1. Introduction
The characterisation of biomarkers and bio-indicators that reflect the physiological health status of plants and their productivity is a major focus for agricultural development (i.e., breeding tolerant genotypes). Renewed focus on plant chemistry recognises the variation and complexity of plant physio-chemical responses to the dynamic environment, an aspect emphasised under field conditions. Despite this, few chemical biomarkers exist for the functional characterisation of plant stress responses. Notable examples are that of polyols as indicators of abiotic stress [
1] and volatile compounds as indicators of plant disease [
2].
The partitioning of carbon among compound classes has recently received renewed attention as an indicator of plant growth and productivity [
1]. These findings build upon a large volume of work amassed over recent decades investigating individual chemical entities—often termed ‘stress metabolites’ in response to single stress elicitors [
3]. Alternatively, the emerging discipline of ‘metabolomics’ offers unique insight into the response of the whole plant chemical network, yielding a more comprehensive understanding of its response to changing conditions [
4,
5,
6]. However, such investigations remain costly and are often conducted under controlled conditions [
7]. Few investigations adopt the strategic approach of focusing on subsets of plant metabolism that reflect important branch-points in the biochemical network.
Evidence suggests that sugar alcohols impart an array of osmolytic and osmo-protective functions to mitigate the effects of stressful conditions [
8,
9]. Therefore, the allocation of carbon to the synthesis of sugar alcohols is commonly termed an accumulation of stress metabolites. Despite these functions, very few papers [
10] have sought to characterise the accumulation of this compound class under field conditions. The elicitation of sugar alcohol biosynthesis across a range of taxa has been demonstrated under controlled conditions, most commonly as a result of the combined effects of water, heat, or light stress, [
11]. The dynamics of this elicitation under field conditions is limited to correlative relationships with environmental gradients [
10], such as mean annual rainfall, with little regard for concomitant physiological responses or changes in plant chemistry attributable to developmental processes.
The dynamic of plant water use is a crucial characteristic that supports both plant productivity and survival. The characterisation of water use efficiency (WUE) is a common bio-indicator for both crop management and plant improvement programs. The use of carbon isotopes to predict WUE is an established approach [
12,
13] that is now transferring to industrial applications [
14]. Improving the relationship between stochastic collection of gas exchange parameters and time integrated WUE modelled from isotope fractionation models has been achieved via a range of considerations, such as assimilation weighting gas exchange measures [
15] and a broader consideration of fractionation events both pre- and post-carboxylation [
15,
16,
17]. It is well-established that post carboxylation, biochemical processes of fractionation and partitioning lead to the unequal distribution of carbon isotopes throughout the biochemical network [
18]. Predicting WUE on the basis of carbon isotope composition (expressed as δ
13C) is therefore susceptible to divergence with actual WUE attributable to post-photosynthetic fractionation and the uneven distribution of isotopes among compound classes. It has been previously shown that the δ
13C of sugar alcohols can differ substantially from more commonly abundant metabolites (such as sucrose and glucose) in
Phaseolus vulgaris [
19,
20]. It is not clear if these patterns are ubiquitous among legumes and whether the scope and magnitude of these patterns exhibit a significant influence over predictions of WUE.
Here, a set of commercially available chickpea (Cicer arietinum L.) genotypes were used to characterise the accumulation of carbohydrates and sugar alcohols in leaf tissues in field under two levels of water availability and at three different developmental stages. Changes in concentrations of carbohydrates and sugar alcohols as well as their carbon isotope composition at the compound specific level was determined in order to quantify their subsequent influence over predictions of WUE. Specifically, the following objectives were addressed: (1) to observe changes in carbon allocation to different compounds at the leaf level under field conditions; and (2) determine how these changes differ with different chickpea genotype, irrigation treatment, and developmental stage. It was hypothesized that the δ13C of D-pinitol will differ from the δ13C of other metabolites sampled from the soluble leaf fraction, that the δ13C of D-pinitol will increase in response to lower water availability—similar to the other metabolites from the soluble fraction—and that the magnitude of changes in concentration will influence predictions of WUE.
2. Materials and Methods
2.1. Experimental Design
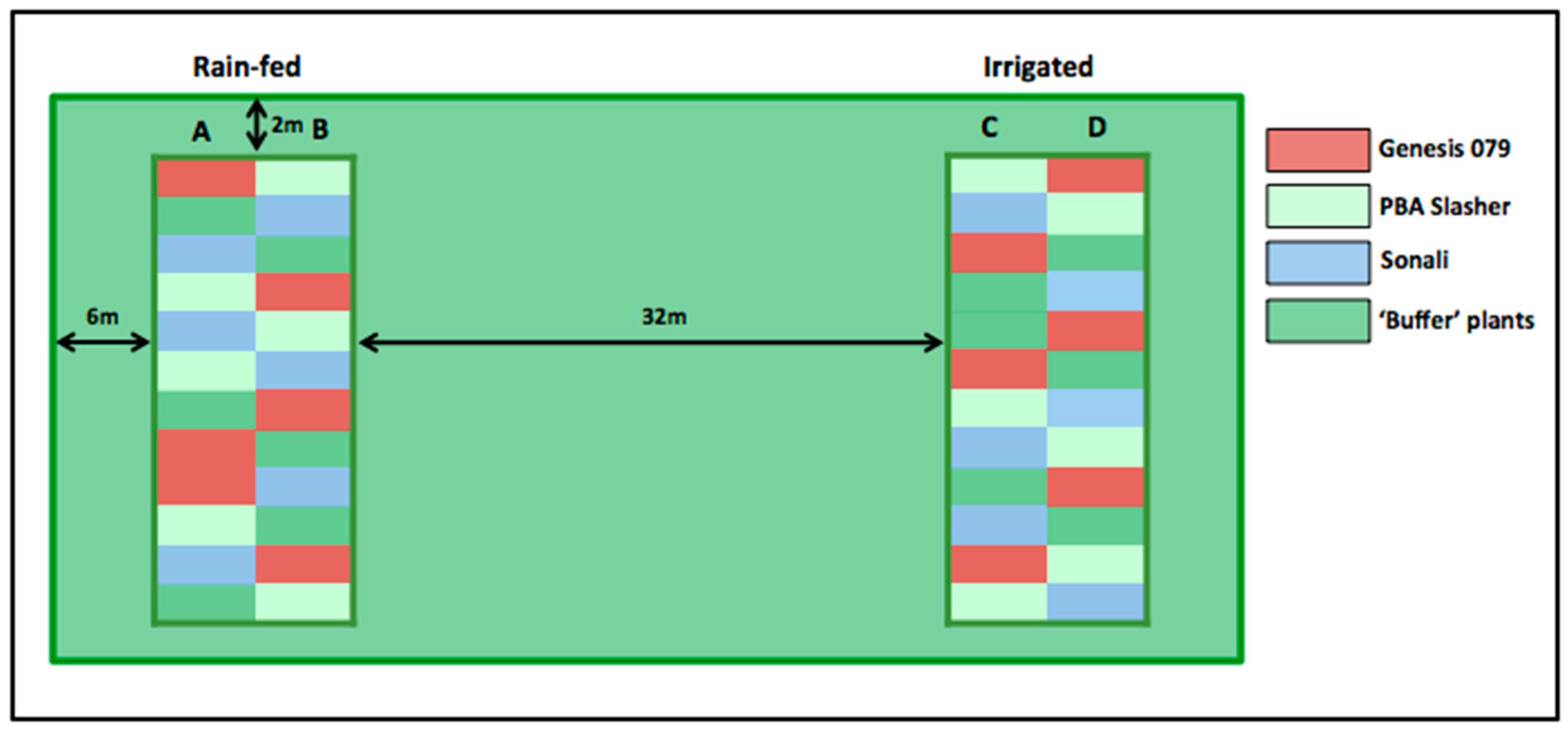
Three chickpea genotypes Genesis 079, PBA Slasher, and Sonali were grown in the field at the I.A. Watson Wheat Research Centre in Narrabri, New South Wales, Australia (30°16′27.0″ S 149°48′33.5″ E) on Black Vertosol in 2015 (11 June to 11 November). All three chickpea genotypes had similar phenology and reached 50% flowering within 15 days and first pod development with one day difference between genotypes.
