Unfolding the Endoplasmic Reticulum of a Social Amoeba: Dictyostelium discoideum as a New Model for the Study of Endoplasmic Reticulum Stress

 ,
, 

Abstract
:1. Dictyostelium as a Model Organism for Experimental Biology Research
2. The Endoplasmic Reticulum of a Social Amoeba
2.1. A Membranous Network with an Intricate Structure
2.2. The ER Is a Dynamic Structure Continuously Rearranged
2.3. A Well-Connected Membranous System
2.4. The Main Source of Lipid Synthesis
2.5. A Perfect Compartment to Fold and Modify Proteins
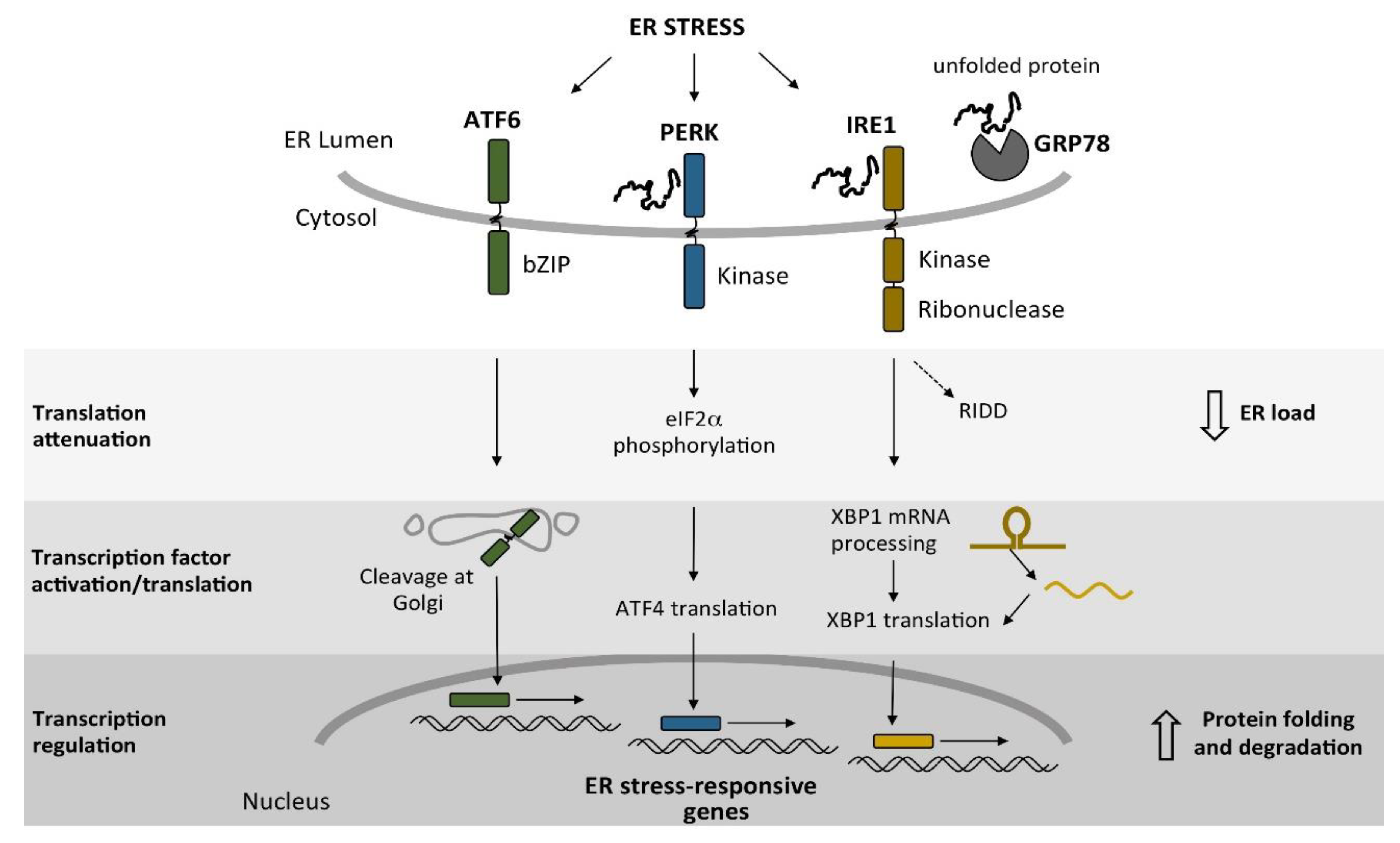
3. Endoplasmic Reticulum Stress and the Unfolded Protein Response in a Social Amoeba
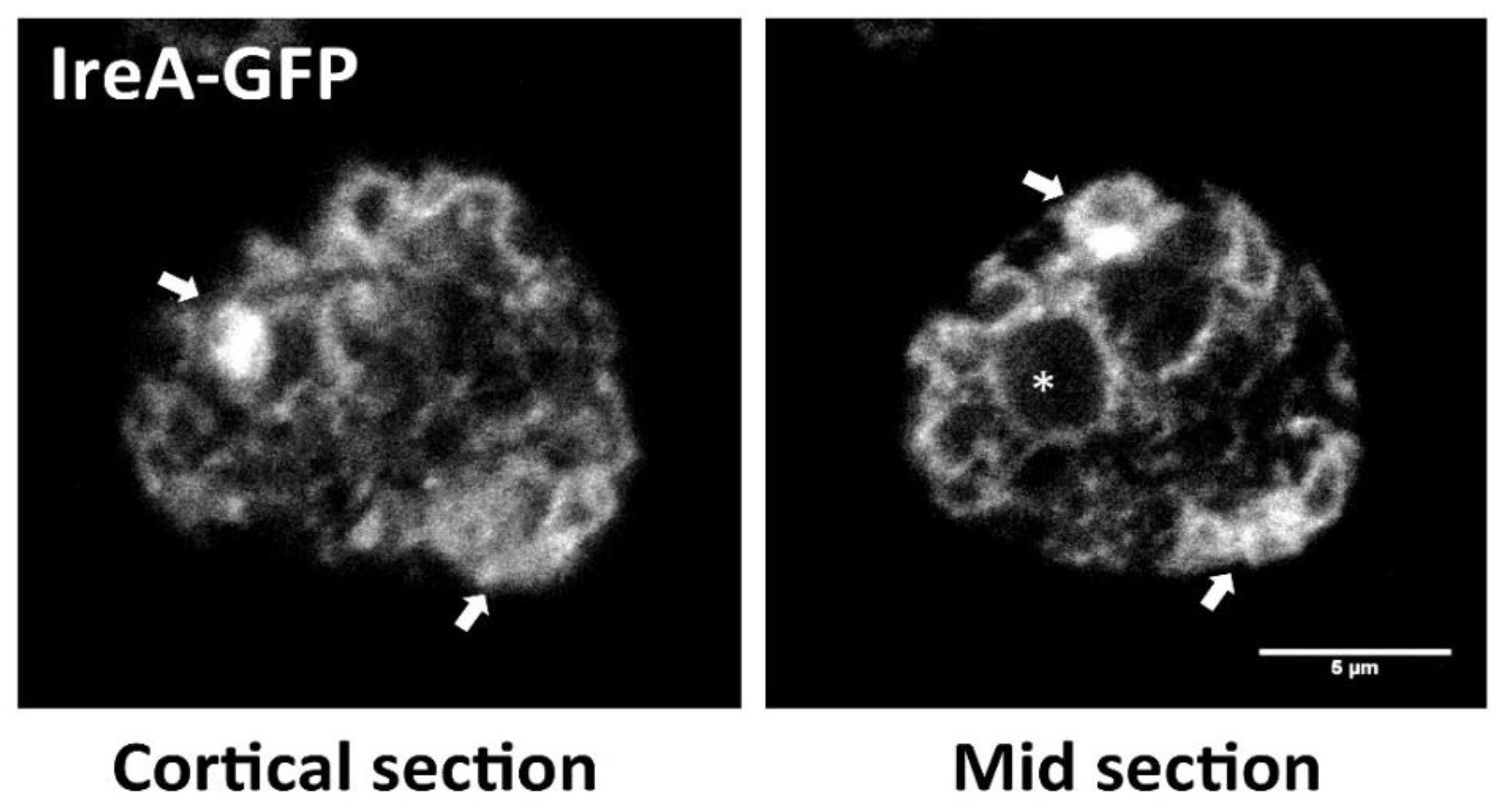
