Good Riddance? Breaking Disease Susceptibility in the Era of New Breeding Technologies


Chair of Phytopathology, TUM School of Life Sciences Weihenstephan, Technical University of Munich, 85354 Freising, Germany
*
Author to whom correspondence should be addressed.
Agronomy 2018, 8(7), 114; https://doi.org/10.3390/agronomy8070114
Submission received: 11 June 2018
/
Revised: 29 June 2018
/
Accepted: 2 July 2018
/
Published: 5 July 2018
(This article belongs to the Special Issue Biotic and Abiotic Stress Responses in Crop Plants)
{kind=link}
{kind=link}
Abstract
:Despite a high abundance and diversity of natural plant pathogens, plant disease susceptibility is rare. In agriculture however, disease epidemics often occur when virulent pathogens successfully overcome immunity of a single genotype grown in monoculture. Disease epidemics are partially controlled by chemical and genetic plant protection, but pathogen populations show a high potential to adapt to new cultivars or chemical control agents. Therefore, new strategies in breeding and biotechnology are required to obtain durable disease resistance. Generating and exploiting a genetic loss of susceptibility is one of the recent strategies. Better understanding of host susceptibility genes (S) and new breeding technologies now enable the targeted mutation of S genes for genetic plant protection. Here we summarize biological functions of susceptibility factors and both conventional and DNA nuclease-based technologies for the exploitation of S genes. We further discuss the potential trade-offs and whether the genetic loss of susceptibility can provide durable disease resistance.
1. Introduction
In crop production systems, plant diseases are controlled by standard field management practices (e.g., crop rotation, ploughing), usage of disease-resistant cultivars and pesticide applications. However, disease resistance and pesticide efficacy are often not durable because pathogen populations rapidly adapt to the selection pressure that is exerted by these disease control mechanisms. This and potentially harmful effects of pesticides on off-target organisms can render plant protection unsustainable, necessitating novel approaches to combat plant pathogens. In recent years, fundamental research on molecular plant-microbe interactions has revealed new insights on how plants defend themselves against pathogens and how pathogens subvert plant immunity. This knowledge and the development of new breeding technologies holds the potential for innovative approaches in genetic plant protection, which could complement the limitations of conventional technologies to provide greater resistance durability [1].
In plants, invading pathogens are challenged at several levels of plant-pathogen interactions [2]. Preformed defensive barriers together with pathogen-induced plant defense responses successfully restrict parasitic growth on resistant plants. Induced plant defenses have, however, led to the adaptation of pathogens to certain host species and the evolution of host-specific virulence strategies. This includes the secretion of proteinaceous and non-proteinaceous pathogenicity factors that support pathogen virulence. Since these so-called effector molecules are required to actively overcome host immune barriers, the term “effector-triggered susceptibility” (ETS) was coined [1]. Most reports on effector activities show that effectors function as suppressors of plant immune receptor functions, signal transduction and defense reactions [3]. Hence, the suppression of plant defense appears to be pivotal for virulence. Effectors can either specifically modulate host immune processes or more broadly influence host physiology. The latter often contributes to the development of disease symptoms. Such effectors may even provoke strong symptoms when applied as pure substances to plants. In this case, they are considered as toxins that can act either host-specifically or host-nonspecifically. Some effectors influence plant development and some pathogens produce plant hormones or hormone analogs to manipulate host development or physiology.
Pathogen detection either takes place at the plant cell surface, where surface receptor complexes function, or in the host plant cytoplasm or nucleoplasm by intracellular receptor proteins or receptor complexes [1,4]. Plant immune receptors (so-called resistance proteins encoded by major disease resistance genes [R]) detect the presence of effector proteins in a race-cultivar specific manner as determined by monogenic inheritance in both the host and parasite. At the molecular level, this classical gene-for-gene model is described by the term “effector-triggered immunity” (ETI) [1]. A more basal, race-nonspecific type of immunity operates within the broader context of ETS and ETI and it is mediated by the detection of a broad spectrum of non-self or altered-self molecules. This is collectively summarized as pattern-triggered immunity (PTI) [4,5].
2. The Principle of Susceptibility Genes and How to Find Them
Host immune components are encoded by dominantly inherited genes, which show either major (qualitative) or minor (quantitative) effects on disease resistance. However, the observation that disease resistance can also be recessively inherited indicates that pathogens can also profit from dominantly inherited host functions or susceptibility factors [1,6,7]. The corresponding dominantly inherited genes are called susceptibility genes (S). Recessive s genes have been successfully used in conventional and marker assisted plant breeding for the improvement of disease resistance. Recessive mlo (mildew locus o), several virus resistance genes and ToxA-insensitive tsn1 genes are prominent examples for this [8,9]. Here we discuss the mechanisms of disease susceptibility and how provoking and exploiting genetic loss of susceptibility can aid durable disease resistance.
Farmers and breeders have selected naturally occurring mutations of s genes for the improvement of crop health. Additionally, breeders and researchers searching for mutagenesis-induced resistance in crop and model plants have identified a broad variety of recessive disease resistances. High research effort over the last three decades enabled the identification of several of the corresponding mutations in S genes and first insights into the mechanisms of disease susceptibility.
The mlo-mediated powdery mildew resistance is perhaps the most prominent example of recessive plant disease resistance. It is of particular interest, because it is race-nonspecific and durable in the field. The MLO-gene was originally characterized in spring barley but it seems to function in all interactions in which MLO S-gene functions have been studied in detail [8]. Ethiopian highland farmers may have originally selected the barley mlo-11 allele in old land races, collected during expeditions in the 1930s, and used later in European plant breeding in the 1970s [10]. However, the first description of mlo goes back to an X-ray induced powdery mildew mutant generated in the 1940s [11]. Across multiple plant species, many mutagenesis-derived loss-of-function mlo alleles exist and MLO null-mutants are generally resistant to powdery mildew assuming no genetic redundancy with other MLO family members exists. The exact biochemical function of the f protein is not understood, but it may act as a negative regulator of pathogen-triggered and spontaneous defense reactions, putting mlo mutants in a primed defensive status [8,12].
The model plant Arabidopsis thaliana has been instrumental for the identification of many more susceptibility factors. For instance, forward genetic screens for powdery mildew resistance (pmr) or gene expression studies of compatible interactions with diverse biotrophic pathogens, followed by reverse genetic approaches have identified several candidate S genes [6,13,14,15]. Additionally, educated guesses and translational approaches have proved similarly successful in discovering S genes in crop plants [16,17]. In vivo protein-protein interaction screens are yet another suitable approach to identify susceptibility factors using effector proteins as bait [18,19]. Candidate susceptibility genes were also identified via host gene expression profiling, as it was shown with Hyaloperonospora arabidopsidis-infected Arabidopsis thaliana and Phytophthora cinnamoni-infected Castanea [20,21]. Once an S gene is identified, studying the physiological function of the susceptibility factor and genetic or physical interactions can identify susceptibility mechanisms or associated pathways and thereby new susceptibility factors [22,23].
3. Biological Functions of Susceptibility Genes
Considering the role of susceptibility genes in compatible plant-microbe interactions, the question arises as to what physiological function host susceptibility factors (or compatibility factors in terms of microbial symbiosis) may exert in healthy and microbe-attacked plants (Figure 1). Some plant susceptibility factors are regulators of host defense responses or cell death. Depending on whether the pathogen is a biotroph, hemibiotroph, or a necrotroph, it can be more or less sensitive to individual plant defense reactions or even profit from host cell-death. Biotrophs often profit from negative regulators of host defense reactions or cell death whereas necrotrophs can profit from host programmed cell-death. This might explain why certain host susceptibility factors show an ambivalent character and can turn into a resistance factor in interaction with another pathogen (see also chapter 5 for trade-offs below). Similarly, individual plant hormone pathways can positively or negatively influence plant-pathogen interactions, depending on the pathogen’s lifestyle [24]. In other cases, susceptibility factors do not have reported functions in regulating plant defense. They could be involved in physiological reprogramming of the susceptible host to establish and maintain a compatible interaction. This is particularly well described for the interaction with biotrophic pathogens that show a tight parasitic symbiosis with their host plants and appear to depend on many host functions for disease development. An increasing amount of publications support that successful obligate biotrophs not only successfully inhibit plant immunity, but also heavily rely on and reorganize host cell physiology and development. In the next paragraphs we discuss some prominent examples of S gene functions. For more comprehensive overviews, we refer to other review articles [25,26,27].
3.1. Host Cues for Recognition by the Pathogen
Upon first contact of a pathogen with the host aerial surface or rhizosphere, silent pathogen genes need to be activated e.g., for germination, directed growth, the development of infection structures, and for secretion of virulence factors. Gene activation requires recognition of host cues that trigger pathogen development. For example, epicuticular waxes and cutins of plants provide such cues for germination and formation of appressoria (the pathogen organ for surface adherence and cell wall penetration) by oomycetes, powdery mildew, anthracnose, and rust fungi. Correspondingly, plant mutants that show alterations in leaf wax composition can be less susceptible to fungal invasion [28,29,30,31,32,33]. Wild type forms of mutated genes that function in or interfere with biosynthesis of wax or cutin components can be considered as S genes. There are also reports about other types of chemical cues from the host (e.g., volatiles, flavonoids, acetosyringone, etc.), which are considered to be responsible for metabolic compatibility with adapted pathogens [34,35,36], and corresponding biosynthetic pathways may contain S factors. Additionally, plant surface hydrophobicity and topology trigger early pathogen development on susceptible plants [37,38].
