Danger: High Voltage—The Role of Voltage-Gated Calcium Channels in Central Nervous System Pathology



1
Institute of Anatomy and Cell Biology, University of Würzburg, 97070 Würzburg, Germany
2
Institute of Anatomy and Cell Biology, Friedrich-Alexander University Erlangen-Nürnberg, 91054 Erlangen, Germany
*
Author to whom correspondence should be addressed.
Cells 2017, 6(4), 43; https://doi.org/10.3390/cells6040043
Submission received: 14 September 2017
/
Revised: 10 November 2017
/
Accepted: 13 November 2017
/
Published: 15 November 2017
Abstract
:Voltage-gated calcium channels (VGCCs) are widely distributed within the central nervous system (CNS) and presumed to play an important role in the pathophysiology of a broad spectrum of CNS disorders including Alzheimer’s and Parkinson’s disease as well as multiple sclerosis. Several calcium channel blockers have been in clinical practice for many years so that their toxicity and side effects are well studied. However, these drugs are primarily used for the treatment of cardiovascular diseases and most if not all effects on brain functions are secondary to peripheral effects on blood pressure and circulation. While the use of calcium channel antagonists for the treatment of CNS diseases therefore still heavily depends on the development of novel strategies to specifically target different channels and channel subunits, this review is meant to provide an impulse to further emphasize the importance of future research towards this goal.
Keywords:
calcium; calcium channel antagonists; CNS; EAE; neurodegeneration; MS; regeneration; remyelination1. Calcium and Voltage-Gated Calcium Channels (VGCCs)
Calcium is one of the most important intracellular second messengers in the central nervous system (CNS). It regulates numerous cellular processes due to its electrogenic properties. These processes include neurotransmitter release, excitation, cell growth, proliferation, gene expression, long-term potentiation, plasticity and apoptosis [1,2,3]. In order to trigger and maintain Ca2+-dependent processes, an influx of cytosolic calcium from the extracytoplasmic space is required. This is achieved by calcium release from internal calcium stores or by entry of calcium ions via the cell membrane. In electrically excitable cells, voltage-gated calcium channels (VGCCs) are the main route for calcium entry into the cell after depolarization of the membrane. Despite mediating calcium influx, VGCCs also regulate intracellular processes depending on their localization. In cardiomyocytes, VGCCs regulate contraction processes; in endocrine cells, they control the secretion of hormones and in the CNS, they modulate the release of neurotransmitters [4]. Structurally, VGCCs are heteromultimeric complexes consisting of a central pore-forming Cavα1 subunit, which is conductive for ions. The central pore-forming subunit is convoyed by several auxiliary subunits (α2δ1-4, β1-4 and γ1-8) [5,6]. So far, ten different Cavα1 subunits