Direct Detection of T- and B-Memory Lymphocytes by ImmunoSpot® Assays Reveals HCMV Exposure that Serum Antibodies Fail to Identify


 and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Human Subjects and PBMC
2.2. CD4 and CD8 Depletion of PBMC
2.3. Antigens
2.4. Human Interferon-γ ImmunoSpot® Assay
2.5. B-Cell ImmunoSpot® Assay
2.6. Statistical Analysis of ImmunoSpot(R) SFU Counts
2.7. HCMV PCR Assay
3. Results
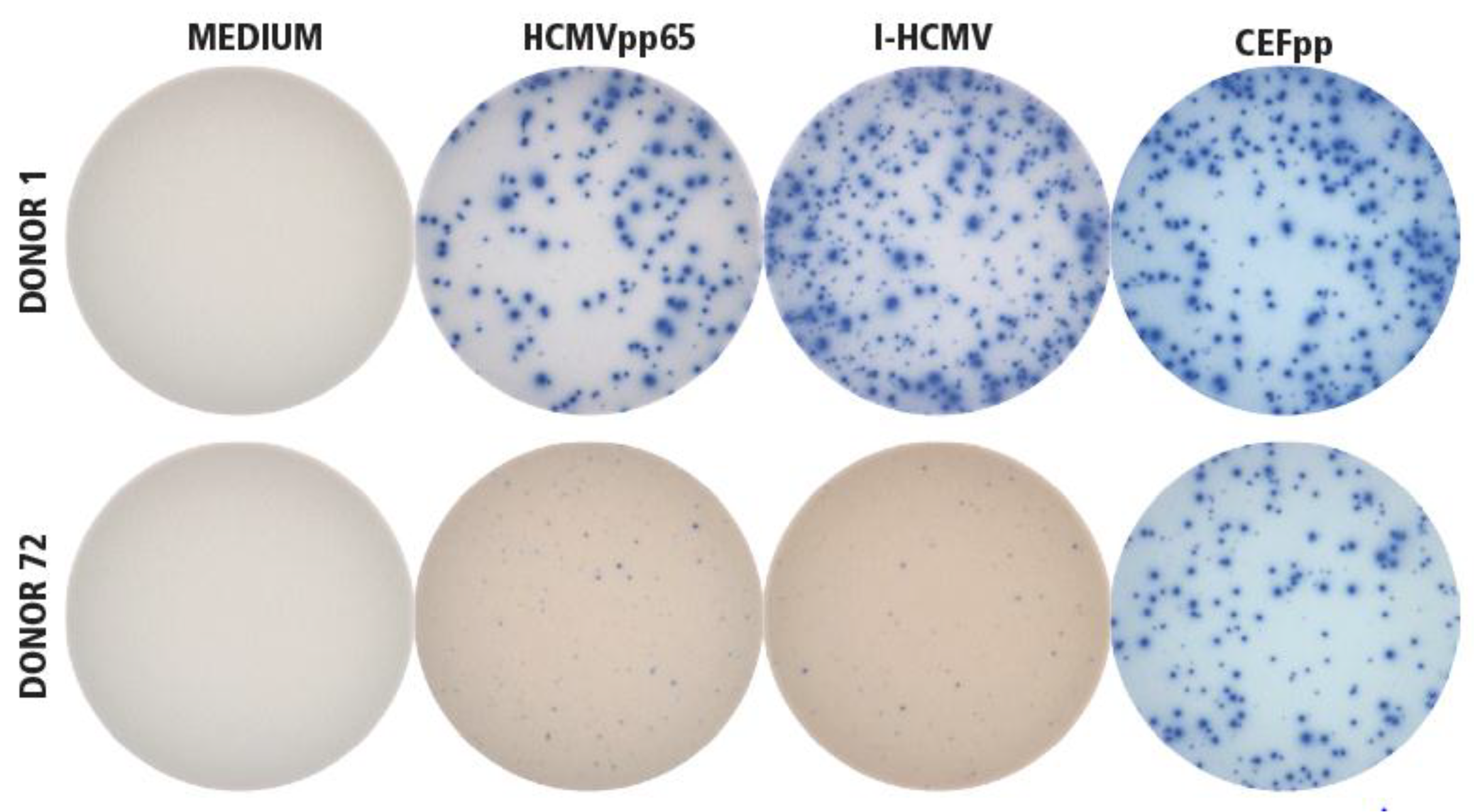
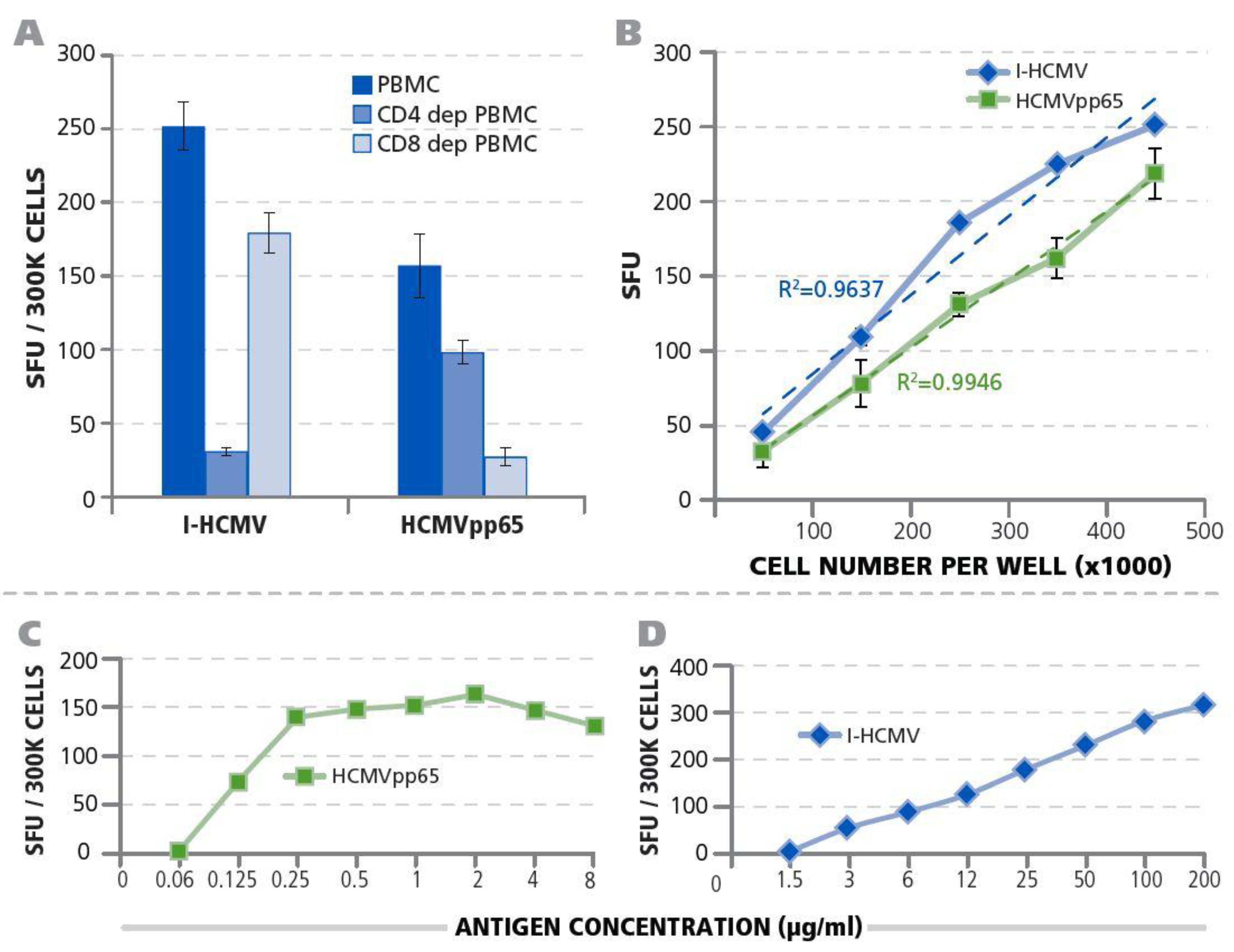
3.1. Detection of HCMV-Specific CD4 and CD8 Memory T Cells
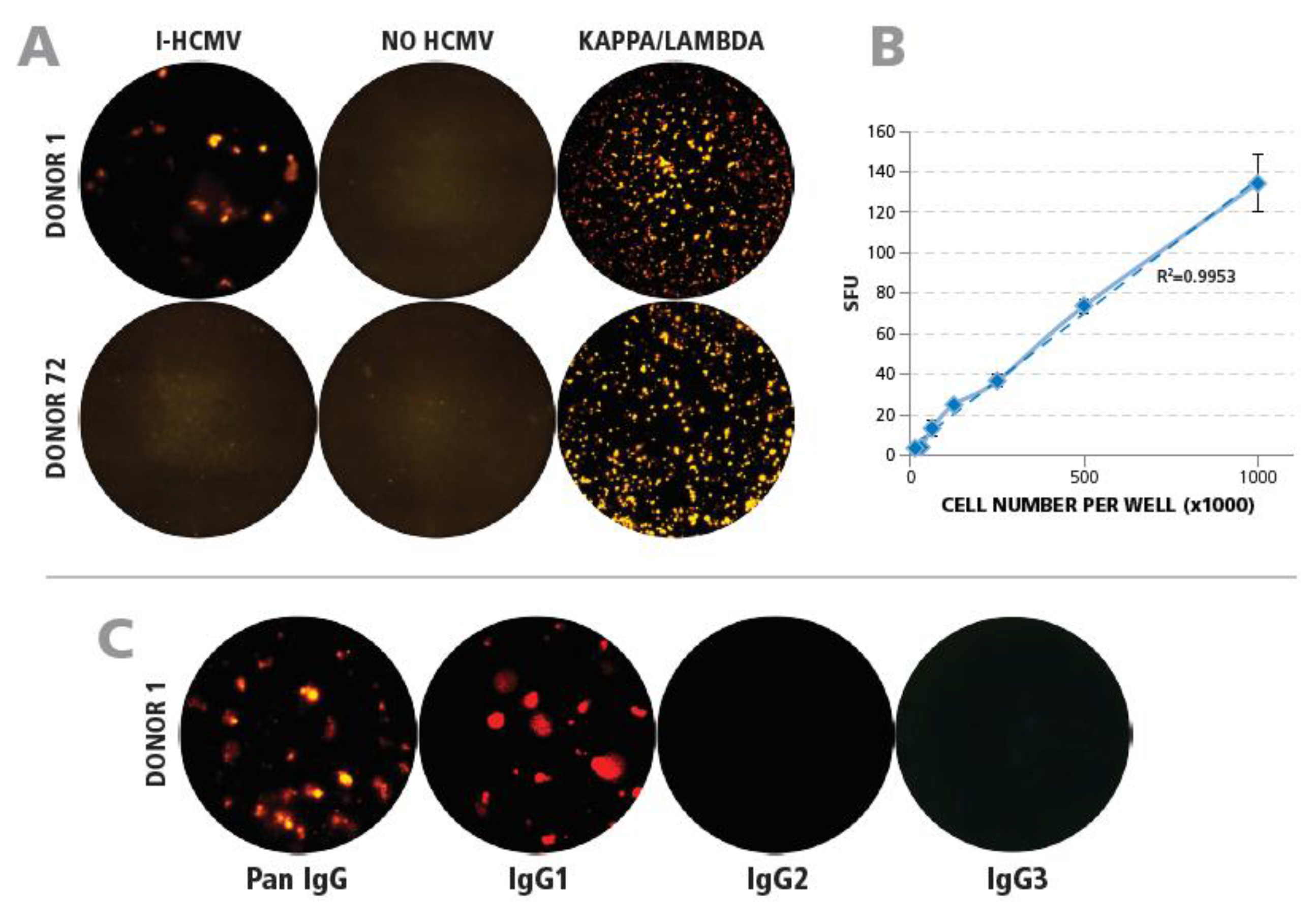
3.2. Detection of HCMV-Specific Memory B Cells
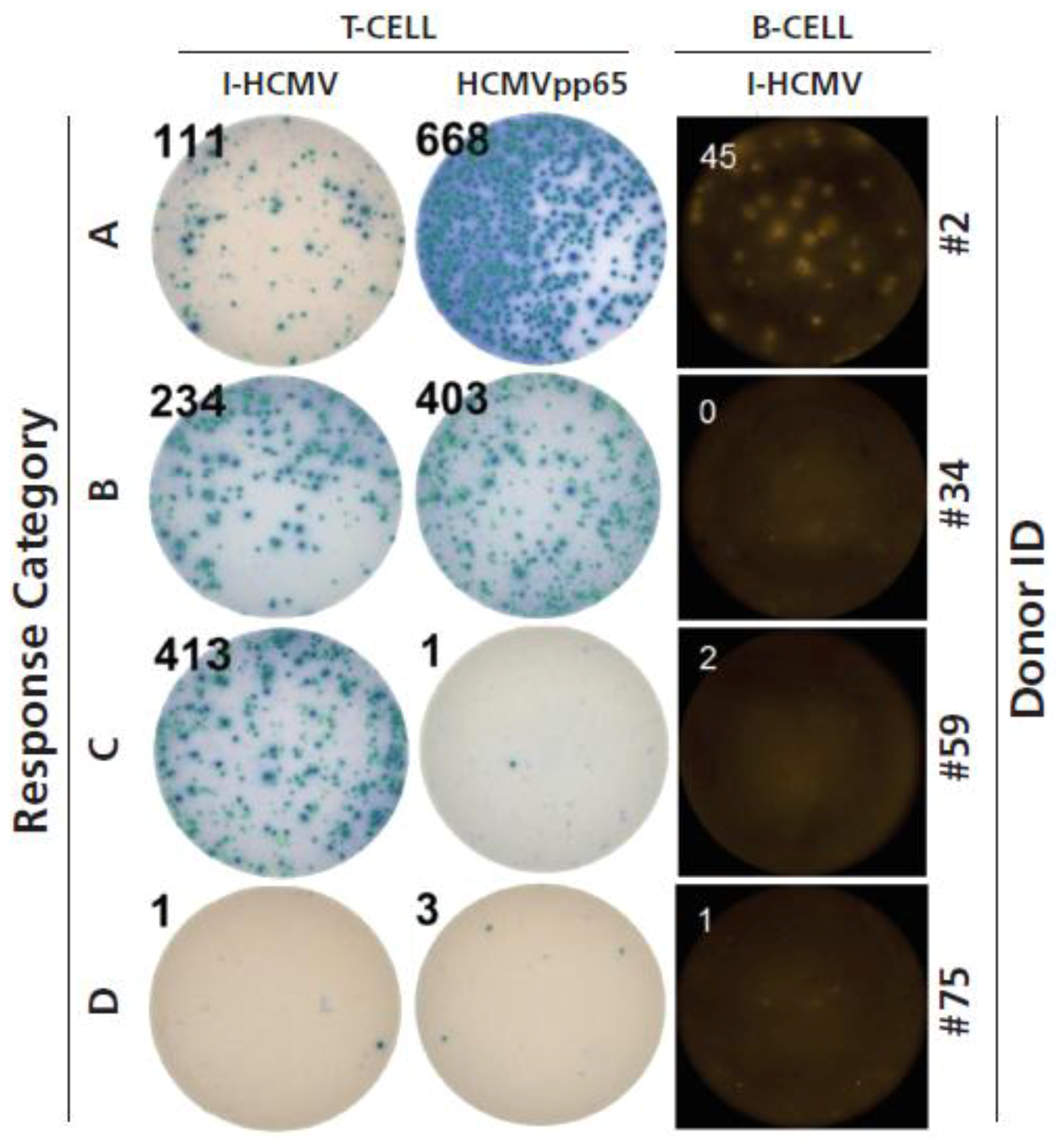
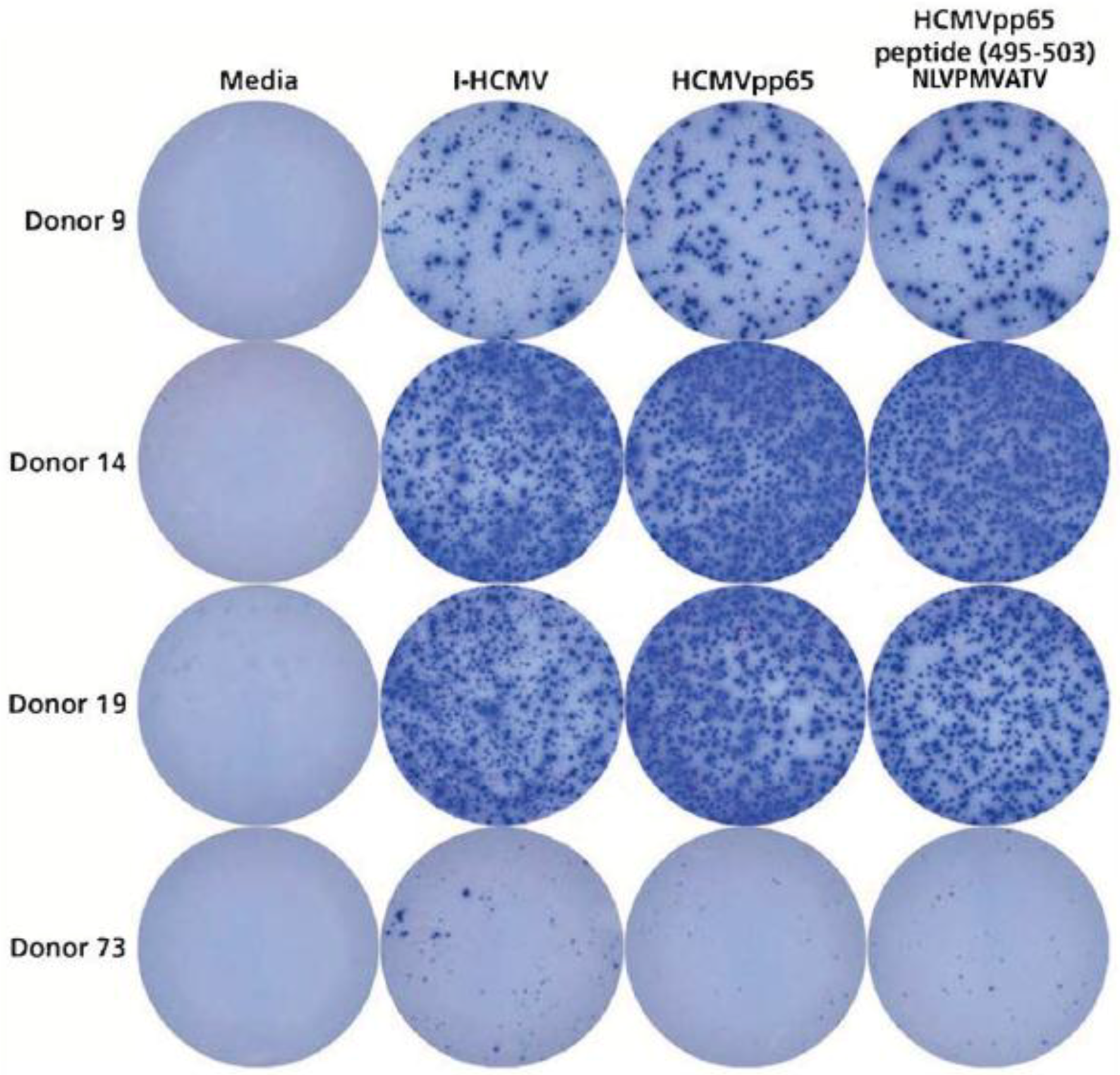
3.3. Testing HCMV Seronegative Human Donors for HCMV-Specific CD4+ T Cells, CD8+ T Cells, and B Cells
3.4. Detection of HCMVpp65 (495-503) Peptide-Specific T Cells in HLA-A*02:01-Positive Donors
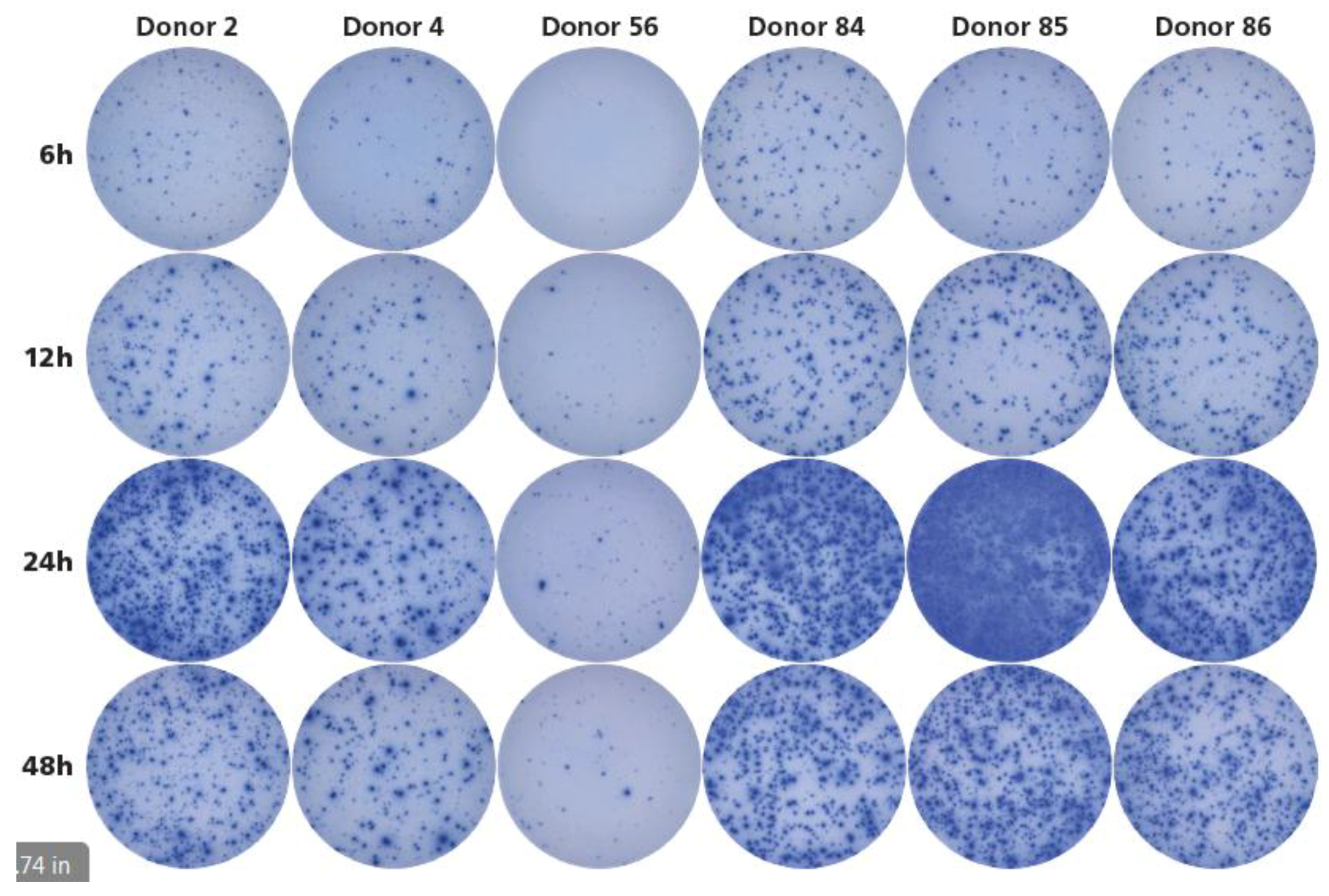
3.5. Twenty-Four Hours Antigen Stimulation Is Needed to Detect HCMV-Reactive Memory T Cells.
4. Discussion
Author Contributions
Acknowledgments
Conflicts of Interest
References