3.2. Support of Pathogen Demands
Host-adapted necrotrophs can usually tolerate preformed and induced plant defensive barriers, but biotrophs, in particular, require additional support from the host because they lack saprophytic potential [39]. This host support means that, for example, the plant is involved in establishing pathogen feeding structures (haustoria or analogous feeding hyphae) in living host cells. Additionally, the host actively provides nutrients, e.g., by changes in carbohydrate metabolism, sugar transport, or carbohydrate source-sink transitions. Some obligate biotrophs have lost certain biosynthetic pathways, and hence, they might depend on host metabolite supply for primary or secondary metabolite biosynthesis. Thus, the pathogen requires plant components, whilst simultaneously suppressing the same plant’s defense. In plant-virus interactions, host primary metabolism and basic cell functions are involved in susceptibility because they provide the building blocks and biochemical machinery for synthesis of the virus itself. For instance, several components of the plant translation machinery contribute to virus replication and they are S factors in virus diseases [40]. An example for a host protein that supports fungal infection is the ROP GTPase RACB of barley, which is required for the susceptibility to fungal growth into epidermal cells and the expansion of haustoria of the powdery mildew pathogen Blumeria graminis f.sp. hordei. The physiological function of RACB in a healthy plant lies in polar cell development of leaf and root epidermal cells. RACB supports the outgrowth of root hairs from trichoblasts and the ingrowth of haustoria in leaf epidermal cells [41,42]. Another example is SWEET proteins; SWEET sugar transporters transport sucrose out of plant cells for reallocation of sugars. SWEETS are S factors, because they can be overexpressed in pathogen interactions and function in providing nutrients for the pathogen [43]. In summary, host cellular processes support certain demands of pathogens that feed from live tissue and the components of these processes can be S factors.
3.3. Control of Plant Defense Responses
Many S genes encode negative regulators of plant defense responses. Consequently, corresponding homozygous loss-of-function-mutants show either instantaneous defense responses or stronger defense responses after pathogen contact, which can be considered as a genetic priming mechanism. Mutant screens provided several lesion mimic or constitutive defense gene expression mutants, and in many of these mutants, stress hormone signaling is imbalanced. Prominent examples are lesion-simulating disease 1 (lsd1) or constitutive expressor of PR genes (e.g., cpr1 or cpr5). These mutants are usually less susceptible to biotrophic pathogens. In fewer cases, such mutants show a resistance to necrotrophs or broad-spectrum resistance [44]. Powdery mildew resistant mlo mutants show primed defense in young tissues and spontaneous defense in older tissues. However, in this particular case, it remains unclear as to whether deregulated defense is decisive for disease resistance because double mutants in mlo and stress hormone pathways lose spontaneous defense yet they retain pathogen resistance [45]. Genetic studies have shown that secondary indole metabolism appears crucial for mlo-mediated resistance in Arabidopsis [46], and vesicle fusion involved in protein secretion appears to be generally crucial for mlo-mediated resistance [45,47].
4. Effector Targets
Plant pathogenic microbes benefit from a certain repertoire of secreted effector proteins that interact with host molecules aiming to create a more favorable environment for the microbe (Figure 1). Hence, any host protein that directly or indirectly supports the susceptibility to any given phytopathogen represents an attractive potential effector target. Our knowledge of microbial effector proteins and susceptibility factors as genuine targets of these effectors has increased significantly over the last years [3,26], and some examples are described below.
4.1. Hub Proteins
Proteins at the center of signaling networks constitute hubs that are by definition key players during plant development and hence represent potential effector targets. In a recent study, a protein-protein interaction network was generated based on Arabidopsis thaliana host proteins and virulence effector proteins of biotrophic (Golovinomyces orontii and Hyaloperonospora arabidopsidis) and hemibiotrophic (Pseudomonas syringae) phytopathogens. The authors showed that certain plant proteins are targeted simultaneously by effector proteins from bacterial, fungal, and oomycete pathogens, thereby demonstrating exemplary effector convergence on key targets [19]. Several of these concurrently targeted plant proteins have also been shown to be susceptibility factors. Mutants of the COP9 signalosome complex subunit 5A (CSN5A), for example, showed enhanced disease resistance against both biotrophic and hemibiotrophic pathogens, suggesting a role for CSN5A in facilitating pathogen sustenance in the host. In contrast, transcription factor TCP14, being the most effector-targeted protein, seems to act as a susceptibility factor only in hemibiotrophic interactions, similar to other TCPs. Likewise, APC8, a protein involved in cell-cycle phase transitions, is one of the five most targeted hub proteins and acts as susceptibility factor in hemibiotrophic interactions [19]. The C3HC4 RING finger protein HUB1 was found to be targeted by only one pathogen effector protein. However, its function as susceptibility factor was demonstrated in hub1 mutant plants that showed enhanced disease resistance against biotrophic pathogens [19].
RIN4 (RPM1-INTERACTING PROTEIN 4) is another excellent and well-studied example of an effector-targeted susceptibility factor. This negative regulator of plant immune responses is also guarded by R proteins RPM1 (RESISTANCE TO PSEUDOMONAS SYRINGAE PV. MACULICOLA 1) and RPS2 (RESISTANT TO PSEUDOMONAS SYRINGAE 2), which are activated upon perception of the effectors AvrB-, AvrRpm1- or AvrRpt2-induced state modifications of RIN4 to subsequently trigger ETI. RIN4 nicely demonstrates that guarding of susceptibility factors by R-proteins sometimes is the plant’s only efficient way for protection, as opposed to S-gene mutation or the entire elimination from the genetic background, which regarding RIN4 would have detrimental pleiotropic consequences [48,49].
4.2. Bacterial Effector Targets
Certain race-specific susceptibility genes are targeted by transcription activator-like effectors (TALEs) of phytopathogenic bacteria from the genus Xanthomonas. They are known to drive host gene expression in a sequence-specific manner, leading to enhanced disease symptoms [50]. For example, AvrXa7 and PthXo3 activate the expression of sugar transporter OsSWEET14 in rice cultivars by directly binding to the effector binding element (EBE), which is located in the promoter region of OsSWEET14, OsSWEET11, and OsSWEET13, like OsSWEET14, are likewise major susceptibility genes and targets of TAL effectors [51,52]. Intriguingly, host transcription factors seem to be an attractive target for TALEs, as several other studies have now shown [53,54,55]. With the increasing amount of EBEs found in nature, the next years will continue to enrich the arsenal of TALE-targeted susceptibility genes.
4.3. Effector Targets of (Hemi)Biotrophic Pathogens
In contrast to bacteria, filamentous pathogens like fungi and oomycetes possess a plethora of effector proteins, indicating probable effector function redundancy. Biotrophs might, furthermore, require multiple effectors for manipulating host S factors, as suggested recently [56]. Barley MLO has not been described yet as a powdery mildew effector target, in contrast to its functional ortholog MLO2 from Arabidopsis thaliana, which is targeted by bacterial Pseudomonas syringae effector HopZ2 [8,57]. Likewise, the barley S factor ROP GTPase RACB was recently shown to interact with the ROP-interactive peptide 1 (ROPIP1) from Blumeria graminis f.sp. hordei. ROPIP1 is encoded on repetitive DNA, supports fungal penetration, and can provoke host cell microtubule disorganization. It may therefore represent an unconventional powdery mildew effector [58].
Hemibiotrophic pathogens require living host tissue during the initial stage of infection, suggesting that any host proteins that are involved in suppressing early resistance-associated cell death reactions might constitute susceptibility factors. For example, the effector Avr3a from the late blight pathogen Phytophthora infestans targets and stabilizes the E3 ubiquitin ligase CMPG1. CMPG1 degradation is required for INF1 elicitor-mediated cell death, which would, in turn, obstruct further pathogen spreading during the biotrophic phase of the infection [59]. A similar stabilization, followed by an enhanced infection, was observed with the putative potato K-homology (KH) RNA-binding protein KRBP1 that is targeted by P. infestans effector Pi04089 [60]. Targeting S factors is a well-exploited strategy that oomycetes like P. infestans follow, as additional studies have shown [61,62].
4.4. Effector Targets of Necrotrophic Pathogens
The pathogenicity of necrotrophs can also rely on secreted effectors that interact with host susceptibility factors to establish foliar necrosis and/or chlorosis in a cultivar-specific manner. For example, it was recently reported that some subunits of the membrane tethering exocyst complex from solanaceous plants seem to act as susceptibility factors for the grey mold pathogen Botrytis cinerea [63]. Profound knowledge of necrotrophic effector proteins was gained from studying effector proteins ToxA and ToxB from the tan spot pathogen Pyrenophora tritici-repentis, with ToxA being the best studied to date. The ToxA gene was acquired by horizontal gene transfer from the leaf blotch pathogen Parastagonospora nodorum [64]. ToxA itself interacts directly with a chloroplast-localised protein, ToxABP1. The presence of both, ToxA and ToxABP1, or the silencing of ToxABP1 in the absence of ToxA leads to similar necrotic phenotypes, indicating ToxA altering ToxABP1 function [65,66]. ToxA genetically interacts with the product of the toxin sensitivity gene Tsn1, which encodes a protein that resembles canonical Resistance proteins [9,67]. Functional TSN1 is involved in triggering ToxA-dependent cell death, which favors necrotrophic pathogen growth, and is thus a major S factor with agronomic relevance for both tan spot and leaf blotch in wheat [68].
