Characterization of Spontaneous Immune Responses against Long Peptides Derived from Bcl-X(L) in Cancer Patients Using Elispot


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Donors
2.2. Peptides
2.3. Generation of Dendritic Cells
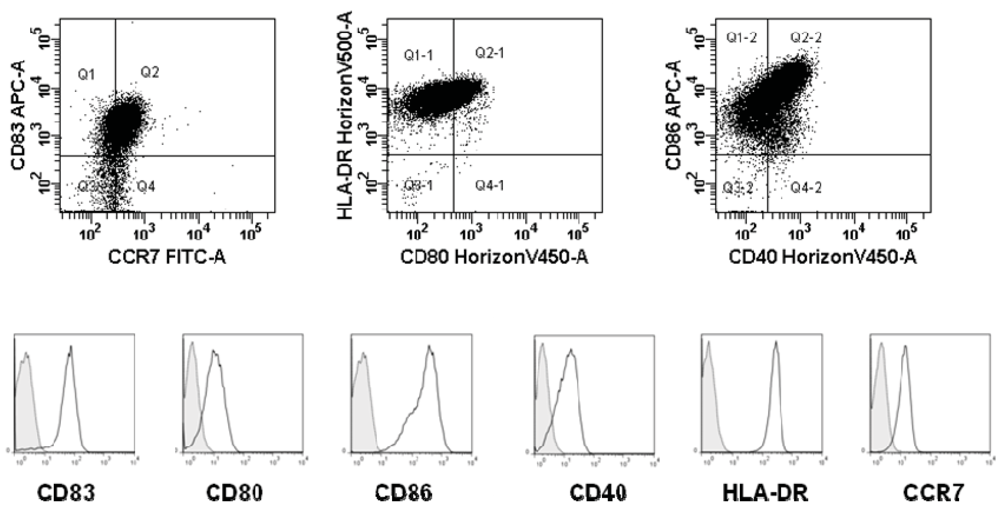
2.4. Phenotype of Mature DCs by Flow Cytometry
2.5. ELISPOT Assay
3. Results
3.1. Dendritic Cells

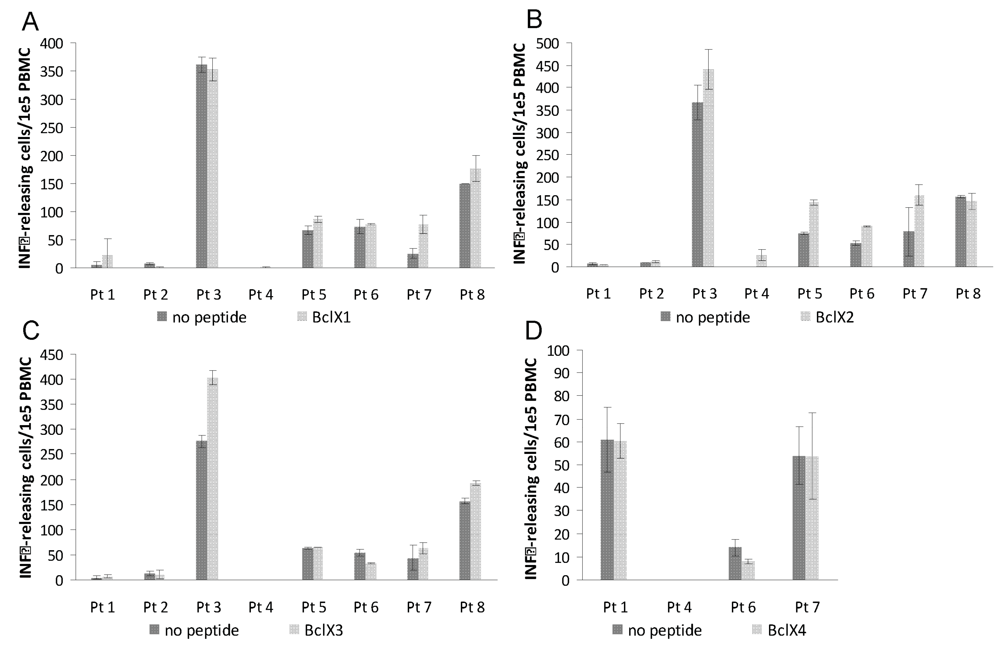
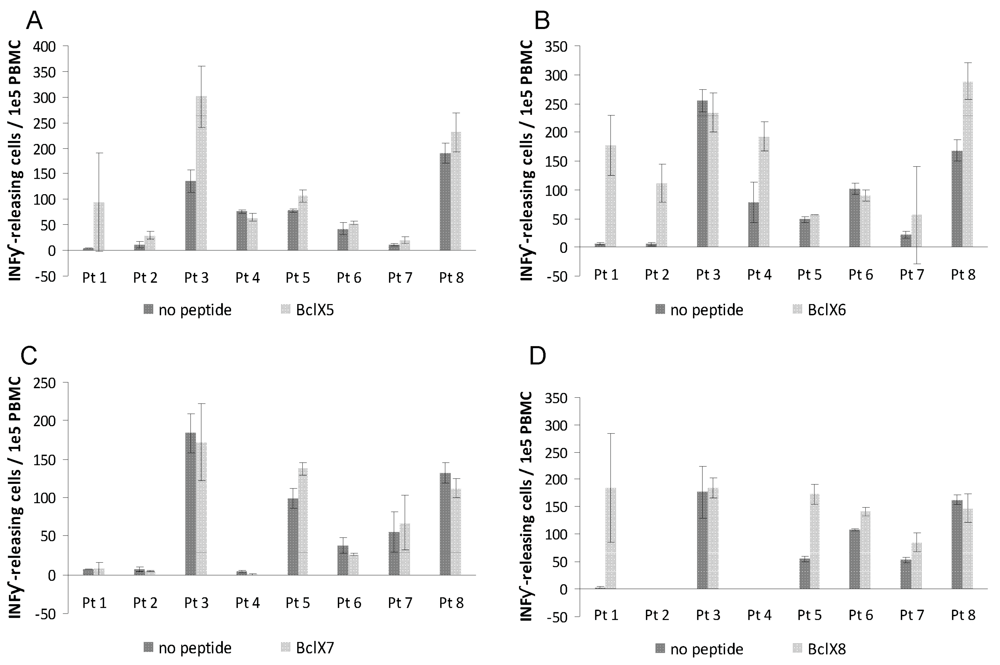
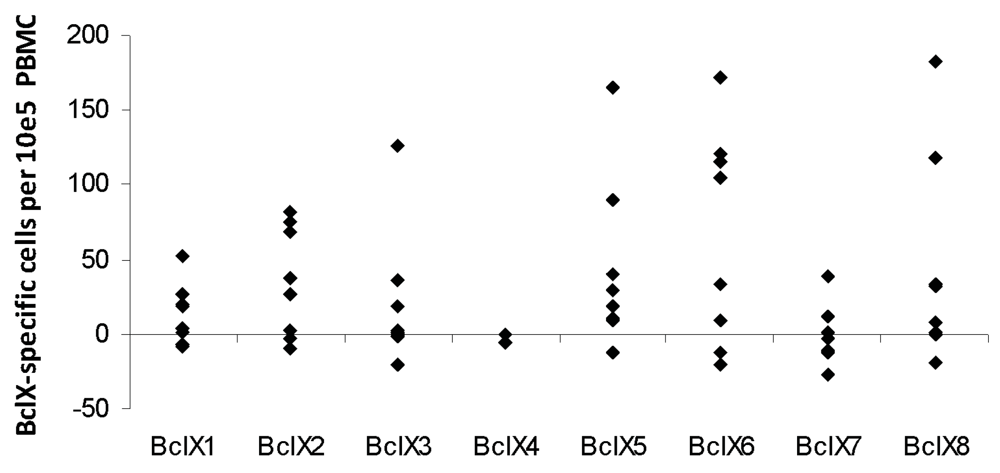
3.2. ELISPOT Responses against Long Bcl-X(L) Derived Peptides



4. Discussion
Acknowledgements
References
- Keilholz, U.; Weber, J.; Finke, J.H.; Gabrilovich, D.I.; Kast, W.M.; Disis, M.L.; Kirkwood, J.M.; Scheibenbogen, C.; Schlom, J.; Maino, V.C.; et al. Immunologic monitoring of cancer vaccine therapy: Results of a workshop sponsored by the Society for Biological Therapy. J. Immunother. 2002, 25, 97–138. [Google Scholar] [CrossRef]
- Herr, W.; Schneider, J.; Lohse, A.W.; Meyer zum Buschenfelde, K.H.; Wolfel, T. Detection and quantification of blood-derived CD8+ T lymphocytes secreting tumor necrosis factor alpha in response to HLA-A2.1-binding melanoma and viral peptide antigens. J. Immunol. Methods 1996, 191, 131–142. [Google Scholar] [CrossRef]
- Andersen, M.H.; Pedersen, L.O.; Becker, J.C.; thor Straten, P. Identification of a Cytotoxic T Lymphocyte Response to the Apoptose Inhibitor Protein Survivin in Cancer Patients. Cancer Res. 2001, 61, 869–872. [Google Scholar]
- Scheibenbogen, C.; Sun, Y.; Keilholz, U.; Song, M.; Stevanovic, S.; Asemissen, A.M.; Nagorsen, D.; Thiel, E.; Rammensee, H.G.; Schadendorf, D. Identification of known and novel immunogenic T-cell epitopes from tumor antigens recognized by peripheral blood T cells from patients responding to IL-2-based treatment. Int. J. Cancer. 2002, 20, 409–414. [Google Scholar]
