Challenges in Using Precision Agriculture to Optimize Symbiotic Nitrogen Fixation in Legumes: Progress, Limitations, and Future Improvements Needed in Diagnostic Testing

Abstract
:1. Introduction
2. Current SNF Diagnostic Methods That Have Potential as Precision Agriculture Tools
2.1. Shoot Nitrogen Status
2.2. Nitrogen Difference Method
2.3. Nitrogen Isotope-Based Methods
3. Current SNF Diagnostic Methods That Require Major Technical Advances to Facilitate Their Application to Precision Agriculture
3.1. Nitrogen Balance Method
3.2. Ureide Assays
3.3. Acetylene Reduction Assay
3.4. Nodulation Traits
3.5. GlnLux Assay
4. Opportunities to Use Precision Agriculture to Diagnose Soil Traits That Affect SNF
4.1. Current Soil Tests That Hold Promise as Precision Agriculture Tools to Optimize SNF
4.1.1. Available Soil Nitrogen
4.1.2. Available Soil Phosphorus
4.1.3. Soil pH
4.1.4. Salinity
4.2. Current Soil Diagnostics That Require Major Technical Advances to Facilitate Their Application as Precision Agriculture Tools for SNF Optimization
4.2.1. Soil Rhizobia Detection
4.2.2. Available Soil Micronutrients
5. Perspectives on the Future of Precision Agriculture to Improve SNF in Field Legumes
Author Contributions
Acknowledgments
Conflicts of Interest
References
- Herridge, D.F.; Peoples, M.B.; Boddey, R.M. Global inputs of biological nitrogen fixation in agricultural systems. Plant Soil 2008, 311, 1–18. [Google Scholar] [CrossRef]
- Ledgard, S.F.; Steele, K.W. Biological nitrogen-fixation in mixed legume grass pastures. Plant Soil 1992, 141, 137–153. [Google Scholar] [CrossRef]
- Dakora, F.D.; Keya, S.O. Contribution of legume nitrogen fixation to sustainable agriculture in sub-saharan Africa. Soil Biol. Biochem. 1997, 29, 809–817. [Google Scholar] [CrossRef]
- Udvardi, M.; Poole, P.S. Transport and metabolism in legume-Rhizobia symbioses. Annu. Rev. Plant Biol. 2013, 64, 781–805. [Google Scholar] [CrossRef] [PubMed]
- Graham, P.H.; Vance, C.P. Legumes: Importance and constraints to greater use. Plant Physiol. 2003, 131, 872–877. [Google Scholar] [CrossRef] [PubMed]
- Foyer, C.H.; Lam, H.; Nguyen, H.T.; Siddique, K.H.M.; Varshney, R.K.; Colmer, T.D.; Cowling, W.; Bramley, H.; Mori, T.A.; Hodgson, J.M.; et al. Neglecting legumes has compromised human health and sustainable food production. Nat. Plants 2016, 2, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Thilakarathna, M.S.; Mcelroy, M.S.; Chapagain, T.; Papadopoulos, Y.A.; Raizada, M.N. Belowground nitrogen transfer from legumes to non-legumes under managed herbaceous cropping systems. A review. Agron. Sustain. Dev. 2016, 36, 58. [Google Scholar] [CrossRef]
- Fustec, J.; Lesuffleur, F.; Mahieu, S.; Cliquet, J.B. Nitrogen rhizodeposition of legumes. A review. Agron. Sustain. Dev. 2010, 30, 57–66. [Google Scholar] [CrossRef]
- Thilakarathna, M.S.; Serran, S.; Lauzon, J.; Janovicek, K.; Deen, B. Management of manure nitrogen using cover crops. Agron. J. 2015, 107, 1595–1607. [Google Scholar] [CrossRef]
- Zahran, H.H. Rhizobium-legume symbiosis and nitrogen fixation under severe conditions and in an arid climate. Microbiol. Mol. Biol. Rev. 1999, 63, 968–989. [Google Scholar] [PubMed]
- Giller, K.E. Nitrogen Fixation in Tropical Cropping Systems; CABI: Wallingford, UK, 2001. [Google Scholar]
- Hungria, M.; Vargas, M.A.T. Environmental factors affecting N2 fixation in grain legumes in the tropics, with an emphasis on Brazil. Field Crop. Res. 2000, 65, 151–164. [Google Scholar] [CrossRef]
- Thilakarathna, M.S.; Raizada, M.N. A meta-analysis of the effectiveness of diverse rhizobia inoculants on soybean traits under field conditions. Soil Biol. Biochem. 2017, 105, 177–196. [Google Scholar] [CrossRef]
- Walsh, K.B. Physiology of the legume nodule and its response to stress. Soil Biol. Biochem. 1995, 27, 637–655. [Google Scholar] [CrossRef]
- Graham, P.H. Some problems of nodulation and symbiotic nitrogen fixation in Phaseolus vulgaris L.: A review. Field Crop. Res. 1981, 4, 93–112. [Google Scholar] [CrossRef]
- Aranjuelo, I.; Arrese-Igor, C.; Molero, G. Nodule performance within a changing environmental context. J. Plant Physiol. 2014, 171, 1076–1090. [Google Scholar] [CrossRef] [PubMed]
