm).

Figure 3.
Callus induction frequency for four Malaysian rice varieties cultured on different basal media MS, N6, and LS media. Mean values marked with the same letters do not differ significantly (p ≤ 0.05) in Tukey’s test. Vertical bars represent ± SD (n = 3).
Figure 3.
Callus induction frequency for four Malaysian rice varieties cultured on different basal media MS, N6, and LS media. Mean values marked with the same letters do not differ significantly (p ≤ 0.05) in Tukey’s test. Vertical bars represent ± SD (n = 3).

Figure 4.
Callus induction frequency of MR220, MR220-Cl2, MR232, and Bario varieties cultured on MS media supplemented with 3 mg/L 2,4-D treated with different carbon sources were evaluated. Mean values marked with the same letters do not differ significantly (p ≤ 0.05) in Tukey’s test. Vertical bars represent ± SD (n = 3).
Figure 4.
Callus induction frequency of MR220, MR220-Cl2, MR232, and Bario varieties cultured on MS media supplemented with 3 mg/L 2,4-D treated with different carbon sources were evaluated. Mean values marked with the same letters do not differ significantly (p ≤ 0.05) in Tukey’s test. Vertical bars represent ± SD (n = 3).

Figure 5.
Callus morphology of 30 g/L Maltose (A) and sucrose (B) on MS media supplemented with 3 mg/L 2,4-D. Bar = 1 mm.
Figure 5.
Callus morphology of 30 g/L Maltose (A) and sucrose (B) on MS media supplemented with 3 mg/L 2,4-D. Bar = 1 mm.

Figure 6.
Scanning electron microscopic observation of callus culture of 4 varieties (A) MR220; (B) MR220-CL2; (C) MR232, and (D) Bario showing the globular and unfolded callus structure to differentiate the cells of the embryogenic callus from the 4 week culture. Bar (A,C,D) =100 µm, B = 200 µm.
Figure 6.
Scanning electron microscopic observation of callus culture of 4 varieties (A) MR220; (B) MR220-CL2; (C) MR232, and (D) Bario showing the globular and unfolded callus structure to differentiate the cells of the embryogenic callus from the 4 week culture. Bar (A,C,D) =100 µm, B = 200 µm.

Figure 7.
(A–D) Histological analysis of calli derived from in vitro tissue cultures on MS media containing 3 mg/L 2,4-D on MS media for (A) MR220; (B) MR220-CL2; (C) MR232, and (D) Bario. Calli inner region containing both small meristematic cells with stained nucleus in the mitotic cell zone indicate embryogenic callus (E) and vacuolated large cells indicate non-embryogenic callus (NE). Magnification 4 × 10, Bar = 100 µm.
Figure 7.
(A–D) Histological analysis of calli derived from in vitro tissue cultures on MS media containing 3 mg/L 2,4-D on MS media for (A) MR220; (B) MR220-CL2; (C) MR232, and (D) Bario. Calli inner region containing both small meristematic cells with stained nucleus in the mitotic cell zone indicate embryogenic callus (E) and vacuolated large cells indicate non-embryogenic callus (NE). Magnification 4 × 10, Bar = 100 µm.

Figure 8.
Plant regeneration through somatic embryogenesis of four Oryza sativa indica rice varieties which were derived from MS medium supplemented with 3 mg/L 2,4-D and 30 g/L maltose. A–D: Morphological features of callus on regeneration media under light microscope after 10 days of regeneration; E–H: green spot initiation after 21 days of regeneration; I–L: regenerated shoot; M–P: plantlet regeneration of MR220, MR220-CL2, MR232, and Bario respectively. Bar: 10 mm (A–L); and Bar: 1 mm (M–P).
Figure 8.
Plant regeneration through somatic embryogenesis of four Oryza sativa indica rice varieties which were derived from MS medium supplemented with 3 mg/L 2,4-D and 30 g/L maltose. A–D: Morphological features of callus on regeneration media under light microscope after 10 days of regeneration; E–H: green spot initiation after 21 days of regeneration; I–L: regenerated shoot; M–P: plantlet regeneration of MR220, MR220-CL2, MR232, and Bario respectively. Bar: 10 mm (A–L); and Bar: 1 mm (M–P).


