Challenges and Prospects for Building Resilient Disease Management Strategies and Tactics for the New York Table Beet Industry


Abstract
:1. Introduction
2. Diseases Affecting Table Beet Production
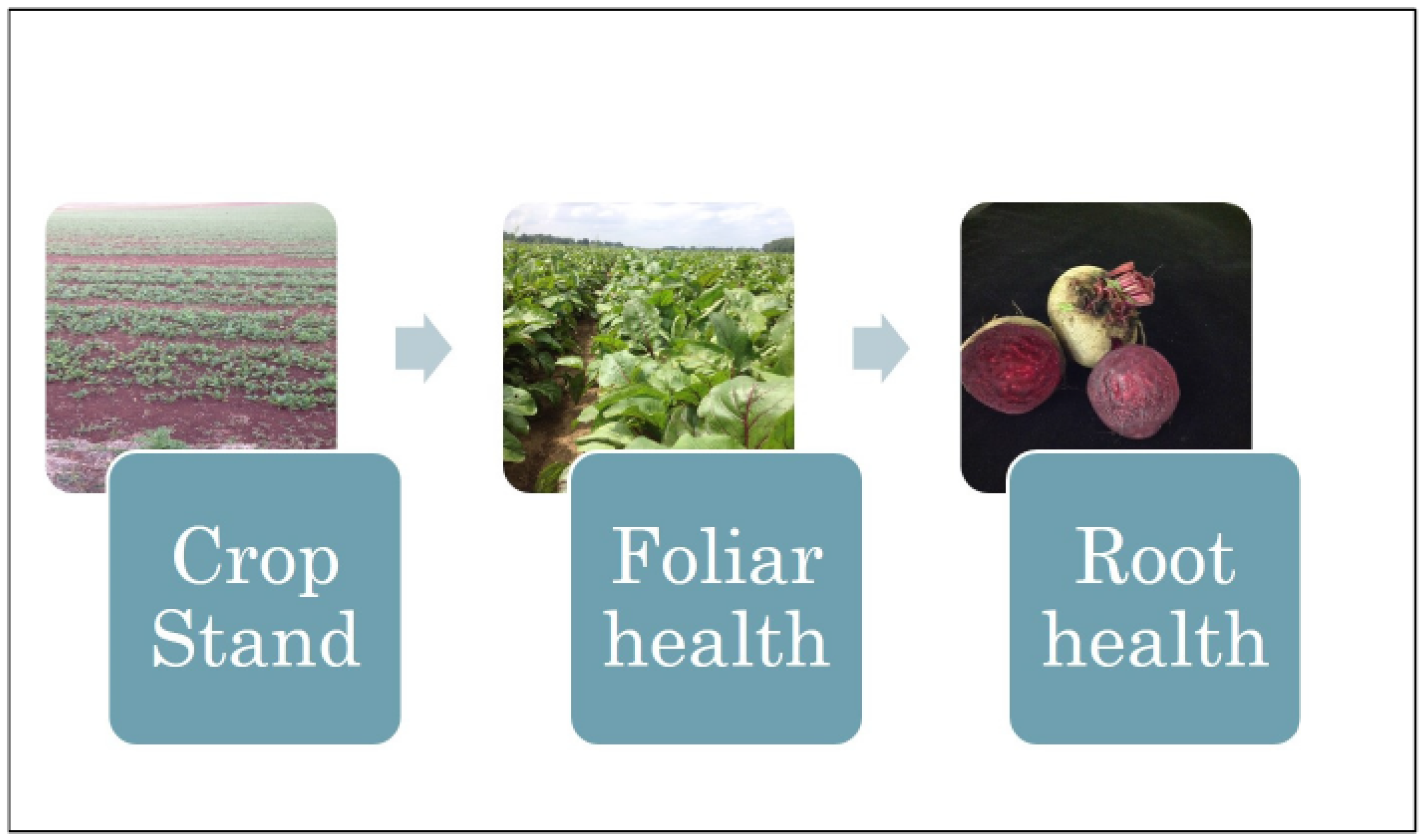
2.1. Diseases Affecting Crop Stand
2.2. Diseases Affecting Foliar Health—Cercospora Leaf Spot
Other Foliar Diseases
2.3. Root Diseases
3. Conclusions and Opportunities
Funding
Acknowledgments
Conflicts of Interest
References
- USDA-NASS. United States Department of Agriculture National Agriculture Statistics Service. 2016. Available online: https://quickstats.nass.usda.gov/ (accessed on 31 March 2018).