Each genotype was grown on 6 × 2 m plots replicated six times in a randomized block design within two treatments (rain-fed and irrigated). Buffer chickpea plots were planted between treatments as well as around the outer perimeter of the trial (
Figure 1). Total monthly precipitation recorded at the I.A. Watson Wheat Research Centre over the course of the field trial was as follows: 41.16 mm in from 11–30 June, 33.88 mm for the month of July, 42.56 mm for the month of August, 22.12 mm for the month of September, 17.64 mm for the month of October, and 82.6 mm from 1–11 November. Irrigation occurred on 25 September 2015 and 15 October 2015 with 36 mm and 26 mm applied respectively. The herbicides Terbyne 750 wg (1 kg ha
−1) and Balance 750 wg (100 g ha
−1) were applied on 13 June 2015. The fungicides Unite 27 (0.5 L ha
−1) and Ridomil (2.5 kg ha
−1) were applied on 21 August 2015 and 22 August 2015 respectively and the insecticide Krate Zion (0.036 L ha
−1) was applied on 4 October 2015.
Plants were sampled systematically throughout reproductive stages of development. Three campaign style measurements, each of two days duration, were made and coincided with major developmental stages in reproduction: (1) flowering, (2) pod development, and (3) pod fill. Each stage was defined as:
Flowering: All four genotypes contain flowers. Flowers are in full bloom, but with no visible signs of developing pods (sampling took place 90 and 91 days after planting)
Pod Development: All genotypes still contain flowers, but most blooms have been replaced with developing pods (sampling took place 102 and 103 days after planting)
Pod Fill: All genotypes have pods that have become larger and more round due to seed growth and filling (sampling took place 118 and 119 days after planting)
2.2. Physiological Measurements
Leaf gas exchange of the first fully developed leaf was determined using the WALZ GFS-3000 portable infra-red gas analyzer (Walz Heinz GmbH, Effeltrich, Germany), commencing at 08:30. The measurement included net photosynthetic rate (A), stomatal conductance to H
2O (g
s) and the ratio of intercellular to atmospheric CO
2 concentration (c
i/c
a). Conditions within the gas exchange chamber were set to track ambient PAR and temperature. Average daily parameters such as temperature and relative humidity for each sampling campaign are depicted in
Table 1. All six replicate plots per treatment were measured within two days (three replicates each day).
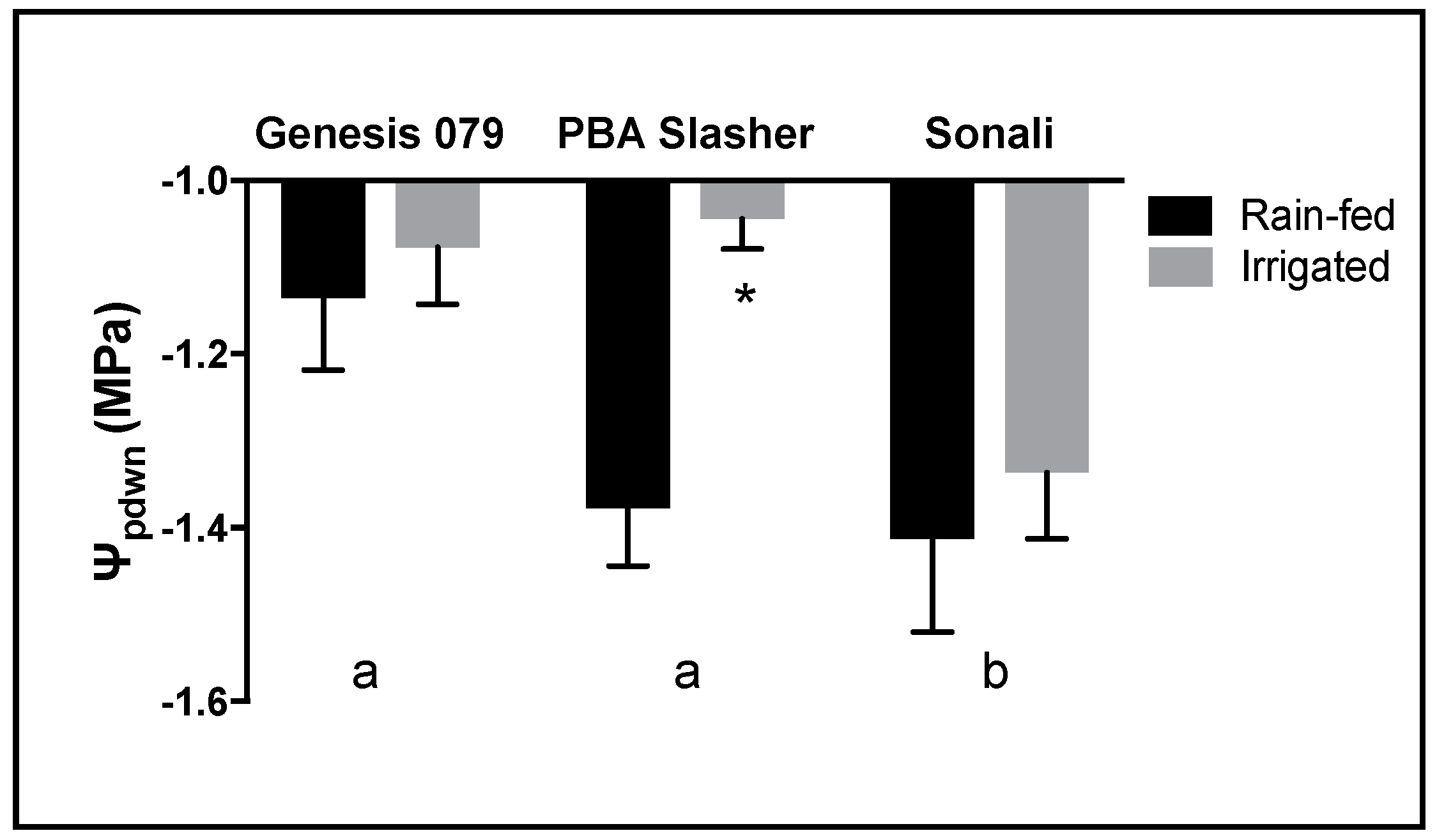
Predawn leaf water potentials were measured at pod fill (Model 3000F01, Soilmoisture Equipment Corp, Santa Barbara, CT, USA) from 04:00 to 06:00. Leaves adjacent to those used for gas exchange measurements were collected for metabolomic analysis at midday between the hours of 12:30 and 14:00 and immediately stored in a portable freezer at −10 °C, awaiting sample microwave treatment according to [
21]. Samples were then dried in an oven at 70 °C overnight and stored until analysis took place.
2.3. Analysis of Metabolite Concentrations
Dried leaf samples were ground and hot water extractions were performed according to [
22]. In order to analyse non-polar analytes using GC-QQQ, samples were derivatized according to [
23]. The separation and quantification of target metabolites was completed using an Agilent 6890A gas chromatograph with QQQ 7000 mass selective detector in scan mode using 50–500 AMU (70 eV) (Agilent Technologies, Santa Clara, CA, USA). Quantification was achieved using MassHunter
TM software (Agilent Technologies, Santa Clara, CA, USA) according to the protocol detailed in [
23].
2.4. Compound Specific Isotope Analysis
Compound specific isotope analysis was completed according to [