have been described and classified according to their pharmacological and electrophysiological properties into high-voltage activated (HVA) and low-voltage activated (LVA) Ca2+ channels [4,5]. HVA Ca2+ channels include dihydropyridine-sensitive L (“long-lasting”)-type Cav1.1–1.4 and non-L-type Cav2.1–2.3 channels, which are less sensitive for DHP. Compared to LVA channels, which consist of the T (“transient”)-type Ca2+ channels Cav3.1–3.3, HVA channels require much stronger depolarization to reach the activation threshold. Additionally, they show prolonged channel opening [4,5,6] (Table 1).
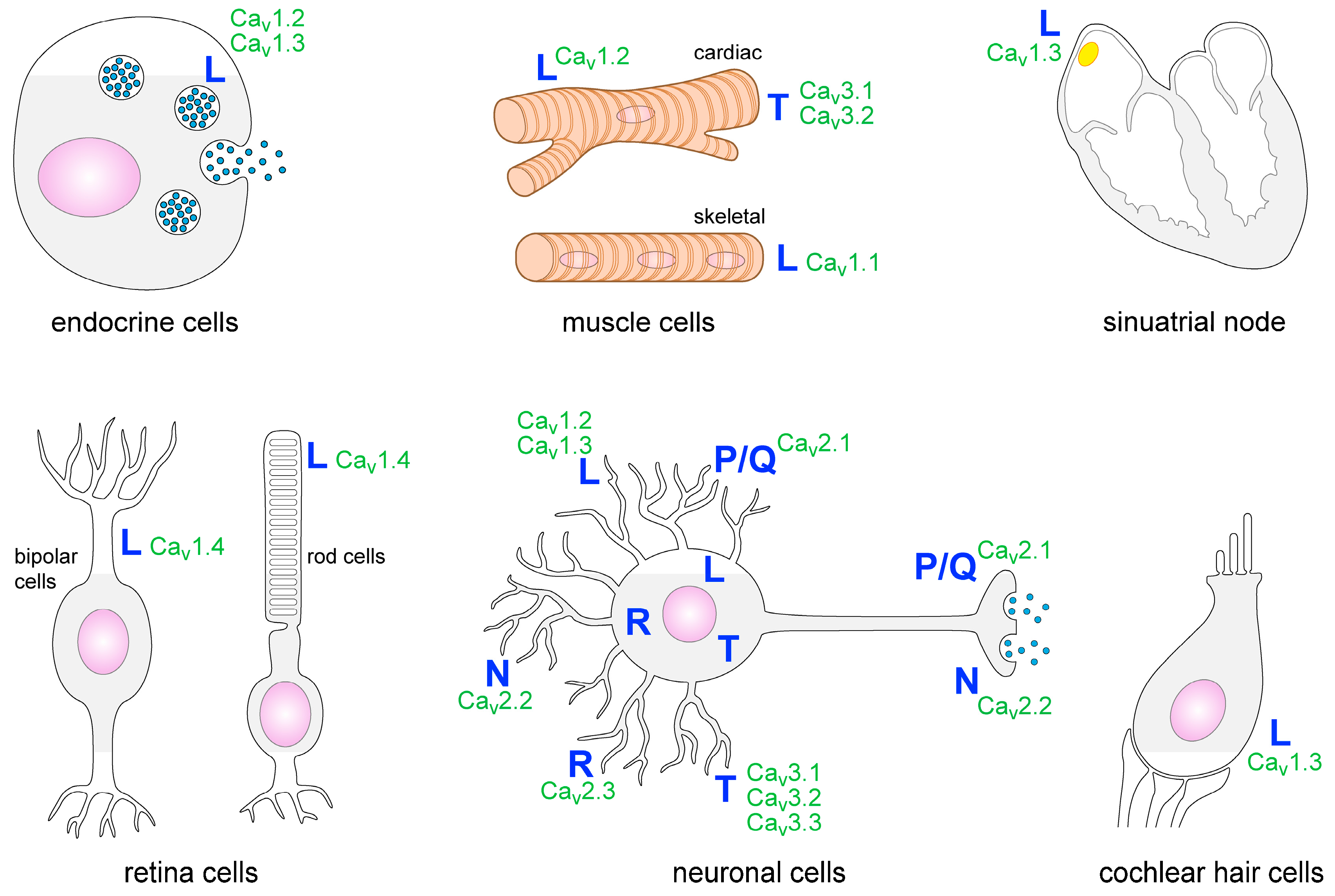
Within the nervous system, several types of VGCCs are expressed. They are detectable in many brain areas such as the cortex, thalamus and the hippocampus. P/Q-, T- and N-type VGCCs are the most common ones in the CNS [7]. Presynaptic P/Q- and N-type VGCCs induce neurotransmitter release and T-type VGCCs facilitate rhythmic burst firing of neurons. L-type VGCCs are localized on neuronal cell bodies as well as on dendrites and spines. Postsynaptic L-type VGCCs regulate gene expression and neuronal excitability (Figure 1) [4]. Some types of glial cells such as astrocytes, oligodendrocytes and glial precursor cells have also been shown to express VGCCs [2,8,9,10,11]. Outside the nervous system the heart, skeletal muscle cells, cells of the retina, endocrine cells, cochlear hair cells and cells of the immune system have been reported to express VGCCs or VGCC-like channels (Figure 1) [12,13,14,15].
2. Signs of Calcium-Mediated Cellular Damage
Intracytoplasmic calcium levels have to be strictly regulated in order to prevent cellular damage. In the CNS, neuronal organelles such as neurofilaments—and in particular mitochondria—are vulnerable to cytotoxicity [3,16,17,18,19]. Mitochondria play different roles in organisms, which comprise cellular respiration, temporary calcium storage, calcium buffering, maintenance of structural integrity and mediation of apoptosis [19,20,21]. Mitochondrial function can be directly influenced by extracellular signalling molecules. Increased nitric oxide (NO) levels, for instance, can alter gene expression and induce dysfunction of mitochondria. This in turn causes dysregulation of calcium homeostasis, resulting in enhanced cellular degeneration [19] and finally apoptosis of neurons and oligodendrocytes. Mitochondrial dysfunction can be detected ultrastructurally by an increased size (swelling) of mitochondria, reflecting an enhanced local energy demand [20,22,23]. Increased intracytoplasmic calcium levels also weaken neuronal integrity as they promote breakdown of the cytoskeleton, including actin, tubulin and intermediate filaments. This becomes evident both histologically and ultrastructurally as cytoplasmic blebbing and accumulation or dissolution of filaments [24,25]. Other detectable signs of calcium-mediated damage are dilatation of the endoplasmic reticulum and cytosolic shrinkage [24,25].