- Herr, W.; Ranieri, E.; Gambotto, A.; Kierstead, L.S.; Amoscato, A.A.; Gesualdo, L.; Storkus, W.J. Identification of naturally processed and HLA-presented Epstein-Barr virus peptides recognized by CD4(+) or CD8(+) T lymphocytes from human blood. Proc. Natl. Acad. Sci. USA 1999, 96, 12033–12038. [Google Scholar]
- Andersen, M.H.; Tan, L.; Sondergaard, I.; Zeuthen, J.; Elliott, T.; Haurum, J.S. Poor correspondence between predicted and experimental binding of peptides to class I MHC molecules. Tissue Antigens. 2000, 55, 519–531. [Google Scholar]
- Andersen, M.H.; Reker, S.; Kvistborg, P.; Becker, J.C.; thor Straten, P. Spontaneous immunity against Bcl-X(L) in cancer patients. J. Immunol. 2005, 175, 2709–2714. [Google Scholar]
- Boise, L.H.; Gonzalez-Garcia, M.; Postema, C.E.; Ding, L.; Lindsten, T.; Turka, L.A.; Mao, X.; Nunez, G.; Thompson, C.B. bcl-x, a bcl-2-related gene that functions as a dominant regulator of apoptotic cell death. Cell 1993, 74, 597–608. [Google Scholar]
- McCutcheon, M.; Wehner, N.; Wensky, A.; Kushner, M.; Doan, S.; Hsiao, L.; Calabresi, P.; Ha, T.; Tran, T.V.; Tate, K.M.; et al. A sensitive ELISPOT assay to detect low-frequency human T lymphocytes. J. Immunol. Methods 1997, 210, 149–166. [Google Scholar] [CrossRef]
- Shangary, S.; Johnson, D.E. Recent advances in the development of anticancer agents targeting cell death inhibitors in the Bcl-2 protein family. Leukemia 2003, 17, 1470–1481. [Google Scholar]
- Kawasaki, H.; Altieri, D.C.; Lu, C.D.; Toyoda, M.; Tenjo, T.; Tanigawa, N. Inhibition of apoptosis by survivin predicts shorter survival rates in colorectal cancer. Cancer Res. 1998, 58, 5071–5074. [Google Scholar]
- Nakagawara, A. Molecular basis of spontaneous regression of neuroblastoma: Role of neurotrophic signals and genetic abnormalities. Hum. Cell 1998, 11, 115–124. [Google Scholar]