24] and took place at the Terrestrial Ecosystem Research laboratory at the University of Vienna in Austria. Dried and weighed leaf samples were transported to the University of Vienna where hot water extractions were first performed according to [
22] and then passed over ion exchange resins in order to remove ionic components. Metabolite standards with δ
13C values ranging from −27‰ to −10‰ and referenced as pure chemicals by EA/IRMS (EA 1110, CE Instruments, Milan, Italy coupled to a Finnigan MAT Delta Plus IRMS, Thermo Fisher Scientific, Waltham, MA, USA) were prepared at different concentrations and analysed alongside samples. To correct the offset that occurred to the δ
13C of samples, the following multiple regression equation [
24] had to be applied
δ13Coffline and δ13Conline represent the δ13C values of the standards measured by EA/IRMS and HPLC/IRMS respectively. δ13C of every sample were corrected by plugging a, b, and c (calculated from standards), peak area and δ13Conline into the equation.
2.5. Modelling Carbon Isotope Discimination and Water Use Efficiency
Photosynthetic carbon isotope discrimination (Δ) for individual metabolites was calculated using the equation
The δ
13C
metabolite represents measured carbon isotope composition of metabolites from leaf samples and δ
13C
air represents the carbon isotope composition of atmospheric CO
2 (−8‰). Modelled intrinsic water use efficiency was calculated according to [
12] originally formulated by [
25]
where
a is fractionation caused by gaseous diffusion through the stomata (4.4‰), and
b’ is the effective fractionation caused by carboxylating enzymes RuBisCO and PEP carboxylase (approximately 27‰). The factor 1.6 originates from the ratio of the diffusion rate of H
2O to CO
2. Weighted water use efficiency, or modelled intrinsic water use efficiency incorporating all metabolite concentrations and indicating water use efficiency for the total soluble fraction of the leaf samples collected was also calculated.
2.6. Statistical Analysis
A restricted maximum likelihood (REML) analysis was run on GenStat 15th Edition (VSN International, Hemel Hempstead, UK) to determine if a significant temporal, treatment and genotypic effect on leaf water potential, metabolite concentration, δ13C, and water use efficiency. A least significant difference (LSD) test was then done to determine similar groups and see where significant temporal, treatment, and genotypic differences had occurred. A linear mixed effect model was used for statistical analysis of gas exchange, including the irrigation treatment and genotypes as fixed effects and the day of measurement and repeated readings per leaf as random effects using the nlme package in R (version 3.4.3). To check for differences between days of measurement of gas exchange, the irrigation treatment and genotype were defined as random effects of the linear mixed effects model. A significance level of 0.05 was set for all statistical tests.
4. Discussion
The accumulation of soluble metabolites in plant tissues has been observed across many genera in response to stressful conditions. Many of these metabolites belong to a set of compound classes commonly referred to as stress metabolites [
3,
8]. Here, we have not observed an accumulation of sugars,
myo-inositol, or D-pinitol in response to the applied irrigation treatment under field conditions. This indicates that the applied irrigation treatment introduced a very mild drought treatment and that variation in these characteristics under field conditions is lower than that suggested by previous glasshouse based experiments [see for example 1]. This study identified D-pinitol accumulation in chickpea to have implications for the use of soluble metabolite δ
13C values in the prediction of WUE. This trial also lays a strong foundation for future studies to further investigate the use of D-pinitol as an indicator of stress tolerance.
