“Alperujo” Compost Improves the Ascorbate (Vitamin C) Content in Pepper (Capsicum annuum L.) Fruits and Influences Their Oxidative Metabolism

 ,
,  , and
, and 
Abstract
:
1. Introduction
2. Materials and Methods
2.1. Agrochemical Characterization of the AL Compost
2.2. Pepper Growth Conditions
2.3. Experimental Design and Statistical Analysis
- (1)
- In the First experiment, AL compost was added as an organic fertiliser (Exp. No. 1). According to this, three treatments were assayed: NS (no AL compost added and watered with the modified Hewitt nutrient solution mentioned before), C (50% AL compost, v/v, and only watered with distilled water) and C + NS (50% AL compost, v/v, and watered with the modified Hewitt nutrient solution mentioned before). In C and NS treatments, similar total nitrogen concentration was added with differences in its chemical forms: in the former, compost nitrogen was predominantly in the organic form (Table 1), whereas, in the latter treatment (NS), nitrogen was added under nitrate form (easily assimilable). The total amount of NO3– added in NS and C + NS treatments was 2.77 g. Finally, a combination of both treatments (C + NS) was also done to test synergistic effect between AL compost and nitrate. According to physiological evolution, flowering of pepper plants started at 78 days after seeds sowing (first harvest) and the pepper fruits production was monitored at 107 days (second harvest).
- (2)