5. Trade-offs and Prediction of Durability
S gene-based resistance has been suggested to be more durable than the widely applied R gene-based resistance [25]. Indeed, there are many reasons why R gene resistances, even when combined, might not be durable [69]. Thus, loss-of-susceptibility could potentially be a way to create more durable resistance. However, little is known about general S gene durability with only a few examples of long term durability being reported: Mlo resistance in barley has been in use for over 35 years and eIF4E-mediated virus resistance in pepper for almost 60 [11,70]. It is often thought that a combination of (functional) redundancy and pleiotropic effects contribute to the durability of S genes. Yet, which factors specifically contribute to this durability remains unclear. However, the non-race-specific nature of s gene-mediated resistance (e.g., mlo-mediated powdery mildew resistance) makes it less likely that population dynamics rapidly select for single races that are capable of circumventing the loss of susceptibility.
5.1. Broad Spectrum Resistance & Functionality
It has been hypothesized that S genes are more durable because of the central role they play in facilitating the initial infection stages. This could be tightly linked to biological functions and the physical environment of S factors. As described above, S factors can generally be considered as (indirect) negative regulators of immunity [71]. In s mutant genotypes, it is therefore possible that the whole plant is in a primed state or that certain defense-associated processes are more active, preventing the pathogen from establishing infection [13].
S factors are often intra-cellular and well-embedded in physiological pathways. It has been shown that certain S genes can be classified as protein hubs [19]. They are therefore unlikely to steer a single physiological process, which, when missing, can be easily circumvented by the pathogen. In the case of host S gene null mutants, pathogens would have to evolve new functions to overcome loss of susceptibility. This would be particularly relevant if an S factor serves an essential requirement for the pathogen. Such evolution of a new biological function can be considered to be slow or even impossible, according to complexity. In addition, the subcellular location and the presence of many possible guard and decoy proteins will impede even rapidly evolving effectors to reach and modify their intracellular target, as opposed to cell-surface receptors or apoplastic R proteins [72]. Thus, it is easier for a pathogen to lose or modify a single R-protein-recognized effector to tackle ETI than to overcome the loss of a host susceptibility gene.
5.2. Pleiotropy and Trade offs
S genes often have key physiological functions within the host. This implies a high chance of pleiotropic effects of S gene mutation. On the one hand, if the recessive s gene of interest displays deleterious phenotypes, even in a different genetic background, it would not be easily exploitable [26]. On the other hand, mutants that do not show pleiotropic effects are extremely valuable and they could provide sustainable solutions in resistance breeding. Interestingly, pleiotropic effects also complicate the assessment of both durability and the broadness of the resistance spectrum. For example, elF4E1 provides virus resistance in cultivated tomato (S. lycopersicum) and the wild species S. habrochaites, yet the null mutant in S. habrochaites show stronger and broader resistance than the S. lycopersicum mutant. The addition of an independent mutation in a related gene, elF4E2, did increase the strength and breadth of resistance, but lead to detrimental growth [73], although elF4E2 mutants alone do not confer resistance. Also, Mlo genes occur as gene families and in A. thaliana, independent mutations of three orthologs are required for mlo resistance, each having their own major or minor effect [46]. A trade-off has been shown between mlo resistance to powdery mildew and increased susceptibility to Magnaporthe grisea [74] or the toxins of the necrotroph Bipolaris sorokiniana [75]. On the other side, reduced pleiotropic effects in barley have been reported in moderate mlo variants [76]. So far, no trade-offs have been detected in tsn1 plants, but recent work by See et al. [67] suggested that the tan spot infection could develop in some wheat backgrounds, independent of the tsn1 allele. Moreover, the strong day/night rhythm of Tsn1 gene expression might indicate an additional function in wheat.
5.3. Decoys and Protection
Many S genes have allelic variants in the genome that may act as a “sponge” decoy [72], attracting effectors and triggering defense responses. In addition, the molecular inclusion of S gene domains in R genes has been reported, thus combining recessive resistance with active defense signaling. Pi21 is a gene that encodes an HMA domain containing protein that suppresses the plant defense response in rice [77]. HMA domains are virulence targets for multiple pathogens [78]. Additionally, HMA domains have also been found as integrated domains in R genes. Effectors from the rice blast fungus Magnaporthe oryzae bind to the HMA domain of the R protein Pik and trigger defense responses [79]. Many more integrated domains have been described to date [80], yet the question remains how many of these are “orthologs” of true s genes.
5.4. S Gene Diversity
Durability of S genes could possibly contribute to the fact that different homologs and s alleles are present in different plant cultivars or ecotypes. Assuming lack of pleiotropic effects, it will be theoretically possible to maintain many different s alleles in a population. Yet, evidence seems inconclusive. Additional elF4E1 alleles have been identified in wild tomato species, albeit with a limited resistance spectrum [81]. Conversely, in chili peppers (Capsicum annuum), many variants confer resistance and are thought to have originated from loci under positive selection. Nevertheless, it is striking that elF4E2 shows no polymorphisms in chili pepper [73]. In barley, the viral S factor HvEIF4E also occurs in many diverse resistance-conferring haplotypes. For another unrelated virus S gene, HvPDIL5-1, many haplotypes are present in wild barley and landraces, yet none of these are actual virus resistance conferring alleles (rym-alleles) [82,83]. The fact that several barley cultivars with different resistance conferring alleles exist, does suggest that the s alleles arose by mutation during domestication in an area where the virus was also present [83]. Also, Tsn1 shows large variety in spelt and other grasses, but in turn, it has hardly any polymorphisms in bread and durum wheat [9].
6. Exploitation of Susceptibility Genes
6.1. Breeding for Reduced Susceptibility/Loss of Susceptibility
In several cases, naturally occurring s genes have been identified in breeding material without actually knowing the nature of the corresponding dominant gene. In many other instances, random mutagenesis and selfing have produced disease resistant offspring with mutant s genes (Figure 2). Recessive resistance is best identified in inbreeding plant species or artificial double haploid plants. Cloning of S genes from crop plants has been successful in several cases and has sometimes facilitated the identification of related genes in other crop or model plant species. MLO is an s gene with many mlo resistance alleles being identified in diverse plant species, making it a “universal weapon” against powdery mildew [8]. Likewise, once ToxA and the Tsn1 tan spot S-gene of wheat were identified, it became straightforward to eliminate the susceptibility from breeding material. Even the phenotypic identification of Tsn1 genotypes became possible because ToxA, which is as a host-genotype specific toxin, could be directly applied to distinguish Tsn1 from tsn1 genotypes by differentially ToxA-provoked symptoms [67].
Another breeding strategy for loss of susceptibility starts from a known S gene and looks for natural or induced allelic diversity by TILLING (Targeting Induced Local Lesions In Genomes). TILLING has the advantage that it starts from mutagenesis of any desired genotype, followed by S gene re-sequencing, and can identify mutant s alleles even in complex genomes, such as that of hexaploid wheat [84]. The hexaploidy of bread wheat and the presence of three barley Mlo orthologues (TaMlo-A1, TaMlo-B1 and TaMlo-D1) make the natural occurrence of mlo mutants including pathogen resistance quite unlikely for bread wheat. Using TILLING technology to select partial loss-of-function alleles of TaMlo however enhanced powdery mildew resistance in some lines without negative pleiotropic effects [84]. Naturally occurring variation of S genes can be identified by EcoTILLING: re-sequencing S genes in natural populations of crop progenitors or land races. EcoTILLING identified a new allele of eIF4E for melon necrotic spot virus resistance [85], and similar to our previous example of HvPDIL5-1, a TILLING approach further confirmed the identity of the mutated S gene in rym1 genotypes [83].
6.2. Gene Editing Possibilities for Targeted Exploitation of Susceptibility Genes
Besides targeted S gene-associated traditional breeding strategies, there are several additional opportunities for exploiting S gene function (Figure 2). In fundamental research, susceptibility factors can be used as bait to either find antagonists that are involved in resistance or downstream interactors that themselves act as susceptibility factors. If the molecular mode-of-action of an S factor has already been elucidated, one way to inhibit its function can be the application of a biocompatible chemical treatment. Alternatively, the ectopic expression of dominant negative S gene variants might outcompete and eventually eliminate the endogenous S factor function. Genome editing using sequence-specific nuclease mutagenesis technologies has gained much attention over the last ten years, as it provides researchers with constantly improving tools like zinc finger nucleases (ZFNs), TALE nucleases (TALENs), and clustered regularly interspaced short palindrome repeats (CRISPR)/CRISPR-associated systems (Cas) to create transgene-free genetically modified crops [86,87,88]. CRISPR/Cas9-targeted editing has been successfully applied on the citrus susceptibility gene CsLOB1 and its promoter, resulting in resistance to Xanthomonas citri subsp. citri (Xcc), and also for creating novel alleles of rice eIF4G, which conferred resistance to Rice tungro spherical virus [89,90,91]. A strong disease resistance against powdery mildew has been achieved in tomato by transgene-free genetically CRISPR/Cas-mutated slmlo1, one of 16 Mlo genes in tomato [92], exemplifying again how powerful and versatile these novel genome editing approaches can be.
