1. Introduction
Humoral immune responses represent one of the strongest known correlates of protection against various microbial and viral pathogens, as well as toxins [
1]. Traditionally, the presence of antibodies in serum has been measured for the assessment of humoral immunity. Serum antibodies, however, provide only indirect and incomplete insights into the functions of the B cell system [
2]. In vivo, antibody molecules have a rather short half-life in serum: for IgG1, IgG2, and IgG4 it is 20 to 29 days, while for IgG3 it is 7 to 15 days [
3]. Therefore, the presence of serum antibodies in vivo depends on their continuous production by plasma cells—immune memory that has been imprinted in the past through infection or immunization will reveal itself in serum antibody measurement only if such plasma B cells are present and continue to produce antibodies long after the antigen has been cleared [
2,
4].
The presence of antibodies in serum of individuals may or may not accurately reflect on the existence of humoral (or cellular) long term immunological memory (i.e., the presence of memory B or T cells in a host). On one hand, life-long antibody persistence has been documented following smallpox and flu immunizations, long after the clearance of the respective viruses [
5,
6]. On the other, antibody titers elicited following vaccinations against measles, tetanus toxoid, diphtheria, and poliomyelitis are known to wane over time, requiring regular booster immunizations to sustain protective antibody levels [
7,
8]. In yet a third scenario, memory B cells can be present in a host, capable of engaging in secondary antibody responses, however, in the absence of serum antibodies [
9,
10]. Thus, in addition to monitoring serum antibody levels, ascertaining the magnitude and diversity of long-lived B-cell memory populations can provide a more complete understanding of immune protection by antibodies following the re-exposure to the antigen.
The detection and enumeration of antigen-specific B cell memory cells in blood is best performed by ELISPOT [
11,
12]. Using this approach, not only the frequency of these cells can be established within peripheral blood mononuclear cells (PBMC) revealing the extent of their clonal sizes, but the assay is also suited to reveal the antibody classes and subclasses that these B cells produce, providing insights into the effector functions of B cell memory. While tetramers and other multimers can be used for the detection and study of rare antigen-specific T cells in PBMC [
13], ELISPOT has been the primary approach for B cell immune monitoring and has been used to assess B cell memory in various antigenic and pathogenic systems [
14,
15,
16,
17].
During an immune response, naïve B cells, which occur in undetectably low numbers in PBMC, proliferate and differentiate into antibody secreting (B) cells (ASC), also called plasma cells, or B cell blasts. The first generation of antibodies produced by such ASC are IgM antibodies. Subsequently, immunoglobulin (Ig) class switching occurs, giving rise to ASC and memory cells capable of producing other Ig classes and subclasses. ASC in freshly isolated blood that are actively secreting antibody can only be observed during an ongoing immune encounter, and in the first weeks following the clearance of the antigen [
5]. Therefore, detecting such ASC in freshly isolated blood, directly ex vivo, provides an important immunodiagnostic marker for identifying ongoing immune processes in the body vs. serum antibodies or memory B cells that in most cases do not permit to distinguish between long term immune memory and an actively ongoing antigen encounter [
18]. Active B cell blasts (ASC) can be detected in the so called Direct B Cell ELISPOT assay, in which freshly isolated PBMC are plated in the assay, without additional activation. One central question that the experiments reported here addresses is whether such spontaneously secreting B cell blasts can be cryopreserved in PBMC, and subsequently thawed and cultured without disrupting their ongoing Ig secretion process.
Upon successful elimination of an antigen, circulating ASC are no longer detectable in peripheral blood [
5]. Instead, the memory B cells become resting lymphocytes that do not secrete antibody unless re-stimulated [
12]. Thus, the detection of such quiescent memory B cells in PBMC requires that they be first activated to become ASC, and thus can be detected in the so called Indirect B Cell ELISPOT assay. Over the years, a variety of approaches for polyclonal B cell stimulation has been developed [
19], of which a protocol reported by Pinna et al. has proven to be the most effective [
19]: it relies on R848 (a TLR 7/8 agonist) and Interleukin 2 (IL-2) to achieve stimulation [
20]. Comparing different protocols, we too found the combination of R848 plus IL-2 to provide the most potent polyclonal B cell stimulation (R.C., unpublished data), and used it for the present study. The second principal question addressed here is whether resting memory B cells in PBMC can be cryopreserved and subsequently thawed without disrupting their ability to re-engage in antibody production after polyclonal B cell stimulation in vitro.