- Stivers, L. Crop Profile: Beets in New York. Cornell Cooperative Extension Bulletin. Available online: http://pmep.cce.cornell.edu/fqpa/crop-profiles/beet.html (accessed on 25 March 2018).
- Thakore, Y. The biopesticide market for global agricultural use. Ind. Biotechnol. 2006, 2, 194–208. [Google Scholar] [CrossRef]
- Governor, A.M. Cuomo. Love Beets USA to be Located at Eastman Business Park. Available online: https://www.governor.ny.gov/news/governor-cuomo-announces-140-new-jobs-rochester-lidestri-foods-and-gs-fresh-create-partnership (accessed on 3 July 2018).
- Clifford, T.; Howatson, G.; West, D.J.; Stevenson, E.J. The potential benefits of red beetroot supplementation in health and disease. Nutrients 2015, 7, 2801–2822. [Google Scholar] [CrossRef] [PubMed]
- Gilchrist, M.; Winyard, P.G.; Fulford, J.; Anning, C.; Shore, A.C.; Benjamin, N. Dietary nitrate supplementation improves reaction time in type 2 diabetes: Development and application of a novel nitrate-depleted beetroot juice placebo. Nitrate Oxide 2014, 40, 67–74. [Google Scholar] [CrossRef] [PubMed]
- Hobbs, D.A.; George, T.W.; Lovegrove, J.A. The effects of dietary nitrate on blood pressure and endothelial function: A review of human intervention studies. Nutr. Res. Rev. 2013, 26, 210–222. [Google Scholar] [CrossRef] [PubMed]
- Georgiev, V.G.; Weber, J.; Kneschke, E.M.; Denev, P.N.; Bley, T.; Pavlov, A.I. Antioxidant activity and phenolic content of betalain extracts from intact plants and hairy root cultures of the red beetroot Beta vulgaris cv. Detroit Dark Red. Plant Foods Hum. Nutr. 2010, 65, 105–111. [Google Scholar] [CrossRef] [PubMed]
- Vanhatalo, A.; Bailey, S.J.; Blackwell, J.R.; di Menna, F.J.; Pavey, T.G.; Wilderson, D.P.; Benjamin, N.; Winyard, P.B.; Jones, A.M. Acute and chronic effects of dietary nitrate supplementation on blood pressure and the physiological responses to moderate-intensity and incremental exercise. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2010, 299, R1121–R1131. [Google Scholar] [CrossRef] [PubMed]
- Szalaty, M. Physiological roles and bioavailability of betacyanins. Postep. Fitoter. 2008, 1, 20–25. [Google Scholar]
- Lewellen, R.T.; Panella, L.W.; Harveson, R. Introduction—Botany of the beet plant. In Compendium of Beet Diseases and Insects; APS Press: St. Paul, MN, USA, 2009; ISBN 978-0-89054-365-8. [Google Scholar]
- Warne, L.G.G. Effects of close spacing on the growth of garden beet. Nature 1953, 172, 506. [Google Scholar] [CrossRef]
- Benjamin, L.R. The relative importance of cluster size, sowing depth, time of seedling emergence, and between-plant spacing on variation in plant size in red beet (Beta vulgaris L.) crops. J. Agric. Sci. 1987, 108, 221–230. [Google Scholar] [CrossRef]
- Benjamin, L.R.; Bell, N. The influence of seed type and plant density on variation in plant size of red beet (Beta vulgaris L.) crops. J. Agric. Sci. 1985, 105, 563–571. [Google Scholar] [CrossRef]
- Benjamin, L.R.; Sutherland, R.A.; Senior, D. The influence of sowing rate and row spacing on the plant density and yield of red beet. J. Agric. Sci. 1985, 104, 615–624. [Google Scholar] [CrossRef]
- Kikkert, J.R.; Reiners, S.; Gugino, B.K. Row width, population density, and harvest dates on marketable yield of table beet. HortTechnology 2010, 20, 560–567. [Google Scholar]