4.1. Physiological Responses
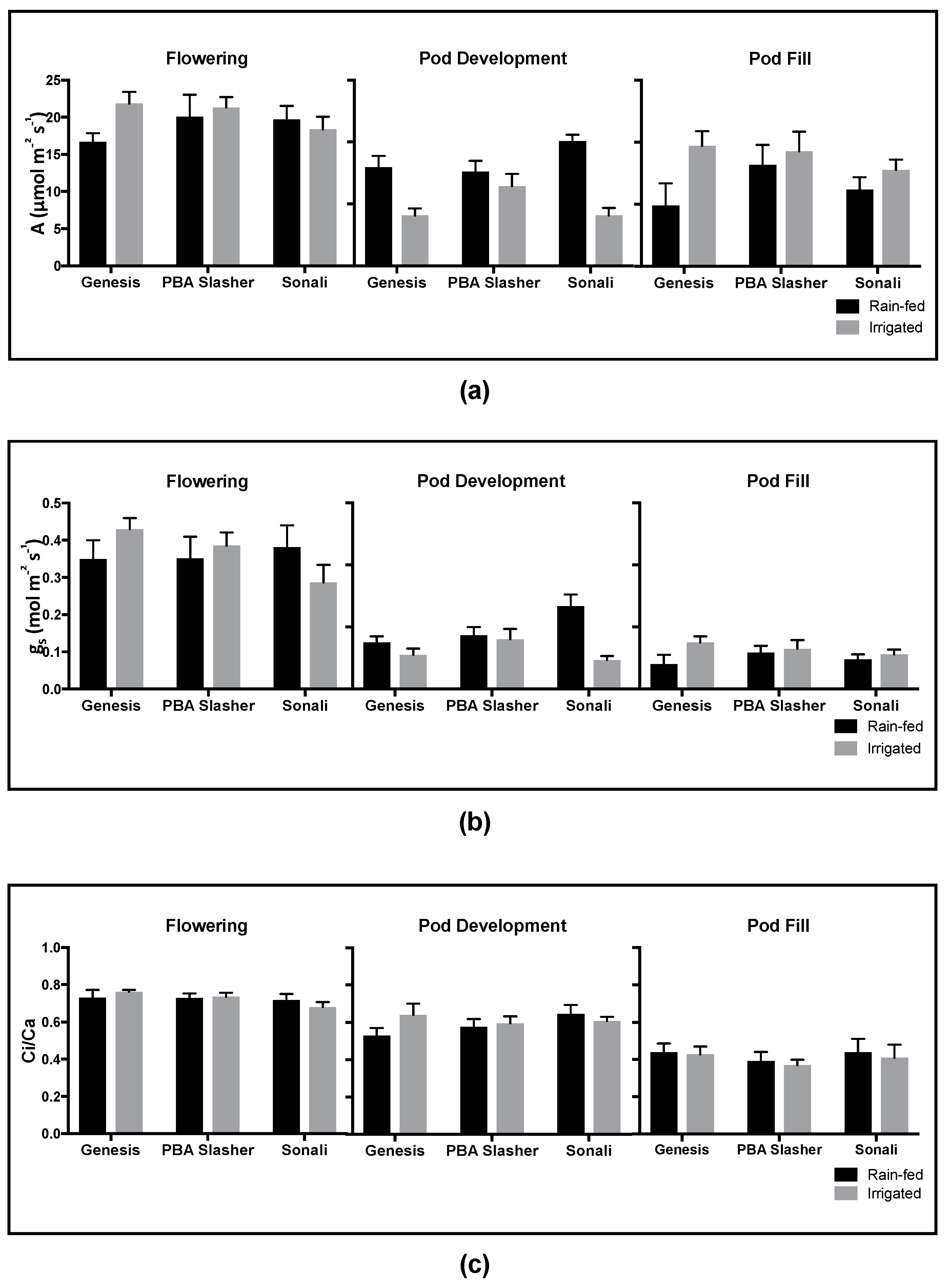
Stress responses within the plant system are complex and highly variable, responding to a range of adverse environmental conditions with changes to physiology, metabolism, and gene expression. One of the earliest strategies employed by plants is through changes in physiology, particularly that of changes in stomatal aperture (i.e.
, closure). Stomatal closure is the first line of defence against dehydration in response to a range of environmental conditions [
26]. At no time point was the expected stomatal closure for plants on the rain-fed plots observed. Instead, g
s responded to day to day and diurnal variation in field conditions (i.e.
, heat, light intensity).
Physiological measurements may therefore not be as accurate or stable when screening plants in natural conditions. For the present study, this day to day variation was observed in physiological measurements collected during pod development. Significant differences in gas exchange values between sampling days were observed for A and gs and were likely caused by daily variations in temperature or relative humidity and do not accurately represent the overall plant performance. For this reason, it is critical to take a more integrative approach (such as the using carbon isotope abundances) when assessing plant health and performance in field in order to gain a more accurate understanding of plant health status.
Small and largely non-significant changes in gas exchange (A, g
s, c
i/c
a) and Ψ
pdwn between treatments likely reflect the subtlety of the water deficit treatment produced in this study. Nevertheless, the irrigation treatment increased yield significantly compared to the rain-fed (data not shown). As a result of field conditions such as larger soil volumes, good water holding capacity of the Black Vertosol, conservative soil water use, and (perhaps) lateral flow of water may have contributed to reducing the severity of the water deficit exhibited in this study. Nevertheless, this phenomenon is a consequence of realistic field conditions and serves as a suitable context for the biochemical patterns observed. In addition, specifically for gas exchange parameters, the lack of significant differences in rates of A, g
s, and c
i/c
a may have been due to adaptive characteristics (such as modification of water potential [
27]) to the slow onset of water deficit experienced as a consequence of working under field conditions. Biochemical patterns that were observed are therefore informative of what is likely to be experienced in broader, field based screening efforts.
4.2. Biochemical Responses
Sampling of metabolites under field conditions enables the measurement of plant processes in a realistic cropping context. Both magnitude and variability of light, temperature, humidity, and soil quality can influence biochemical processes in both the short and long term [
28]. The interpretation of the data must therefore consider the cascade of processes resulting from a higher level of complexity in growth conditions.