Furthermore, the EBE of OsSWEET14 is targeted by TALEs AvrXa7, PthXo3, TalC and Tal5 from Xanthomonas oryzae pv. oryzae. TALEN-mediated genome editing of AvrXa7 and Tal5 EBEs conferred resistance to bacterial strains relying on the corresponding TALE [93]. Nature itself teaches us of how to benefit from the EBE-promoter repertoire. To counteract TALE activity, several R proteins have recently been found in pepper and in rice that contain corresponding EBEs for Xanthomonas TALEs AvrBs3, AvrXa10, AvrXa23, and AvrXa27, displaying a promoter-trap strategy in a decoy-like manner to confer disease resistance [94,95,96,97]. Using TALENs or genome-based engineering, artificial EBE-promoter traps could be generated using EBEs of S gene promoter regions upstream of known R genes. Moreover, it is even feasible to combine different EBEs and R genes in a consecutive manner aiming for broad-spectrum resistance against a range of microbial pathogens [50]. Additionally, with the help of genome editing technology, broad resistance to different phytopathogenic fungi may be achieved by generating loss-of-function alleles of genes encoding HMA domain-containing proteins, like plant defense suppressor Pi21 [77,98].
6.3. Other Possibilities to Exploit S Factors
In the event that the constitutive knock-out or the silencing of susceptibility genes by genome editing is rendered impossible due to deleterious pleiotropic phenotypes, “silencing on demand” using pathogen-inducible promoters can be an alternative approach. In barley, the pathogen-inducible Hv-Ger4c promoter has been successfully used to control the expression of Ta-Lr34res, an ABC transporter that confers resistance against several fungal pathogens in wheat [99].
S genes can also be modified to give rise to artificial decoys that inform R proteins to trigger ETI. This neofunctionalization is of course only applicable for susceptibility factors that are effector targets. Targeting of the artificial decoy by the particular effector protein would consequentially lead to a dead end for this particular effector function. Artificial decoys based on susceptibility genes could eventually be even used to switch plant immunity between pathogen kingdoms, as it was recently shown for artificial R proteins. RPS5, which is an intracellular R protein from Arabidopsis thaliana, is normally activated upon the recognition of AVRPPHB SUSCEPTIBLE1 (PBS1) cleavage by Pseudomonas syringae effector AvrPphB, with PBS1 serving as a decoy. The AvrPphB cleavage site within PBS1 was substituted with cleavage sites for other pathogen protease effectors, e.g., protease effectors of Turnip mosaic virus, thereby conferring resistance to different pathogens [100].
7. Future Direction
We have discussed several methods and trade-offs for S gene exploitation (Figure 2). For the optimal exploitation of S genes, future research should focus on further unraveling the molecular mechanisms of S gene resistance. This is essential to identify novel susceptibility factors to increase our breeding capacities. Furthermore, intensive research is required to take full advantage of S gene exploitation by controlling and, in the best case, diminishing pleiotropic effects. Additionally, whole genome resequencing studies could reveal the diversity and variability of S genes in wild crop relatives and heirloom varieties. Combined with large scale protein-protein interaction studies, these findings can be put in a larger S gene defense signaling context. Such information will help to understand the durability of s gene resistance when compared to R gene resistance. One could also try to identify partial S gene mutants. Such genes might confer less, but still sufficient field resistance while suffering less from pleiotropic effects. Such genes might be found in natural populations, where they have been selected for, or they might be created by random or knowledge-based approaches.
These new findings can be used for modern breeding and genome editing technologies. In fact, transgenic and marker-assisted breeding have already been utilized for over several decades. More recently, new mutagenesis and gene editing approaches have also been shown to generate strong and functional s genes. Thus, the targeted exploitation of susceptibility factors forms a credible and potentially durable route for future resistance breeding.
Author Contributions
S.E., R.S. and R.H. designed and wrote the manuscript.
Funding
Experimental work on susceptibility factors in the laboratory of R.H. is funded by the German Research Foundation in frame of the collaborative research center SFB924 and HU886/8.
Acknowledgments
We thank Alexander Coleman (TU Munich) for critical reading of this manuscript.
Conflicts of Interest
The authors declare no conflict of interest. The founding sponsors had no role in the writing of the manuscript, and in the decision to publish.
References
- Dangl, J.L.; Horvath, D.M.; Staskawicz, B.J. Pivoting the plant immune system from dissection to deployment. Science 2013, 341, 746–751. [Google Scholar] [CrossRef] [PubMed]
- Thordal-Christensen, H. Fresh insights into processes of nonhost resistance. Curr. Opin. Plant Biol. 2003, 6, 351–357. [Google Scholar] [CrossRef]
- Presti, L.L.; Lanver, D.; Schweizer, G.; Tanaka, S.; Liang, L.; Tollot, M.; Zuccaro, A.; Reissmann, S.; Kahmann, R. Fungal effectors and plant susceptibility. Annu. Rev. Plant Biol. 2015, 66, 513–545. [Google Scholar] [CrossRef] [PubMed]
- Boller, T.; Felix, G. A renaissance of elicitors: Perception of microbe-associated molecular patterns and danger signals by pattern-recognition receptors. Annu. Rev. Plant Biol. 2009, 60, 379–406. [Google Scholar] [CrossRef] [PubMed]
- Ranf, S. Sensing of molecular patterns through cell surface immune receptors. Curr. Opin. Plant Biol. 2017, 38, 68–77. [Google Scholar] [CrossRef] [PubMed]
- Schulze-Lefert, P.; Vogel, J. Closing the ranks to attack by powdery mildew. Trends Plant Sci. 2000, 5, 343–348. [Google Scholar] [CrossRef]
- Hückelhoven, R. Powdery mildew susceptibility and biotrophic infection strategies. FEMS Microbiol. Lett. 2005, 245, 9–17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kusch, S.; Panstruga, R. mlo-based resistance: An apparently universal “weapon” to defeat powdery mildew disease. Mol. Plant Microbe Interact. 2017, 30, 179–189. [Google Scholar] [CrossRef] [PubMed]
- Faris, J.D.; Zhang, Z.C.; Lu, H.J.; Lu, S.W.; Reddy, L.; Cloutier, S.; Fellers, J.P.; Meinhardt, S.W.; Rasmussen, J.B.; Xu, S.S.; et al. A unique wheat disease resistance-like gene governs effector-triggered susceptibility to necrotrophic pathogens. Proc. Natl. Acad. Sci. USA 2010, 107, 13544–13549. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Piffanelli, P.; Ramsay, L.; Waugh, R.; Benabdelmouna, A.; D’Hont, A.; Hollricher, K.; Jørgensen, J.H.; Schulze-Lefert, P.; Panstruga, R. A barley cultivation-associated polymorphism conveys resistance to powdery mildew. Nature 2004, 430, 887–891. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jørgensen, H. Discovery, characterization and exploitation of Mlo powdery mildew resistance in barley. Euphytica 1992, 63, 141–152. [Google Scholar] [CrossRef]
- Peterhansel, C.; Freialddenhoven, A.; Kurth, J.; Kolsch, R.; Schulze-Lefert, P. Interaction analyses of genes required for resistance responses to powdery mildew in barley reveal distinct pathways leading to leaf cell death. Plant Cell 1997, 9, 1397–1409. [Google Scholar] [CrossRef] [PubMed]