3. VGCCs in the Pathophysiology and Treatment of CNS Diseases
Studies of human diseases, mouse, rat and cell culture models indicate an important contribution of VGCCs to several neurological and psychiatric disorders, blindness and pain (Table 2) [26]. Of these conditions in particular Parkinson’s and Alzheimer’s disease have been in the focus of research mainly due to their tremendous socioeconomic relevance. In Parkinson’s disease, it has been demonstrated that dihydropyridines—potent VGCC antagonists—reduce the overall population risk in humans [27,28]. Evaluation of the pathogenesis of Alzheimer’s disease has revealed that pathogenic amyloid β (Aβ) peptides elevate L-type VGCC activity in cell cultures [29,30,31,32,33,34,35,36]. In addition, there was increased radiolabel binding to L-type VGCCs in the brains of Alzheimer’s disease patients post mortem [37]. Along these lines, L-type VGCC activity has been reported to be elevated during aging [38,39] and it is assumed to be involved in age-related alterations of synaptic function [38,40], membrane excitability [41] and cognition [42,43]. Yet, there is some controversy because studies of a mouse model of Alzheimer’s disease rather observed a decrease in L-type VGCC currents, suggesting a complex interplay between several factors including aging, the amount of circulating Aβ, Ca2+ dysregulation and Ca2+ release from the endoplasmic reticulum [44].
The continuous interest in using VGCCs as therapeutic targets to treat CNS disorders is also reflected by currently ongoing clinical trials, of which the majority uses L-type calcium channel antagonists. On the one hand, the L-type calcium channel blocker amlodipine is tested in a trial to reduce the risk for Alzheimer’s disease (NCT02913664) and a phase III trial on the use of the L-type calcium channel antagonist nilvadipine to treat Alzheimer’s disease was recently completed (NCT02017340) with results that are still expected. On the other hand, there are ongoing studies on the efficacy of isradipine in early Parkinson’s disease (NCT02168842). Yet, studies in a mouse model of Parkinson’s disease ask for caution since the plasma concentrations of isradipine approved for therapy were not neuroprotective, most likely due to the fact that the drug fails to reduce somatic calcium oscillations of dopaminergic neurons of the substantia nigra [45]. Isradpine is also currently investigated for cognitive enhancement in schizophrenia and schizoaffective disorder (NCT01658150). In addition, the drug is tested for the treatment of nicotine dependence (NCT03083353). There is one trial using the novel drug CX-8998, a T-type VGCC antagonist for the treatment of essential tremor (NCT03101241). Most recently, nimodipine—a Cav1.2 antagonist—was shown to be neuroprotective in the setting of experimental autoimmune encephalomyelitis—the most common animal model for multiple sclerosis—by limiting microglia-mediated damage of the CNS and promoting remyelination [46]. Interestingly, microglia are devoid of the Cav1.2 channel [46] so that the exact mechanism by which nimodipine acts on microglia still has to be elucidated in future studies. Table 2 summarizes hallmark diseases/syndromes and symptoms that are thought to be associated with VGCC involvement.
Overall, several studies regarding the role of VGCCs in CNS pathology exist and several attempts have been made to use VGCC antagonists as therapeutic targets in this context. While historically, VGCCs were targets of the first synthesized drugs [6], the establishment of ion channel-specific therapies for CNS disorders has so far proven to be difficult. Yet, the availability of a broad range of modern technologies such as RNAi, function-blocking antibodies and gene-editing present promising therapeutic avenues, which may be of particular importance for several still incurable and devastating CNS disorders including Alzheimer’s’ and Parkinson’s disease as well as multiple sclerosis.
Acknowledgments
We would like to thank Ursula Lebherz for critical reading of the manuscript and language revision. We are grateful to Michael Christof for the design of Figure 1.