Sugar alcohols have been shown to follow taxonomical patterns [
29,
30] and accumulate in response to environmental stress [
9,
11,
31,
32,
33], making them excellent candidates for use in breeding programs and assessing plant health in field [
1]. Despite substantial evidence documenting sugar alcohol accumulation under field conditions is limited to only a handful of studies [
10]. In this field study, D-pinitol and
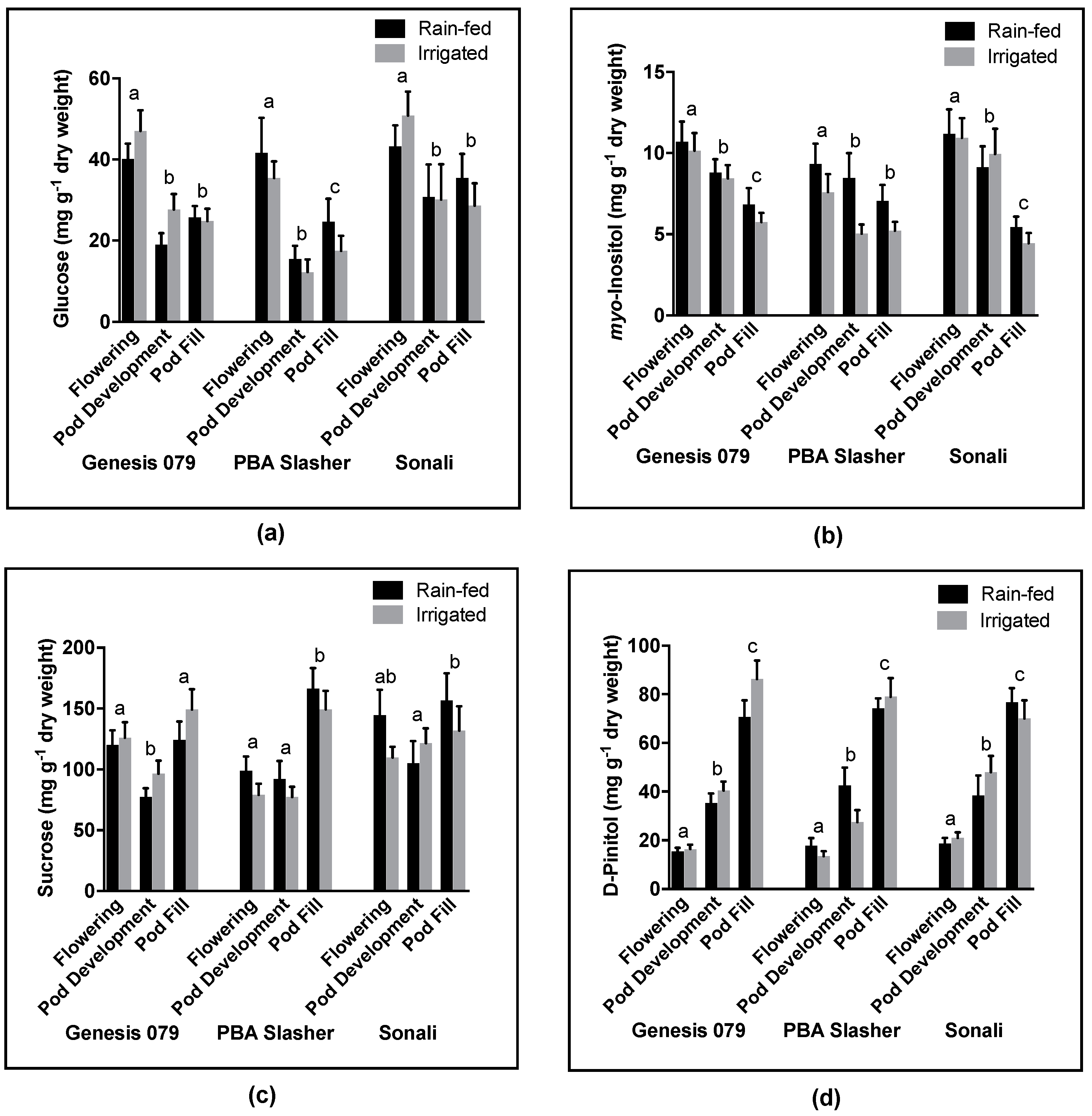
myo-inositol concentrations showed consistent changes in concentrations over the three development stages with D-pinitol increasing and
myo-inositol (one of D-pinitol’s precursors) decreasing upon each subsequent sampling trip (
Figure 4). This suggests that the accumulation of D-pinitol is strongly influenced by developmental processes (such as desiccation during seed maturity). Interestingly, despite this observed inverse relationship in the accumulation patterns of
myo-inositol and D-pinitol, concentrations were positively correlated on a plant by plant basis (
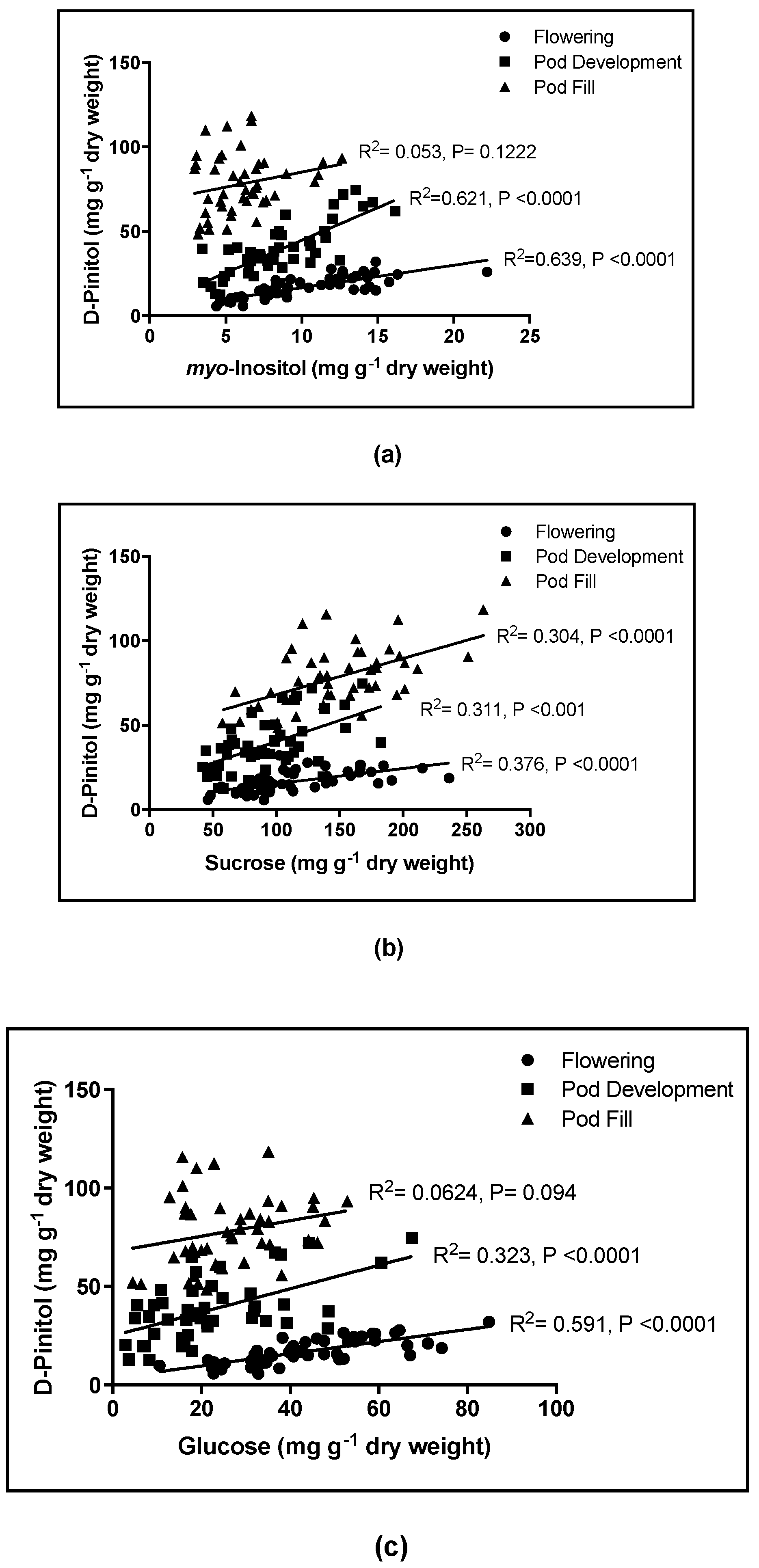
Figure 5a), suggesting plants that produce higher concentrations of
myo-inositol will accumulate higher concentrations of D-pinitol. However, this relation became weaker with plant development, possibly due to metabolites being allocated from the leaf and into the developing pod. Further studies should aim to elucidate the mechanisms responsible for these observed patterns in metabolite accumulation, focusing on the influence of plant development as well as water availability.
In a rare example of field based investigation of sugar alcohol accumulation, Streeter et al. [