- Tyler, F.T.; Adas, L.; Benjamin, L.R. Spacing red beets for high returns. Grower 1982, 97, 19–23. [Google Scholar]
- Becker, R. The beet goes on, this stalwart vegetable keeps getting better. Horticulture 1987, 10, 51–55. [Google Scholar]
- Du Toit, L. Crop Profile for Table Beet Seed in Washington; Washington State University Extension Bulletin Number MISC0356E; Washington State University Press: Pullman, WA, USA, 2007. [Google Scholar]
- Hipp, B.W. Influence of nitrogen and length of growing season on yield and size distribution of table beets. J. Am. Soc. Hortic. Sci. 1977, 102, 598–601. [Google Scholar]
- Mampa, S.S.; Maboko, M.M.; Soundy, P.; Sivakumar, D. Nitrogen application and leaf harvesting improves yield and nutritional quality of beetroot. HortTechnology 2017, 27, 337–343. [Google Scholar] [CrossRef]
- Stagnari, F.A.; Galieni, A.; Speca, S.; Pisante, M. Water stress effects on growth, yield and quality traits of red beet. Sci. Hortic. 2013. [Google Scholar] [CrossRef]
- Penuelas, J.; Filella, I.; Gamon, J.A. Assessment of photosynthetic radiation-use efficiency with spectral reflectance. New Phytol. 1995, 131, 291–296. [Google Scholar] [CrossRef] [Green Version]
- Stagnari, F.A.; Galieni, A.; Cafiero, G.; Pisante, M. Application of photoselective films to manipulate wavelength of transmitted radiation and photosynthate composition in red beet (Beta vulgaris var. conditiva Alef.). J. Sci. Food Agric. 2013. [Google Scholar] [CrossRef]
- Stagnari, F.A.; Galieni, A.; Speca, S.; Cafiero, G.; Pisante, M. Effect of light and water supply on morphological and physiological leaf traits of red beet. Agron. J. 2014, 106, 459–468. [Google Scholar] [CrossRef]
- USDA. United States Standards for Grades of Canned Beets (81 FR 93572). Effective 20 January 2017. Available online: https://www.ams.usda.gov/sites/default/files/media/CannedBeetsStandard.pdf (accessed on 28 March 2018).
- Byford, W.J.; Gambogi, P. Phoma and other fungi on beet seed. Trans. Br. Mycol. Soc. 1985, 84, 21–28. [Google Scholar] [CrossRef]
- Abawi, G.S.; Crosier, D.C.; Becker, R.F. Symptomatology and etiology of root rot of table beets in New York. Phytopathology 1974, 1, 132. [Google Scholar]
- Olaya, G.; Abawi, G.S. Occurrence of Thanatephorus cucumeris on table beets in New York State (Abstr.). Phytopathology 1991, 81, 1186. [Google Scholar]
- Olaya, G.; Abawi, G.S. Characteristics of Rhizoctonia solani and binucleate Rhizoctonia species causing foliar blight and root rot on table beets in New York State. Plant Dis. 1994, 78, 800–804. [Google Scholar] [CrossRef]
- Olaya, G.; Abawi, G.S. Influence of inoculum type and moisture on development of Rhizoctonia solani on foliage of table beets. Plant Dis. 1994, 78, 805–810. [Google Scholar] [CrossRef]
- Shah, D.A.; Stivers-Young, L. Factors associated with decay in table beets identified by a statistical analysis of grower and processor records. HortTechnology 2004, 14, 25–29. [Google Scholar]
- Hoch, H.C.; Abawi, G.S. Biological control of Pythium root rot of table beet with Corticium sp. Phytopathology 1979, 69, 417–419. [Google Scholar] [CrossRef]
- Sadeghi, A.; Hesan, A.R.; Askari, H.; Qomi, D.N.; Farsi, M.; Hervan, E.M. Biocontrol of Rhizoctonia solani damping-off of sugar beet with native Streptomyces strains under field conditions. Biocontrol Sci. Technol. 2009, 19, 985–991. [Google Scholar] [CrossRef]