- Based on the obtained results in Exp. No. 1, a Second experiment (Exp. No. 2) was developed without any nitrogen limitations. Then, four treatments were performed: Control (no AL compost added), C1 (8% of AL compost, v/v), C2 (17% of AL compost, v/v) and C3 (33% of AL compost, v/v). Control treatment was watered using the modified Hewitt mineral solution described before, and C1, C2 and C3 were watered only with NO3– 14.92 mM. For each treatment, the total amount of NO3– added was 3.33 g, respectively. Pepper plant flowering started at 98 days after seeds sowing (first harvest) and pepper fruits were harvested at 127 days (second harvest).
2.4. Chemical and Biochemical Analysis
3. Results
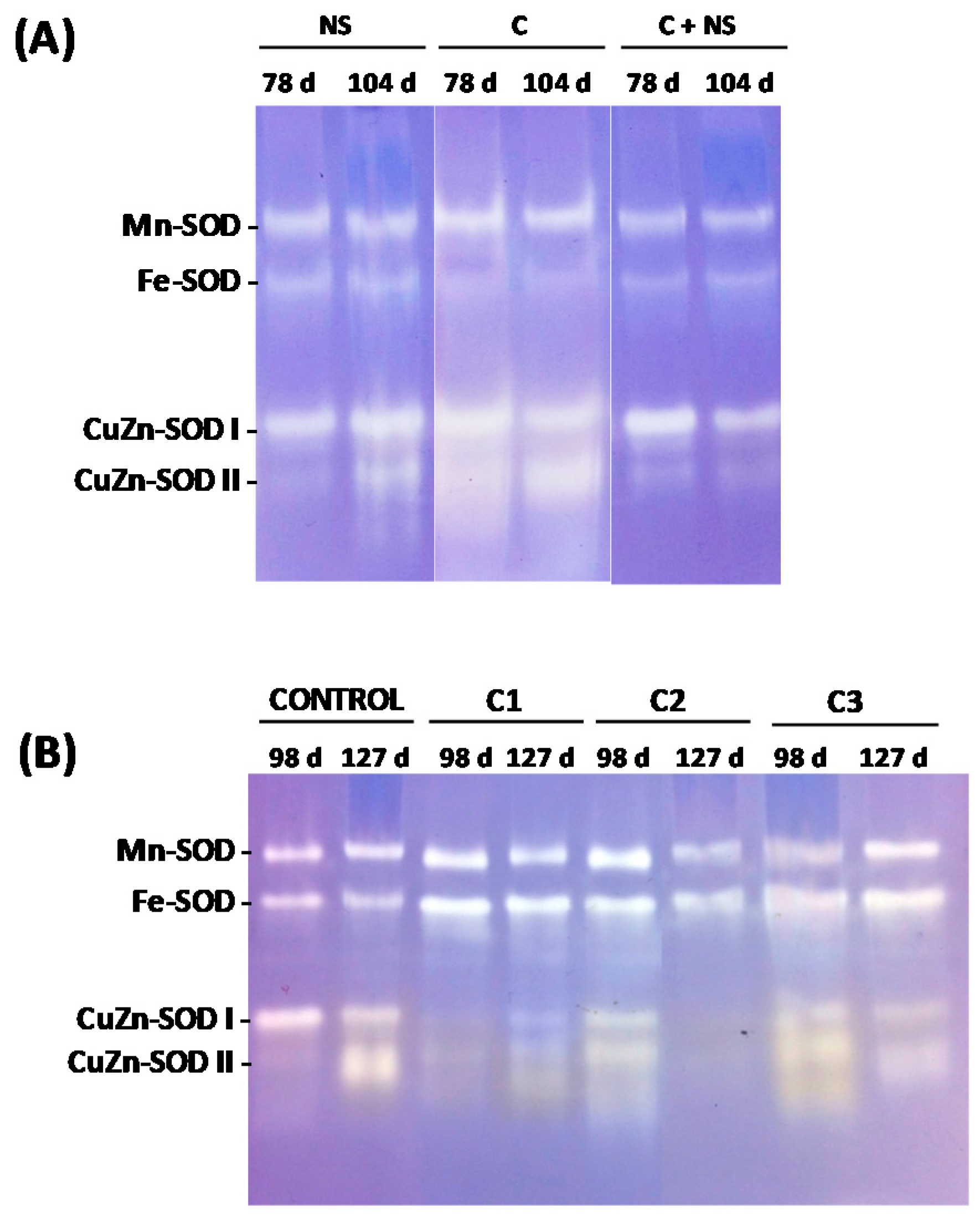
3.1. AL Compost as an Organic Fertiliser (Exp. No. 1)
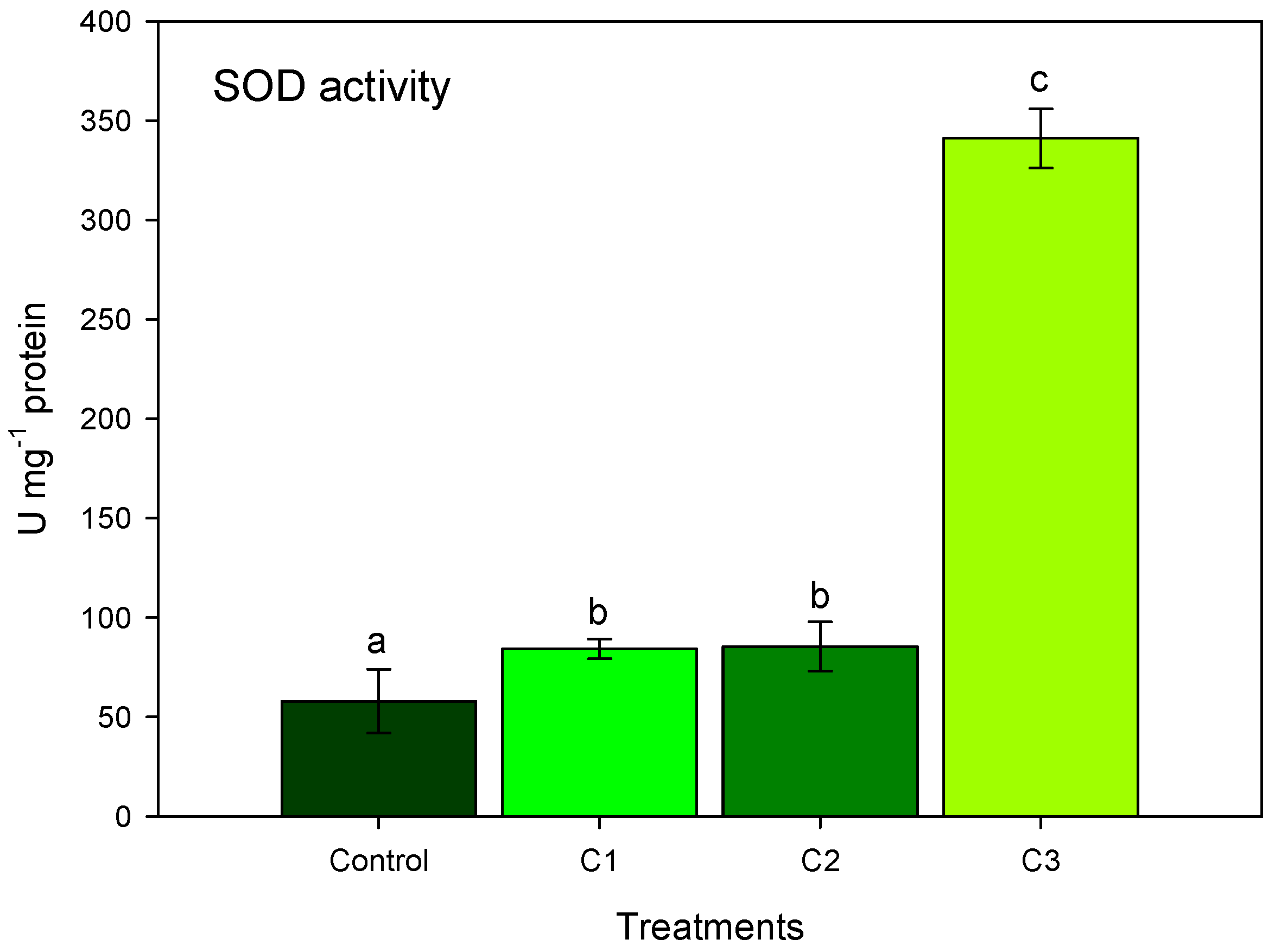
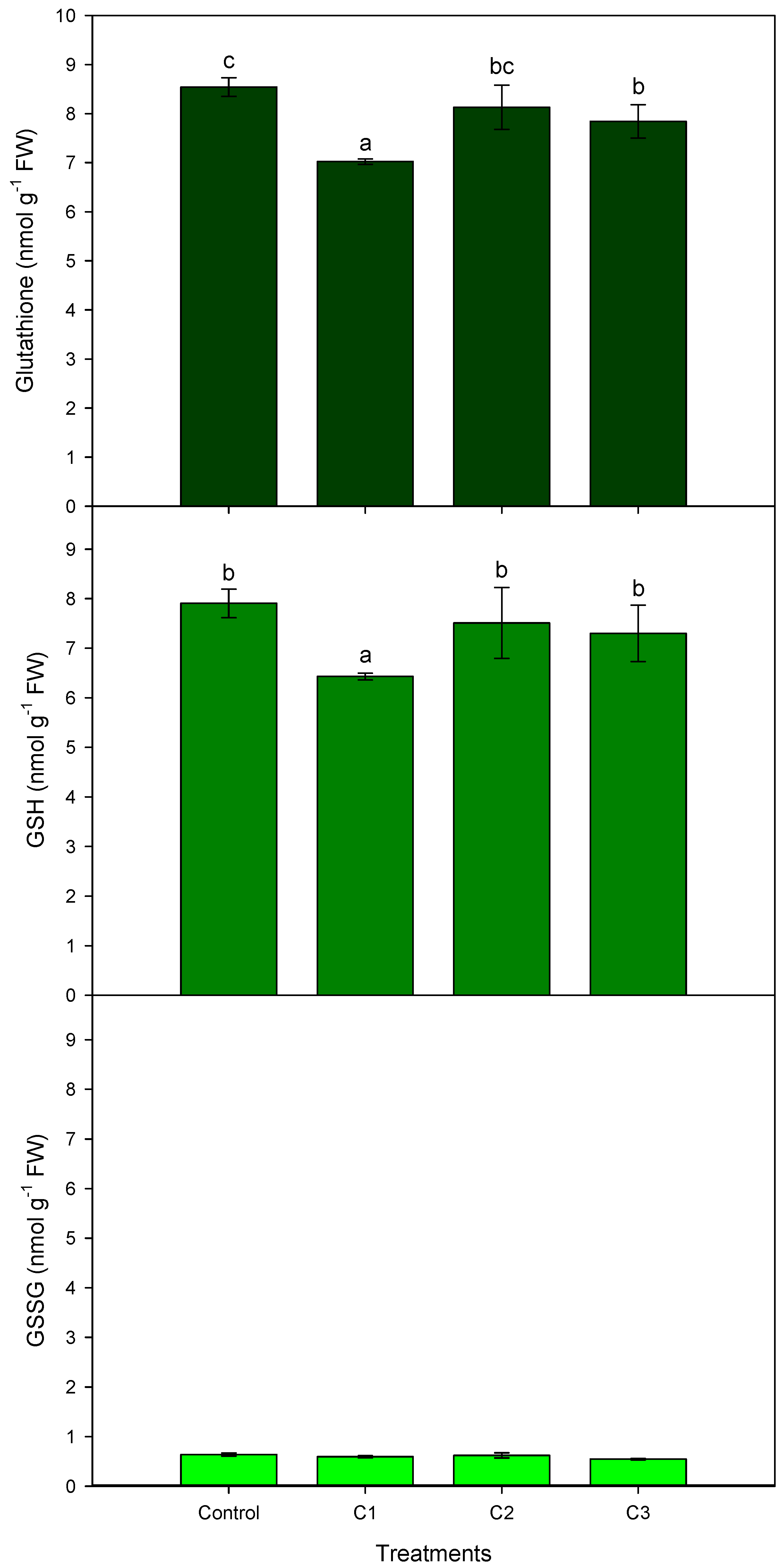
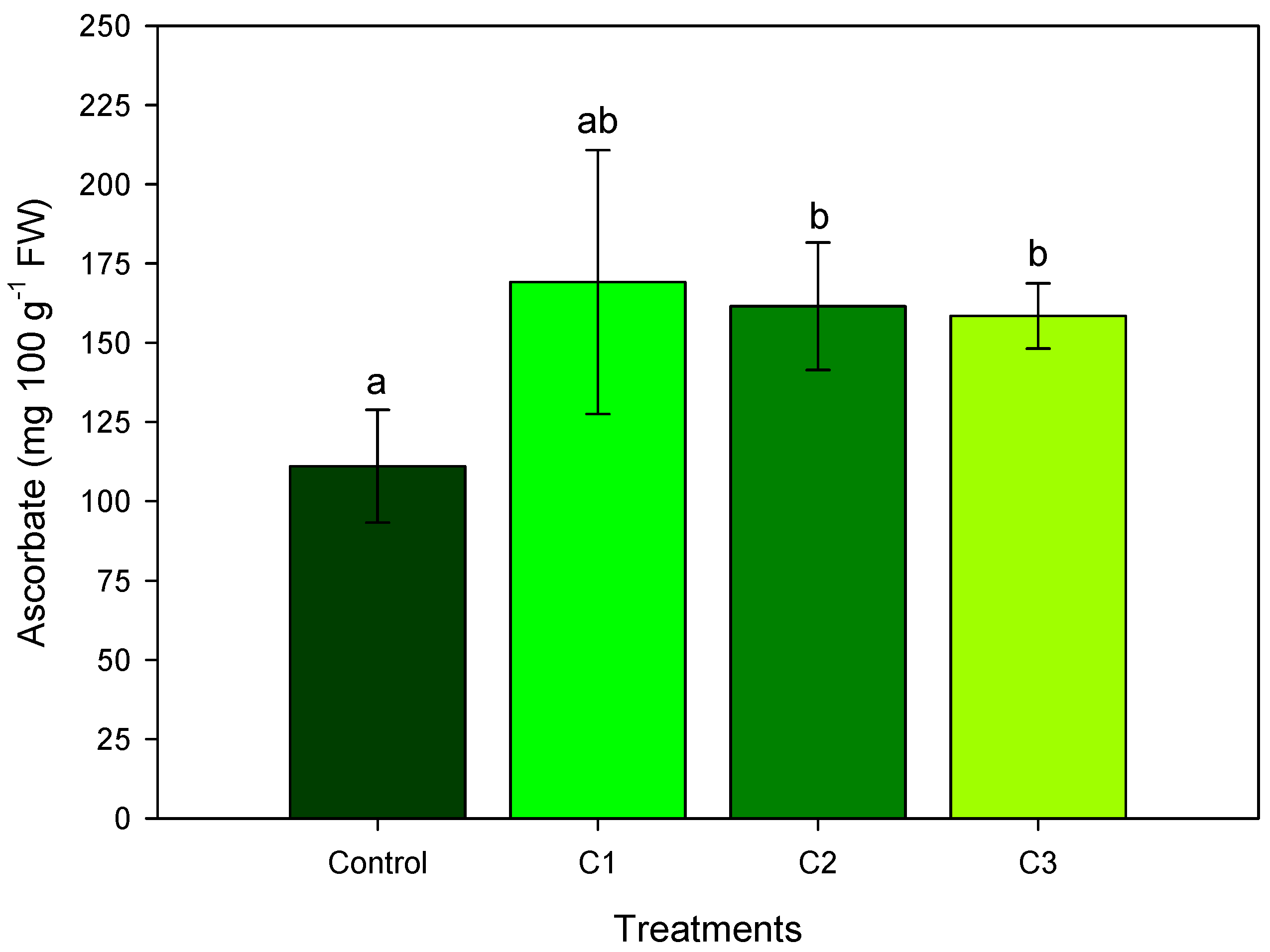
3.2. AL Compost Supplemented with Nitrate (Exp. No. 2)
4. Discussion
5. Conclusions
Author Contributions
Acknowledgments
Conflicts of Interest
References
- White, P.J.; Brown, P.H. Plant nutrition for sustainable development and global health. Ann. Bot. 2010, 105, 1073–1080. [Google Scholar] [CrossRef] [PubMed]
- Food and Agriculture Organization of the United Nations (FAO). World Fertiliser Trends and Outlook to 2019 (Summary Report). 2016. Available online: http://www.fao.org/3/a-i5627e.pdf (accessed on 15 November 2017).
- European Commission. Implementation of Council Directive 91/676/EEC Concerning the Protection of Waters against Pollution Caused by Nitrates from Agricultural Sources; Official Publications of the European Communities, 2002; Available online: http://ec.europa.eu/environment/water/water-nitrates/pdf/91_676_eec_en.pdf (accessed on 15 November 2017).