- Vogel, J.; Somerville, S. Isolation and characterization of powdery mildew-resistant Arabidopsis mutants. Proc. Natl. Acad. Sci. USA 2000, 97, 1897–1902. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Radakovic, Z.S.; Anjam, M.S.; Escobar, E.; Chopra, D.; Cabrera, J.; Silva, A.C.; Escobar, C.; Sobczak, M.; Grundler, F.M.W.; Siddique, S. Arabidopsis HIPP27 is a host susceptibility gene for the beet cyst nematode Heterodera schachtii. Mol. Plant Pathol. 2018. [Google Scholar] [CrossRef] [PubMed]
- Douchkov, D.; Lück, S.; Johrde, A.; Nowara, D.; Himmelbach, A.; Rajaraman, J.; Stein, N.; Sharma, R.; Kilian, B.; Schweizer, P. Discovery of genes affecting resistance of barley to adapted and non-adapted powdery mildew fungi. Genome Biol. 2014, 15, 518. [Google Scholar] [CrossRef] [PubMed]
- Eichmann, R.; Bischof, M.; Weis, C.; Shaw, J.; Lacomme, C.; Schweizer, P.; Duchkov, D.; Hensel, G.; Kumlehn, J.; Hückelhoven, R. BAX INHIBITOR-1 is required for full susceptibility of barley to powdery mildew. Mol. Plant Microbe Interact. 2010, 23, 1217–1227. [Google Scholar] [CrossRef] [PubMed]
- Sun, K.; Wolters, A.A.M.; Vossen, J.H.; Rouwet, M.E.; Loonen, A.E.H.M.; Jacobsen, E.; Visser, R.G.F.; Bai, Y.L. Silencing of six susceptibility genes results in potato late blight resistance. Transgenic Res. 2016, 25, 731–742. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mukhtar, M.S.; Carvunis, A.R.; Dreze, M.; Epple, P.; Steinbrenner, J.; Moore, J.; Tasan, M.; Galli, M.; Hao, T.; Nishimura, M.T.; et al. Independently evolved virulence effectors converge onto hubs in a plant immune system network. Science 2011, 333, 596–601. [Google Scholar] [CrossRef] [PubMed]
- Weßling, R.; Epple, P.; Altmann, S.; He, Y.J.; Yang, L.; Henz, S.R.; McDonald, N.; Wiley, K.; Bader, K.C.; Gläßer, C.; et al. Convergent targeting of a common host protein-network by pathogens effectors from three kingdoms of life. Cell Host Microbe 2014, 16, 364–375. [Google Scholar] [CrossRef] [PubMed]
- Hok, S.; Danchin, E.G.J.; Allasia, V.; Panabières, F.; Attard, A.; Keller, H. An Arabidopsis (malectin-like) leucine-rich repeat receptor-like kinase contributes to downy mildew disease. Plant Cell Environ. 2011, 34, 1944–1957. [Google Scholar] [CrossRef] [PubMed]
- Santos, C.; Duarte, S.; Tedesco, S.; Fevereiro, P.; Costa, R.L. Expression profiling of Castanea genes during resistant and susceptible interactions with the oomycete pathogen Phytophthora cinnamomi reveal possible mechanisms of immunity. Front. Plant Sci. 2017, 8, 515. [Google Scholar] [CrossRef] [PubMed]
- Schultheiss, H.; Preuss, J.; Pircher, T.; Eichmann, R.; Hückelhoven, R. Barley RIC171 interacts with RACB in planta and supports entry of the powdery mildew fungus. Cell Microbiol. 2008, 10, 1815–1826. [Google Scholar] [CrossRef] [PubMed]
- Schnepf, V.; Vlot, A.C.; Kugler, K.; Hückelhoven, R. Barley susceptibility factor RACB modulates transcript levels of signalling protein genes in compatible interaction with Blumeria graminis f. sp. hordei. Mol. Plant Pathol. 2018, 19, 393–404. [Google Scholar] [CrossRef] [PubMed]
- Shigenaga, M.; Berens, M.L.; Tsuda, K.; Argueso, C.T. Towards engineering of hormonal crosstalk in plant immunity. Curr. Opin. Plant Biol. 2017, 38, 164–172. [Google Scholar] [CrossRef] [PubMed]
- Pavan, S.; Jacobsen, E.; Visser, R.F.; Bai, Y. Loss of susceptibility as a novel breeding strategy for durable and broad-spectrum resistance. Mol. Breed. 2010, 25, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Van Schie, C.C.N.; Takken, F.L.W. Susceptibility genes 101: How to be a good host. Annu. Rev. Phytopathol. 2014, 52, 551–581. [Google Scholar] [CrossRef] [PubMed]
- Lapin, D.; Van den Ackerveken, G. Susceptibility to plant disease: More than a failure of host immunity. Trends Plant Sci. 2013, 18, 546–554. [Google Scholar] [CrossRef] [PubMed]
- Uppalapati, S.R.; Ishiga, Y.; Doraiswamy, V.; Bedair, M.; Mittal, S.; Chen, J.H.; Nakashima, J.; Tang, Y.H.; Tadege, M.; Ratet, P.; et al. Loss of abaxial leaf epicuticular wax in Medicago truncatula irg1/palm1 mutants results in reduced spore differentiation of anthracnose and nonhost rust pathogens. Plant Cell 2012, 24, 353–370. [Google Scholar] [CrossRef] [PubMed]
- Hansjakob, M.; Riederer, U. Hildebrandt, Appressorium morphogenesis and cell cycle progression are linked in the grass powdery mildew fungus Blumeria graminis. Fungal Biol. 2012, 116, 890–901. [Google Scholar] [CrossRef] [PubMed]
- Wang, E.; Schornack, S.; Marsh, J.F.; Gobbato, E.; Schwessinger, B.; Eastmond, P.; Schultze, M.; Kamoun, S.; Oldroyd, G.E.D. A common signaling process that promotes mycorrhizal and oomycete colonization of plants. Curr. Biol. 2012, 22, 2242–2246. [Google Scholar] [CrossRef] [PubMed]
- Weis, C.; Hildebrandt, U.; Hoffmann, T.; Hemetsberger, C.; Pfeilmeier, S.; König, C.; Schwab, W.; Eichmann, R.; Hückelhoven, R. CYP83A1 is required for metabolic compatibility of Arabidopsis with the adapted powdery mildew fungus Erysiphe cruciferarum. New Phytol. 2014, 202, 1310–1319. [Google Scholar] [CrossRef] [PubMed]
- Weidenbach, D.; Jansen, M.; Franke, R.B.; Hensel, G.; Weissgerber, W.; Ulferts, S.; Jansen, I.; Schreiber, L.; Korzun, V.; Pontzen, R.; et al. Evolutionary conserved function of barley and Arabidopsis 3-KETOACYL-CoA SYNTHASES in providing wax signals for germination of powdery mildew fungi. Plant Physiol. 2014, 166, 1621–1633. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Haslam, T.M.; Krüger, A.; Schneider, L.M.; Mishina, K.; Samuels, L.; Yang, H.X.; Kunst, L.; Schaffrath, U.; Nawrath, C.; et al. The beta-ketoacyl-CoA Synthase HvKCS1, encoded by Cer-zh, plays a key role in synthesis of barley leaf wax and germination of barley powdery mildew. Plant Cell Physiol. 2018, 59, 811–827. [Google Scholar] [CrossRef] [PubMed]
- Tyler, B.M. Molecular basis of recognition between Phytophthora pathogens and their hosts. Annu. Rev. Phytopathol. 2002, 40, 137–167. [Google Scholar] [CrossRef] [PubMed]
- Mendgen, K.; Wirsel, S.G.; Jux, A.; Hoffmann, J.; Boland, W. Volatiles modulate the development of plant pathogenic rust fungi. Planta 2006, 224, 1353–1361. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hooykaas, P.; Beijersbergen, A.G. The virulence system of Agrobacterium tumefaciens. Annu. Rev. Phytopathol. 1994, 32, 157–181. [Google Scholar] [CrossRef]
- Read, N.; Kellock, L.; Knight, H.; Trewavas, A. Contact sensing during infection by fungal pathogens. Perspect. Plant Cell Recog. 1992, 48, 137–172. [Google Scholar]
- Mendgen, K.; Hahn, M.; Deising, H. Morphogenesis and mechanisms of penetration by plant pathogenic fungi. Annu. Rev. Phytopathol. 1996, 34, 367–386. [Google Scholar] [CrossRef] [PubMed]
- Hückelhoven, R.; Eichmann, R.; Weis, C.; Hoefle, C.; Proels, R.K. Genetic loss of susceptibility: A costly route to disease resistance? Plant Pathol. 2013, 62, 56–62. [Google Scholar] [CrossRef]
- Sanfacon, H. Plant translation factors and virus resistance. Viruses 2015, 7, 3392–3419. [Google Scholar] [CrossRef] [PubMed]
- Scheler, B.; Schnepf, V.; Galgenmüller, C.; Ranf, S.; Hückelhoven, R. Barley disease susceptibility factor RACB acts in epidermal cell polarity and positioning of the nucleus. J. Exp. Bot. 2016, 67, 3263–3275. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hoefle, C.; Huesmann, C.; Schultheiss, H.; Börnke, F.; Hensel, G.; Kumlehn, J.; Hückelhoven, R. A barley ROP GTPase ACTIVATING PROTEIN associates with microtubules and regulates entry of the barley powdery mildew fungus into leaf epidermal cells. Plant Cell 2011, 23, 2422–2439. [Google Scholar] [CrossRef] [PubMed]