- Webb, K.M.; Harveson, R.M.; West, M.S. Evaluation of Rhizoctonia zeae as a potential biological control option for fungal root diseases of sugar beet. Ann. Appl. Biol. 2015, 167, 67–89. [Google Scholar] [CrossRef]
- Dube, B.; Smart, G.C. Biological control of Meloidogyne incognita by Paecilomyces lilacinus and Pasteuria penetrans. J. Nematol. 1987, 19, 222–227. [Google Scholar] [PubMed]
- Xu, X.M.; Jeffries, P.; Pautasso, M.; Jeger, M.J. Combined use of biological control agents to management plant diseases in theory and practice. Phytopathology 2011, 101, 1024–1031. [Google Scholar] [CrossRef] [PubMed]
- Weiland, J.; Koch, G. Sugarbeet leaf spot disease (Cercospora beticola Sacc.). Mol. Plant Pathol. 2004, 5, 157–166. [Google Scholar] [CrossRef] [PubMed]
- Pethybridge, S.J.; Vaghefi, N.; Kikkert, J.R. Management of Cercospora leaf spot in conventional and organic table beet production. Plant Dis. 2017, 101, 1642–1651. [Google Scholar] [CrossRef]
- McKay, M.B.; Pool, V.W. Field studies of Cercospora beticola. Phytopathology 1918, 8, 119–136. [Google Scholar]
- Vereijssen, J.; Schneider, H.J.H.M.; Termorshuizen, A.A.J. Possible root infection of Cercospora beticola in sugar beet. Eur. J. Plant Pathol. 2004, 110, 103–106. [Google Scholar] [CrossRef]
- Vereijssen, J.; Schneider, H.J.H.M.; Termorshuizen, A.A.J. Root infection of sugar beet by Cercospora beticola in a climate chamber and in the field. Eur. J. Plant Pathol. 2005, 112, 201–210. [Google Scholar] [CrossRef]
- Giannopolitis, C.N. Lesions on sugar beet roots caused by Cercospora beticola. Plant Dis. Rep. 1978, 62, 424–427. [Google Scholar]
- Shane, W.W.; Teng, P.S. Impact of Cercospora leaf spot on root weight, sugar yield and purity of Beta vulgaris. Plant Dis. 1982, 76, 812–820. [Google Scholar] [CrossRef]
- Wolf, P.F.J.; Weis, F.J.; Verreet, J.A. Influence of different cropping systems and threshold values on the epidemiological behaviour of Cercospora beticola in sugar beet. Meded. Fac. Landbouwkd. Toegep. Biol. Wet. 1995, 60, 431–438. [Google Scholar]
- Lawrence, J.S.; Meredith, D.S. Wind dispersal of conidia of Cercospora beticola. Phytopathology 1970, 60, 1076–1078. [Google Scholar] [CrossRef]
- Vereijssen, J.; Schneider, J.H.M.; Stein, A.; Jeger, M.J. Spatial pattern of Cercospora leaf spot of sugar beet in fields in long- and recently-established area. Eur. J. Plant Pathol. 2006, 116, 187–198. [Google Scholar] [CrossRef]
- Vereijssen, J.; Schneider, J.H.; Jeger, M.J. Epidemiology of Cercospora leaf spot on sugar beet: Modeling disease dynamics within and between plants. Phytopathology 2007, 97, 1550–1557. [Google Scholar] [CrossRef] [PubMed]
- Lartey, R.T.; Caesar-TonThat, T.C.; Caesar, A.J.; Shelver, W.L.; Sol, N.I.; Berman, J.W. Safflower: A new host of Cercospora beticola. Plant Dis. 2005, 89, 797–801. [Google Scholar] [CrossRef]
- Groenewald, M.; Groenewald, J.Z.; Braun, U.; Crous, P. Host range of Cercospora apii and C. beticola and description of C. apiicola, a novel species from celery. Mycologia 2006, 98, 275–285. [Google Scholar] [CrossRef] [PubMed]
- Vestal, E.F. Pathogenicity, host response and control of Cercospora leaf-spot in sugar beets. Res. Bull. 1933, 168, 43–72. [Google Scholar]
- Knight, N.L.; Vaghefi, N.; Hansen, Z.R.; Kikkert, J.R.; Pethybridge, S.J. Temporal genetic differentiation of Cercospora beticola populations in New York table beet fields. Plant Dis. 2018. [Google Scholar] [CrossRef]