10] examined the D-pinitol accumulation among soybean lines planted across China, noting three-fold changes in concentration between genotypes growing in areas of high rainfall and areas of low rainfall. In the present study, no difference in D-pinitol levels was observed between soil moisture treatments, suggesting that under mild water stress other field conditions (i.e., sunlight, wind conditions, temperature) might have a stronger influence on D-pinitol levels. D-Pinitol accumulation showed distinct patterns across time, with relatively little variation among lines. This suggests that the accumulation of D-pinitol is developmentally and environmentally regulated, although a greater variety of lines must be sampled to elucidate how strongly accumulation is determined on a genetic basis.
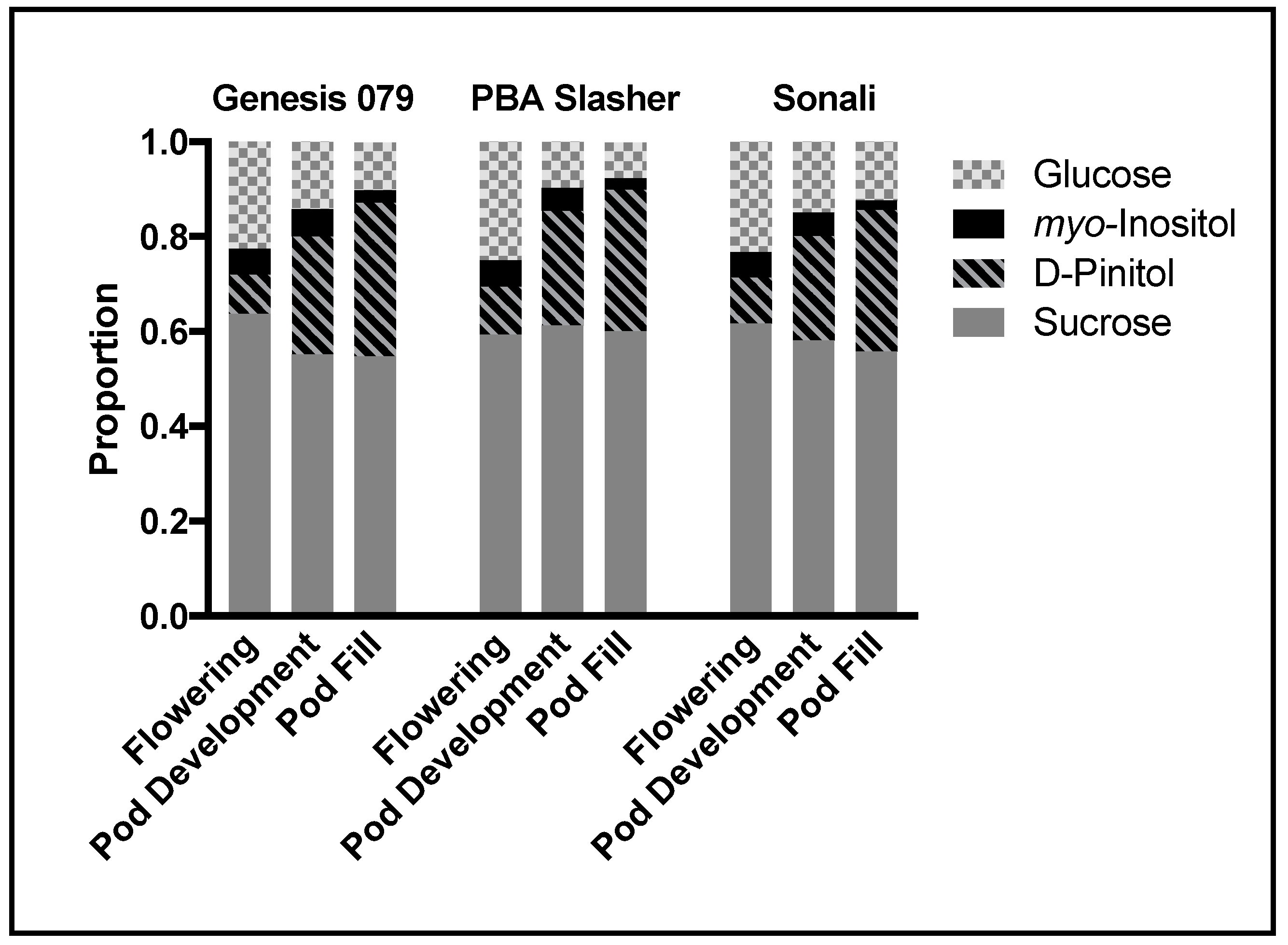
In the present study, D-pinitol was found to constitute a major component of the plant soluble fraction that was measured, being second only to sucrose at the pod fill stage (
Figure 6). Allocating carbon to the D-pinitol pool and away from metabolites such as starch and sucrose may have influences over plant growth. Substantial evidence now exists for the role(s) of sugar alcohols as stress metabolites, attributable to their their physiochemical properties as stress metabolites [
3]. The accumulation of sugar alcohols may also acts as a carbon sink that is removed from ‘primary metabolism, thereby mitigating the effects of sugar-mediated repression of photosynthetic reactions under stress conditions [
1]. By shuttling carbon into the polyol pool, plants are able to remove carbon from the carbohydrate pool, diminishing unwanted feedbacks on primary photosynthesis [
34]. Avoiding down regulation of photosynthesis is thought to be an important strategy used by plants to tolerate resource availability and influence rates of primary metabolism upstream [
35]. Our study suggests that limited variation among genotypes may impede breeding efforts to improve the stress tolerance of chickpea lines attributable to D-pinitol accumulation, although scanning a wider range of genotypes under field conditions is necessary.