- Smith, P.; Martino, D.; Cai, Z.; Gwary, D.; Janzen, H.; Kumar, P.; McCarl, B.; Ogle, S.; O’Mara, F.; Rice, C.; et al. Greenhouse gas mitigation in agriculture. Phil. Trans. R. Soc. B 2008, 363, 789–813. [Google Scholar] [CrossRef] [PubMed]
- Tortosa, G.; Correa, D.; Sánchez-Raya, A.J.; Delgado, A.; Sánchez-Monedero, M.A.; Bedmar, E.J. Effects of nitrate contamination and seasonal variation on the denitrification and greenhouse gas production in La Rocina Stream (Doñana National Park, SW Spain). Ecol. Eng. 2011, 37, 539–548. [Google Scholar] [CrossRef] [Green Version]
- Cayuela, M.L.; Aguilera, E.; Sanz-Cobena, A.; Adams, D.C.; Abalos, D.; Barton, L.; Ryals, R.; Silver, W.L.; Alfaro, M.A.; Pappa, V.A.; et al. Direct nitrous oxide emissions in Mediterranean climate cropping systems: Emission factors based on a meta-analysis of available measurement data. Agric. Ecosyst. Environ. 2017, 238, 25–35. [Google Scholar] [CrossRef]
- Food and Agriculture Organization of the United Nations (FAO). Status of the World’s Soil Resources Technical Summary; Food and Agriculture Organization of the United Nations: Rome, Italy, 2015; ISBN 978-92-5-108960-6. [Google Scholar]
- Minasny, B.; Malone, B.P.; McBratney, A.B.; Angers, D.A.; Arrouays, D.; Chambers, A.; Chaplot, V.; Chen, Z.S.; Cheng, K.; Das, B.S.; et al. Soil carbon 4 per mille. Geoderma 2017, 292, 59–86. [Google Scholar] [CrossRef]