Conflicts of Interest
The authors declare no conflict of interest.
Abbreviations
| Aβ | Amyloid β |
| CNS | Central nervous system |
| HVA | High-voltage activated |
| LVA | Low-voltage activated |
| VGCC | Voltage-gated calcium channel |
References
- Yagami, T.; Kohma, H.; Yagamoto, Y. L-type voltage-dependent calcium channels as therapeutic targets for neurodegenerative diseases. Curr. Med. Chem. 2012, 19, 4816–4827. [Google Scholar] [CrossRef] [PubMed]
- Braet, K.; Cabooter, L.; Paemeleire, K.; Leybaert, L. Calcium signal communication in the central nervous system. Biol. Cell 2004, 96, 79–91. [Google Scholar] [CrossRef] [PubMed]
- Young, W. Role of calcium in central nervous system injuries. J. Neurotrauma 1992, 9 (Suppl. S1), S9–S25. [Google Scholar] [PubMed]
- Catterall, W.A.; Perez-Reyes, E.; Snutch, T.P.; Striessnig, J. International Union of Pharmacology. XLVIII. Nomenclature and structure-function relationships of voltage-gated calcium channels. Pharmacol. Rev. 2005, 57, 411–425. [Google Scholar] [CrossRef] [PubMed]
- Wormuth, C.; Lundt, A.; Henseler, C.; Müller, R.; Broich, K.; Papazoglou, A.; Weiergräber, M. Cav2.3 R-type voltage-gated Ca2+ channels—Functional implications in convulsive and non-convulsive seizure activity. Open Neurol. J. 2016, 10, 99–126. [Google Scholar] [CrossRef] [PubMed]
- Waszkielewicz, A.M.; Gunia, A.; Szkaradek, N.; Słoczyńska, K.; Krupińska, S.; Marona, H. Ion channels as drug targets in central nervous system disorders. Curr. Med. Chem. 2013, 20, 1241–1285. [Google Scholar] [CrossRef] [PubMed]
- Schlick, B.; Flucher, B.E.; Obermair, G.J. Voltage-activated calcium channel expression profiles in mouse brain and cultured hippocampal neurons. Neuroscience 2010, 167, 786–798. [Google Scholar] [CrossRef] [PubMed]
- Verkhratsky, A.; Kettenmann, H. Calcium signalling in glial cells. Trends Neurosci. 1996, 19, 346–352. [Google Scholar] [CrossRef]
- Casamassima, F.; Hay, A.C.; Benedetti, A.; Lattanzi, L.; Cassano, G.B.; Perlis, R.H. L-type calcium channels and psychiatric disorders: A brief review. Am. J. Med. Genet. B Neuropsychiatr. Genet. 2010, 153B, 1373–1390. [Google Scholar] [CrossRef] [PubMed]
- Triggle, D.J. Calcium channel antagonists: Clinical uses—Past, present and future. Biochem. Pharmacol. 2007, 74, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Silei, V.; Fabrizi, C.; Venturini, G.; Salmona, M.; Bugiani, O.; Tagliavini, F.; Lauro, G.M. Activation of microglial cells by PrP and beta-amyloid fragments raises intracellular calcium through L-type voltage sensitive calcium channels. Brain Res. 1999, 818, 168–170. [Google Scholar] [CrossRef]
- Seitter, H.; Koschak, A. Relevance of tissue specific subunit expression in channelopathies. Neuropharmacology 2017. [Google Scholar] [CrossRef] [PubMed]
- Lyubchenko, T. Ca2+ signalling in B cells. Sci. World J. 2010, 10, 2254–2264. [Google Scholar] [CrossRef] [PubMed]
- Sadighi Akha, A.A.; Willmott, N.J.; Brickley, K.; Dolphin, A.C.; Galione, A.; Hunt, S.V. Anti-Ig-induced calcium influx in rat B lymphocytes mediated by cGMP through a dihydropyridine-sensitive channel. J. Biol. Chem. 1996, 13, 7297–7300. [Google Scholar] [CrossRef]
- Mesirca, P.; Torrente, A.G.; Mangoni, M.E. Functional role of voltage gated Ca(2+) channels in heart automaticity. Front. Physiol. 2015, 6, 19. [Google Scholar] [CrossRef] [PubMed]
- Stys, P.K.; Jiang, Q. Calpain-dependent neurofilament breakdown in anoxic and ischemic rat central axons. Neurosci. Lett. 2002, 328, 150–154. [Google Scholar] [CrossRef]
- Takano, Y.; Ohguro, H.; Dezawa, M.; Ishikawa, H.; Ohguro, I.; Mamiya, K.; Metoki, T.; Ishikawa, F.; Nakazawa, M. Study of drug effects of calcium channel blockers on retinal degeneration of rd mouse. Biochem. Biophys. Res. Commun. 2004, 313, 1015–1022. [Google Scholar] [CrossRef] [PubMed]
- Damaj, M.I.; Martin, B.R. Calcium agonists and antagonists of the dihydropyridine type: Effect on nicotine-induced antinociception and hypomotility. Drug Alcohol Depend. 1993, 32, 73–79. [Google Scholar] [CrossRef]