4.3. Implications for Predicting WUE Based on Compound Specific Isotope Analysis
Stable carbon isotope abundance is an established proxy for predicting WUE within the plant system [
25]. The carbon isotope composition of plant tissues is influenced by a well-studied set of fractionation events governing the diffusion and incorporation of carbon into primary photosynthates [
12]. Less studied are the processes of post-carboxylation fractionation [
36]. Changes in the ratio of intercellular to ambient CO
2 concentrations are reflected in ∆
13C due to the strong impact of diffusional fractionation into the stomatal cavity. Thus, environmental conditions, especially soil water availability, have a strong impact on ∆
13C, which can be used to monitor temporal and spatial trends in carbon–water relations [
12]. As there was no irrigation treatment effect on c
i/c
a, it is expected that the carbon isotope abundance in the measured metabolites was not influenced by the irrigation treatment either. Post-carboxylation processes imparting fractionation of carbon isotopes (often termed heterotrophic fractionation) give rise to variation in ∆
13C among plant organs and compounds within metabolite networks [
37]. As these processes are less understood, heterotrophic fractionation is considered problematic in predicting WUE based on isotope ratios obtained from carbon pools that are considered ‘remote’ to the primary photosynthetic reactions.
In addition to challenges in predicting WUE due to heterotrophic fractionation, the temporal integration of WUE by isotope abundance is reliant on the source tissue. Sampling for ∆
13C is often constrained to aboveground plant tissue, particularly to leaves. Bulk leaf carbon isotope composition [
25] provides a time integrated measure for the growing period of the leaf material. In contrast, when only the carbon isotope composition of total soluble compounds [
38] is analysed, the integrated signal reflects a shorter time frame over which the pool of carbon was fixed into plant tissues.
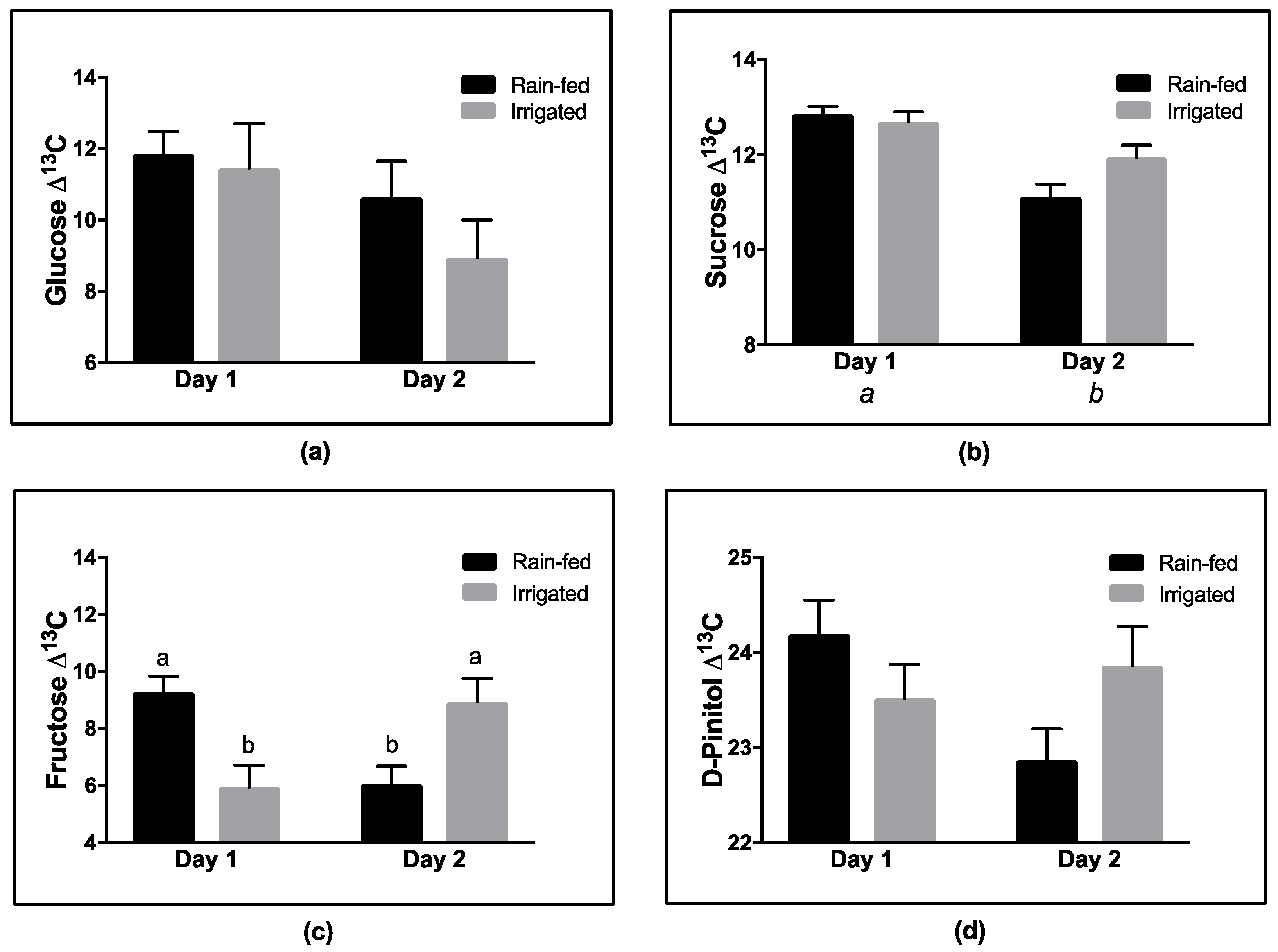
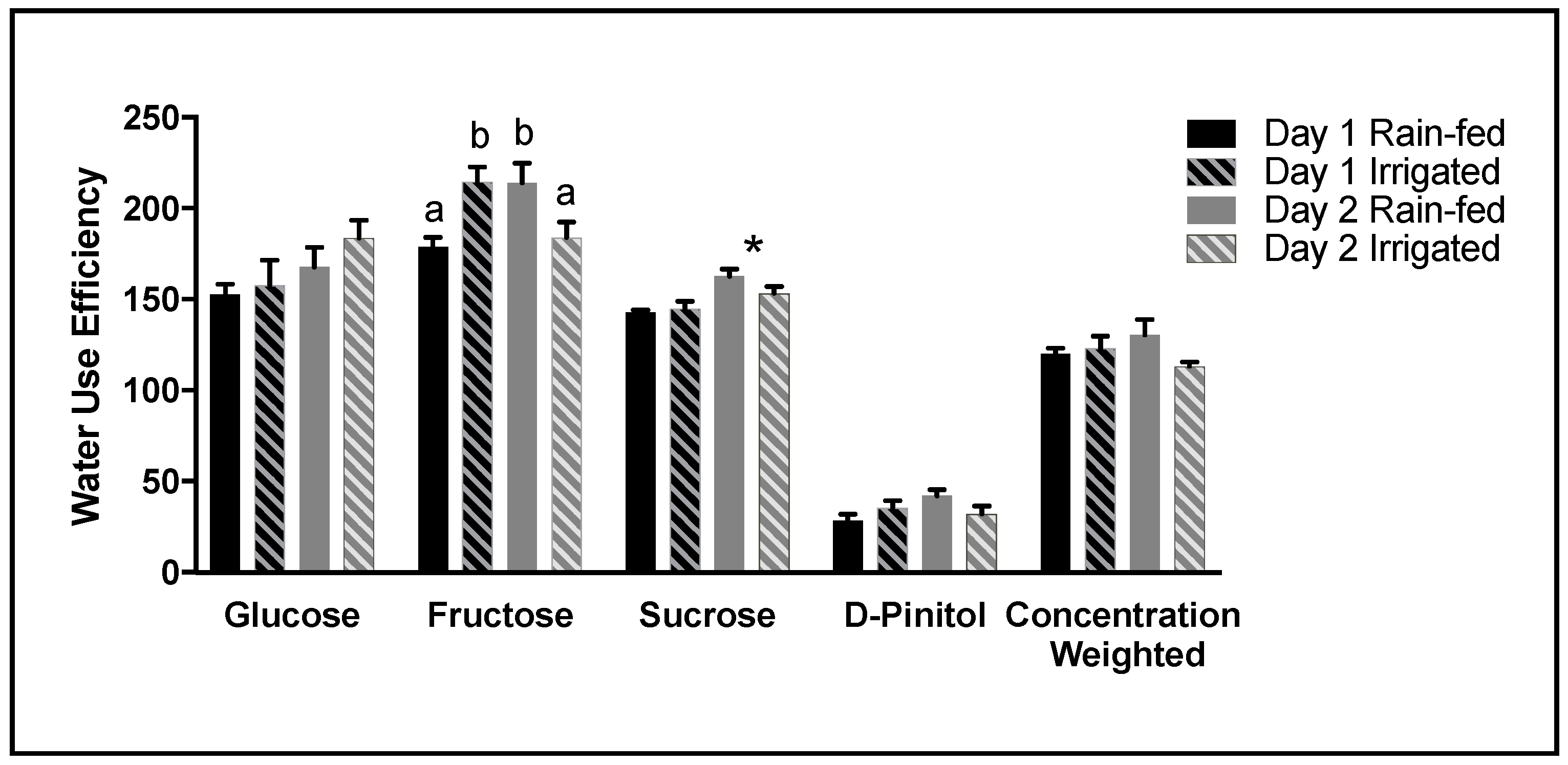