- McBratney, A.B.; Whelan, B.; Ancev, T.; Bouma, J. Future directions of precision agriculture. Precis. Agric. 2005, 6, 7–23. [Google Scholar] [CrossRef]
- Gebbers, R.; Adamchuk, V.I. Precision agriculture and food security. Science 2010, 327, 828–830. [Google Scholar] [CrossRef] [PubMed]
- Hedley, C. The role of precision agriculture for improved nutrient management on farms. J. Sci. Food Agric. 2015, 95, 12–19. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Kovacs, J.M. The application of small unmanned aerial systems for precision agriculture: A review. Precis. Agric. 2012, 13, 693–712. [Google Scholar] [CrossRef]
- Zhang, N.; Wang, M.; Wang, N. Precision agriculture—A worldwide overview. Comput. Electron. Agric. 2002, 36, 113–132. [Google Scholar] [CrossRef]
- Franzen, D.; Long, D.; Sims, A.; Lamb, J.; Casey, F.; Staricka, J.; Halvorson, M.; Hofman, V. Evaluation of methods to determine residual soil nitrate zones across the northern Great Plains of the USA. Precis. Agric. 2011, 12, 594–606. [Google Scholar] [CrossRef]
- Shanahan, J.F.; Kitchen, N.R.; Raun, W.R.; Schepers, J.S. Responsive in-season nitrogen management for cereals. Comput. Electron. Agric. 2008, 61, 51–62. [Google Scholar] [CrossRef]
- Schmidt, J.; Beegle, D.; Zhu, Q.; Sripada, R. Improving in-season nitrogen recommendations for maize using an active sensor. Field Crop. Res. 2011, 120, 94–101. [Google Scholar] [CrossRef]
- Samborski, S.M.; Gozdowski, D.; Stepień, M.; Walsh, O.S.; Leszczyńska, E. On-farm evaluation of an active optical sensor performance for variable nitrogen application in winter wheat. Eur. J. Agron. 2016, 74, 56–67. [Google Scholar] [CrossRef]
- Yost, M.A.; Kitchen, N.R.; Sudduth, K.A.; Sadler, E.J.; Drummond, S.T.; Volkmann, M.R. Long-term impact of a precision agriculture system on grain crop production. Precis. Agric. 2017, 18, 823–842. [Google Scholar] [CrossRef]
- Peoples, M.B.; Unkovich, M.J.; Herridge, D.F. Measuring Symbiotic Nitrogen Fixation by Legumes. In Nitrogen Fixation in Crop Production; Emerich, D.W., Krishnan, H.B., Eds.; American Society of Agronomy, Crop Science Society of America, Soil Science Society of America: Madison, WI, USA, 2009; pp. 125–127. [Google Scholar]
- Unkovich, M.; Herridge, D.; Peoples, M.; Cadisch, G.; Boddey, B.; Giller, K.; Alves, B.; Chalk, P. Measuring Plant-Associated Nitrogen Fixation in Agricultural Systems; Australian Centre for International Agricultural Research: Canberra, Australia, 2008.
- Hardarson, G.; Danso, S.K.A. Methods for measuring biological nitrogen fixation in grain legumes. Plant Soil 1993, 152, 19–23. [Google Scholar] [CrossRef]
- Howieson, J.G.; Dilworth, M.J. Working with Rhizobia; Australian Centre for International Agricultural Research: Canberra, Australia, 2016.
- Thilakarathna, M.S.; Moroz, N.; Raizada, M.N. A biosensor-based leaf punch assay for glutamine correlates to symbiotic nitrogen fixation measurements in legumes to permit rapid screening of rhizobia inoculants under controlled conditions. Front. Plant Sci. 2017, 8, 1714. [Google Scholar] [CrossRef] [PubMed]
- Filella, I.; Serrano, L.; Serra, J.; Peñuelas, J. Evaluating wheat nitrogen status with canopy reflectance indexes and discriminant-analysis. Crop Sci. 1995, 35, 1400–1405. [Google Scholar] [CrossRef]
- Herridge, D.F. Relative abundance of ureides and nitrate in plant tissues of soybean as a quantitative assay of nitrogen fixation. Plant Physiol. 1982, 70, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Stewart, B.W.D.P.; Fitzgerald, G.P.; Burris, R.H. In situ studies on N2 fixation using the acetylene reduction technique. Proc. Natl. Acad. Sci. USA 1967, 58, 2071–2078. [Google Scholar] [CrossRef] [PubMed]
- Lira, M.A., Jr.; Smith, D.L. Use of a standard TWAIN scanner and software for nodule number determination on different legume species. Soil Biol. Biochem. 2000, 32, 1463–1467. [Google Scholar] [CrossRef]
- Jordan, D.C.; Garrard, E.H. Studies on the legume root nodule bacteria. Can. J. Bot. 1951, 29, 361–372. [Google Scholar] [CrossRef]