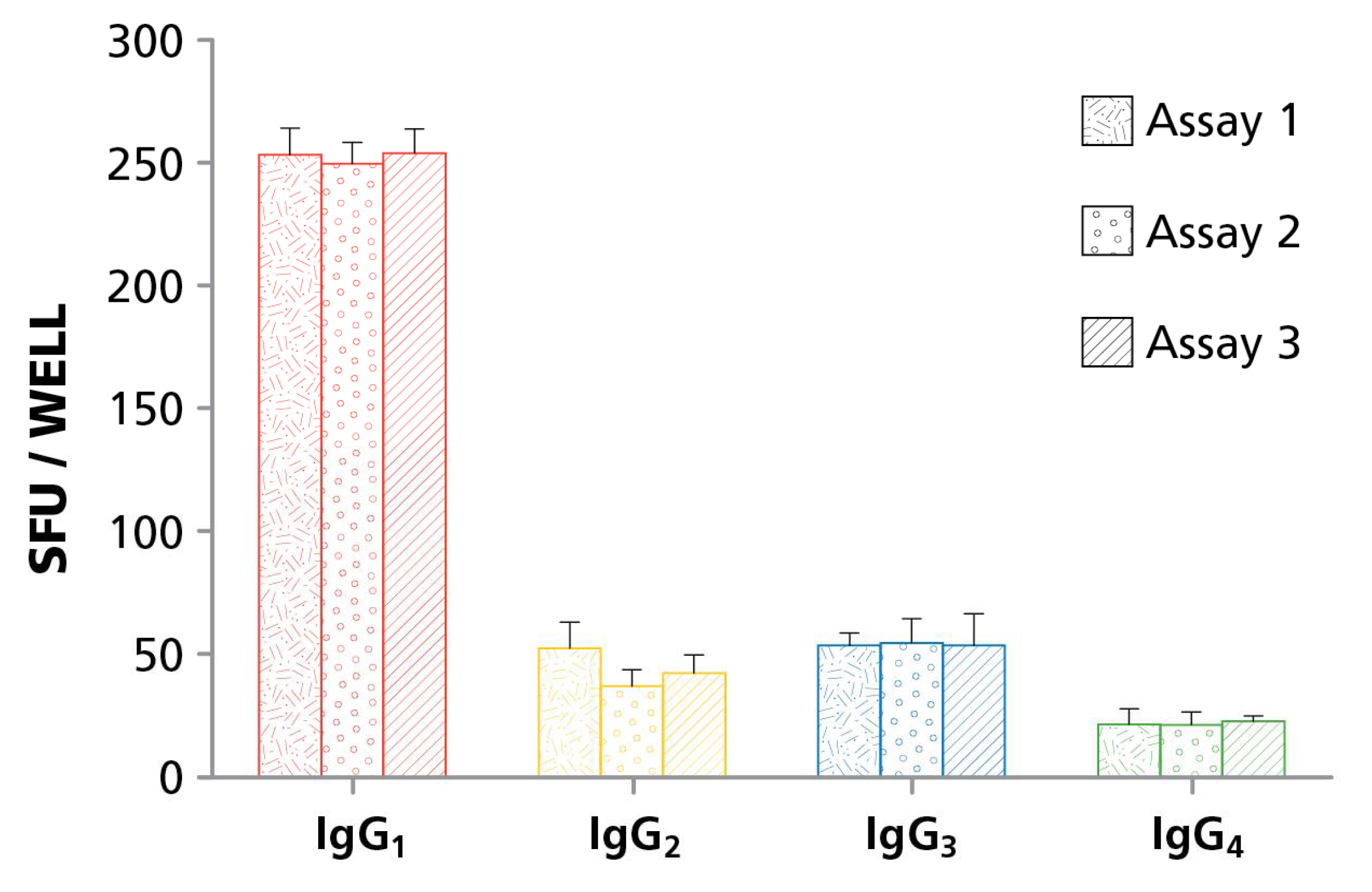
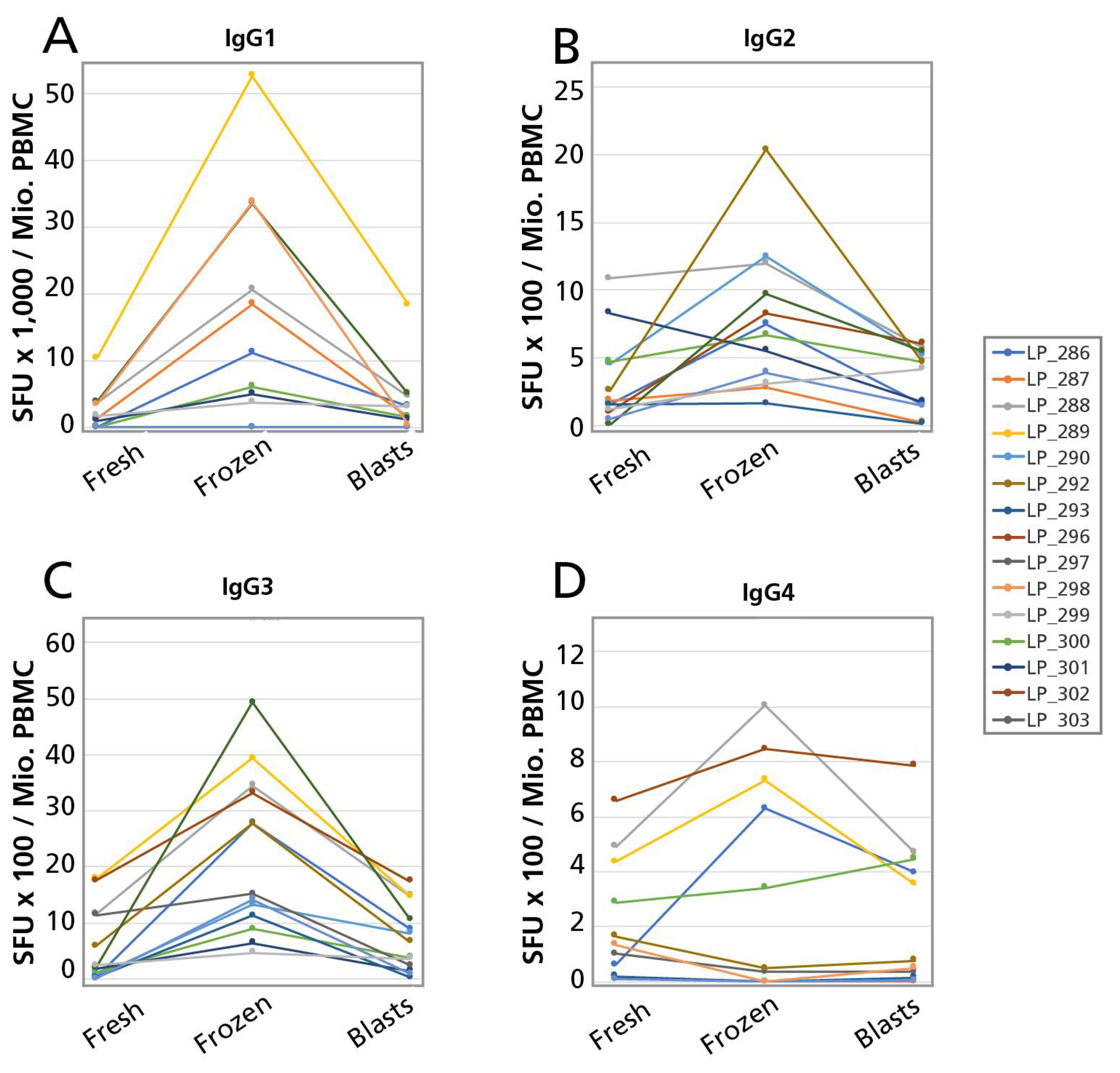
B cells secrete antibodies that fall within different Ig classes, namely, IgM, IgG, IgA, and IgE, of which IgG is the most abundant in human serum. While these classes show high levels of conservation in the primary amino acid sequence, they carry key differences in their discordant Fc-regions, which contribute to their ability to bind antigen polyvalently, different tissue distributions, half-life, and overall effector functions. Functionally, IgM is associated with the initial response of naïve B cells to antigen, the IgA class protects mucosal surfaces and IgE triggers degranulation of mast cells. The bulk of the adaptive immune response to pathogens, however, is mediated by IgG which in humans occur in four subclasses: IgG1; IgG2; IgG3; and IgG4. Because each of these Ig classes and subclasses convey different effector functions it is important for immune monitoring purposes to accurately define the numbers of B cells programmed to secrete exclusively each of them (individual B cells can produce Ig molecules belonging to one class or subclass only). The original B cell ELISPOT assays were done in single color, permitting only the detection of one antibody class or subclass per test. For comprehensive immune monitoring, therefore, seven single color B cell ELISPOT assays would need to be run in parallel (if pan IgG is not measured as such, assessing the four IgG subclasses provides that information). We describe here a four color B cell ELISPOT assay that operates with the same sensitivity as single color assays, yet require one fourth of the cells, reagents, and labor. After validating this four color B cell ImmunoSpot® assay, we set out to apply it to test whether PBMC can be cryopreserved before enumerating active ASC, and those reactivated by polyclonal stimulation.