- Vaghefi, N.; Nelson, S.C.; Kikkert, J.R.; Pethybridge, S.J. Genetic structure of Cercospora beticola populations on Beta vulgaris in New York and Hawaii. Sci. Rep. 2017, 7, 1726. [Google Scholar] [CrossRef] [PubMed]
- Groenewald, M.; Linde, C.C.; Groenewald, J.Z.; Crous, P.W. Indirect evidence for sexual reproduction in Cercospora beticola populations from sugar beet. Plant Pathol. 2008, 57, 25–32. [Google Scholar] [CrossRef]
- Groenewald, M.; Groenewald, J.Z.; Harrington, T.C.; Abeln, E.C.A.; Crous, P.W. Mating type gene analysis in apparently asexual Cercospora species is suggestive of cryptic sex. Fungal Genet. Biol. 2006, 43, 813–825. [Google Scholar] [CrossRef] [PubMed]
- Bolton, M.D.; Secor, G.A.; Rivera, V.; Weiland, J.J.; Rudolph, K.; Birla, K.; Rengifo, J.; Campbell, L.G. Evaluation of the potential for sexual reproduction in field populations of Cercospora beticola from USA. Fungal Biol. 2012, 116, 511–521. [Google Scholar] [CrossRef] [PubMed]
- Bolton, M.D.; de Jonge, R.; Inderbitzen, P.; Liu, Z.; Birla, K.; Van de Peer, Y.; Subbarao, K.V.; Thomma, B.P.H.J.; Secor, G.A. The heterothallic sugarbeet pathogen Cercospora beticola contains exon fragments of both MAT genes that are homogenized by concerted evolution. Fungal Genet. Biol. 2014, 62, 43–54. [Google Scholar] [CrossRef] [PubMed]
- Vaghefi, N.; Hay, F.S.; Kikkert, J.R.; Pethybridge, S.J. Genotypic diversity and resistance to azoxystrobin of Cercospora beticola on processing table beet in New York. Plant Dis. 2016, 100, 1466–1473. [Google Scholar] [CrossRef]
- Vaghefi, N.; Kikkert, J.R.; Bolton, M.D.; Hanson, L.E.; Secor, G.A.; Nelson, S.C.; Pethybridge, S.J. Global genotype flow in Cercospora beticola populations confirmed through genotyping-by sequencing. PLoS ONE 2017, 12, e0186488. [Google Scholar] [CrossRef] [PubMed]
- Khan, J.; del Rio, L.E.; Nelson, R.; Rivera-Varas, V.; Secor, G.A.; Khan, M.F.R. Survival, dispersal and primary infection site for Cercospora beticola in sugar beet. Plant Dis. 2008, 92, 741–745. [Google Scholar] [CrossRef]
- Pethybridge, S.J.; Vaghefi, N.; Kikkert, J.R. Horticultural characteristics and susceptibility of table beet cultivars to Cercospora leaf spot in New York. HortTechnology 2017, 27, 530–538. [Google Scholar] [CrossRef]
- Abawi, G.S.; Kikkert, J.R.; Moktan, K.; Lange, H.W.; Smart, C.D. First report of resistance to Quadris among populations of Cercospora beticola causing leaf spot of table beet in New York State. (Abstr.) Phytopathology 2014, 104, 1. [Google Scholar]
- Bartlett, D.W.; Clough, J.M.; Godwin, J.R.; Hall, A.A.; Hamer, M.; Parr-Dobrzanski, B. The strobilurin fungicides. Pest Manag. Sci. 2002, 58, 649–662. [Google Scholar] [CrossRef] [PubMed]
- Fisher, N.; Meunier, B. Molecular basis of resistance to cytochrome bc1 inhibitors. FEMS Yeast Res. 2008, 8, 183–192. [Google Scholar] [CrossRef] [PubMed]
- Pethybridge, S.J. Fungicide Resistance Monitoring in C. beticola; Cornell University: Geneva, NY, USA, 2017; Unpublished data. [Google Scholar]
- Gugino, B.K.; Ludwig, J.W.; Abawi, G.S. Evaluation of fungicides for the control of pocket rot and leaf spot of table beet, 2007. Plant Dis. Manag. Rep. 2008, 2, V156. [Google Scholar]
- McGrath, M.T. What are fungicides? Plant Health Instr. 2004. [Google Scholar] [CrossRef]