- Alburquerque, J.A.; Gonzálvez, J.; García, D.; Cegarra, J. Agrochemical characterisation of ‘‘alperujo”, a solid by-product of the two-phase centrifugation method for olive oil extraction. Bioresour. Technol. 2004, 91, 195–200. [Google Scholar] [CrossRef]
- Alburquerque, J.A.; Gonzálvez, J.; Tortosa, G.; Baddi, G.A.; Cegarra, J. Evaluation of ‘‘alperujo” composting based on organic matter degradation, humification and compost quality. Biodegradation 2009, 20, 257–270. [Google Scholar] [CrossRef] [PubMed]
- Tortosa, G.; Alburquerque, J.A.; Ait Baddi, G.; Cegarra, J. The production of commercial organic amendments and fertilisers by composting of two-phase olive mill waste (‘‘alperujo”). J. Clean. Prod. 2012, 26, 48–55. [Google Scholar] [CrossRef]
- Tortosa, G.; Alburquerque, J.A.; Bedmar, E.J.; Ait Baddi, G.; Cegarra, J. Strategies to produce commercial liquid organic fertilisers from “alperujo” composts. J. Clean. Prod. 2014, 82, 37–44. [Google Scholar] [CrossRef]
- Stevenson, F.J. Humus Chemistry: Genesis, Composition, Reactions, 2nd ed.; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 1994; ISBN 978-0-471-59474-1. [Google Scholar]
- Govindarajan, V.S.; Salzer, U.J. Capsicum production, technology, chemistry, and quality part 1: History, botany, cultivation, and primary processing. Crit. Rev. Food Sci. Nutr. 1985, 22, 109–176. [Google Scholar] [CrossRef] [PubMed]
- FAO-STAT. Food and Agriculture Organization of the United Nations (FAO). 2017. Available online: http://www.fao.org/faostat (accessed on 15 November 2017).