- Chandran, D. Co-option of developmentally regulated plant SWEET transporters for pathogen nutrition and abiotic stress tolerance. IUBMB Life 2015, 67, 461–471. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lorrain, S.; Vailleau, F.; Balagué, C.; Roby, D. Lesion mimic mutants: Keys for deciphering cell death and defense pathways in plants? Trends Plant Sci. 2003, 8, 263–271. [Google Scholar] [CrossRef]
- Consonni, C.; Humphry, M.E.; Hartmann, H.A.; Livaja, M.; Durner, J.; Westphal, L.; Vogel, J.; Lipka, V.; Kemmerling, B.; Schulze-Lefert, P.; et al. Conserved requirement for a plant host cell protein in powdery mildew pathogenesis. Nat. Genet. 2006, 38, 716–720. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Consonni, C.; Bednarek, P.; Humphry, M.; Francocci, F.; Ferrari, S.; Harzen, A.; van Themaat, E.V.L.; Panstruga, R. Tryptophan-derived metabolites are required for antifungal defense in the Arabidopsis mlo2 mutant. Plant Physiol. 2010, 152, 1544–1561. [Google Scholar] [CrossRef] [PubMed]
- Collins, N.C.; Thordal-Christensen, H.; Lipka, V.; Bau, S.; Kombrink, E.; Qiu, J.L.; Hückelhoven, R.; Stein, M.; Freialdenhoven, A.; Somerville, S.C.; et al. SNARE-protein-mediated disease resistance at the plant cell wall. Nature 2003, 425, 973–977. [Google Scholar] [CrossRef] [PubMed]
- Khan, M.; Subramaniam, R.; Desveaux, D. Of guards, decoys, baits and traps: Pathogen perception in plants by type III effector sensors. Curr. Opin. Microbiol. 2016, 29, 49–55. [Google Scholar] [CrossRef] [PubMed]
- Kaundal, A.; Ramu, V.; Oh, S.; Lee, S.; Pant, B.D.; Lee, H.K.; Rojas, C.; Senthil-Kumar, M.; Mysore, K.S. GENERAL CONTROL NONREPRESSIBLE4 degrades 14-3-3 and the RIN4 complex to regulate stomatal aperture with implications on nonhost diseases resistance and drought tolerance. Plant Cell 2017, 29, 2233–2248. [Google Scholar] [CrossRef] [PubMed]
- Boch, J.; Bonas, U.; Lahaye, T. TAL effectors—Pathogen strategies and plant resistance engineering. New Phytol. 2014, 204, 823–832. [Google Scholar] [CrossRef] [PubMed]
- Yang, B.; Sugio, A.; White, F.F. Os8N3 is a host disease-susceptibility gene for bacterial blight of rice. Proc. Natl. Acad. Sci. USA 2006, 103, 10503–10508. [Google Scholar] [CrossRef] [PubMed]
- Huang, S.; Antony, G.; Li, T.; Liu, B.; Obasa, K.; Yang, B.; White, F.F. The broadly effective resistance gene xa5 of rice is a virulence effector-dependent quantitative trait for bacterial blight. Plant J. 2016, 86, 186–194. [Google Scholar] [CrossRef] [PubMed]
- Kay, S.; Hahn, S.; Marois, E.; Hause, G.; Bonas, U. A bacterial effector acts as a plant transcription factor and induces a cell size regulator. Science 2007, 318, 648–651. [Google Scholar] [CrossRef] [PubMed]
- Duan, S.; Jia, H.; Pang, Z.Q.; Teper, D.; White, F.; Jones, J.; Zhou, C.Y.; Wang, N. Functional characterization of the citrus canker susceptibility gene CsLOB1. Mol. Plant Pathol. 2018. [Google Scholar] [CrossRef] [PubMed]
- Ma, L.; Wang, Q.; Yuan, M.; Zou, T.T.; Yin, P.; Wang, S.P. Xanthomonas TAL effectors hijack host basal transcription factor IIA α and γ subunits for invasion. Biochem. Biophys. Res. Commun. 2018, 496, 608–613. [Google Scholar] [CrossRef] [PubMed]
- Thordal-Christensen, H.; Birch, P.R.J.; Spanu, P.D.; Panstruga, R. Why did filamentous plant pathogens evolve the potential to secrete hundreds of effectors to enable disease? Mol. Plant Pathol. 2018, 19, 781–785. [Google Scholar] [CrossRef] [PubMed]
- Lewis, J.D.; Wan, J.; Ford, R.; Gong, Y.C.; Fung, P.; Nahal, H.; Wang, P.W.; Desveaux, D.; Guttman, D.S. Quantitative interactor screening with next-generation sequencing (QIS-Seq) identifies Arabidopsis thaliana MLO2 as a target of the Pseudomonas syringae type III effector HopZ2. BMC Genom. 2012, 13, 8. [Google Scholar] [CrossRef] [PubMed]
- Nottensteiner, M.; Zechmann, B.; McCollum, C.; Hückelhoven, R. A barley powdery mildew fungus non-autonomous retrotransposon encodes a peptide that supports penetration success on barley. J. Exp. Bot. 2018. [Google Scholar] [CrossRef] [PubMed]
- Bos, J.I.B.; Armstrong, M.R.; Gilroy, E.M.; Boevink, P.C.; Hein, I.; Taylor, R.M.; Tian, J.D.; Engelhardt, S.; Vetukuri, R.R.; Harrower, B.; et al. Phytophthora infestans effector Avr3a is essential for virulence and manipulates plant immunity by stabilizing host E3 ligase CMPG1. Proc. Natl. Acad. Sci. USA 2010, 107, 9909–9914. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.D.; Boevink, P.; McLellan, H.; Armstrong, M.; Bukharova, T.; Qin, Z.W.; Birch, P.R. A host KH RNA-binding protein is a susceptibility factor targeted by an RXLR effector to promote late blight disease. Mol. Plant 2015, 8, 1385–1395. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; McLellan, H.; Naqvi, S.; He, Q.; Boevink, P.C.; Armstrong, M.; Giuliani, L.M.; Zhang, W.; Tian, Z.D.; Zhan, J.S.; et al. Potato NPH3/RPT2-like protein StNRL1, targeted by a Phytophthora infestans RXLR effector, is a susceptibility factor. Plant Physiol. 2016, 171, 645–657. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Murphy, F.; He, Q.; Armstrong, M.; Giuliani, L.M.; Boevink, P.C.; Zhang, W.; Tian, Z.; Birch, P.R.J.; Gilroy, E.M. The potato MAP3K StVIK is required for the Phytophthora infestans RXLR effector Pi17316 to promote disease. Plant Physiol. 2018. [Google Scholar] [CrossRef] [PubMed]
- Du, Y.; Overdijk, E.J.R.; Berg, J.A.; Govers, F.; Bouwmeester, K. Solanaceous exocyst subunits are involved in immunity to diverse plant pathogens. J. Exp. Bot. 2018, 69, 655–666. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Friesen, T.L.; Stukenbrock, E.H.; Liu, Z.H.; Meinhardt, S.; Ling, H.; Faris, J.D.; Rasmussen, J.B.; Solomon, P.S.; McDonald, B.A.; Oliver, R.P. Emergence of a new disease as a result of interspecific virulence gene transfer. Nat. Genet. 2006, 38, 953–956. [Google Scholar] [CrossRef] [PubMed]
- Manning, V.A.; Hardison, L.K.; Ciuffetti, L.M. Ptr ToxA interacts with a chloroplast-localised protein. Mol. Plant Microbe Interact. 2007, 20, 168–177. [Google Scholar] [CrossRef] [PubMed]
- Manning, V.A.; Chu, A.L.; Scofield, S.R.; Ciuffetti, L.M. Intracellular expression of a host-selective toxin, ToxA, in diverse plants phenocopies silencing of a ToxA-interacting protein, ToxABP1. New Phytol. 2010, 187, 1034–1047. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- See, P.T.; Marathamuthu, P.A.; Iagallo, E.M.; Oliver, R.P.; Moffat, C.S. Evaluating the importance of the tan spot ToxA-Tsn1 interaction in Australian wheat varieties. Plant Pathol. 2018. [Google Scholar] [CrossRef]
- Virdi, S.K.; Liu, Z.; Overlander, M.E.; Zhang, Z.; Xu, S.S.; Friesen, T.L.; Faris, J.D. New insights into the roles of host gene-necrotrophic effector interactions in governing susceptibility of durum wheat to tan spot and septoria nodorum blotch. G3 (Bethesda) 2016, 6, 4139–4150. [Google Scholar] [CrossRef] [PubMed]
- Stam, R.; McDonald, B.A. When resistance gene pyramids are not durable—The role of pathogen diversity. Mol. Plant Pathol. 2018, 19, 521–524. [Google Scholar] [CrossRef] [PubMed]
- Cook, A.A. A mutation for resistance to potato virus Y in pepper. Phytopathology 1961, 51, 550–552. [Google Scholar]
- Humphry, M.; Reinstädler, A.; Ivanov, S.; Bisseling, T.; Panstruga, R. Durable broad-spectrum powdery mildew resistance in pea er1 plants is conferred by natural loss-of-function mutations in PsMLO1. Mol. Plant Pathol. 2011, 12, 866–878. [Google Scholar] [CrossRef] [PubMed]
- Paulus, J.K.; van der Hoorn, R.A.L. Tricked or trapped—Two decoy mechanisms in host-pathogen interactions. PLoS Pathog. 2018, 14, e1006761. [Google Scholar] [CrossRef] [PubMed]