- Cannon, M.J.; Schmid, D.S.; Hyde, T.B. Review of cytomegalovirus seroprevalence and demographic characteristics associated with infection. Rev. Med. Virol. 2010, 20, 202–213. [Google Scholar] [CrossRef] [PubMed]
- Britt, W. Manifestations of human cytomegalovirus infection: Proposed mechanisms of acute and chronic disease. Curr. Top. Microbiol. Immunol. 2008, 325, 417–470. [Google Scholar] [PubMed]
- Panjwani, D.D.; Ball, M.G.; Berry, N.J.; Wimperis, J.Z.; Blacklock, H.A.; Prentice, H.G.; Hoffbrand, A.V.; Griffiths, P.D. Virological and serological diagnosis of cytomegalovirus infection in bone marrow allograft recipients. J. Med. Virol. 1985, 16, 357–365. [Google Scholar] [CrossRef] [PubMed]
- Quinnan, G.V., Jr.; Masur, H.; Rook, A.H.; Armstrong, G.; Frederick, W.R.; Epstein, J.; Manischewitz, J.F.; Macher, A.M.; Jackson, L.; Ames, J.; et al. Herpesvirus infections in the acquired immune deficiency syndrome. JAMA 1984, 252, 72–77. [Google Scholar] [CrossRef] [PubMed]
- Fiala, M.; Payne, J.E.; Berne, T.V.; Moore, T.C.; Henle, W.; Montgomerie, J.Z.; Chatterjee, S.N.; Guze, L.B. Epidemiology of cytomegalovirus infection after transplantation and immunosuppression. J. Infect. Dis. 1975, 132, 421–433. [Google Scholar] [CrossRef] [PubMed]