- Senesi, N. Composted Materials as organic fertilisers. Sci. Total Environ. 1985, 81/82, 521–542. [Google Scholar] [CrossRef]
- Quilty, J.R.; Cattle, S.R. Use and understanding of organic amendments in Australian agriculture: A review. Soil Res. 2011, 49, 1–26. [Google Scholar] [CrossRef]
- Martínez-Blanco, J.; Lazcano, C.; Christensen, T.H.; Muñoz, P.; Rieradevall, J.; Møller, J.; Antón, A.; Boldrin, A. Compost benefits for agriculture evaluated by life cycle assessment. A review. Agron. Sustain. Dev. 2013, 33, 721–732. [Google Scholar] [CrossRef]
- Del Río, L.A. ROS and RNS in plant physiology: An overview. J. Exp. Bot. 2015, 66, 2827–2837. [Google Scholar] [CrossRef] [PubMed]
- Mittler, R. ROS Are Good. Trends Plant Sci. 2017, 22, 11–19. [Google Scholar] [CrossRef] [PubMed]
- Halliwell, B. The antioxidant paradox: Less paradoxical now? Br. J. Clin. Pharmacol. 2012, 75, 637–644. [Google Scholar] [CrossRef] [PubMed]
- Palma, J.M.; Sevilla, F.; Jiménez, A.; del Río, L.A.; Corpas, F.J.; Álvarez de Morales, P.; Camejo, D.M. Physiology of pepper fruit and the metabolism of antioxidants: Chloroplasts, mitochondria and peroxisomes. Ann. Bot. 2015, 116, 627–636. [Google Scholar] [CrossRef] [PubMed]
- Palma, J.M.; Corpas, F.J.; del Río, L.A. Plant vitamin antioxidants and their influence on the human diet. In Fruit and vegetable consumption and health; Papareschi, A., Eppolito, H., Eds.; Nova Science Publishers Inc.: New York, NY, USA, 2009; pp. 127–138. [Google Scholar]
- Howard, L.R.; Talcott, S.T.; Brenes, C.H.; Villalon, B. Changes in Phytochemical and Antioxidant Activity of Selected Pepper Cultivars (Capsicum Species) As Influenced by Maturity. J. Agric. Food Chem. 2000, 48, 1713–1720. [Google Scholar] [CrossRef] [PubMed]
- Chowdhury, A.K.M.M.B.; Akratos, C.S.; Vayenas, D.V.; Pavlou, S. Olive mill waste composting: A review. Int. Biodeterior. Biodegrad. 2013, 85, 108–119. [Google Scholar] [CrossRef]
- Del Río, L.A.; Sandalio, L.M.; Yáñez, J.; Gómez, M. Induction of a manganese-containing superoxide dismutase in leaves of Pisum sativum L. by high nutrient levels of zinc and manganese. J. Inorg. Biochem. 1985, 24, 25–34. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1986, 72, 248–254. [Google Scholar] [CrossRef]
- Aebi, H. Catalase in vitro. Methods Enzymol. 1984, 105, 121–126. [Google Scholar] [PubMed]
- McCord, J.M.; Fridovich, I. Superoxide dismutase: An enzymic function for erythrocuprein. J. Biol. Chem. 1969, 244, 6049–6055. [Google Scholar] [PubMed]
- Beauchamp, C.O.; Fridovich, I. Superoxide dismutase: Improved assays and an assay applicable to acrylamide gels. Anal. Biochem. 1971, 44, 276–287. [Google Scholar] [CrossRef]
- Buege, J.A.; Aust, S.D. Microsomal lipid peroxidation. Methods Enzymol. 1978, 52, 302–310. [Google Scholar] [PubMed]
- Rodríguez-Ruiz, M.; Mateos, R.M.; Codesido, V.; Corpas, F.J.; Palma, J.M. Characterization of the galactono-1,4-lactone dehydrogenase from pepper fruits and its modulation in the ascorbate biosynthesis. Role of nitric oxide. Redox Biol. 2017, 12, 171–181. [Google Scholar] [CrossRef] [PubMed]
- Airaki, M.; Leterrier, M.; Mateos, R.M.; Valderrama, R.; Chaki, M.; Barroso, J.B.; Del Río, L.A.; Palma, J.M.; Corpas, F.J. Metabolism of reactive oxygen species and reactive nitrogen species in pepper (Capsicum annuum L.) plants under low temperature stress. Plant Cell Environ. 2011, 35, 281–295. [Google Scholar] [CrossRef] [PubMed]