- Karaoglanidis, G.S.; Bardas, G. Control of benzimidazole- and DMI-resistant strains of Cercospora beticola with strobilurin fungicides. Plant Dis. 2006, 90, 419–424. [Google Scholar] [CrossRef]
- Secor, G.A.; Rivera, V.V.; Khan, M.F.R.; Gudmestad, N.C. Monitoring fungicide sensitivity of Cercospora beticola of sugar beet for disease management decisions. Plant Dis. 2010, 94, 1272–1282. [Google Scholar] [CrossRef]
- Bolton, M.D.; Birla, K.; Rivera-Varas, V.; Rudolph, K.D.; Secor, G.A. Characterization of CbCyp51 from field isolates of Cercospora beticola. Phytopathology 2012, 102, 298–305. [Google Scholar] [CrossRef] [PubMed]
- Trueman, C.J.; Rosenzweig, N.; Somohano, P.; Hanson, L.E. First report of demethylase inhibitor fungicide resistance in Cercospora beticola from sugar beet in Ontario, Canada. New Dis. Rep. 2017, 36, 20. [Google Scholar] [CrossRef]
- Wightwick, A.M.; Mollah, M.R.; Partington, D.L.; Allinson, G. Copper fungicide residues in Australian vineyard soils. J. Agric. Food Chem. 2008, 56, 2457–2464. [Google Scholar] [CrossRef] [PubMed]
- Gent, D.H.; Mahaffee, W.F.; McRoberts, N.; Pfender, W. The use and role of predictive systems in disease management. Annu. Rev. Phytopathol. 2013, 51, 267–289. [Google Scholar] [CrossRef] [PubMed]
- Khan, J.; del Rio, L.E.; Nelson, R.; Khan, M.F.R. Improving the Cercospora leaf spot management model for sugar beet in Minnesota and North Dakota. Plant Dis. 2007, 91, 1105–1108. [Google Scholar] [CrossRef]
- Windels, C.E.; Lamey, H.A.; Dave, H.; Widner, J.; Knudsen, T.A. Cercospora leaf spot model for sugar beet in practice by an industry. Plant Dis. 1998, 82, 716–726. [Google Scholar] [CrossRef]
- Rossi, V.; Battilani, P. CERCOPRI: A forecasting model for primary infections of Cercospora leaf spot of sugarbeet. EPPO Bull. 1991, 21, 527–531. [Google Scholar] [CrossRef]
- Wolf, P.F.J.; Verreet, J.A. An integrated pest management system in Germany for the control of fungal leaf diseases in sugar beet. The IPM sugar beet model. Plant Dis. 2002, 86, 336–344. [Google Scholar] [CrossRef]
- Jacobsen, B. Bacterial leaf spot. In Compendium of Beet Diseases and Other Pests; APS Press: St. Paul, MN, USA, 2009; ISBN 978-0-89054-365-8. [Google Scholar]
- O’Brien, R.G.; Sparshott, P.E. Bacterial blight in Queensland beetroot crops. Australas. Plant Pathol. 1999, 28, 182. [Google Scholar] [CrossRef]
- Ark, P.A.; Leach, L.D. Seed transmission of bacterial blight of sugar beet. Phytopathology 1946, 36, 549–553. [Google Scholar] [PubMed]
- Riffaud, C.M.-H.; Morris, C.E. Detection of Pseudomonas syringae pv. aptata in irrigation water retention basins by immunofluorescence colony-staining. Eur. J. Plant Pathol. 2002, 108, 539–545. [Google Scholar] [CrossRef]
- Pethybridge, S.J.; Kikkert, J.R. Control of Foliar Disease of Table Beet Using OMRI-Approved Products; Cornell University: Geneva, NY, USA, 2017; Unpublished data. [Google Scholar]
- Pool, V.W.; McKay, M.B. Phoma betae on the leaves of sugar beet. J. Agric. Res. 1915, 4, 169–177. [Google Scholar]
- Monte, E.; Garcia-Acha, I. Vegetative and reproductive structures of Phoma betae in vitro. Trans. Br. Mycol. Soc. 1998, 90, 233–245. [Google Scholar] [CrossRef]
- Vaghefi, N.; Kikkert, J.R.; Pethybridge, S.J. Incidence and pathogenicity of Didymella americana on table beet in New York. Plant Health Prog. 2016, 17, 270–272. [Google Scholar] [CrossRef]