- Kotton, C.N. CMV: Prevention, Diagnosis and Therapy. Am. J. Transplant. 2013, 13, 24–40. [Google Scholar] [CrossRef] [PubMed]
- Ross, S.A.; Novak, Z.; Pati, S.; Boppana, S.B. Overview of the diagnosis of cytomegalovirus infection. Infect. Disord. Drug Targets 2011, 11, 466–474. [Google Scholar] [CrossRef] [PubMed]
- Humar, A.; Mazzulli, T.; Moussa, G.; Razonable, R.R.; Paya, C.V.; Pescovitz, M.D.; Covington, E.; Alecock, E. Clinical utility of cytomegalovirus (CMV) serology testing in high-risk CMV D+/R− transplant recipients. Am. J. Transplant. 2005, 5, 1065–1070. [Google Scholar] [CrossRef] [PubMed]
- Litjens, N.H.R.; Huang, L.; Dedeoglu, B.; Meijers, R.W.J.; Kwekkeboom, J.; Betjes, M.G.H. Protective cytomegalovirus (CMV)-specific T-cell immunity is frequent in kidney transplant patients without serum anti-CMV antibodies. Front. Immunol. 2017, 8, 1137. [Google Scholar] [CrossRef] [PubMed]
- Sester, M.; Gartner, B.C.; Sester, U.; Girndt, M.; Mueller-Lantzsch, N.; Kohler, H. Is the cytomegalovirus serologic status always accurate? A comparative analysis of humoral and cellular immunity. Transplantation 2003, 76, 1229–1230. [Google Scholar] [CrossRef] [PubMed]
- Zhu, J.; Paul, W.E. CD4 T cells: Fates, functions, and faults. Blood 2008, 112, 1557–1569. [Google Scholar] [CrossRef] [PubMed]
- Bunde, T.; Kirchner, A.; Hoffmeister, B.; Habedank, D.; Hetzer, R.; Cherepnev, G.; Proesch, S.; Reinke, P.; Volk, H.D.; Lehmkuhl, H.; et al. Protection from cytomegalovirus after transplantation is correlated with immediate early 1-specific CD8 T cells. J. Exp. Med. 2005, 201, 1031–1036. [Google Scholar] [CrossRef] [PubMed]
- Wrammert, J.; Ahmed, R. Maintenance of serological memory. Biol. Chem. 2008, 389, 537–539. [Google Scholar] [CrossRef] [PubMed]
- Crotty, S.; Ahmed, R. Immunological memory in humans. Semin. Immunol. 2004, 16, 197–203. [Google Scholar] [CrossRef] [PubMed]
- Bottiger, M.; Gustavsson, O.; Svensson, A. Immunity to tetanus, diphtheria and poliomyelitis in the adult population of Sweden in 1991. Int. J. Epidemiol. 1998, 27, 916–925. [Google Scholar] [CrossRef] [PubMed]