- Pascual, I.; Azcona, I.; Guirreolea, J.A.; Morales, F.; Corpas, F.J.; Palma, J.M.; Rellán-Álvarez, R.; Sánchez-Díaz, M.J. Growth, Yield, and Fruit Quality of Pepper Plants Amended with Two Sanitized Sewage Sludges. J. Agric. Food Chem. 2010, 58, 6951–6959. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.Y.; Lin, H.S. Compost as a Soil Supplement Increases the Level of Antioxidant Compounds and Oxygen Radical Absorbance Capacity in Strawberries. J. Agric. Food Chem. 2003, 51, 6844–6850. [Google Scholar] [CrossRef] [PubMed]
- Arancon, N.Q.; Edwards, C.A.; Atiyeh, R.; Metzger, J.D. Effects of vermicomposts produced from food waste on the growth and yields of greenhouse peppers. Bioresour. Technol. 2004, 93, 139–144. [Google Scholar] [CrossRef] [PubMed]
- Galloway, J.N.; Aber, J.D.; Erisman, J.W.; Seitzinger, S.P.; Howarth, R.W.; Cowling, E.B.; Cosby, B.J. The Nitrogen Cascade. BioScience 2003, 53, 341–356. [Google Scholar] [CrossRef]
- Good, A.G.; Beatty, P.H. Fertilizing Nature: A Tragedy of Excess in the Commons. PLoS Biol. 2011, 9, e1001124. [Google Scholar] [CrossRef] [PubMed]
- Martins, O.; Dewes, T. Loss of Nitrogenous Compounds during Composting of Animal Wastes. Bioresour. Technol. 1992, 42, 103–111. [Google Scholar] [CrossRef]
- Tiquia, S.M.; Tam, N.F.Y. Fate of nitrogen during composting of chicken litter. Environ. Pollut. 2000, 110, 535–541. [Google Scholar] [CrossRef]
- Herridge, D.F.; Peoples, M.B.; Boddey, R.M. Global inputs of biological nitrogen fixation in agricultural systems. Plant Soil. 2008, 311, 1–18. [Google Scholar] [CrossRef]
- Bhat, T.A.; Ahmad, L.; Ganai, M.A.; Haq, S.U.; Khan, O.A. Nitrogen Fixing Biofertilisers; Mechanism and Growth Promotion: A Review. J. Pure Appl. Microbiol. 2015, 9, 1675–1690. [Google Scholar]
- Adamczyk, B.; Smolander, A.; Kitunen, V.; Godlewski, M. Proteins as nitrogen source for plants. A short story about exudation of proteases by plant roots. Plant Signal. Behav. 2010, 5, 817–819. [Google Scholar] [CrossRef]
- Shin, R.; Berg, R.H.; Schachtman, D.P. Reactive Oxygen Species and Root Hairs in Arabidopsis Root Response to Nitrogen, Phosphorus and Potassium Deficiency. Plant Cell Physiol. 2005, 46, 1350–1357. [Google Scholar] [CrossRef] [PubMed]
- Tartoura, K.A.H.; Youssef, S.A. Stimulation of ROS-scavenging systems in squash (Cucurbita pepo L.) plants by compost supplementation under normal and low temperature conditions. Sci. Hortic. 2011, 130, 862–868. [Google Scholar] [CrossRef]
- Youssef, S.A.; Tartoura, K.A.H. Compost enhances plant resistance against the bacterial wilt pathogen Ralstonia solanacearum via up-regulation of ascorbate-glutathione redox cycle. Eur. J. Plant Pathol. 2013, 137, 821–834. [Google Scholar] [CrossRef]
- Tartoura, K.A.H.; Youssef, S.A.; Tartoura, E.A.A. Compost alleviates the negative effects of salinity via up-regulation of antioxidants in Solanum lycopersicum L. plants. Plant Growth Regul. 2014, 74, 299–310. [Google Scholar] [CrossRef]
- Rimmer, D.L. Free radicals, antioxidants, and soil organic matter recalcitrance. Eur. J. Soil Sci. 2006, 57, 91–94. [Google Scholar] [CrossRef]
- Masmoudi, S.; Jarboui, R.; Feki, H.E.; Gea, T.; Medhioub, K.; Ammar, E. Characterization of olive mill wastes composts and their humic acids: Stability assessment within different particle size fractions. Environ. Technol. 2013, 34, 787–797. [Google Scholar] [CrossRef] [PubMed]