- Shafer-Weaver, K.; Sayers, T.; Strobl, S.; Derby, E.; Ulderich, T.; Baseler, M.; Malyguine, A. The Granzyme B ELISPOT assay: an alternative to the 51Cr-release assay for monitoring cell-mediated cytotoxicity. J. Transl. Med. 2003, 1, 14. [Google Scholar]
- Toes, R.E.; Offringa, R.; Blom, R.J.J.; Melief, C.J.M.; Kast, W.M. Peptide vaccination can leade to enhanced tumor growth through specific T-cell tolerance induction. Proc. Natl. Acad. Sci. USA 1996, 93, 7855–7860. [Google Scholar]
- Zwaveling, S.; Ferreira Mota, S.C.; Nouta, J.; Johnson, M.; Lipford, G.B.; Offringa, R.; van der Burg, S.H.; Melief, C.J.M. Established human papillomavirus type 16-expressing tumors are effectively eradicated following vaccination with long peptides. J. Immunol. 2002, 169, 350–358. [Google Scholar]
- Kennedy, R.; Celis, E. Multiple roles for CD4+ T cells in anti-tumor immune responses. Immunol. Rev. 2008, 222, 129–144. [Google Scholar]
- Knutson, K.L.; Disis, M.L. Augmenting T helper cell immunity in cancer. Curr. Drug Targets Immune Endocr. Metabol. Disord. 2005, 5, 365–371. [Google Scholar]
© 2012 by the authors; licensee MDPI, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Larsen, S.K.; Hansen, M.; Svane, I.M.; Straten, P.T.; Andersen, M.H. Characterization of Spontaneous Immune Responses against Long Peptides Derived from Bcl-X(L) in Cancer Patients Using Elispot. Cells 2012, 1, 51-60. https://doi.org/10.3390/cells1020051
Larsen SK, Hansen M, Svane IM, Straten PT, Andersen MH. Characterization of Spontaneous Immune Responses against Long Peptides Derived from Bcl-X(L) in Cancer Patients Using Elispot. Cells. 2012; 1(2):51-60. https://doi.org/10.3390/cells1020051
Chicago/Turabian StyleLarsen, Stine Kiaer, Morten Hansen, Inge Marie Svane, Per Thor Straten, and Mads Hald Andersen. 2012. "Characterization of Spontaneous Immune Responses against Long Peptides Derived from Bcl-X(L) in Cancer Patients Using Elispot" Cells 1, no. 2: 51-60. https://doi.org/10.3390/cells1020051