- Gonsette, R.E. Oxidative stress and excitotoxicity: A therapeutic issue in multiple sclerosis? Mult. Scler. J. 2008, 14, 22–34. [Google Scholar] [CrossRef] [PubMed]
- Waxman, S.G. Axonal conduction and injury in multiple sclerosis: The role of sodium channels. Nat. Rev. Neurosci. 2006, 7, 932–941. [Google Scholar] [CrossRef] [PubMed]
- Zambonin, J.L.; Zhao, C.; Ohno, N.; Campbell, G.R.; Engeham, S.; Ziabreva, I.; Schwarz, N.; Lee, S.E.; Frischer, J.M.; Turnbull, D.M.; et al. Increased mitochondrial content in remyelinated axons: Implications for multiple sclerosis. Brain 2011, 134, 1901–1913. [Google Scholar] [CrossRef] [PubMed]
- Dhib-Jalbut, S.; Arnold, D.L.; Cleveland, D.W.; Fisher, M.; Friedlander, R.M.; Mouradian, M.M.; Przedborski, S.; Trapp, B.D.; Wyss-Coray, T.; Yong, V.W. Neurodegeneration and neuroprotection in multiple sclerosis and other neurodegenerative diseases. J. Neuroimmunol. 2006, 176, 198–215. [Google Scholar] [CrossRef] [PubMed]
- Dutta, R.; McDonough, J.; Yin, X.; Peterson, J.; Chang, A.; Torres, T.; Gudz, T.; Macklin, W.B.; Lewis, D.A.; Fox, R.J.; et al. Mitochondrial dysfunction as a cause of axonal degeneration in multiple sclerosis patients. Ann. Neurol. 2006, 59, 478–489. [Google Scholar] [CrossRef] [PubMed]
- Trump, B.F.; Berezesky, I.K. Calcium-mediated cell injury and cell death. FASEB J. 1995, 9, 219–228. [Google Scholar] [PubMed]
- Soellner, I.A.; Rabe, J.; Mauri, V.; Kaufmann, J.; Addicks, K.; Kuerten, S. Differential aspects of immune cell infiltration and neurodegeneration in acute and relapse experimental autoimmune encephalomyelitis. Clin. Immunol. 2013, 149, 519–529. [Google Scholar] [CrossRef] [PubMed]
- Heyes, S.; Pratt, W.S.; Rees, E.; Dahimene, S.; Ferron, L.; Owen, M.J.; Dolphin, A.C. Genetic disruption of voltage-gated calcium channels in psychiatric and neurological disorders. Prog. Neurobiol. 2015, 134, 36–54. [Google Scholar] [CrossRef] [PubMed]
- Ritz, B.; Rhodes, S.L.; Qian, L.; Schernhammer, E.; Olsen, J.H.; Friis, S. L-type calcium channel blockers and Parkinson disease in Denmark. Ann. Neurol. 2010, 67, 600–606. [Google Scholar] [CrossRef] [PubMed]
- Surmeier, D.J.; Schumacker, P.T.; Guzman, J.D.; Ilijic, E.; Yang, B.; Zampese, E. Calcium and Parkinson’s disease. Biochem. Biophys. Res. Commun. 2017, 483, 1013–1019. [Google Scholar] [CrossRef] [PubMed]
- Davidson, R.M.; Shajenko, L.; Donta, T.S. Amyloid beta-peptide (A beta P) potentiates a nimodipine-sensitive L-type barium conductance in N1E-115 neuroblastoma cells. Brain Res. 1994, 643, 324–327. [Google Scholar] [CrossRef]
- Fu, H.; Li, W.; Lao, Y.; Luo, J.; Lee, N.T.; Kan, K.K.; Tsang, H.W.; Tsim, K.W.; Pang, Y.; Li, Z.; et al. Bis(7)-tacrine attenuates beta amyloid-induced neuronal apoptosis by regulating L-type calcium channels. J. Neurochem. 2006, 98, 1400–1410. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.; Rhim, H. Effects of amyloid-beta peptides on voltage-gated L-type Ca(V)1.2 and Ca(V)1.3 Ca (2+) channels. Mol. Cells 2011, 32, 289–294. [Google Scholar] [CrossRef] [PubMed]
- Ramsden, M.; Henderson, Z.; Pearson, H.A. Modulation of Ca2+ channel currents in primary cultures of rat cortical neurones by amyloid beta protein (1–40) is dependent on solubility status. Brain Res. 2002, 956, 254–261. [Google Scholar] [CrossRef]
- Scragg, J.L.; Fearon, I.M.; Boyle, J.P.; Ball, S.G.; Varadi, G.; Peers, C. Alzheimer’s amyloid peptides mediate hypoxic up-regulation of L-type Ca2+ channels. FASEB J. 2005, 19, 150–152. [Google Scholar] [CrossRef] [PubMed]
- Ueda, K.; Shinohara, S.; Yagami, T.; Asakura, K.; Kawasaki, K. Amyloid beta protein potentiates Ca2+ influx through L-type voltage-sensitive Ca2+ channels: A possible involvement of free radicals. J. Neurochem. 1997, 68, 265–271. [Google Scholar] [CrossRef] [PubMed]
- Webster, N.J.; Ramsden, M.; Boyle, J.P.; Pearson, H.A.; Peers, C. Amyloid peptides mediate hypoxic increase of L-type Ca2+ channels in central neurons. Neurobiol. Aging 2006, 27, 439–445. [Google Scholar] [CrossRef] [PubMed]