- Calderín-García, A.; Olaetxea, M.; Santos, L.A.; Mora, V.; Baigorri, R.; Fuentes, M.; Zamarreño, A.M.; Louro-Berbara, R.L.; Garcia-Mina, J.M. Involvement of Hormone- and ROS-Signaling Pathways in the Beneficial Action of Humic Substances on Plants Growing under Normal and Stressing Conditions. BioMed Res. Int. 2016, 3747501. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameters 1 | Mean | Standard Deviation | Coefficient of Variation (%) |
|---|---|---|---|
| Moisture | 38.5 | 2.2 | 6 |
| pH 2 | 7.67 | 0.23 | 3 |
| EC 2 (dS m−1) | 1.11 | 0.08 | 7 |
| OM (%) | 54.5 | 1.8 | 3 |
| Lignin (%) | 31.9 | 4.5 | 14 |
| Cellulose (%) | 15.8 | 2.2 | 14 |
| Hemicellulose (%) | 10.7 | 2.1 | 20 |
| TOC (%) | 25.27 | 0.03 | 0 |
| TN (%) | 1.50 | 0.30 | 20 |
| TOC/TN | 17.16 | 3.41 | 20 |
| Fat content (%) | 0.35 | 0.07 | 20 |
| WSC (%) | 1.70 | 0.14 | 9 |
| WSCH (%) | 0.30 | 0.14 | 47 |
| WSPH (%) | 0.15 | 0.07 | 47 |
| HR | 28.85 | 2.33 | 8 |
| HD | 79.65 | 1.06 | 1 |
| PHA (%) | 78.30 | 3.39 | 4 |
| P (%) | 0.32 | 0.04 | 13 |
| K (%) | 1.14 | 0.17 | 15 |
| Ca (%) | 2.99 | 0.16 | 5 |
| Mg (%) | 0.63 | 0.12 | 19 |
| Na (%) | 0.10 | 0.03 | 28 |
| S (%) | 0.17 | 0.03 | 17 |
| Fe (%) | 0.86 | 0.01 | 2 |
| Cu (mg kg−1) | 30 | 2 | 7 |
| Mn (mg kg−1) | 221 | 30 | 13 |
| Zn (mg kg−1) | 55 | 4 | 6 |
| Pb (mg kg−1) | 16 | 1 | 9 |
| Cr (mg kg−1) | 35 | 6 | 18 |
| Ni (mg kg−1) | 17 | 2 | 13 |
| Cd (mg kg−1) | <0.025 | - | - |
| Hg (mg kg−1) | <0.025 | - | - |
| GI | 93 | 1 | 2 |
| Exp. No. 1 | Treatment | SDW (g) | RDW (g) | Number of Fruits per Plant | Yield of Pepper Fruits per Plant (g) |
|---|---|---|---|---|---|
| Harvest | |||||
| 78 days | NS | 3.9 b | 1.9 b | 0.0 | 0.0 |
| C | 1.3 a | 3.7 c | 0.0 | 0.0 | |
| C + NS | 5.5 c | 1.2 a | 0.0 | 0.0 | |
| 107 days | NS | 15.9 b | 4.9 a | 3.1 a | 43.1 a |
| C | 2.0 a | 5.1 a | 0.0 | 0.0 | |
| C + NS | 20.3 c | 7.2 b | 3.3 b | 45.6 b | |
| Exp. No. 2 | |||||
| Harvest | |||||
| 98 days | Control | 1.6 a | 1.5 a | 0.0 | 0.0 |
| C1 | 3.5 b | 3.2 b | 0.0 | 0.0 | |
| C2 | 5.1 c | 5.7 c | 0.0 | 0.0 | |
| C3 | 6.8 d | 5.9 c | 0.0 | 0.0 | |
| 127 days | Control | 7.7 a | 7.9 a | 1.3 a | 19.0 a |
| C1 | 8.0 a | 7.5 a | 2.0 c | 37.4 b | |
| C2 | 10.6 b | 12.4 c | 1.6 b | 47.6 b | |
| C3 | 9.2 b | 10.6 b | 2.0 c | 72.5 c | |
| Exp. No. 1 | ||||||
|---|---|---|---|---|---|---|
| Harvest | 78 days | 107 days | ||||
| Plant Organ | Shoots | |||||
| Nutrients 1 | NS | C | C + NS | NS | C | C + NS |
| TOC (%) | 38.03 a | 40.43 a | 38.47 a | 41.00 a | 40.47 a | 40.67 a |
| TN (%) | 5.17 b | 2.32 a | 5.54 c | 2.81 b | 1.29 a | 3.18 c |
| P (g kg−1) | 3.0 c | 1.7 a | 2.2 b | 2.2 b | 1.6 a | 2.0 b |
| K (g kg−1) | 35.9 b | 27.7 a | 38.2 c | 22.8 a | 24.1 a | 23.7 a |
| Ca (g kg−1) | 17.2 b | 6.5 a | 16.1 b | 11.2 b | 6.5 a | 12.5 b |
| Mg (g kg−1) | 10.7 b | 5.9 a | 11.1 b | 5.2 a | 5.1 a | 6.6 b |
| Na (g kg−1) | 0.1 a | 0.1 a | 0.1 a | 0.1 a | 0.1 a | 0.1 a |
| S (g kg−1) | 4.7 b | 2.3 a | 5.1 b | 3.5 b | 1.8 a | 3.8 b |
| Fe (mg kg−1) | 238 c | 92 a | 213 b | 100 b | 40 a | 98 b |
| Cu (mg kg−1) | 24 b | 10 a | 31 b | 11 a | 5 a | 16 a |
| Mn (mg kg−1) | 192 c | 128 a | 159 b | 86 a | 128 b | 95 a |
| Zn (mg kg−1) | 13 a | 14 a | 16 a | 13 a | 15 a | 11 a |
| Exp. No. 1 Harvest | Treatments | Catalase Activity (µmol H2O2 min−1 mg−1 Protein) | Ascorbate (µmol g−1 FW) | Glutathione (nmol g−1 FW) | GSH (nmol g−1 FW) | GSSG (nmol g−1 FW) | GSH/GSSG |