- Kreher, C.R.; Dittrich, M.T.; Guerkov, R.; Boehm, B.O.; Tary-Lehmann, M. CD4+ and CD8+ cells in cryopreserved human PBMC maintain full functionality in cytokine Elispot assays. J. Immunol. Methods 2003, 278, 79–93. [Google Scholar] [CrossRef]
- Ramachandran, H.; Laux, J.; Moldovan, I.; Caspell, R.; Lehmann, P.V.; Subbramanian, R.A. Optimal thawing of cryopreserved peripheral blood mononuclear cells for use in high-throughput human immune monitoring studies. Cells 2012, 1, 313–324. [Google Scholar] [CrossRef] [PubMed]
- Currier, J.R.; Kuta, E.G.; Turk, E.; Earhart, L.B.; Loomis-Price, L.; Janetzki, S.; Ferrari, G.; Birx, D.L.; Cox, J.H. A panel of MHC class I restricted viral peptides for use as a quality control for vaccine trial Elispot assays. J. Immunol. Methods 2002, 260, 157–172. [Google Scholar] [CrossRef]
- Koch, S.; Solana, R.; Dela Rosa, O.; Pawelec, G. Human cytomegalovirus infection and T cell immunosenescence: A mini review. Mech. Ageing Dev. 2006, 127, 538–543. [Google Scholar] [CrossRef] [PubMed]
- Immunospot Resources, ELISPOT Protocol. Available online: http://www.immunospot.com/resources/protocols/ELISPOT-protocol.htm (accessed on 26 April 2018).