- Remmler, L.; Clairmont, L.; Rolland-Lagan, A.G.; Guinel, F.C. Standardized mapping of nodulation patterns in legume roots. New Phytol. 2014, 202, 1083–1094. [Google Scholar] [CrossRef] [PubMed]
- Jaramillo, P.M.D.; Guimarães, A.A.; Florentino, L.A.; Silva, K.B.; Nóbrega, R.S.A.; Moreira, F.M.S. Symbiotic nitrogen-fixing bacterial populations trapped from soils under agroforestry systems in the Western Amazon. Sci. Agric. 2013, 70, 397–404. [Google Scholar] [CrossRef]
- Muñoz-Huerta, R.F.; Guevara-Gonzalez, R.G.; Contreras-Medina, L.M.; Torres-Pacheco, I.; Prado-Olivarez, J.; Ocampo-Velazquez, R.V. A review of methods for sensing the nitrogen status in plants: advantages, disadvantages and recent advances. Sensors 2013, 13, 10823–10843. [Google Scholar] [CrossRef] [PubMed]
- Tremblay, N.; Wang, Z.; Cerovic, Z.G. Sensing crop nitrogen status with fluorescence indicators. A review. Agron. Sustain. Dev. 2012, 32, 451–464. [Google Scholar] [CrossRef]
- Peñuelas, J.; Filella, L. Visible and near-infrared reflectance techniques for diagnosing plant physiological status. Trends Plant Sci. 1998, 3, 151–156. [Google Scholar] [CrossRef]
- Hansen, P.M.; Schjoerring, J.K. Reflectance measurement of canopy biomass and nitrogen status in wheat crops using normalized difference vegetation indices and partial least squares regression. Remote Sens. Environ. 2003, 86, 542–553. [Google Scholar] [CrossRef]
- Li, F.; Miao, Y.; Feng, G.; Yuan, F.; Yue, S.; Gao, X.; Liu, Y.; Liu, B.; Ustin, S.L.; Chen, X. Improving estimation of summer maize nitrogen status with red edge-based spectral vegetation indices. Field Crop. Res. 2014, 157, 111–123. [Google Scholar] [CrossRef]
- Sharma, L.K.; Bu, H.; Denton, A.; Franzen, D.W. Active-optical sensors using red NDVI compared to red edge NDVI for prediction of corn grain yield in north Dakota, U.S.A. Sensors 2015, 15, 27832–27853. [Google Scholar] [CrossRef] [PubMed]
- Tahmasbian, I.; Xu, Z.; Abdullah, K.; Zhou, J.; Esmaeilani, R.; Nguyen, T.T.N.; Hosseini Bai, S. The potential of hyperspectral images and partial least square regression for predicting total carbon, total nitrogen and their isotope composition in forest litterfall samples. J. Soils Sediments 2017, 17, 2091–2103. [Google Scholar] [CrossRef]
- Yi, Q.-X.; Huang, J.-F.; Wang, F.-M.; Wang, X.-Z.; Liu, Z.-Y. Monitoring rice nitrogen status using hyperspectral reflectance and artificial neural network. Environ. Sci. Technol. 2007, 41, 6770–6775. [Google Scholar] [CrossRef] [PubMed]
- Knyazikhin, Y.; Schull, M.A.; Stenberg, P.; Mottus, M.; Rautiainen, M.; Yang, Y.; Marshak, A.; Latorre Carmona, P.; Kaufmann, R.K.; Lewis, P.; et al. Hyperspectral remote sensing of foliar nitrogen content. Proc. Natl. Acad. Sci. USA 2013, 110, E185–E192. [Google Scholar] [CrossRef] [PubMed]
- Smith, M.L.; Martin, M.E.; Plourde, L. Analysis of hyperspectral data for estimation of temperate forest canopy nitrogen concentration: Comparison between an airborne (AVIRIS) and a spaceborne (Hyperion) sensors. Geosci. Remote Sens. 2003, 41, 1332–1337. [Google Scholar] [CrossRef]
- Matese, A.; Toscano, P.; Gennaro, S.F.D.; Genesio, L.; Vaccari, F.P.; Primicerio, J.; Belli, C.; Zaldei, A.; Bianconi, R.; Gioli, B. Intercomparison of UAV, aircraft and satellite remote sensing platforms for precision viticulture. Remote Sens. 2015, 7, 2971–2990. [Google Scholar] [CrossRef]
- Erdle, K.; Mistele, B.; Schmidhalter, U. Comparison of active and passive spectral sensors in discriminating biomass parameters and nitrogen status in wheat cultivars. Field Crop. Res. 2011, 124, 74–84. [Google Scholar] [CrossRef]
- Solari, F.; Shanahan, J.; Ferguson, R.; Schepers, J.; Gitelson, A. Active sensor reflectance measurements of corn nitrogen status and yield potential. Agron. J. 2008, 100, 571–579. [Google Scholar] [CrossRef]
- Hardin, P.J.; Jensen, R.R. Small-scale unmanned aerial vehicles in environmental remote sensing: Challenges and opportunities. GISci. Remote Sens. 2011, 48, 99–111. [Google Scholar] [CrossRef]
- Gil-Quintana, E.; Larrainzar, E.; Arrese-Igor, C.; González, E.M. Is N-feedback involved in the inhibition of nitrogen fixation in drought-stressed Medicago truncatula? J. Exp. Bot. 2012, 63, 281–292. [Google Scholar] [CrossRef]