|---|---|---|---|---|---|---|---|
| 78 days | NS | 11.40 a | 1.240 a | 142 b | 129 b | 13 a | 10 |
| C | 35.46 b | 1.492 a | 110 a | 97 a | 12 a | 8 | |
| C + NS | 11.28 a | 1.112 a | 164 c | 151 c | 13 a | 12 | |
| 107 days | NS | 19.34 a | 0.905 a | 91 b | 11 a | 80 b | 0.1 |
| C | 25.13 b | 1.278 a | 71 a | 22 a | 49 a | 0.4 | |
| C +NS | 13.18 a | 1.021 a | 52 a | 17 a | 35 a | 0.5 |
| Exp. No. 2 | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Harvest | 98 days | 127 days | ||||||||||
| Plant Organ | Shoots | Shoots | Fruits | |||||||||
| Nutrients 1 | Control | C1 | C2 | C3 | Control | C1 | C2 | C3 | Control | C1 | C2 | C3 |
| TOC (%) | 36.95 a | 37.45 b | 37.50 b | 38.20 b | 38.90 a | 37.90 a | 39.00 b | 37.90 a | 50.30 c | 44.90 a | 47.20 b | 45.80 a |
| TN (%) | 4.79 c | 4.72 c | 3.76 b | 2.78 a | 2.53 b | 2.71 c | 2.31 a | 2.10 a | 1.31 a | 1.72 c | 1.93 d | 1.46 b |
| P (g kg−1) | 4.9 c | 2.4 a | 3.0 b | 2.6 a | 1.4 c | 0.8 a | 1.2 b | 1.2 b | 2.3 b | 1.4 a | 2.2 b | 2.5 c |
| K (g kg−1) | 39.9 d | 36.1 b | 37.7 c | 31.7 a | 29.0 a | 28.6 a | 30.2 a | 29.6 a | 19.4 b | 17.3 a | 18.5 a | 17.7 a |
| Ca (g kg−1) | 12.7 b | 11.6 a | 13.5 b | 11.1 a | 9.9 a | 14.6 b | 14.0 b | 16.3 c | 0.6 a | 1.0 b | 1.0 b | 0.9 b |
| Mg (g kg−1) | 12.4 c | 9.9 b | 9.7 b | 5.5 a | 8.0 b | 7.8 b | 6.8 b | 5.6 a | 1.3 a | 1.1 a | 1.3 a | 1.2 a |
| Na (g kg−1) | 0.3 b | 0.1 a | 0.1 a | 0.0 a | 0.0 a | 0.0 a | 0.0 a | 0.0 a | 0.0 a | 0.0 a | 0.0 a | 0.0 a |
| S (g kg−1) | 5.3 c | 3.6 b | 3.4 b | 2.2 a | 4.6 b | 2.5 a | 2.2 a | 2.0 a | 1.8 c | 1.5 b | 1.6 b | 1.3 a |
| Fe (mg kg−1) | 386 d | 120 b | 143 c | 81 a | 157 d | 66 b | 82 c | 56 a | 100 b | 79 a | 94 b | 73 a |
| Cu (mg kg−1) | 20 c | 9 b | 8 b | 6 a | 12 b | 5 a | 5 a | 5 a | 12 b | 6 a | 6 a | 6 a |
| Mn (mg kg−1) | 124 d | 57 b | 68 c | 42 a | 87 c | 47 a | 59 b | 59 b | 14 b | 11 a | 14 b | 13 b |
| Zn (mg kg−1) | 44 c | 28 a | 32 b | 29 a | 34 b | 18 a | 17 a | 31 b | 24 a | 30 b | 44 c | 41 c |
| Exp. No. 2 Harvest | Treatments | Catalase Activity (µmol H2O2 min−1 mg−1 Protein) | Lipid Peroxidation (µM MDA g−1 FW) |
|---|---|---|---|
| 98 days | Control | 39.65 a | 0.33 a |
| C1 | 31.20 a | 0.35 a | |
| C2 | 43.37 b | 0.41 a | |
| C3 | 73.96 c | 0.47 a | |
| 127 days | Control | 68.34 b | 0.34 a |
| C1 | 47.35 a | 0.32 a | |
| C2 | 69.57 b | 0.38 a | |
| C3 | 71.26 c | 0.41 a |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tortosa, G.; González-Gordo, S.; Ruiz, C.; Bedmar, E.J.; Palma, J.M. “Alperujo” Compost Improves the Ascorbate (Vitamin C) Content in Pepper (Capsicum annuum L.) Fruits and Influences Their Oxidative Metabolism. Agronomy 2018, 8, 82. https://doi.org/10.3390/agronomy8060082
Tortosa G, González-Gordo S, Ruiz C, Bedmar EJ, Palma JM. “Alperujo” Compost Improves the Ascorbate (Vitamin C) Content in Pepper (Capsicum annuum L.) Fruits and Influences Their Oxidative Metabolism. Agronomy. 2018; 8(6):82. https://doi.org/10.3390/agronomy8060082
Chicago/Turabian StyleTortosa, Germán, Salvador González-Gordo, Carmelo Ruiz, Eulogio J. Bedmar, and José M. Palma. 2018. "“Alperujo” Compost Improves the Ascorbate (Vitamin C) Content in Pepper (Capsicum annuum L.) Fruits and Influences Their Oxidative Metabolism" Agronomy 8, no. 6: 82. https://doi.org/10.3390/agronomy8060082