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Effect of 3 mg/L 2,4-D (2, 4-dichlorophenoxyacetic acid)and different concentrations of NAA (1-naphthalene acetic acid) for callus induction frequency after 3 weeks of culture.
Table 1.
Effect of 3 mg/L 2,4-D (2, 4-dichlorophenoxyacetic acid)and different concentrations of NAA (1-naphthalene acetic acid) for callus induction frequency after 3 weeks of culture.
| Treatment (mg/L) | Variety | ||||
|---|---|---|---|---|---|
| 2,4-D | NAA | MR220 | MR220-CL2 | MR232 | Bario |
| 3.0 | 0.0 | 68 | 82.0 | 78.0 | 28.0 a |
| 3.0 | 2.5 | 70 | 70.3 | 71.0 | 32.8 a |
| 3.0 | 5.0 | 68.0 | 74.3 | 68.5 | 25.0 b |
| 3.0 | 7.5 | 66.0 | 74.0 | 60.0 | 25.6 b |
| 3.0 | 10 | 60.0 | 64.5 | 59.4 | 22.3 c |
| p | NS | NS | NS | 0.001 | |
Mean values (n = 3) marked with the same letters do not differ significantly (p ≤ 0.05) in Tukey’s test. NS indicates as non-significant.
Table 2.
The effect of different concentration (mg/L) of BAP (6-benzylaminopurine) + Kinetin + 0.5 NAA in MS (Murashige and Skoog) medium on plantlet regeneration (%) from the calli of four different Malaysian indica rice varieties.
Table 2.
The effect of different concentration (mg/L) of BAP (6-benzylaminopurine) + Kinetin + 0.5 NAA in MS (Murashige and Skoog) medium on plantlet regeneration (%) from the calli of four different Malaysian indica rice varieties.
| Treatment Name | Treatment (mg/L) | Plantlet Regeneration (%) | Number of Plantlets Regenerated | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| BAP | Kinetin | NAA | MR220 | MR220-CL2 | MR232 | Bario | MR220 | MR220-CL2 | MR232 | Bario | |
| RM1 | 1.0 | 1.0 | 0.5 | 24 c | 27 cd | 16 c | 10 c | 4 b | 4 b | 4.2 | 4.6 |
| RM2 | 2.0 | 1.0 | 0.5 | 28 bc | 38 c | 28.6 b | 14 c | 7 a | 4.1 b | 5.1 | 5.4 |
| RM3 | 3.0 | 1.0 | 0.5 | 34 b | 43 c | 33 b | 24 b | 4.5 b | 5.3 b | 4.2 | 3.9 |
| RM4 | 1.0 | 2.0 | 0.5 | 23 c | 24.1 d | 32 b | 12 c | 4 b | 2.7 c | 5.4 | 3.8 |
| RM5 | 2.0 | 2.0 | 0.5 | 68 a | 82 a | 68 a | 40 a | 7 a | 9.1 a | 6.1 | 3.8 |
| RM6 | 3.0 | 2.0 | 0.5 | 62 a | 71 ab | 70 a | 29 ab | 5 a | 6.1 ab | 4.1 | 4.2 |
| p | 0.041 | 0.001 | 0.034 | 0.045 | 0.001 | 0.001 | N.S. | N.S. | |||
Means followed by different letters were significantly different at p ≤ 0.05 in Tukey’s test (n = 3), calli were induced n MS medium supplemented with 3 mg/L 2,4-D and 30 g/L maltose.
Table 3.
Survival percentage of in vitro raised plantlets from RM5 (2 BAP+ 2 Kinetin+ 0.5 NAA) and RM6 (3 BAP+ 2 Kinetin+ 0.5 NAA) under natural environmental conditions (Malaysian condition) in pots after 2 weeks.
Table 3.
Survival percentage of in vitro raised plantlets from RM5 (2 BAP+ 2 Kinetin+ 0.5 NAA) and RM6 (3 BAP+ 2 Kinetin+ 0.5 NAA) under natural environmental conditions (Malaysian condition) in pots after 2 weeks.
| Variety | MR220 | MR220-CL2 | MR232 | Bario |
|---|---|---|---|---|
| Survival percentage from RM5 | 90.0 a | 92.0 a | 88 a | 84 a |
| Survival percentage from RM6 | 80.0 b | 80.0 b | 70.0 b | 72 b |
| p | 0.001 | 0.002 | 0.001 | 0.009 |
Means (n = 3) with common letters within a column are not significantly different (p ≤ 0.05) in Tukey’s test.
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Binte Mostafiz, S.; Wagiran, A. Efficient Callus Induction and Regeneration in Selected Indica Rice. Agronomy 2018, 8, 77. https://doi.org/10.3390/agronomy8050077
AMA Style
Binte Mostafiz S, Wagiran A. Efficient Callus Induction and Regeneration in Selected Indica Rice. Agronomy. 2018; 8(5):77. https://doi.org/10.3390/agronomy8050077
Chicago/Turabian StyleBinte Mostafiz, Suraiya, and Alina Wagiran. 2018. "Efficient Callus Induction and Regeneration in Selected Indica Rice" Agronomy 8, no. 5: 77. https://doi.org/10.3390/agronomy8050077
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.