- Liese, R.; Schulze, J.; Cabeza, R.A. Nitrate application or P deficiency induce a decline in Medicago truncatula N2-fixation by similar changes in the nodule transcriptome. Sci. Rep. 2017, 7, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Hellmann, C.; Große-Stoltenberg, A.; Lauströ, V.; Oldeland, J.; Werner, C. Retrieving nitrogen isotopic signatures from fresh leaf reflectance spectra: disentangling δ15N from biochemical and structural leaf properties. Front. Plant Sci. 2015, 6, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Elmore, A.J.; Craine, J.M. Spectroscopic analysis of canopy nitrogen and nitrogen isotopes in managed pastures and hay land. IEEE Trans. Geosci. Remote Sens. 2011, 49, 2491–2498. [Google Scholar] [CrossRef]
- Wang, L.; Okin, G.S.; Wang, J.; Epstein, H.; Macko, S.A. Predicting leaf and canopy 15N compositions from reflectance spectra. Geophys. Res. Lett. 2007, 34, 2–5. [Google Scholar] [CrossRef]
- Jin, X.; Song, K.; Du, J.; Liu, H.; Wen, Z. Comparison of different satellite bands and vegetation indices for estimation of soil organic matter based on simulated spectral configuration. Agric. For. Meteorol. 2017, 244–245, 57–71. [Google Scholar] [CrossRef]
- Tegeder, M. Transporters involved in source to sink partitioning of amino acids and ureides: Opportunities for crop improvement. J. Exp. Bot. 2014, 65, 1865–1878. [Google Scholar] [CrossRef] [PubMed]
- Goos, R.J.; Abdraimova, N.; Johnson, B.E. Method for determination of ureides in soybean tissues. Commun. Soil Sci. Plant Anal. 2015, 46, 424–429. [Google Scholar] [CrossRef]
- Ladrera, R.; Marino, D.; Larrainzar, E.; González, E.M.; Arrese-Igor, C. Reduced carbon availability to bacteroids and elevated ureides in nodules, but not in shoots, are involved in the nitrogen fixation response to early drought in soybean. Plant Physiol. 2007, 145, 539–546. [Google Scholar] [CrossRef] [PubMed]
- Nouwen, N.; Arrighi, J.F.; Cartieaux, F.; Chaintreuil, C.; Gully, D.; Klopp, C.; Giraud, E. The role of rhizobial (NifV) and plant (FEN1) homocitrate synthases in Aeschynomene/photosynthetic Bradyrhizobium symbiosis. Sci. Rep. 2017, 7, 448. [Google Scholar] [CrossRef] [PubMed]
- Riar, M.K.; Cerezini, P.; Manandhar, A.; Sinclair, T.R.; Li, Z.; Carter, T.E. Expression of drought-tolerant N fixation in heterogeneous inbred families derived from PI471938 and hutcheson soybean. Crop Sci. 2018, 58, 364–369. [Google Scholar] [CrossRef]
- Cassar, N.; Tang, W.; Gabathuler, H.; Huang, K. A method for high frequency underway N fixation measurements: Flow-through incubation Acetylene Reduction Assays by Cavity ring down laser Absorption Spectroscopy (FARACAS). Anal. Chem. 2018. acs.analchem.7b04977. [Google Scholar] [CrossRef] [PubMed]
- Unkovich, M.J.; Pate, J.S. An appraisal of recent field measurements of symbiotic N2 fixation by annual legumes. Field Crops. Res. 2000, 65, 211–228. [Google Scholar] [CrossRef]
- Lodwig, E.M.; Hosie, A.H.F.; Bourdès, A.; Findlay, K.; Allaway, D.; Karunakaran, R.; Downie, J.A.; Poole, P.S. Amino-acid cycling drives nitrogen fixation in the legume—Rhizobium symbiosis. Nature 2006, 422, 722–726. [Google Scholar] [CrossRef] [PubMed]
- Zablotowicz, R.M.; Reddy, K.N. Nitrogenase activity, nitrogen content, and yield responses to glyphosate in glyphosate-resistant soybean. Crop Prot. 2007, 26, 370–376. [Google Scholar] [CrossRef]
- Metzner, R.; Eggert, A.; van Dusschoten, D.; Pflugfelder, D.; Gerth, S.; Schurr, U.; Uhlmann, N.; Jahnke, S. Direct comparison of MRI and X-ray CT technologies for 3D imaging of root systems in soil: Potential and challenges for root trait quantification. Plant Methods 2015, 11, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mairhofer, S.; Sturrock, C.; Wells, D.M.; Bennett, M.J.; Mooney, S.J.; Pridmore, T.P. On the evaluation of methods for the recovery of plant root systems from X-ray computed tomography images. Funct. Plant Biol. 2014, 42, 460–470. [Google Scholar] [CrossRef]