- Karulin, A.Y.; Karacsony, K.; Zhang, W.; Targoni, O.S.; Moldovan, I.; Dittrich, M.; Sundararaman, S.; Lehmann, P.V. Elispots produced by CD8 and CD4 cells follow log normal size distribution permitting objective counting. Cells 2015, 4, 56–70. [Google Scholar] [CrossRef] [PubMed]
- Czerkinsky, C.C.; Nilsson, L.A.; Nygren, H.; Ouchterlony, O.; Tarkowski, A. A solid-phase enzyme-linked immunospot (Elispot) assay for enumeration of specific antibody-secreting cells. J. Immunol. Methods 1983, 65, 109–121. [Google Scholar] [CrossRef]
- Hesse, M.D.; Karulin, A.Y.; Boehm, B.O.; Lehmann, P.V.; Tary-Lehmann, M. A T cell clone’s avidity is a function of its activation state. J. Immunol. (Baltim. Md. 1950) 2001, 167, 1353–1361. [Google Scholar] [CrossRef]
- Wills, M.R.; Carmichael, A.J.; Mynard, K.; Jin, X.; Weekes, M.P.; Plachter, B.; Sissons, J.G. The human cytotoxic T-lymphocyte (CTL) response to cytomegalovirus is dominated by structural protein pp65: Frequency, specificity, and T-cell receptor usage of pp65-specific CTL. J. Virol. 1996, 70, 7569–7579. [Google Scholar] [PubMed]
- Duechting, A.; Przybyla, A.; Kuerten, S.; Lehmann, P.V. Delayed activation kinetics of Th2- and Th17 cells compared to Th1 cells. Cells 2017, 6. [Google Scholar] [CrossRef] [PubMed]
- Kotton, C.N.; Kumar, D.; Caliendo, A.M.; Asberg, A.; Chou, S.; Danziger-Isakov, L.; Humar, A. Transplantation Society International, CMV Consensus Group. Updated international consensus guidelines on the management of cytomegalovirus in solid-organ transplantation. Transplantation 2013, 96, 333–360. [Google Scholar] [CrossRef] [PubMed]
- Zhu, J.; Shearer, G.M.; Marincola, F.M.; Norman, J.E.; Rott, D.; Zou, J.P.; Epstein, S.E. Discordant cellular and humoral immune responses to cytomegalovirus infection in healthy blood donors: Existence of a Th1-type dominant response. Int. Immunol. 2001, 13, 785–790. [Google Scholar] [CrossRef] [PubMed]
- Nikolich-Zugich, J. Ageing and life-long maintenance of T-cell subsets in the face of latent persistent infections. Nat. Rev. Immunol. 2008, 8, 512–522. [Google Scholar] [CrossRef] [PubMed]
- Apetoh, L.; Smyth, M.J.; Drake, C.G.; Abastado, J.P.; Apte, R.N.; Ayyoub, M.; Blay, J.Y.; Bonneville, M.; Butterfield, L.H.; Caignard, A.; et al. Consensus nomenclature for CD8+ T cell phenotypes in cancer. Oncoimmunology 2015, 4, e998538. [Google Scholar] [CrossRef] [PubMed]
- Wunsch, M.; Zhang, W.; Hanson, J.; Caspell, R.; Karulin, A.Y.; Recks, M.S.; Kuerten, S.; Sundararaman, S.; Lehmann, P.V. Characterization of the HCMV-specific CD4 T cell responses that are associated with protective immunity. Viruses 2015, 7, 4414–4437. [Google Scholar] [CrossRef] [PubMed]
- Nagy, Z.A.; Lehmann, P.V.; Falcioni, F.; Muller, S.; Adorini, L. Why peptides? Their possible role in the evolution of MHC-restricted T-cell recognition. Immunol. Today 1989, 10, 132–138. [Google Scholar] [CrossRef]
- Rowland-Jones, S.; Sutton, J.; Ariyoshi, K.; Dong, T.; Gotch, F.; McAdam, S.; Whitby, D.; Sabally, S.; Gallimore, A.; Corrah, T.; et al. HIV-specific cytotoxic T-cells in HIV-exposed but uninfected gambian women. Nat. Med. 1995, 1, 59–64. [Google Scholar] [CrossRef] [PubMed]
- Clerici, M.; Levin, J.M.; Kessler, H.A.; Harris, A.; Berzofsky, J.A.; Landay, A.L.; Shearer, G.M. HIV-specific T-helper activity in seronegative health care workers exposed to contaminated blood. JAMA 1994, 271, 42–46. [Google Scholar] [CrossRef] [PubMed]
- De Maria, A.; Cirillo, C.; Moretta, L. Occurrence of human immunodeficiency virus type 1 (HIV-1)-specific cytolytic T cell activity in apparently uninfected children born to HIV-1-infected mothers. J. Infect. Dis. 1994, 170, 1296–1299. [Google Scholar] [CrossRef] [PubMed]