Conducting immune monitoring trials based on antibody measurements in serum is rather straightforward. After the blood has been drawn, antibody molecules are stable in serum for years, and thus shipment from clinical sites to central testing laboratories does not require complex logistics, nor does the storage of the samples require sophisticated facilities and protocols to permit batch testing. In this way, testing can be done cost and labor efficiently, involving assay runs with large numbers of samples. Also, due to the stability of antibodies, the serum samples can be retested an indefinite number of times thereby facilitating not only assay optimization, development, qualification, and validation, but also the re-testing of clinical samples that have provided ambiguous results, or sharing of samples between different test facilities. In contrast, testing live PBMC for immune monitoring purposes require that the cells are either tested upon their isolation from blood, or cryopreserved in a way that they maintain their functionality upon thawing. Without the ability to cryopreserve samples effectively, clinical trials involving PBMC are close to impossible to conduct.
For decades, it was thought that PBMC could not be cryopreserved without substantial loss in function. In the meantime, protocols have been introduced that—at least for T cells—are suited to maintain the (T) cells’ functionality [
21]. Loss-free cryopreservation of PBMC has permitted the development of reference sample strategies [
22], and enabled the test sample logistics for conducting large scale clinical trials under GLP requirements [
23]. The key to loss-free cryopreservation of PBMC lays in the understanding that it is not primarily DMSO’s toxicity, but its osmotic activity that harms the cells during the cryopreservation and thawing process [
21]. According to the original protocols, ice cold DMSO is added to cells that are chilled on ice. In the chilled state, cells are metabolically inactive, however, and thus cannot actively compensate for the osmotic pressure caused by the addition of DMSO, resulting in lysis or damage to the cells. Similarly, when the cells are thawed, the original protocol requires that ice chilled washing medium is rapidly added to the thawing cells when the “last ice crystal” is still visible, that is, at a time when the cells are still chilled and metabolically inactive. Under these conditions again, the cells cannot actively compensate for the rapid change in osmotic pressure as the concentration of DMSO drops from 10% to essentially zero in seconds as the ice cold washing medium is added rapidly—again, the cells lyse or become damaged. In the new protocol, warm DMSO is slowly added to warm PBMC during the cryopreservation protocol, and when thawing, the cells are brought to 37 °C before warm washing medium is added slowly, permitting the cells to actively compensate for the change in osmolarity as the concentration of DMSO gradually changes. Using the latter protocol, measuring T cell functionality, no difference can be seen when testing fresh versus cryopreserved PBMC [
21]. While this notion is widely accepted and utilized in the meantime for T cell measurements, it is still controversial whether B cells too can be cryopreserved without loss of function. We have addressed here this question for B cells, utilizing the protocol that has proven successful for T cells.
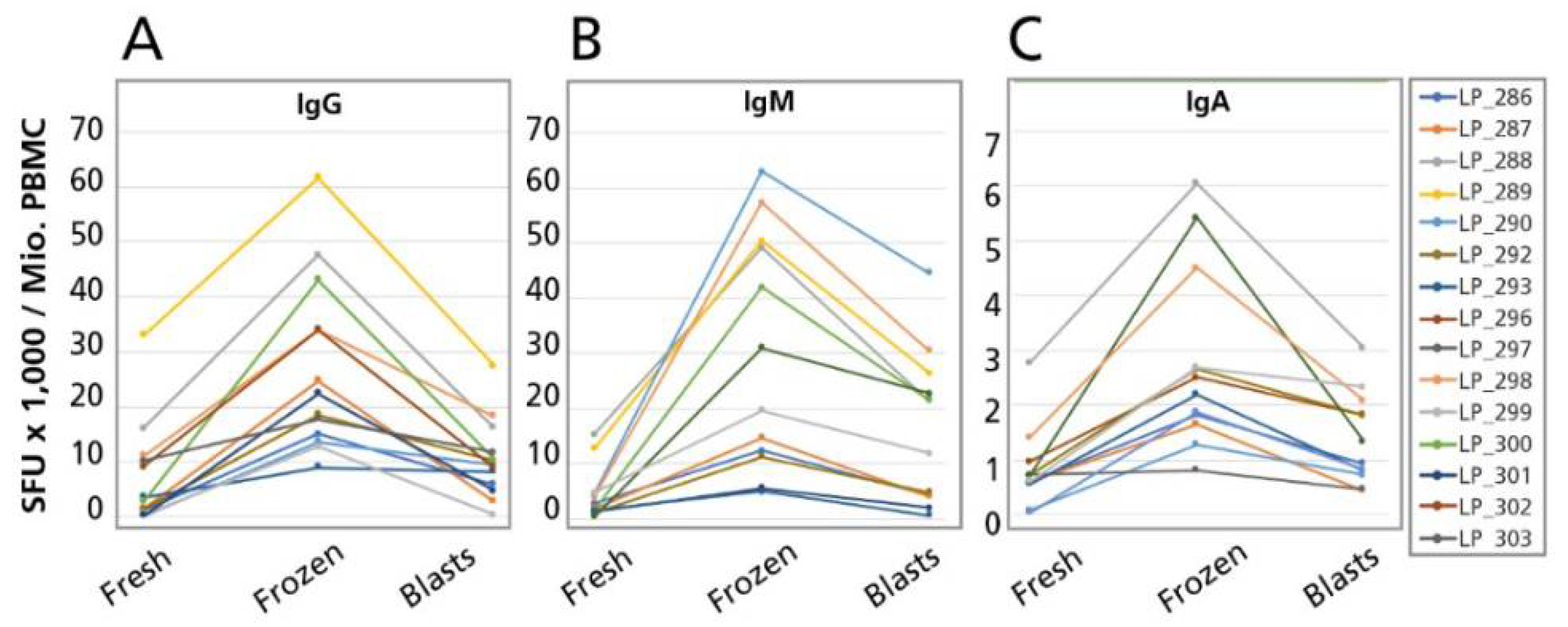