- Sturrock, C.J.; Woodhall, J.; Brown, M.; Walker, C.; Mooney, S.J.; Ray, R.V. Effects of damping-off caused by Rhizoctonia solani anastomosis group 2-1 on roots of wheat and oil seed rape quantified using X-ray Computed Tomography and real-time PCR. Front. Plant Sci. 2015, 6, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Goron, T.L.; Raizada, M.N. Current and future transgenic whole-cell biosensors for plant macro- and micronutrients. Crit. Rev. Plant Sci. 2014, 33, 392–413. [Google Scholar] [CrossRef]
- Moser, I.; Jobst, G.; Urban, G.A. Biosensor arrays for simultaneous measurement of glucose, lactate, glutamate, and glutamine. Biosens. Bioelectron. 2002, 17, 297–302. [Google Scholar] [CrossRef]
- Röhlen, D.L.; Pilas, J.; Schöning, M.J.; Selmer, T. Development of an amperometric biosensor platform for the combined determination of l-Malic, Fumaric, and l-Aspartic cid. Appl. Biochem. Biotechnol. 2017, 183, 566–581. [Google Scholar] [CrossRef] [PubMed]
- Deangelis, K.M.; Ji, P.; Firestone, M.K.; Lindow, S.E. Two novel bacterial biosensors for detection of nitrate availability in the rhizosphere. Appl. Environ. Microbiol. 2005, 71, 8537–8547. [Google Scholar] [CrossRef] [PubMed]
- Olfs, H.W.; Blankenau, K.; Brentrup, F.; Jasper, J.; Link, A.; Lammel, J. Soil- and plant-based nitrogen-fertilizer recommendations in arable farming. J. Plant Nutr. Soil Sci. 2005, 168, 414–431. [Google Scholar] [CrossRef]
- Missouri Agricultural Experiment Station (MAES). Recommended Chemical Soil Test Procedures for the North Central Region; North Central Regional Research Publication No. 221 (Revised); Missouri Agricultural Experiment Station, University of Missouri—Columbia: Columbia, MO, USA, 1999. [Google Scholar]
- Okalebo, J.R.; Gathua, K.W.; Woomer, P.L. Laboratory Methods of Soil and Plant Analysis: A Working Manual, 2nd ed.; Tropical Soil Biology and Fertility Programme: Nairobi, Kenya, 2002; ISBN 9966989218. [Google Scholar]
- Sanchez, C.A. Phosphorus. In Handbook of Plant Nutrition; Barker, A.V., Pilbeam, D.J., Eds.; CRC Press: Boca Raton, FL, USA, 2007; pp. 51–90. [Google Scholar]
- Matthiesen, H. In situ measurement of soil pH. J. Archaeol. Sci. 2004, 31, 1373–1381. [Google Scholar] [CrossRef]
- Schofield, R.K.; Taylor, A.W. The measurement of soil pH. Soil Sci. Soc. Proc. 1955, 19, 164–167. [Google Scholar] [CrossRef]
- Corwin, D.L.; Lesch, S.M. Apparent soil electrical conductivity measurements in agriculture. Comput. Electron. Agric. 2005, 46, 11–43. [Google Scholar] [CrossRef]
- Rhoades, J.D.; Fernando, C. Soil Salinity Assessment: Methods and Interpretation of Electrical Conductivity Measurements; Food & Agriculture Org.: Rome, Italy, 1999; Volume 57. [Google Scholar]
- Hamlin, R.L. Molybdenum. In Handbook of Plant Nutrition; Barker, A.V., Pilbeam, D.J., Eds.; CRC Press: Boca Raton, FL, USA, 2007; pp. 375–409. [Google Scholar]
- Gupta, U.C. Boron. In Handbook of Plant Nutrition; Barker, A.V., Pilbeam, D.J., Eds.; CRC Press: Boca Raton, FL, USA, 2007; pp. 241–277. [Google Scholar]
- Salvagiotti, F.; Cassman, K.G.; Specht, J.E.; Walters, D.T.; Weiss, A.; Dobermann, A. Nitrogen uptake, fixation and response to fertilizer N in soybeans: A review. Field Crop. Res. 2008, 108, 1–13. [Google Scholar] [CrossRef]
- Sahrawat, K.L. Factors affecting nitrification in soils. Commun. Soil Sci. Plant Anal. 2008, 39, 1436–1446. [Google Scholar] [CrossRef]
- Adamchuk, V.I.; Hummel, J.W.; Morgan, M.T.; Upadhyaya, S.K. On-the-go soil sensors for precision agriculture. Comput. Electron. Agric. 2004, 44, 71–91. [Google Scholar] [CrossRef]
- Sinfield, J.V.; Fagerman, D.; Colic, O. Evaluation of sensing technologies for on-the-go detection of macro-nutrients in cultivated soils. Comput. Electron. Agric. 2010, 70, 1–18. [Google Scholar] [CrossRef]
- Divito, G.A.; Sadras, V.O. How do phosphorus, potassium and sulphur affect plant growth and biological nitrogen fixation in crop and pasture legumes? A meta-analysis. Field Crop. Res. 2014, 156, 161–171. [Google Scholar] [CrossRef]
- Valentine, A.J.; Kleinert, A.; Benedito, V.A. Adaptive strategies for nitrogen metabolism in phosphate deficient legume nodules. Plant Sci. 2017, 256, 46–52. [Google Scholar] [CrossRef] [PubMed]