- Lucia, M.; Luque, S.; Crespo, E.; Melilli, E.; Cruzado, J.M.; Martorell, J.; Jarque, M.; Gil-Vernet, S.; Manonelles, A.; Grinyo, J.M.; et al. Preformed circulating HLA-specific memory B cells predict high risk of humoral rejection in kidney transplantation. Kidney Int. 2015, 88, 874–887. [Google Scholar] [CrossRef] [PubMed]
- Bonilla, F.A. Pharmacokinetics of immunoglobulin administered via intravenous or subcutaneous routes. Immunol. Allergy Clin. N. Am. 2008, 28, 803–819. [Google Scholar] [CrossRef] [PubMed]
- Rawlinson, W.D. Diagnosis of human cytomegalovirus infection and disease. Pathology 1999, 31, 109–115. [Google Scholar] [CrossRef] [PubMed]
- De la Hoz, R.E.; Stephens, G.; Sherlock, C. Diagnosis and treatment approaches of CMV infections in adult patients. J. Clin. Virol. 2002, 25 (Suppl. 2), S1–S12. [Google Scholar] [CrossRef]
- Wong, P.; Pamer, E.G. CD8 T cell responses to infectious pathogens. Annu. Rev. Immunol. 2003, 21, 29–70. [Google Scholar] [CrossRef] [PubMed]
- Lieberman, J. The ABCs of granule-mediated cytotoxicity: New weapons in the arsenal. Nat. Rev. Immunol. 2003, 3, 361–370. [Google Scholar] [CrossRef] [PubMed]
- Oehen, S.; Brduscha-Riem, K. Differentiation of naive CTL to effector and memory CTL: Correlation of effector function with phenotype and cell division. J. Immunol. 1998, 161, 5338–5346. [Google Scholar] [PubMed]
- Weninger, W.; Manjunath, N.; von Andrian, U.H. Migration and differentiation of CD8+ T cells. Immunol. Rev. 2002, 186, 221–233. [Google Scholar] [CrossRef] [PubMed]
- Wherry, E.J.; Ahmed, R. Memory CD8 T-cell differentiation during viral infection. J. Virol. 2004, 78, 5535–5545. [Google Scholar] [CrossRef] [PubMed]
- Nowacki, T.M.; Kuerten, S.; Zhang, W.; Shive, C.L.; Kreher, C.R.; Boehm, B.O.; Lehmann, P.V.; Tary-Lehmann, M. Granzyme B production distinguishes recently activated CD8(+) memory cells from resting memory cells. Cell. Immunol. 2007, 247, 36–48. [Google Scholar] [CrossRef] [PubMed]
- Solomon, G.F.; Benton, D.; Harker, J.O.; Bonavida, B.; Fletcher, M.A. Prolonged asymptomatic states in HIV-seropositive persons with fewer than 50 CD4+ T cells per MM3. Psychoneuroimmunologic findings. Ann. N. Y. Acad. Sci. 1994, 741, 185–190. [Google Scholar] [CrossRef] [PubMed]
- Nowacki, T.M.; Bettenworth, D.; Ross, M.; Heidemann, J.; Lehmann, P.V.; Lugering, A. Cytomegalovirus (CMV)-specific perforin and granzyme B Elispot assays detect reactivation of CMV infection in inflammatory bowel disease. Cells 2012, 1, 35–50. [Google Scholar] [CrossRef] [PubMed]
- Germain, R.N. The biochemistry and cell biology of antigen presentation by MHC class I and class II molecules. Implications for development of combination vaccines. Ann. N. Y. Acad. Sci. 1995, 754, 114–125. [Google Scholar] [CrossRef] [PubMed]
- Engelhard, V.H. Structure of peptides associated with class I and class II MHC molecules. Ann. Rev. Immunol. 1994, 12, 181–207. [Google Scholar] [CrossRef] [PubMed]
- Bernasconi, N.L.; Traggiai, E.; Lanzavecchia, A. Maintenance of serological memory by polyclonal activation of human memory B cells. Science 2002, 298, 2199–2202. [Google Scholar] [CrossRef] [PubMed]
- Schiller, A.; Zhang, T.; Li, R.; Duechting, A.; Sundararaman, S.; Przybyla, A.; Kuerten, S.; Lehmann, P.V. A positive control for detection of functional CD4 T cells in PBMC: The CPI pool. Cells 2017, 6. [Google Scholar] [CrossRef] [PubMed]
- Varani, S.; Landini, M.P. Cytomegalovirus-induced immunopathology and its clinical consequences. Herpesviridae 2011, 2, 6. [Google Scholar] [CrossRef] [PubMed]
- Rothe, K.; Quandt, D.; Schubert, K.; Rossol, M.; Klingner, M.; Jasinski-Bergner, S.; Scholz, R.; Seliger, B.; Pierer, M.; Baerwald, C.; et al. Latent CMV infection in rheumatoid arthritis increases frequencies of cytolytic lir-1+ CD8+ T cells. Arthritis Rheumatol. 2015. [Google Scholar] [CrossRef]