When cryopreserving B cells, one needs to distinguish between two fundamentally different activation states of these cells. Memory B cells are resting lymphocytes (with a rudimentary cytoplasm) that operate at minimal metabolic activity: as described above, such resting memory B cells can be detected only in Indirect B Cell ELISPOT Assays after they have been activated in vitro through a four day polyclonal stimulation culture to become B cell blasts that secrete antibody. Successful cryopreservation of resting memory B cells therefore requires that, after thawing, these cells can proliferate and differentiate to become ASC.
B cell blasts (ASC), in contrast, are highly activated lymphocytes that actively synthesize and secrete antibody molecules (and because of their high protein-synthesis activity they have an enlarged cytoplasm with a highly developed ER, which inspired them being named plasma cells, and plasma blasts). As they constitutively secrete antibody without the need for additional stimulation, they can be detected in Direct B Cell ELISPOT Assays. As B cell blasts are present in ex vivo PBMC only during ongoing infections, or shortly after vaccinations, they can rarely be detected in unstimulated PBMC of healthy donors. The PBMC library we tested was from healthy donors in whom we could not detect spontaneous ASC using the Direct B Cell ELISPOT Assay approach. Therefore, to be able to test whether B cell blasts can be frozen without loss of function, we cryopreserved PBMC at the end of the four day polyclonal stimulation culture, when B cell blasts/ASC are abundant, and compared the ASC numbers before and after thawing these cells (See
Figure S1).
2. Materials and Methods
2.1. Human Subjects and PBMC
All 15 human subjects tested in this study were healthy adults ages 22–45 and were recruited as a part of larger PBMC banking involving over 298 individuals at Cellular Technology Ltd. (CTL, Cleveland, OH, USA). Blood donors were recruited by Hemacare (Van Nuys, CA, USA) under Hemacare IRB where the PBMC were isolated by leukopheresis. The PBMC were either tested without cryopreservation upon receipt of the cells as “fresh cells”, or were cryopreserved as described below and stored in vapor phase liquid nitrogen until testing in ELISPOT assays as “thawed cells”. The PBMC had been stored between 2 weeks and 13 years before testing, during which, at least as far as T cell functionality goes, no change was seen upon repeated testing of aliquots over time. Detailed methods of PBMC thawing, washing, and counting of the cryopreserved PBMC have been previously described [
24]. PBMC were transferred to polyclonal B-cell stimulation cultures within 2 h of thawing for subsequent use in ELISPOT assays without involving “overnight resting”, as we did not find that to be of any advantage [
25].
2.2. Polyclonal B-Cell Stimulation
B cells do not spontaneously secrete antibody. Therefor their detection requires polyclonal stimulation for several days, during which resting memory B cells differentiate into ASC that can be detected in B cell ELISPOT assays [
20]. For polyclonal stimulation, fresh, or freshly thawed PBMC were resuspended in CTL-Test B™-media (CTLTB-010, CTL, Cleveland, OH, USA) supplemented with polyclonal B cell stimulator (CTL-BPOLY200, CTL, which contains R848 and human IL-2 and is part of the ImmunoSpot
® kits) according to the manufacturer’s instructions. The PBMC were at 4 million cells/mL, in 25 cm
2 tissue culture flasks and were cultured in an incubator at 37 °C, 5% CO
2 for four days.