- Wittry, D.J.; Mallarino, A.P. Comparison of uniform-and variable-rate phosphorus fertilization for corn-soybean rotations. Agron. J. 2004, 96, 26–33. [Google Scholar] [CrossRef]
- Weisz, R.; Heiniger, R.; White, J.G.; Knox, B.; Reed, L. Long-term variable rate lime and phosphorus application for piedmont no-till field crops. Precis. Agric. 2003, 4, 311–330. [Google Scholar] [CrossRef]
- Maleki, M.R.; Mouazen, A.M.; De Ketelaere, B.; Ramon, H.; De Baerdemaeker, J. On-the-go variable-rate phosphorus fertilisation based on a visible and near-infrared soil sensor. Biosyst. Eng. 2008, 99, 35–46. [Google Scholar] [CrossRef]
- Mahajan, G.R.; Pandey, R.N.; Sahoo, R.N.; Gupta, V.K.; Datta, S.C.; Kumar, D. Monitoring nitrogen, phosphorus and sulphur in hybrid rice (Oryza sativa L.) using hyperspectral remote sensing. Precis. Agric. 2017, 18, 736–761. [Google Scholar] [CrossRef]
- Moonrungsee, N.; Pencharee, S.; Jakmunee, J. Colorimetric analyzer based on mobile phone camera for determination of available phosphorus in soil. Talanta 2015, 136, 204–209. [Google Scholar] [CrossRef] [PubMed]
- Adamchuk, V.I.; Lund, E.D.; Reed, T.M.; Ferguson, R.B. Evaluation of an on-the-go technology for soil pH mapping. Precis. Agric. 2007, 8, 139–149. [Google Scholar] [CrossRef]
- Wang, Y.; Huang, T.; Liu, J.; Lin, Z.; Li, S.; Wang, R.; Ge, Y. Soil pH value, organic matter and macronutrients contents prediction using optical diffuse reflectance spectroscopy. Comput. Electron. Agric. 2015, 111, 69–77. [Google Scholar] [CrossRef]
- Mouazen, A.M.; Maleki, M.R.; De Baerdemaeker, J.; Ramon, H. On-line measurement of some selected soil properties using a VIS-NIR sensor. Soil Tillage Res. 2007, 93, 13–27. [Google Scholar] [CrossRef]
- Tekin, Y.; Kuang, B.; Mouazen, A.M. Potential of on-line visible and near infrared spectroscopy for measurement of pH for deriving variable rate lime recommendations. Sensors 2013, 13, 10177–10190. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bianchini, A.A.; Mallarino, A.P. Soil-sampling alternatives and variable-rate liming for a soybean-corn rotation. Agron. J. 2002, 94, 1355–1366. [Google Scholar] [CrossRef]
- Pierce, F.J.; Warncke, D.D. Soil and crop response to variable-rate liming for two michigan fields. Soil Sci. Soc. Am. J. 2000, 64, 774. [Google Scholar] [CrossRef]
- Pitman, M.G.; Läuchli, A. Global impact of salinity and agricultural ecosystems. In Salinity: Environment—Plants—Molecules; Läuchli, A., Lüttge, U., Eds.; Springer: Dordrecht, The Netherlands, 2002; pp. 3–20. ISBN 978-1-4020-0492-6. [Google Scholar]
- Allbed, A.; Kumar, L. Soil salinity mapping and monitoring in arid and semi-arid regions using remote sensing technology: A review. Adv. Remote Sens. 2013, 2, 373–385. [Google Scholar] [CrossRef]
- Thies, J.E.; Singleton, P.W.; Bohlool, B.B. Influence of the size of indigenous rhizobial populations on establishment and symbiotic performance of introduced rhizobia on field-grown legumest. Appl. Environ. Microbiol. 1991, 57, 19–28. [Google Scholar] [PubMed]
- Deaker, R.; Roughley, R.J.; Kennedy, I.R. Legume seed inoculation technology—A review. Soil Biol. Biochem. 2004, 36, 1275–1288. [Google Scholar] [CrossRef]
- Chidebe, I.N.; Jaiswal, S.K.; Dakora, F.D. Distribution and phylogeny of microsymbionts associated with cowpea (Vigna unguiculata) nodulation in three agroecological regions of Mozambique. Appl. Environ. Microbiol. 2018, 84, e01712-17. [Google Scholar] [CrossRef] [PubMed]
- Westhoek, A.; Field, E.; Rehling, F.; Mulley, G.; Webb, I.; Poole, P.S.; Turnbull, L.A. Policing the legume-Rhizobium symbiosis: A critical test of partner choice. Sci. Rep. 2017, 7, 1419. [Google Scholar] [CrossRef] [PubMed]
- Alloway, B.J. Micronutrient Deficiencies in Global Crop proDuction; Springer Science + Business Media, B.V.: Dordrecht, The Netherlands, 2008; ISBN 9781402068607. [Google Scholar]
- O’Hara, G.W. Nutritional constraints on root nodule bacteria affecting symbiotic nitrogen fixation: A review. Aust. J. Exp. Agric. 2001, 41, 417–433. [Google Scholar] [CrossRef]