- Ciccocioppo, R.; Racca, F.; Paolucci, S.; Campanini, G.; Pozzi, L.; Betti, E.; Riboni, R.; Vanoli, A.; Baldanti, F.; Corazza, G.R. Human cytomegalovirus and epstein-barr virus infection in inflammatory bowel disease: Need for mucosal viral load measurement. World J. Gastroenterol. 2015, 21, 1915–1926. [Google Scholar] [CrossRef] [PubMed]
- Olson, N.C.; Doyle, M.F.; Jenny, N.S.; Huber, S.A.; Psaty, B.M.; Kronmal, R.A.; Tracy, R.P. Decreased naive and increased memory CD4(+) T cells are associated with subclinical atherosclerosis: The multi-ethnic study of atherosclerosis. PLoS ONE 2013, 8, e71498. [Google Scholar] [CrossRef] [PubMed]
- Swain, S.L.; McKinstry, K.K.; Strutt, T.M. Expanding roles for CD4(+) T cells in immunity to viruses. Nat. Rev. Immunol. 2012, 12, 136–148. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Donor Serial # | T-cells | B-cells | Donor Serial # | T-cells | B-cells | ||
|---|---|---|---|---|---|---|---|
| HCMVpp65 | I-HCMV | I-HCMV | HCMVpp65 | I-HCMV | I-HCMV | ||
| 1 | 122 | 362 | 45 | 42 | 133 | 515 | not perf. |
| 43 | 353 | 661 | not perf. | ||||
| 2 | 668 | 111 | 45 | 44 | 847 | 721 | not perf. |
| 3 | 189 | 77 | 101 | 45 | 281 | 890 | not perf. |
| 4 | 403 | 52 | 21 | 46 | 31 | 20 | 25 |
| 5 | 29 | 465 | 20 | 47 | not perf. | >1000 | 26 |
| 6 | 243 | 440 | 85 | 48 | 196 | 48 | 9 |
| 7 | 345 | 459 | 59 | 49 | 2 | 23 | 21 |
| 8 | 515 | >1000 | 76 | 50 | 27 | 16 | 152 |
| 9 | 280 | 237 | 38 | 51 | 2 | 73 | 11 |
| 10 | 607 | 646 | 62 | 52 | 86 | 17 | 17 |
| 11 | 385 | 764 | 33 | 53 | 21 | 82 | 1 |
| 12 | 357 | 708 | 13 | 54 | 5 | 104 | 2 |
| 13 | 401 | 480 | 13 | 55 | 3 | 169 | 1 |
| 14 | 835 | >1000 | 133 | 56 | 1 | 71 | 1 |
| 15 | 192 | 435 | 29 | 57 | 5 | 148 | 1 |
| 16 | 72 | 522 | 23 | 58 | 1 | 74 | 3 |
| 17 | 551 | 382 | 95 | 59 | 1 | 413 | 2 |
| 18 | 162 | 385 | 72 | 60. | 0 | 83 | not perf. |
| 19 | 792 | 697 | 43 | 61 | 5 | 79 | not perf. |
| 20 | 237 | 336 | 133 | 62 | 3 | 19 | 2 |
| 21 | 506 | 611 | 89 | 63 | 0 | 46 | not perf. |
| 22 | 533 | 473 | 128 | 64 | 1 | 49 | not perf. |
| 23 | 567 | 433 | 78 | 65 | 0 | 40 | 1 |
| 24 | 515 | 422 | 48 | 66 | 0 | 26 | 2 |
| 25 | 23 | 79 | 104 | 67 | 0 | 46 | 1 |
| 26 | 266 | 193 | 110 | 68 | 1 | 23 | 1 |
| 27 | 95 | 194 | 72 | 69 | 2 | 4 | 2 |
| 28 | 465 | 153 | 79 | 70 | 2 | 3 | 1 |
| 29 | 145 | 202 | 8 | 71 | 2 | 4 | 0 |
| 30 | 65 | 239 | 8 | 72 | 2 | 4 | 2 |
| 31 | 298 | 461 | 6 | 73 | 2 | 8 | 3 |
| 32 | 271 | 519 | 3 | 74 | 2 | 8 | 3 |
| 33 | 163 | 50 | 4 | 75 | 3 | 1 | 1 |
| 34 | 403 | 234 | 0 | 76 | 3 | 1 | 6 |
| 35 | 68 | 148 | 2 | 77 | 5 | 4 | not perf. |
| 36 | 301 | 420 | 2 | 78 | 5 | 4 | not perf. |
| 37 | 176 | 87 | 1 | 79 | 3 | 9 | not perf. |
| 38 | 88 | 433 | 4 | 80 | 1 | 8 | not perf. |
| 39 | 90 | 110 | 5 | 81 | 1 | 3 | not perf. |
| 40 | 52 | 766 | 3 | 82 | 0 | 0 | not perf. |
| 41 | 69 | 239 | not perf. | 83 | 1 | 1 | not perf. |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Terlutter, F.; Caspell, R.; Nowacki, T.M.; Lehmann, A.; Li, R.; Zhang, T.; Przybyla, A.; Kuerten, S.; Lehmann, P.V. Direct Detection of T- and B-Memory Lymphocytes by ImmunoSpot® Assays Reveals HCMV Exposure that Serum Antibodies Fail to Identify. Cells 2018, 7, 45. https://doi.org/10.3390/cells7050045
Terlutter F, Caspell R, Nowacki TM, Lehmann A, Li R, Zhang T, Przybyla A, Kuerten S, Lehmann PV. Direct Detection of T- and B-Memory Lymphocytes by ImmunoSpot® Assays Reveals HCMV Exposure that Serum Antibodies Fail to Identify. Cells. 2018; 7(5):45. https://doi.org/10.3390/cells7050045
Chicago/Turabian StyleTerlutter, Fredrik, Richard Caspell, Tobias M. Nowacki, Alexander Lehmann, Ruliang Li, Ting Zhang, Anna Przybyla, Stefanie Kuerten, and Paul V. Lehmann. 2018. "Direct Detection of T- and B-Memory Lymphocytes by ImmunoSpot® Assays Reveals HCMV Exposure that Serum Antibodies Fail to Identify" Cells 7, no. 5: 45. https://doi.org/10.3390/cells7050045