2.3. Cryopreservtion of PBMC
For cryopreservation of PBMC osmotic pressure caused by DMSO (more than DMSO’s intrinsic toxicity) is one of the primary factors that needs to be controlled. Therefore, all reagents should be used at 37 °C. Freezing followed CTL’s protocols and using CTL’s reagents, as follows. In preparation, CTL-Cryo™ A was mixed with CTL-Cryo™ B in an 80% to 20% (v/v) ratio (4:1), by slowly adding CTL-Cryo™ B into CTL-Cryo™ A. The resulting CTL-Cryo™ A-B mixture and CTL-Cryo™ C were warmed up to 37 °C in a CO2 incubator. The PBMC to be cryopreserved were resuspended PBMC in warm CTL-Cryo™ C medium adjusting the cell concentration to 20 × 106 cells/mL, and also placed for 20 min into the incubator at 37 °C. After these preparations, the admixing of the warmed-up cells and the warmed up freezing solution began. The cells were resuspended by gently tapping the tube and slowly, over a time period of approximately two minutes, an equal volume of warm Cryo™ A + B was added, drop-by-drop while gently whirling the tube to ensure complete mixing of the two solutions. Next, the cells were aliquoted into pre-labeled cryovials while pipetting them gently and slowly to minimize shear forces. Next, the cryovials were placed into a room temperature Nalgene® cryofreezing container (Mr. Frosty, Thermo Scientific, Watham, MA, USA) filled with propanol and transferred into a −80 °C freezer for 24 h, after which the vials were transferred into vapor/liquid nitrogen tanks for long term storage.
The thawing of PBMC also followed CTL protocols using CTL reagents. On the day of the experiment, the cryovials were pulled from the liquid nitrogen tanks, and transported on dry ice to the laboratory, where they were immediately placed into a 37 °C water bath for 10 min. During this time, the cells reached 37 °C temperature. Next, the cells were aspirated from the cryovial, pipetting slowly and avoiding shear forces, and transferred into a 50 mL conical tube. To recover the residual cells from the cryovial, 1 mL of 37 °C warm CTL Anti-Aggregate Wash™ Medium was added to each cryovial, aspirated, and added slowly to the rest of the cells. This resulted in the first 1 + 1 dilution of DMSO. Slowly, additional 37 °C warm CTL Anti-Aggregate Wash™ Medium was added to the 50 mL tube while gently swirling the tube: the first 3 mL were added over one minute, followed by 1 mL over five seconds until the cells were resuspended in 10 mL, and thus DMSO has been diluted 1 in 10. Additional 20 mL of warm CTL Anti- Aggregate Wash™ Medium was added over 1 min. The cells were now spun in at 330 g for 10 min at room temperature, with rapid acceleration and the brake on. After decant the supernatant, the pellet was carefully resuspended by tapping the tube (avoid pipetting or vortexing). Ten ml of 37 °C CTL Anti-Aggregate Wash™ Medium was added, then the cells were mixed by inverting the tube twice 180° with the cap tightly closed. A cell sample was taken for cell counting by the CTL’s Live/Dead/Apoptotic cell counting suite using a CTL S6 Ultimate Analyzer (by CTL, Cleveland, OH, USA). After one more washing step using warm CTL Anti-Aggregate Wash Medium, the cells were adjusted to the desired cell concentration in warm CTL-Test™ Medium, and stored in the CO2 incubator until plated into the assays.
2.4. B-Cell ELISPOT Assays
After four days of polyclonal stimulation, the cells were counted with CTL’s Live/Dead/Apoptotic cell counting suite using a CTL S6 Ultimate Analyzer. After washing the cells one time, the cells were adjusted to 2.5 × 106 cells/mL, of which the specified number of cells was plated, in serial dilutions and in quadruplicates, into 96-well plates that were pre-coated with anti-κ/λ capture antibody contained in the kit. Human Four-Color ImmunoSpot® IgM/IgG/IgA/IgE, Three-Color Human ImmunoSpot® IgM/IgG/IgA, and Human Four-Color ImmunoSpot® IgG1/IgG2/IgG3/IgG4 assay kits were used, as specified, all from CTL. The cells were incubated for a period of 24 h at 37 °C, 5% CO2 during which the antibodies that the B cells secreted were captured on the membrane by the capture antibody directly around the secreting cells. Thereafter, the plates were decanted and washed three times with 200 µL sterile PBS. The plate-bound Ig “spots”, each representing the secretory foot print of a single ASC, were visualized using the four anti-Ig class/subclass-specific detection antibodies contained in the kits, following the manufacturer’s specifications.