- Gauffier, C.; Lebaron, C.; Moretti, A.; Constant, C.; Moquet, F.; Bonnet, G.; Caranta, C.; Gallois, J.L. A TILLING approach to generate broad-spectrum resistance to potyviruses in tomato is hampered by eIF4E gene redundancy. Plant J. 2016, 85, 717–729. [Google Scholar] [CrossRef] [PubMed]
- Jarosch, B.; Jansen, M.; Schaffrath, U. Acquired resistance functions in mlo barley, which is hypersusceptible to Magnaporthe grisea. Mol. Plant Microbe Interact. 2003, 16, 107–114. [Google Scholar] [CrossRef] [PubMed]
- Kumar, J.; Hückelhoven, R.; Beckhove, U.; Nagarajan, S.; Kogel, K.H. A compromised Mlo pathway affects the response of barley to the necrotrophic fungus Bipolaris sorokiniana (Teleomorph: Cochliobolus sativus) and its toxins. Phytopathology 2001, 91, 127–133. [Google Scholar] [CrossRef] [PubMed]
- Ge, X.T.; Deng, W.W.; Lee, Z.Z.; Lopez-Ruiz, F.J.; Schweizer, P.; Ellwood, S.R. Tempered mlo broad-spectrum resistance to barley powdery mildew in an Ethiopian landrace. Sci. Rep. 2016, 7, 29558. [Google Scholar] [CrossRef] [PubMed]
- Fukuoka, S.; Saka, N.; Koga, H.; Ono, K.; Shimizu, T.; Ebana, K.; Hayashi, N.; Takahashi, A.; Hirochika, H.; Okuno, K.; et al. Loss of function of a proline-containing protein confers durable disease resistance in rice. Science 2009, 325, 998–1001. [Google Scholar] [CrossRef] [PubMed]
- De Guillen, K.; Ortiz-Vallejo, D.; Gracy, J.; Fournier, E.; Kroj, T.; Padilla, A. Structure analysis uncovers a highly diverse but structurally conserved effector family in phytopathogenic fungi. PLoS Pathog. 2015, 11, e1005228. [Google Scholar] [CrossRef] [PubMed]
- Maqbool, A.; Saitoh, H.; Franceschetti, M.; Stevenson, C.E.; Uemura, A.; Kanzaki, H.; Kamoun, S.; Terauchi, R.; Banfield, M.J. Structural basis of pathogen recognition by an integrated HMA domain in a plant NLR immune receptor. eLife 2015, 4, e08709. [Google Scholar] [CrossRef] [PubMed]
- Sarris, P.F.; Cevik, V.; Dagdas, G.; Jones, J.D.; Krasileva, K.V. Comparative analysis of plant immune receptor architectures uncovers host proteins likely targeted by pathogens. BMC Biol. 2016, 14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lebaron, C.; Rosado, A.; Sauvage, C.; Gauffier, C.; German-Retana, S.; Moury, B.; Gallois, J.L. A new eIF4E1 allele characterized by RNAseq data mining is associated with resistance to potato virus Y in tomato albeit with a low durability. J. Gen. Virol. 2016, 97, 3063–3072. [Google Scholar] [CrossRef] [PubMed]
- Yang, P.; Lüpken, T.; Habekuss, A.; Hensel, G.; Steuernagel, B.; Kilian, B.; Ariyadasa, R.; Himmelbach, A.; Kumlehn, J.; Scholz, U.; et al. PROTEIN DISULFIDE ISOMERASE LIKE 5-1 is a susceptibility factor to plant viruses. Proc. Natl. Acad. Sci. USA 2014, 111, 2104–2109. [Google Scholar] [CrossRef] [PubMed]
- Yang, P.; Habekuß, A.; Hofinger, B.J.; Kanyuka, K.; Kilian, B.; Graner, A.; Ordon, F.; Stein, N. Sequence diversification in recessive alleles of two host factor genes suggests adaptive selection for bymovirus resistance in cultivated barley from East Asia. Theor. Appl. Genet. 2017, 130, 331–344. [Google Scholar] [CrossRef] [PubMed]
- Acevedo-Garcia, J.; Spencer, D.; Thieron, H.; Reinstädler, A.; Hammond-Kosack, K.; Phillips, A.L.; Panstruga, R. mlo-based powdery mildew resistance in hexaploid bread wheat generated by a non-transgenic TILLING approach. Plant Biotechnol. J. 2017, 15, 367–378. [Google Scholar] [CrossRef] [PubMed]
- Nieto, C.; Piron, F.; Dalmais, M.; Marco, C.F.; Moriones, E.; Gómez-Guillamón, M.L.; Truniger, V.; Gómez, P.; Garcia-Mas, J.; Aranda, M.A.; et al. EcoTILLING for the identification of allelic variants of melon eIF4E, a factor that controls virus susceptibility. BMC Plant Biol. 2007, 7, 34. [Google Scholar] [CrossRef] [PubMed]
- Lusser, M.; Parisi, C.; Plan, D.; Rodríguez-Cerezo, E. Deployment of new biotechnologies in plant breeding. Nat. Biotechnol. 2012, 30, 231–239. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Palmgren, M.G.; Edenbrandt, A.K.; Vedel, S.E.; Andersen, M.M.; Landes, X.; Østerberg, J.T.; Falhof, J.; Olsen, L.I.; Christensen, S.B.; Sandøe, P.; et al. Are we ready for back-to-nature crop breeding? Trends Plant Sci. 2015, 20, 155–164. [Google Scholar] [CrossRef] [PubMed]
- Bastet, A.; Robaglia, C.; Gallois, J.L. eIF4E resistance: Natural variation should guide gene editing. Trends Plant Sci. 2017, 22, 411–419. [Google Scholar] [CrossRef] [PubMed]
- Peng, A.H.; Chen, S.C.; Lei, T.G.; Xu, L.Z; He, Y.R.; Wu, L.; Yao, L.X.; Zou, X.P. Engineering canker-resistant plants through CRISPR/Cas9-targeted editing of the susceptibility gene CsLOB1 promoter in rice. Plant Biotechnol. J. 2017, 15, 1509–1519. [Google Scholar] [CrossRef] [PubMed]
- Jia, H.G.; Zhang, Y.Z.; Orbović, V.; Xu, J.; White, F.F.; Jones, J.B.; Wang, N. Genome editing of the disease susceptibility gene CsLOB1 in citrus confers resistance to citrus canker. Plant Biotechnol. J. 2017, 15, 817–823. [Google Scholar] [CrossRef] [PubMed]
- Macovei, A.; Sevilla, N.R.; Cantos, C.; Jonson, G.B.; Slamet-Loedin, I.; Čermák, T.; Voytas, D.F.; Choi, I.R.; Chadha-Mohanty, P. Novel alleles of rice eIF4G generated by CRISPR/Cas9-targeted mutagenesis confer resistance to Rice tungro spherical virus. Plant Biotechnol. J. 2018. [Google Scholar] [CrossRef] [PubMed]
- Nekrasov, V.; Wang, C.; Win, J.; Lanz, C.; Weigel, D.; Kamoun, S. Rapid generation of a transgen-free powdery mildew resistant tomato by genome deletion. Sci. Rep. 2017, 7. [Google Scholar] [CrossRef] [PubMed]
- Blanvillain-Baufumé, S.; Reschke, M.; Solé, M.; Auguy, F.; Doucoure, H.; Szurek, B.; Meynard, D.; Portefaix, M.; Cunnac, S.; Guiderdoni, E.; et al. Targeted promoter editing for rice resistance to Xanthomonas oryzae pv. oryzae reveals differential activities for SWEET14-inducing TAL effectors. Plant Biotechnol. J. 2017, 15, 306–317. [Google Scholar]
- Gu, K.Y.; Yang, B.; Tian, D.S.; Wu, L.F.; Wang, D.J.; Sreekala, C.; Yang, F.; Chu, Z.Q.; Wang, G.L.; White, F.F.; et al. R gene expression induced by a type-III effector triggers disease resistance in rice. Nature 2005, 435, 1122–1125. [Google Scholar] [CrossRef] [PubMed]
- Römer, P.; Strauss, T.; Hahn, S.; Scholze, H.; Morbitzer, R.; Grau, J.; Bonas, U.; Lahaye, T. Recognition of AvrBs3-like proteins is mediated by specific binding to promoters of matching pepper Bs3 alleles. Plant Physiol. 2009, 150, 1697–1712. [Google Scholar] [CrossRef] [PubMed]
- Römer, P.; Strauss, T.; Hahn, S.; Scholze, H.; Morbitzer, R.; Grau, J.; Bonas, U.; Lahaye, T. The rice TAL effector-dependent resistance protein XA10 triggers cell death and calcium depletion in the endoplasmic reticulum. Plant Cell 2014, 26, 497–515. [Google Scholar]
- Wang, C.L.; Zhang, X.P.; Fan, Y.L.; Gao, Y.; Zhu, Q.L.; Zheng, C.K.; Qin, T.F.; Li, Y.Q.; Che, J.Y.; Zhang, M.W.; et al. Xa23 is an executor R protein and confers broad-spectrum disease resistance in rice. Mol. Plant 2015, 8, 290–302. [Google Scholar] [CrossRef] [PubMed]
- Kourelis, J.; van der Hoorn, R.A.L. Defended to the Nines: 25 years of resistance gene cloning identifies nine mechanisms for R protein function. Plant Cell 2018, 30, 285–299. [Google Scholar] [CrossRef] [PubMed]
- Boni, R.; Chauhan, H.; Hensel, G.; Roulin, A.; Sucher, J.; Kumlehn, J.; Brunner, S.; Krattinger, S.G.; Keller, B. Pathogen-inducible Ta-Lr34res expression in heterologues barley confers disease resistance without negative pleiotropic effects. Plant Biotechnol. J. 2018, 16, 245–253. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.H.; Qi, D.; Ashfield, T.; Helm, M.; Innes, R.W. Using decoys to expand the recognition specificity of a plant disease resistance protein. Science 2016, 351, 684–687. [Google Scholar] [CrossRef] [PubMed]
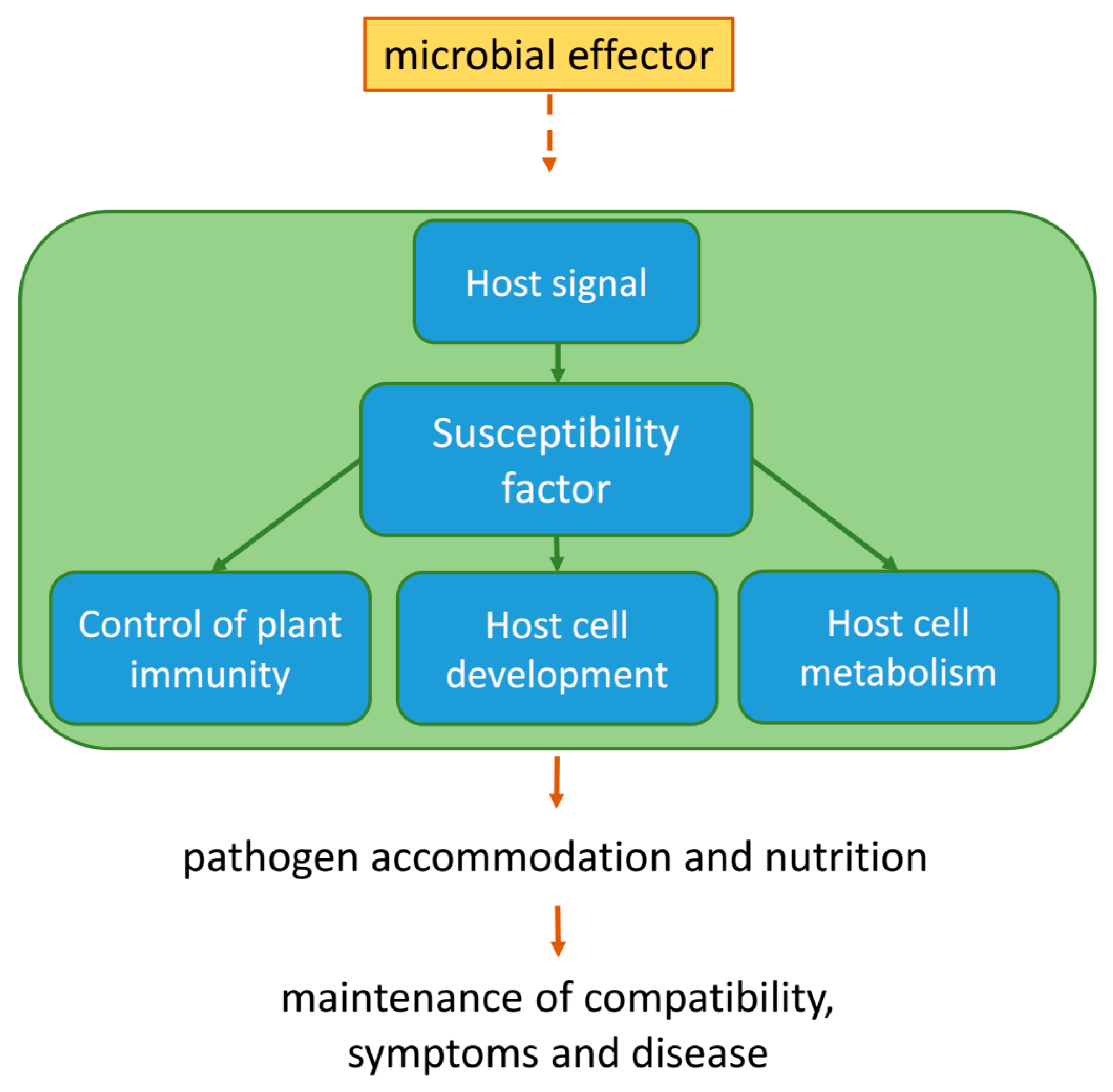
Figure 1.