If substantial heterotrophic fractionation occurs during biosynthesis of a particular compound, such as D-pinitol (
Figure 7), such changes have the potential to influence the predicted WUE (
Figure 8) based upon the soluble fraction of the leaf. Our data illustrates this to be the case. This is especially important when the proportion of D-pinitol in the soluble fraction is high. Hence, the accumulation of D-pinitol with plant development as observed in this study increasingly impacts on the concentration weighted WUE, reducing the value of WUE obtained by predictions based on ∆
13C. This observation has major implications when comparing WUE at different developmental stages, as the change in predicted WUE might arise from a developmental change in chemical composition, rather than reflecting the influence of environmental conditions.
5. Conclusions
The present study sought to evaluate the use of candidate biomarkers for growth and productivity located in the soluble chemistry component of chickpea leaves. One significant aspect of the present study is that it was conducted under realistic field conditions. While controlled environment experiments are useful for closely monitoring shifts in carbon allocation within the plant system, they do not account for a plant’s response to the environment as a whole. Several aspects of growth under field conditions differ from that of controlled conditions, each of which may have an impact on the accumulation and allocation of carbon to different pools in plant tissues.
Here we have identified a major shift in the accumulation of soluble metabolites in leaves under field conditions that also carry a significantly different carbon isotopic composition to that of more commonly measured primary metabolites. The magnitude of both changes in compound concentration and δ13C significantly influences predictions of WUE based on soluble leaf carbon. Importantly, the accumulation of D-pinitol appears to be under developmental control. Future studies should aim to elucidate the influence of environmental stress (such as water availability) on D-pinitol under field conditions.










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}