2.5. Plate Reading and Statistical Analysis
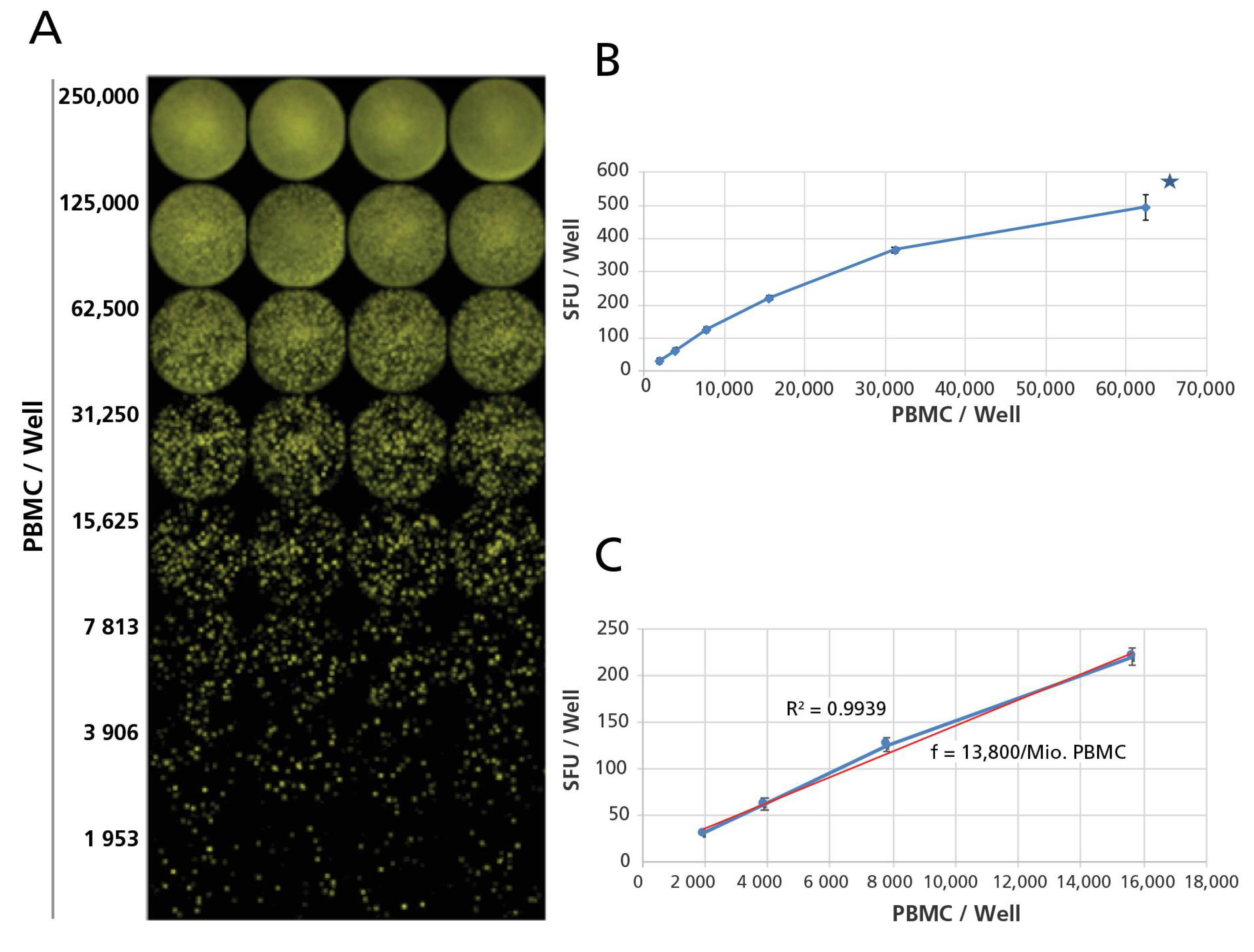
Following the completion of the B cell ELISPOT assay, the plates were air-dried in a laminar flow hood prior to being scanned and counted on an ImmunoSpot® S6 Ultimate Reader. The numbers of B cell Spot Forming Units (SFU), each corresponding to the secretory foot print of an ASC, were established using the BasicCount™-Mode of the ImmunoSpot® Software. Means and standard deviations were calculated for the quadruplicate wells, tested for each test condition. For each PBMC donor, class and subclass, and for fresh, thawed and blast cells of each, the linearity of spot counts was established, based on which the SFU counts were extrapolated to SFU/million using standard functions of Excel. The Student’s t-test, or the ANOVA test were performed to detect statistical differences between two or three groups of Spot Forming Unit (SFU) counts, respectively.