- Weiss, J.H.; Pike, C.J.; Cotman, C.W. Ca2+ channel blockers attenuate beta-amyloid peptide toxicity to cortical neurons in culture. J. Neurochem. 1994, 62, 372–375. [Google Scholar] [CrossRef] [PubMed]
- Coon, A.L.; Wallace, D.R.; Mactutus, C.F.; Booze, R.M. L-type calcium channels in the hippocampus and cerebellum of Alzheimer’s disease brain tissue. Neurobiol. Aging 1999, 20, 597–603. [Google Scholar] [CrossRef]
- Thibault, O.; Landfield, P.W. Increase in single L-type calcium channels in hippocampal neurons during aging. Science 1996, 272, 1017–1020. [Google Scholar] [CrossRef] [PubMed]
- Norris, C.M.; Blalock, E.; Chen, K.C.; Porter, N.M.; Thibault, O.; Kraner, S.D.; Landfield, P.W. Hippocampal ‘zipper’ slice studies reveal a necessary role for calcineurin in the increased activity of L-type Ca(2+) channels with aging. Neurobiol. Aging 2010, 31, 328–338. [Google Scholar] [CrossRef] [PubMed]
- Norris, C.M.; Halpain, S.; Foster, T.C. Reversal of age-related alterations in synaptic plasticity by blockade of L-type Ca2+ channels. J. Neurosci. 1998, 18, 3171–3179. [Google Scholar] [PubMed]
- Moyer, J.R., Jr.; Thompson, L.T.; Black, J.P.; Disterhoft, J.F. Nimodipine increases excitability of rabbit CA1 pyramidal neurons in an age- and concentration-dependent manner. J. Neurophysiol. 1992, 68, 2100–2109. [Google Scholar] [PubMed]
- Deyo, R.A.; Straube, K.T.; Disterhoft, J.F. Nimodipine facilitates associative learning in aging rabbits. Science 1989, 243, 809–811. [Google Scholar] [CrossRef] [PubMed]
- Veng, L.M.; Mesches, M.H.; Browning, M.D. Age-related working memory impairment is correlated with increases in the L-type calcium channel protein alpha1D (Cav1.3) in area CA1 of the hippocampus and both are ameliorated by chronic nimodipine treatment. Brain Res. Mol. Brain Res. 2003, 110, 193–202. [Google Scholar] [CrossRef]
- Thibault, O.; Pancani, T.; Landfield, P.W.; Norris, C.M. Reduction in neuronal L-type calcium channel activity in a double knock-in mouse model of Alzheimer’s disease. Biochim. Biophys. Acta 2012, 1822, 546–549. [Google Scholar] [CrossRef] [PubMed]
- Ortner, N.J.; Bock, G.; Dougalis, A.; Kharitonova, M.; Duda, J.; Hess, S.; Tuluc, P.; Pomberger, T.; Stefanova, N.; Pitterl, F.; et al. Lower affinity of isradipine for L-Type Ca2+ channels during substantia nigra dopamine neuron-like activity: Implications for neuroprotection in Parkinson's disease. J. Neurosci. 2017, 37, 6761–6777. [Google Scholar] [CrossRef] [PubMed]
- Schampel, A.; Volovitch, O.; Koeniger, T.; Scholz, C.J.; Jörg, S.; Linker, R.A.; Wischmeyer, E.; Wunsch, M.; Hell, J.W.; Ergün, S.; et al. Nimodipine fosters remyelination in a mouse model of multiple sclerosis and induces microglia-specific apoptosis. Proc. Natl. Acad. Sci. USA 2017, 114, E3295–E3304. [Google Scholar] [CrossRef] [PubMed]
- Splawski, I.; Timothy, K.W.; Decher, N.; Kumar, P.; Sachse, F.B.; Beggs, A.H.; Sanguinetti, M.C.; Keating, M.T. Severe arrhythmia disorder caused by cardiac L-type calcium channel mutations. Proc. Natl. Acad. Sci. USA 2005, 102, 8089–8096. [Google Scholar] [CrossRef] [PubMed]
- Busquet, P.; Hetzenauer, A.; Sinnegger-Brauns, M.J.; Striessnig, J.; Singewald, N. Role of L-type Ca2+ channel isoforms in the extinction of conditioned fear. Learn. Mem. 2008, 15, 378–386. [Google Scholar] [CrossRef] [PubMed]
- Dao, D.T.; Mahon, P.B.; Cai, X.; Kovacsics, C.E.; Blackwell, R.A.; Arad, M.; Shi, J.; Zandi, P.P.; O’Donnell, P.; Bipolar Genome Study (BiGS) Consortium; et al. Mood disorder susceptibility gene CACNA1C modifies mood-related behaviors in mice and interacts with sex to influence behavior in mice and diagnosis in humans. Biol. Psychiatry 2010, 68, 801–810. [Google Scholar] [CrossRef] [PubMed]
- Ostacher, M.J.; Iosifescu, D.V.; Hay, A.; Blumenthal, S.R.; Sklar, P.; Perlis, R.H. Pilot investigation of isradipine in the treatment of bipolar depression motivated by genome-wide association. Bipolar Disord. 2014, 16, 199–203. [Google Scholar] [CrossRef] [PubMed]
- Radzicki, D.; Yau, H.J.; Pollema-Mays, S.L.; Mlsna, L.; Cho, K.; Koh, S.; Martina, M. Temperature-sensitive Cav1.2 calcium channels support intrinsic firing of pyramidal neurons and provide a target for the treatment of febrile seizures. J. Neurosci. 2013, 33, 9920–9931. [Google Scholar] [CrossRef] [PubMed]