- Shukla, A.K.; Sinha, N.K.; Tiwari, P.K.; Prakash, C.; Behera, S.K.; Lenka, N.K.; Singh, V.K.; Dwivedi, B.S.; Majumdar, K.; Kumar, A.; et al. Spatial distribution and management zones for sulphur and micronutrients in shiwalik mimalayan region of india. Land Degrad. Dev. 2017, 28, 959–969. [Google Scholar] [CrossRef]
- Hansen, T.H.; Laursen, K.H.; Persson, D.P.; Pedas, P.; Husted, S.; Schjoerring, J.K. Micro-scaled high-throughput digestion of plant tissue samples for multi-elemental analysis. Plant Methods 2009, 5, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Franzen, D.W.; Hopkins, D.H.; Sweeney, M.D.; Ulmer, M.K.; Halvorson, A.D. Evaluation of soil survey scale for zone development of site-specific nitrogen management. Agron. J. 2002, 94, 381–389. [Google Scholar] [CrossRef]
- Khosla, R.; Fleming, K.; Delgado, J.A.; Shaver, T.M.; Westfall, D.G. Use of site-specific management zones to improve nitrogen management for precision agriculture. J. Soil Water Conserv. 2002, 57, 513–518. [Google Scholar] [CrossRef]
- Thilakarathna, M.S.; Papadopoulos, Y.A.; Grimmett, M.; Fillmore, S.A.E.; Crouse, M.; Prithiviraj, B. Red clover varieties with nitrogen fixing advantage during the early stages of seedling development. Can. J. Plant Sci. 2018, 98, 517–526. [Google Scholar] [CrossRef]
- Thilakarathna, M.S.; Papadopoulos, Y.A.; Rodd, A.V.; Grimmett, M.; Fillmore, S.A.E.; Crouse, M.; Prithiviraj, B. Nitrogen fixation and transfer of red clover genotypes under legume-grass forage based production systems. Nutr. Cycl. Agroecosyst. 2016, 106, 233–247. [Google Scholar] [CrossRef]
- Yang, X.; Lu, Y.; Tong, Y.; Yin, X. A 5-year lysimeter monitoring of nitrate leaching from wheat-maize rotation system: Comparison between optimum N fertilization and conventional farmer N fertilization. Agric. Ecosyst. Environ. 2015, 199, 34–42. [Google Scholar] [CrossRef]



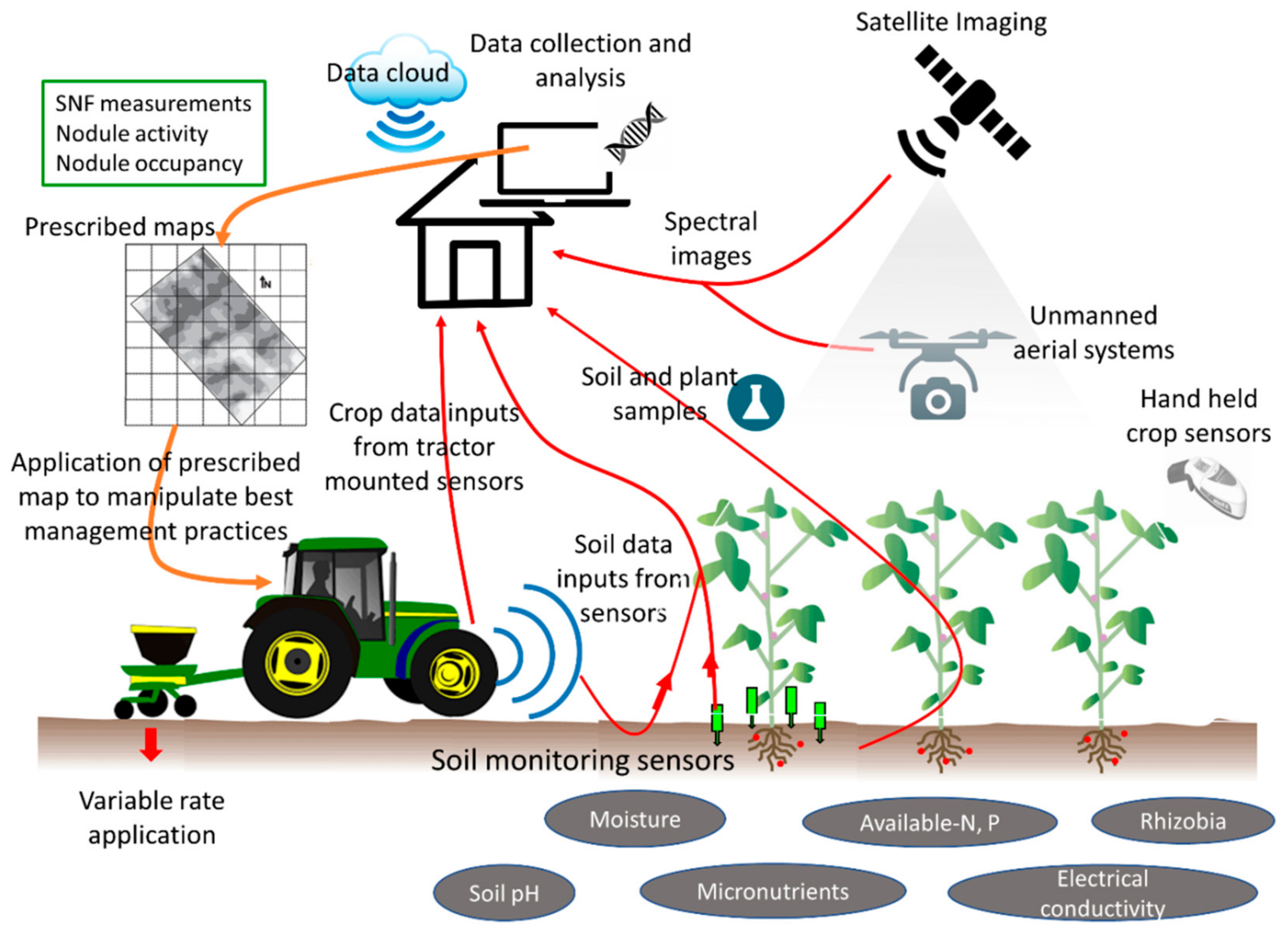{kind=link}
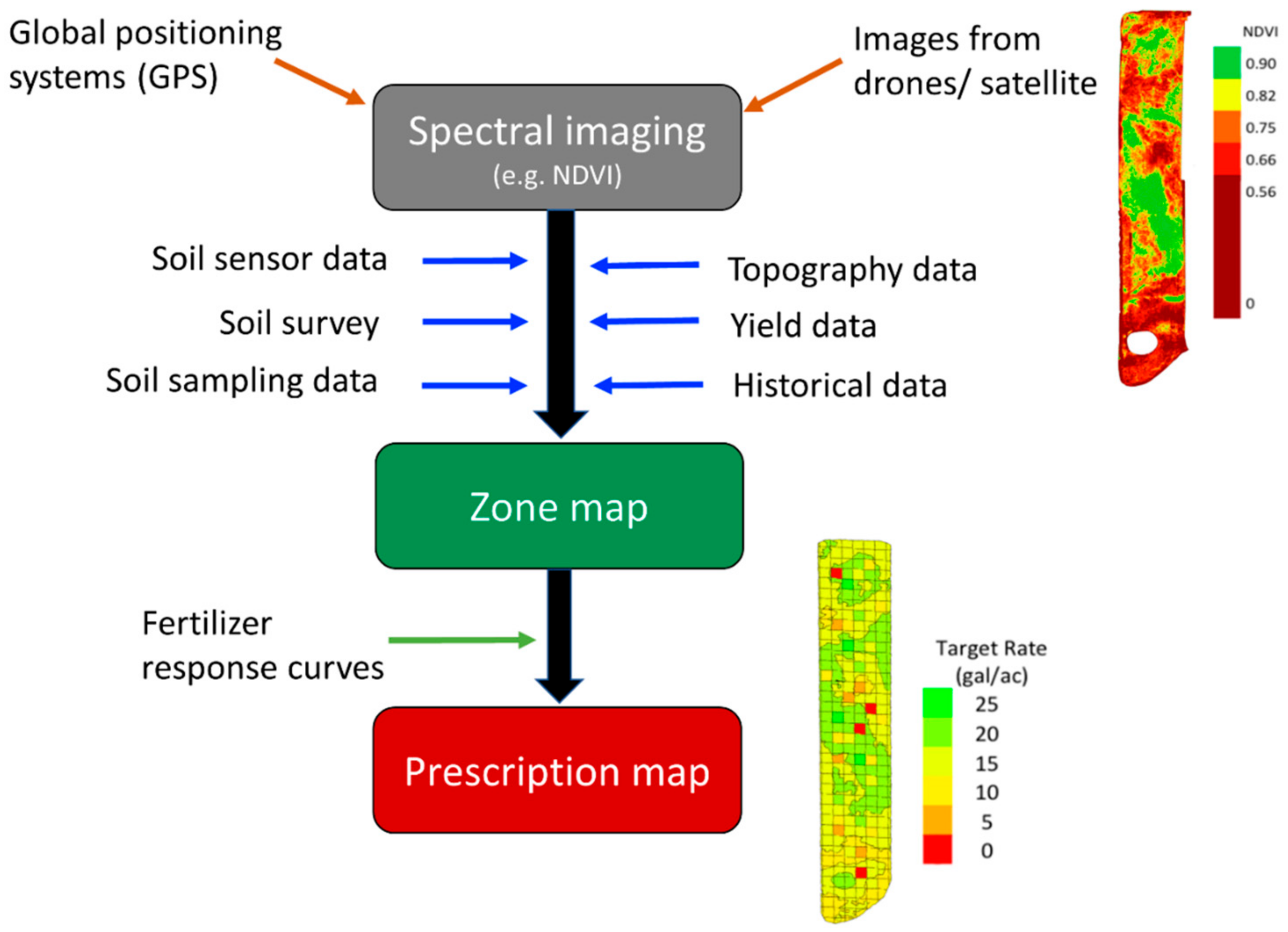{kind=link}
| Method | Principle | Potential to Adapt to Precision Agriculture | |
|---|---|---|---|
| Short–Medium Term | Long Term | ||
| Candidate precision agriculture diagnostics in the short-medium term | |||
| Proximal and remote sensing [32] | Based on the near-infrared spectral region (NDVI or red-edge position (REP)) reflected by plant leaves/canopy. Canopy reflectance has positive correlation with leaf chlorophyll content. | Has tremendous potential to measure SNF at the field level, with the inclusion of control micro plots with different N fertilizer gradients applied within the field. | High probability. |
| Nitrogen difference method [28] | Based on the difference in shoot N accumulated by N2-fixing plants vs. neighboring non-N2-fixing plants (uninoculated or non-nodulating mutant). Assumptions: fixing and non-fixing plants absorb the same amount of N from soil, and surplus N in the fixing plant is from SNF. | Has potential for N-limiting soils if a non-nodulating reference legume genotype is available, and if spectral imaging can distinguish the reference genotype from N-fixing plants. | High probability. |
| 15N natural abundance method [28] | Takes into consideration the natural difference in δ15N abundance between atmospheric N and soil N to measure SNF. SNF calculation based on the difference in δ15N abundance between non-fixing reference plant (resembles soil δ15N) and N-fixing plant. | Use of remote sensing technology to detect plant 15N levels using leaf/canopy reflectance spectra may allow this technology to be used under field conditions. | High probability. |
| 15N isotope dilution method [28] | Plant available soil N is artificially enriched with 15N-enriched fertilizers. N-fixing and non-fixing reference plants are grown in the soil with the same 15N enrichment, and SNF calculated based on the difference in 15N signature of the reference and fixing plants. | Use of remote sensing technology to detect plant 15N levels using leaf/canopy reflectance spectra may allow this technology to be used under field conditions. | High probability. |
| Diagnostics requiring major advances to adapt them for precision agriculture | |||