- Edson, H.A. Seedling diseases of sugar beets and their relation to root-rot and crown-rot. J. Agric. Res. 1915, 4, 135–168. [Google Scholar]
- Herr, L.J. Hot water treatment for elimination of seed-borne Phoma betae and other microbial contaminants from sugar beet seed. J. Am. Soc. Sugar Beet Technol. 1971, 16, 568–574. [Google Scholar] [CrossRef]
- Mangan, A. New method for the detection of Pleospora bjoerlingii infection of sugar beet seed. Trans. Br. Mycol. Soc. 1971, 57, 169–172. [Google Scholar] [CrossRef]
- Bugbee, W.M. A selective medium for the isolation and enumeration of Phoma betae in soil and seed. Phytopathology 1974, 64, 706–708. [Google Scholar] [CrossRef]
- Bugbee, W.M.; Soine, O.C. Survival of Phoma betae in soil. Phytopathology 1974, 64, 1258–1260. [Google Scholar] [CrossRef]
- Maude, R.B.; Vizor, A.S.; Shuring, C.G. The control of fungal seed borne diseases by means of a thiram seed soak. Ann. Appl. Biol. 1969, 64, 245–257. [Google Scholar] [CrossRef]
- Abawi, G.S.; Crosier, D.C.; Cobb, A.C.; Becker, R.F. Root Rot of Table Beets in New York State; N.Y. Food Life Sci. Bull. No. 115; New York State Agricultural Experiment Station: Geneva, NY, USA, 1986. [Google Scholar]
- Scott, P.T.; Martin, H.L.; Boreel, S.M.; Wearing, A.H.; Maclean, D.J. Molecular characterization, pathogenicity, and fungicide sensitivity of Pythium spp. from table beet (Beta vulgaris var. vulgaris) grown in the Lockyer Valley, Queensland. Australas. Plant Pathol. 2005, 34, 361–368. [Google Scholar] [CrossRef]
- Abada, K.A. Fungi causing damping-off and root rot on sugar beet and their biological control with Trichoderma harzianum. Agric. Ecosyst. Environ. 1994, 51, 333–337. [Google Scholar] [CrossRef]
- Martin, F.N.; Loper, J.E. Soilborne plant diseases caused by Pythium spp.: Ecology, epidemiology, and prospects for biological control. Crit. Rev. Plant Sci. 1999, 18, 111–181. [Google Scholar] [CrossRef]
- Ohkura, M.; Abawi, G.S.; Smart, C.D.; Hodge, K.T. Diversity and aggressiveness of Rhizoctonia solani and Rhizoctonia-like fungi on vegetables in New York. Plant Dis. 2009, 93, 615–624. [Google Scholar] [CrossRef]
- Harveson, R.M.; Nielsen, K.A.; Eskridge, K.M. Utilizing a preplant soil test for predicting and estimating root rot severity in sugar beet in the Central High Plains of the United States. Plant Dis. 2014, 98, 1248–1252. [Google Scholar] [CrossRef]
- Ophel-Keller, K.; McKay, A.; Hartley, D.; Curran, J. Development of a routine DNA-based testing service for soilborne diseases in Australia. Australas. Plant Pathol. 2008, 37, 243–253. [Google Scholar] [CrossRef]
- Fox, R.T.V. Rapid methods for diagnosis of soil-borne plant pathogens. Soil Use Manag. 2009, 6, 179–183. [Google Scholar] [CrossRef]
- Tegg, R.; Corkrey, R.; Herdina, H.; McKay, A.C.; Crump, N.S.; de Boer, R.F.; Wiechel, T.J.; Wilson, C.R. Modeling pathogen DNA content and visual disease assessment in seed tubers to inform disease in potato progeny root, stolon and tubers. Plant Dis. 2015, 99, 50–57. [Google Scholar] [CrossRef]
- Hay, F.S.; Herdina, O.-K.K.; Hartley, D.; Pethybridge, S.J. Prediction of potato tuber damage by root-knot nematodes using soilborne DNA. Plant Dis. 2016, 100, 592–600. [Google Scholar] [CrossRef]