Plant physiological functions and microbial utilization of host susceptibility factors (S). Host S factors can have diverse physiological functions (highlighted by the green background box), which would also operate in pathogen-free plants. This can include control of immune responses or normal host cell development and metabolism. In healthy plants, S factors are regulated by host endogenous signals (e.g., hormones, peptides, second messengers, protein-protein interaction, and protein modification). Microbial pathogens can simply profit from S factor functions or they actively take advantage of S factors via virulence effectors. Effectors might directly act on S factors or on their physiological environment.
Figure 1.
Plant physiological functions and microbial utilization of host susceptibility factors (S). Host S factors can have diverse physiological functions (highlighted by the green background box), which would also operate in pathogen-free plants. This can include control of immune responses or normal host cell development and metabolism. In healthy plants, S factors are regulated by host endogenous signals (e.g., hormones, peptides, second messengers, protein-protein interaction, and protein modification). Microbial pathogens can simply profit from S factor functions or they actively take advantage of S factors via virulence effectors. Effectors might directly act on S factors or on their physiological environment.

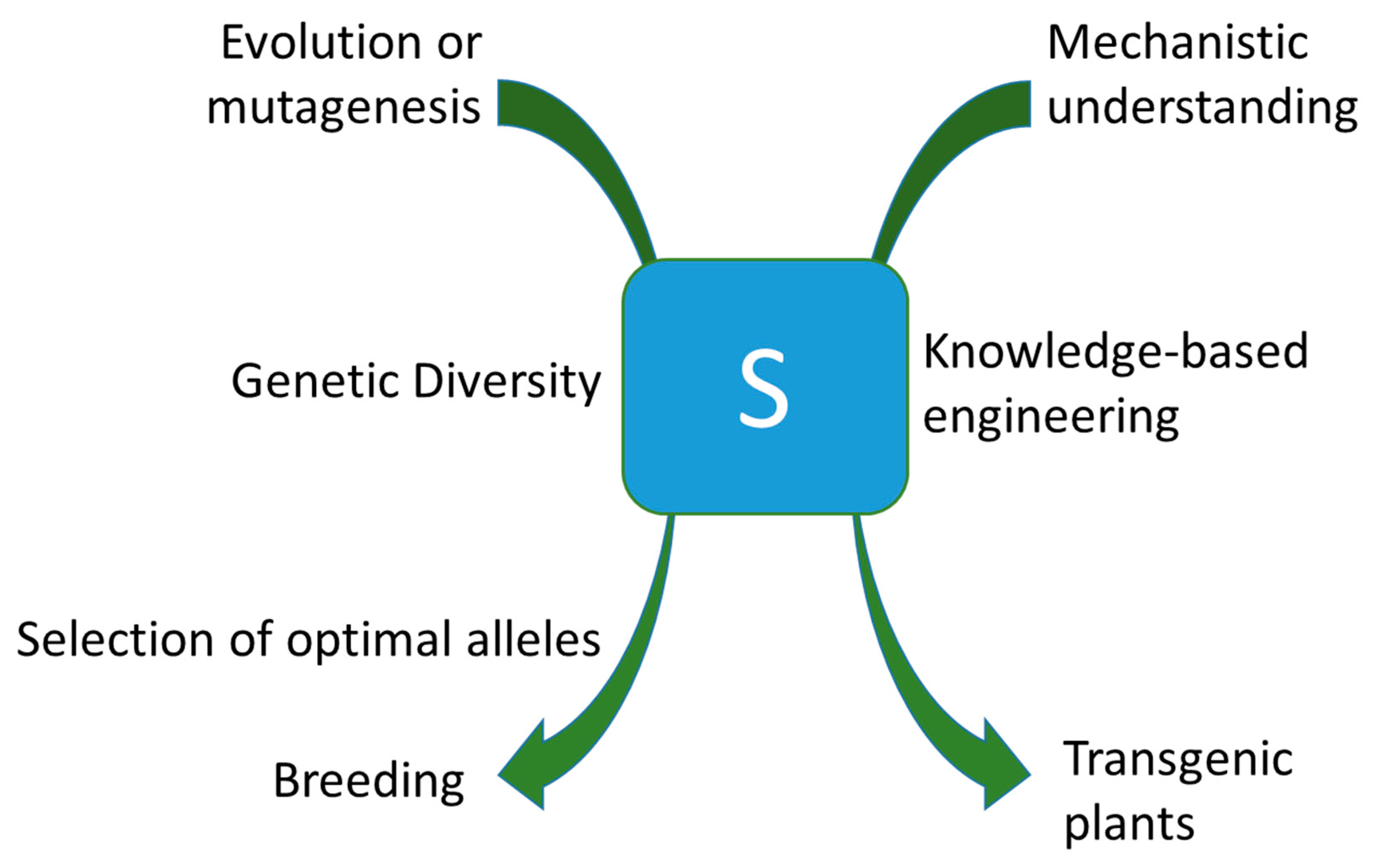
Figure 2.
Plant susceptibility genes can be exploited for loss of susceptibility in breeding or biotechnology. Optimal mutant alleles (left side of the panel) that provide a balance between gain of resistance and potential trade-offs can be identified in natural plant populations by ECOTILLING or after random or targeted mutagenesis by Targeting Induced Local Lesions In Genomes (TILLING) or DNA-endonuclease mediated mutagenesis, respectively. Alternatively, knowledge-based (right side of the panel) approaches are enabled if a mechanistic understanding of S gene functions exists. This might, amongst others, include artificial effector decoys or silencing S genes on demand under control of pathogen-triggered promoters.
Figure 2.
Plant susceptibility genes can be exploited for loss of susceptibility in breeding or biotechnology. Optimal mutant alleles (left side of the panel) that provide a balance between gain of resistance and potential trade-offs can be identified in natural plant populations by ECOTILLING or after random or targeted mutagenesis by Targeting Induced Local Lesions In Genomes (TILLING) or DNA-endonuclease mediated mutagenesis, respectively. Alternatively, knowledge-based (right side of the panel) approaches are enabled if a mechanistic understanding of S gene functions exists. This might, amongst others, include artificial effector decoys or silencing S genes on demand under control of pathogen-triggered promoters.

© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Engelhardt, S.; Stam, R.; Hückelhoven, R. Good Riddance? Breaking Disease Susceptibility in the Era of New Breeding Technologies. Agronomy 2018, 8, 114. https://doi.org/10.3390/agronomy8070114
AMA Style
Engelhardt S, Stam R, Hückelhoven R. Good Riddance? Breaking Disease Susceptibility in the Era of New Breeding Technologies. Agronomy. 2018; 8(7):114. https://doi.org/10.3390/agronomy8070114
Chicago/Turabian StyleEngelhardt, Stefan, Remco Stam, and Ralph Hückelhoven. 2018. "Good Riddance? Breaking Disease Susceptibility in the Era of New Breeding Technologies" Agronomy 8, no. 7: 114. https://doi.org/10.3390/agronomy8070114
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.