| Nitrogen balance method [28] | Calculates the difference between all N inputs (e.g., N-fixation, manure, mineralization, inorganic N) and N outputs (e.g., leaching, volatilization, crop removal, erosion) over a period of time. | Low probability: challenging due to the requirement to measure N in multiple components over a long period of time. | Advances are being made in data collection (e.g., satellite images) and big data management using different software programs. |
| Ureide method [33] | Based on the % ureide N in the xylem sap or stem segments. | Low probability: challenging due to requirement to calibrate different crops, under different growth stages as well as under different stress conditions. | A high throughput low-cost method is needed to analyze ureide compounds (e.g., amperometric biosensor specific for ureides). |
| C2H2 reduction assay (ARA) [34] | Based on the promiscuous activity of nitrogenase enzyme, which can reduce acetylene to ethylene which is then measured by gas chromatography; provides an estimate of N2 fixation activity at a point in time. | Low probability: challenging to measure gas compound under field conditions. | Low probability. |
| Nodule number, dry weight, color [29,35,36,37] | - Manual counting and observation of nodules - Manual separation and dry weight measurement - Software-based analysis of scanned root systems for nodule number | Low probability. | Challenging: need for futuristic underground X-ray and magnetic resonance imaging like technologies to examine root nodule density. |
| GlnLux method [31] | GlnLux assay is based on measuring glutamine output of leaf punch extracts using the GlnLux biosensor, which consists of Escherichia coli cells auxotrophic for glutamine. The assay can be used to measure SNF output of legumes grown under controlled conditions with minimal external N supply. | Low probability. | Holds potential if converted into a field-based technology such as translation into an amperometric biosensor specific for Gln. |
| Chlorophyll meter [38] | The SPAD (soil plant analysis development) index gives an indirect measure of leaf chlorophyll. Leaf SPAD index can be used to measure relative SNF under axenic conditions with limited N supply. | Has potential on N-limiting soils if a non-nodulating reference legume genotype is available, and if SPAD can distinguish the reference genotype from N-fixing plants. | Has potential on N limiting soils with a non-nodulating reference genotype. |
| Soil Trait | Current Diagnostic | Potential to Adapt to Precision Agriculture | |
|---|---|---|---|
| Short–Medium Term | Long-Term | ||
| Candidate precision agriculture diagnostics in the short-medium term | |||
| Available soil nitrogen [76,77] |
| High probability:
| High probability. |
| Available soil phosphorus [78] |
| High probability:
| High probability. |
| Soil pH [79,80] |
| High probability:
| High probability. |
| Salinity [81,82] |
| High probability:
| High probability. |
| Diagnostics requiring major advances to adapt them for precision agriculture | |||
| Competitive soil rhizobia [13] |
| Low probability. |
|
| Available soil molybdenum [83] |
| Low probability. |
|
| Available soil boron [84] |
| Low probability. |
|
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Thilakarathna, M.S.; Raizada, M.N. Challenges in Using Precision Agriculture to Optimize Symbiotic Nitrogen Fixation in Legumes: Progress, Limitations, and Future Improvements Needed in Diagnostic Testing. Agronomy 2018, 8, 78. https://doi.org/10.3390/agronomy8050078
Thilakarathna MS, Raizada MN. Challenges in Using Precision Agriculture to Optimize Symbiotic Nitrogen Fixation in Legumes: Progress, Limitations, and Future Improvements Needed in Diagnostic Testing. Agronomy. 2018; 8(5):78. https://doi.org/10.3390/agronomy8050078
Chicago/Turabian StyleThilakarathna, Malinda S., and Manish N. Raizada. 2018. "Challenges in Using Precision Agriculture to Optimize Symbiotic Nitrogen Fixation in Legumes: Progress, Limitations, and Future Improvements Needed in Diagnostic Testing" Agronomy 8, no. 5: 78. https://doi.org/10.3390/agronomy8050078