- Roca-Lapirot, O.; Radwani, H.; Aby, F.; Nagy, F.; Landry, M.; Fossat, P. Calcium signalling through L-type calcium channels: Role in pathophysiology of spinal nociceptive transmission. Br. J. Pharmacol. 2017. [Google Scholar] [CrossRef] [PubMed]
- Radwani, H.; Lopez-Gonzalez, M.J.; Cattaert, D.; Roca-Lapirot, O.; Dobremez, E.; Bouali-Benazzouz, R.; Eiríksdóttir, E.; Langel, Ü.; Favereaux, A.; Errami, M.; et al. Cav1.2 and Cav1.3 L-type calcium channels independently control short- and long-term sensitization to pain. J. Physiol. 2016, 594, 6607–6626. [Google Scholar] [CrossRef] [PubMed]
- Gadotti, V.M.; Bladen, C.; Zhang, F.X.; Chen, L.; Gündüz, M.G.; Simsek, R.; Safak, C.; Zamponi, G.W. Analgesic effect of a broad-spectrum dihydropyridine inhibitor of voltage-gated calcium channels. Pflugers Arch. 2015, 467, 2485–2493. [Google Scholar] [CrossRef] [PubMed]
- Ripke, S.; O’Dushlaine, C.; Chambert, K.; Moran, J.L.; Kahler, A.K.; Akterin, S.; Bergen, S.E.; Collins, A.L.; Crowley, J.J.; Fromer, M.; et al. Genome-wide association analysis identifies 13 new risk loci for schizophrenia. Nat. Genet. 2013, 45, 1150–1159. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nouvian, R.; Beutner, D.; Parsons, T.D.; Moser, T. Structure and function of the hair cell ribbon synapse. J. Membr. Biol. 2006, 209, 153–165. [Google Scholar] [CrossRef] [PubMed]
- Platzer, J.; Engel, J.; Schrott-Fischer, A.; Stephan, K.; Bova, S.; Chen, H.; Zheng, H.; Striessnig, J. Congenital deafness and sinoatrial node dysfunction in mice lacking class D L-type Ca2+ channels. Cell 2000, 102, 89–97. [Google Scholar] [CrossRef]
- Orthner, N.J.; Striessnig, J. L-type calcium channels as drug targets in CNS disorders. Channels 2016, 10, 7–13. [Google Scholar] [CrossRef] [PubMed]
- Striessnig, J.; Hoda, J.C.; Koschak, A.; Zaghetto, F.; Müllner, C.; Sinnegger-Brauns, M.J.; Wild, C.; Watschinger, K.; Trockenbacher, A.; Pelster, G. L-type Ca2+ channels in Ca2+ channelopathies. Biochem. Biophys. Res. Commun. 2004, 322, 1341–1346. [Google Scholar] [CrossRef] [PubMed]
- Bech-Hansen, N.T.; Naylor, M.J.; Maybaum, T.A.; Pearce, W.G.; Koop, B.; Fishman, G.A.; Mets, M.; Musarella, M.A.; Boycott, K.M. Loss-of-function mutations in a calcium-channel α1 subunit gene in Xp11.23 cause incomplete X-linked congenital stationary night blindness. Nat. Genet. 1998, 19, 264–267. [Google Scholar] [CrossRef] [PubMed]
- Nachbauer, W.; Nocker, M.; Karner, E.; Stankovic, I.; Unterberger, I.; Eigentler, A.; Schneider, R.; Poewe, W.; Delazer, M.; Boesch, S. Episodic ataxia type 2: Phenotype characteristics of a novel CACNA1A mutation and review of the literature. J. Neurol. 2014, 261, 983–991. [Google Scholar] [CrossRef] [PubMed]
- Baloh, R.W.; Jen, J.C. Genetics of familial episodic vertigo and ataxia. Ann. N. Y. Acad. Sci. 2002, 956, 338–345. [Google Scholar] [CrossRef] [PubMed]
- Van den Maagdenberg, A.M.; Pietrobon, D.; Pizzorusso, T.; Kaja, S.; Broos, L.A.; Cesetti, T.; van den Ven, R.C.; Tottene, A.; van der Kaa, J.; Plomp, J.J.; et al. A Cacna1a knockin migraine mouse model with increased susceptibility to cortical spreading depression. Neuron 2004, 41, 701–710. [Google Scholar] [CrossRef]
- Matsuyama, Z.; Kawakami, H.; Maruyama, H.; Izumi, Y.; Komure, O.; Udaka, F.; Kameyama, M.; Nishio, T.; Kuroda, Y.; Nishimura, M.; et al. Molecular features of the CAG repeats of spinocerebellar ataxia 6 (SCA6). Hum. Mol. Genet. 1997, 6, 1283–1287. [Google Scholar] [CrossRef] [PubMed]
- Zhuchenko, O.; Bailey, J.; Bonnen, P.; Ashizawa, T.; Stockton, D.W.; Amos, C.; Dobyns, W.B.; Subramony, S.H.; Zoghbi, H.Y.; Lee, C.C. Autosomal dominant cerebellar ataxia (SCA6) associated with small polyglutamine expansions in the alpha 1A-voltage-dependent calcium channel. Nat. Genet. 1997, 15, 62–69. [Google Scholar] [CrossRef] [PubMed]
- Zaman, T.; Lee, K.; Park, C.; Paydar, A.; Choi, J.H.; Cheong, E.; Lee, C.J.; Shin, H.S. Cav2.3 channels are critical for oscillatory burst discharges in the reticular thalamus and absence epilepsy. Neuron 2011, 70, 95–108. [Google Scholar] [CrossRef] [PubMed]