4. Conclusions
The main goal of B cell immune monitoring is to establish: (a) whether spontaneously antibody secreting B cell blasts are present in PBMC at their isolation, detectable in Direct ELISPOT assays; (b) whether clonally expanded memory B cell populations are present in the test individual; and (c) what type of antibody classes/subclasses these memory B cells secrete upon re-stimulation. Testing polyclonally stimulated B cells, our data show that spontaneously antibody secreting B cell plasma blasts can be detected in cryopreserved PBMC, although their numbers will likely be reduced two to three-fold. In spite of this reduction in numbers, the mere presence of spontaneously secreting ASC in cryopreserved PBMC per se cells should reveal actively ongoing immune responses because active ASC are absent in blood in a quiescent immunological state. Clonally expanded antigen-specific B cell memory populations should also be readily detectable in cryopreserved PBMC because in naïve individuals no antigen-specific B cells can be detected even after polyclonal stimulation. Therefore, in spite of the increased frequencies of ASC detected in cryopreserved PBMC, detectable numbers of ASC per se identify that B memory cells existed in vivo in clonally expanded sizes. Moreover, recently primed B cells can only secrete IgM; it takes Ig class switching in the course of an immune response in vivo to generate (memory) B cells capable of secreting any antibody class other than IgM. Therefore, detecting antigen-specific B cells in PBMC that secrete different antibody classes/subclasses in ELISPOT assays suggests that these B cells have undergone class switching in vivo. Thus, these data suggest that qualitative markers of B cell immunity are maintained in cryopreserved PBMC. This notion was confirmed in a recent study of HCMV-specific T and B memory cells [
27]. Testing cryopreserved PBMC of 82 healthy human donors we found that most individuals who possessed HCMV-specific CD4- and CD8 memory T cells in increased frequencies also possessed HCMV-specific B cells in increased frequencies. Notably, all these B cells secreted IgG1 antibody, the Ig subclass that prevails in sera of subjects with latent HCMV infection [
28]. In contrast, in test subjects in whose cryopreserved PBMC HCMV-specific memory T cells were undetectable, B cells producing HCMV-specific antibodies were also absent, and this finding held up for all Ig classes and subclasses [
27]. In this study, we tested for antigen-specific ASC, that is, by coating the membrane with the antigen itself. An elegant alternative approach for detecting antigen-specific ASC has been introduced by Jahnmatz et al. [
29] in which first all antibodies secreted by ASC are captured on the membrane and the fluorescence labelled antigen is added to visualize the antigen-specific ASC. Our data predict that using either approach the moderate increase of ASC numbers in cryopreserved PBMC should be rather predictable and proportional among donors. If a donor had low SFU counts relative to another donor in the fresh cells, that relationship is maintained proportionally in the cryopreserved cells. This notion represents a major advancement for the perspective of B cell immune monitoring with cryopreserved PBMC compared to the alternative that it can be done reliably with fresh PBMC only. The use of cryopreserved PBMC enables cost-effective, large scale immune monitoring that can be adapted to regulated work, including developing reference PBMC strategies.
Presently, it is unclear why the ASC numbers increase in cryopreserved cells. One possibility is that B cells survive cryopreservation better than other cell populations in PBMC. If such a difference exists, it is not detectable right after thawing of the cells at which time points we compared the fresh vs. freeze-thawed cells (data not shown). Thus, the difference in ASC numbers manifests itself during the four days polyclonal stimulation culture. During these four days, responding to polyclonal stimulation, the resting B cells need to undergo blast transformation and to activate their Ig synthesis machinery. In addition, these B cells will engage in proliferation. All these cellular events are under the influence of a microenvironment defined by cytokines and cell surface molecules contributed of bystander cells present in the PBMC. Thus, the increased ASC numbers after cryopreservation could either result from a direct effect on the B cells, a possibility that these authors consider more unlikely, or could occur as a consequence of affecting third party cells, e.g., cells of the innate immune system that, like the B cells themselves, also become activated by the TLR agonist R848 and IL-2, and apparently responded to these stimuli differently in freeze thawed PBMC, thus providing a modified (more co-stimulatory) microenvironment for B cell proliferation and differentiation than do third party PBMC in freshly isolated PBMC. Another -yet related possibility is that regulatory cells do not withstand freeze thawing, and thus do not exert an inhibitory effect in the cryopreserved PBMC samples.


 and
and 






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}