- Gorny, A.M.; Hay, F.S.; Wang, X.; Pethybridge, S.J. Isolation of nematode DNA from 100 g of soil using Fe3O4 super paramagnetic nanoparticles. Nematology 2018, 20, 271–283. [Google Scholar] [CrossRef]
- Jacobson, B. Scab. In Compendium of Beet Diseases and Pests; APS Press: St. Paul, MN, USA, 2009; ISBN 978-0-89054-365-8. [Google Scholar]
- Braun, S.; Gevens, A.; Charkowski, A.; Allen, C.; Jansky, S. Potato common scab: A review of the causal pathogens, management practices, varietal resistance screening methods, and host resistance. Am. J. Potato Res. 2017, 94, 283–296. [Google Scholar] [CrossRef]
- Harveson, R.M. Southern Sclerotium root rot. In Compendium of Beet Diseases and Pests; APS Press: St. Paul, MN, USA, 2009; ISBN 978-0-89054-365-8. [Google Scholar]
- Pethybridge, S.J. Root Diseases of Table Beet; Cornell University: Geneva, NY, USA, 2018; Unpublished data. [Google Scholar]
- Mullen, J. Southern blight, Southern stem blight, White mold. Plant Health Instr. 2001. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Disease | Pathogen | Inoculum | Management Strategies | Control Tactics |
|---|---|---|---|---|
| Crop Stand | ||||
| Damping-off | Phoma betae | Soil and infested seed | Crop rotation and certified seed | Seed treatment |
| Pocket rot | Rhizoctonia solani | Soil | Crop rotation | Seed treatment and fungicide application (in-furrow and early season) |
| Damping-off | Pythium ultimum | Soil | Crop rotation | Seed treatment and fungicide application (in-furrow and early season) |
| Foliar Health | ||||
| Bacterial leaf spot | Pseudomonas syringae pv. aptata | Soil and infested seed | Certified seed | Pesticide applications (e.g., copper-based products) to foliage |
| Cercospora leaf spot | Cercospora beticola | Unknown | Crop rotation, site selection, cultivar selection, and burial of crop residue | Foliar fungicide applications |
| Root Health | ||||
| Root disease and decay | Rhizoctonia solani | Soil | Crop rotation and cultivar selection | Fungicide application (in-furrow and early season) |
| Pythium ultimum | Soil | Crop rotation | Fungicide seed treatment and in-furrow applications | |
| Phoma betae | Soil and infested seed | Crop rotation and certified seed | Seed treatment | |
| Aphanomyces cochlioides | Soil | Crop rotation | None | |
| Fusarium spp. | Soil | Crop rotation | None | |
| Common scab | Streptomyces spp. | Soil | Crop rotation, irrigation scheduling, and site selection (soil pH < 5.2) | None |
| Southern Sclerotium root rot | Athelia rolfsii | Soil | Crop rotation and site selection | None |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pethybridge, S.J.; Kikkert, J.R.; Hanson, L.E.; Nelson, S.C. Challenges and Prospects for Building Resilient Disease Management Strategies and Tactics for the New York Table Beet Industry. Agronomy 2018, 8, 112. https://doi.org/10.3390/agronomy8070112
Pethybridge SJ, Kikkert JR, Hanson LE, Nelson SC. Challenges and Prospects for Building Resilient Disease Management Strategies and Tactics for the New York Table Beet Industry. Agronomy. 2018; 8(7):112. https://doi.org/10.3390/agronomy8070112
Chicago/Turabian StylePethybridge, Sarah J., Julie R. Kikkert, Linda E. Hanson, and Scot C. Nelson. 2018. "Challenges and Prospects for Building Resilient Disease Management Strategies and Tactics for the New York Table Beet Industry" Agronomy 8, no. 7: 112. https://doi.org/10.3390/agronomy8070112