- Saegusa, H.; Kurihara, T.; Zong, S.; Minowa, O.; Kazuno, A.; Han, W.; Matsuda, Y.; Yamanaka, H.; Osanai, M.; Noda, T.; et al. Altered pain responses in mice lacking α1E subunit of the voltage-dependent Ca2+ channel. Proc. Natl. Acad. Sci. USA 2000, 97, 6132–6137. [Google Scholar] [CrossRef] [PubMed]
- Ernst, W.L.; Zhang, Y.; Yoo, J.W.; Ernst, S.J.; Noebels, J.L. Genetic enhancement of thalamocortical network activity by elevating α1G-mediated low-voltage-activated calcium current induces pure absence epilepsy. J. Neurosci. 2009, 29, 1615–1625. [Google Scholar] [CrossRef] [PubMed]
- Lu, A.T.; Dai, X.; Martinez-Agosto, J.A.; Cantor, R.M. Support for calcium channel gene defects in autism spectrum disorders. Mol. Autism 2012, 3, 18. [Google Scholar] [CrossRef] [PubMed]
- Bourinet, E.; Francois, A.; Laffray, S. T-type channels in neuropathic pain. Pain 2016, 127 (Suppl. S1), S15–S22. [Google Scholar] [CrossRef] [PubMed]
- Tai, C.H.; Yang, Y.C.; Pan, M.K.; Huang, C.S.; Kuo, C.C. Modulation of subthalamic T-type Ca2+ channels remedies locomotor deficits in a rat model of Parkinson disease. J. Clin. Investig. 2011, 121, 3289–3305. [Google Scholar] [CrossRef] [PubMed]
- Nimrich, V.; Eckert, A. Calcium channel blockers and dementia. Br. J. Pharmacol. 2013, 169, 1203–1210. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Distribution of VGCC subtypes on different cell types.


{kind=link}
Table 1.
Classification of voltage-gated calcium channels (VGCCs) according to their voltage-dependent activation.
Table 1.
Classification of voltage-gated calcium channels (VGCCs) according to their voltage-dependent activation.
| High-Voltage Activated | Family |
| L-type (“long-lasting”) VGCC | Cav1.1–Cav1.4 |
| P-type (“Purkinje cell”)/Q-type VGCC | Cav2.1 |
| N-type (“neural”) VGCC | Cav2.2 |
| R-type (“residual”/“resistant”) VGCC | Cav2.3 |
| Low-Voltage Activated | Family |
| T-type (“transient”) VGCC | Cav3.1–Cav3.3 |
Table 2.
Involvement of VGCCs in neurologic and psychiatric disorders.
| Channel | Disease/Symptom | Species |
|---|---|---|
| Cav1.2 | Autism/Timothy syndrome | Human [47] |
| Conditioned fear | Mouse [48] | |
| Depression/Mood disorders | Human, mouse [49,50] | |
| Febrile seizures | Rat [51] | |
| Multiple sclerosis | Mouse [46] | |
| Pain | Mouse, rat [52,53,54] | |
| Parkinson’s disease | Human, mouse (reviewed in [28]) | |
| Schizophrenia | Human [55] | |
| Cav1.3 | Deafness | Mouse [56,57] |
| Depression | Human, mouse [50,58] | |
| Pain | Rat [52,53] | |
| Parkinson’s disease | Human, mouse (reviewed in [28]) | |
| Cav1.4 | (Incomplete X-linked congenital stationary) night blindness | Human [59,60] |
| Cav2.1 | Episodic ataxia type 2 and familiar hemiplegic migraine type 1 | Human [61,62,63] |
| Spinocerebellar ataxia 6 | Human [64,65] | |
| Cav2.2 | Pain | Mouse [54] |
| Cav2.3 | Anxiety | Mouse [66] |
| Absence epilepsy | Mouse [66] | |
| Pain | Mouse [67] | |
| Cav3.1 | Thalamocortical network activity/ absence epilepsy | Mouse [68] |
| Cav3.1–3.3 | Autism/Autism spectrum disorders | Human [69] |
| Pain | Human, mouse, rat (reviewed in [70]) | |
| Parkinson’s disease/locomotor deficits | Rat [71] | |
| Various VGCCs | Alzheimer’s disease/dementia | Mouse, rat, human (reviewed in [72]) |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Schampel, A.; Kuerten, S. Danger: High Voltage—The Role of Voltage-Gated Calcium Channels in Central Nervous System Pathology. Cells 2017, 6, 43. https://doi.org/10.3390/cells6040043
AMA Style
Schampel A, Kuerten S. Danger: High Voltage—The Role of Voltage-Gated Calcium Channels in Central Nervous System Pathology. Cells. 2017; 6(4):43. https://doi.org/10.3390/cells6040043
Chicago/Turabian StyleSchampel, Andrea, and Stefanie Kuerten. 2017. "Danger: High Voltage—The Role of Voltage-Gated Calcium Channels in Central Nervous System Pathology" Cells 6, no. 4: 43. https://doi.org/10.3390/cells6040043
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.