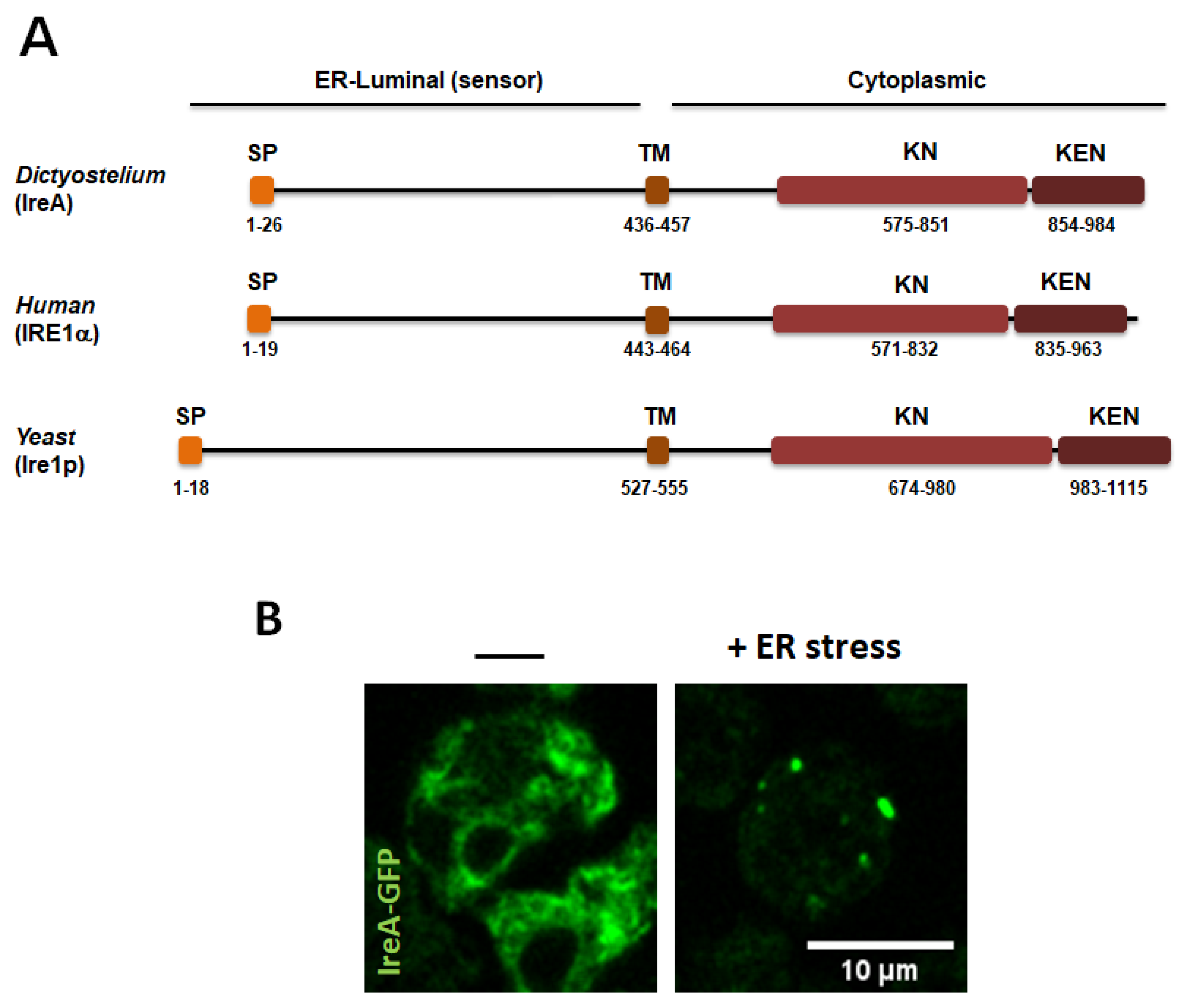
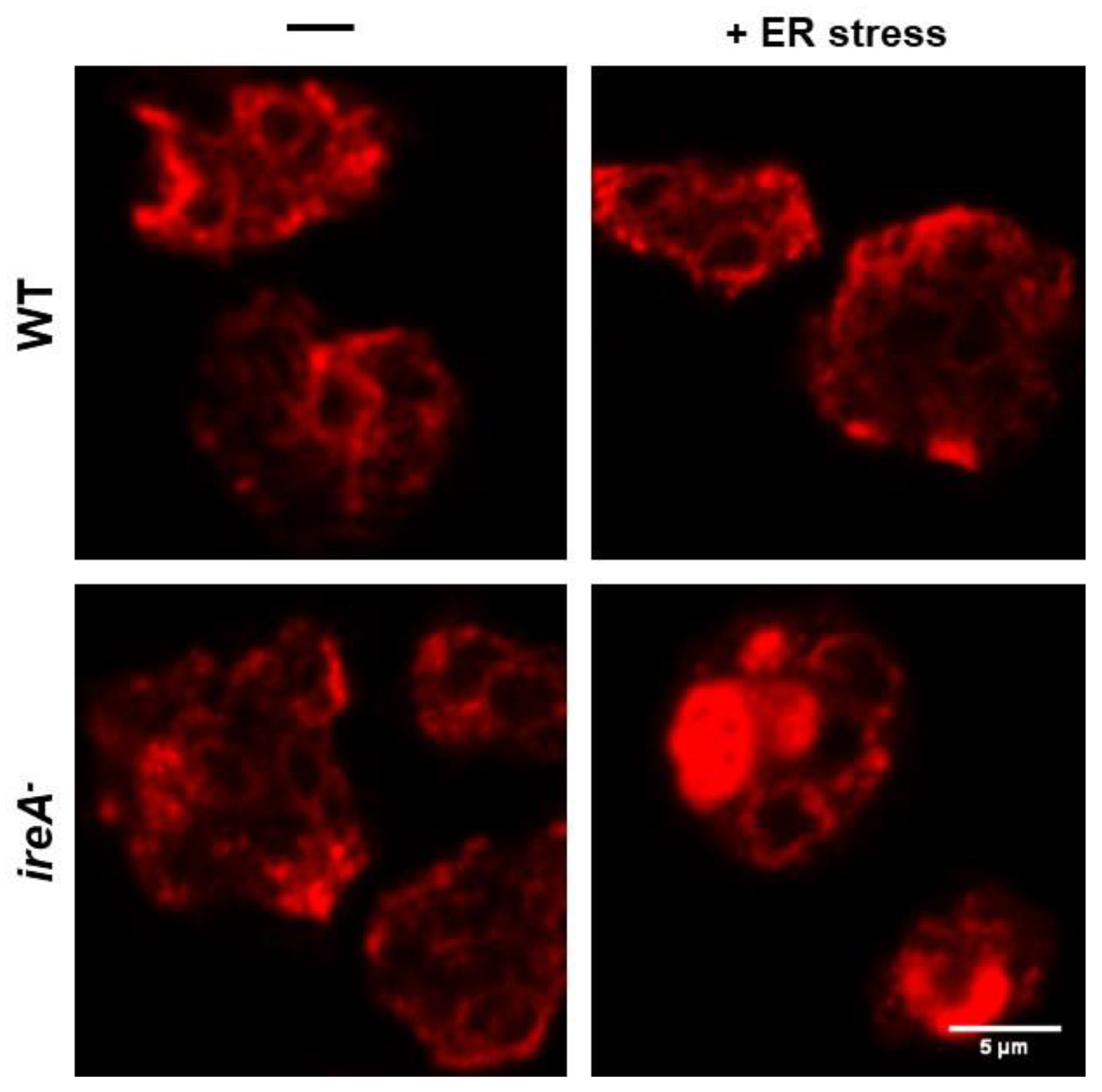
3.1. The IreA Branch
3.2. IreA-Independent UPR Pathways in Dictyostelium
4. ER Stress and the Autophagy Pathway in Dictyostelium
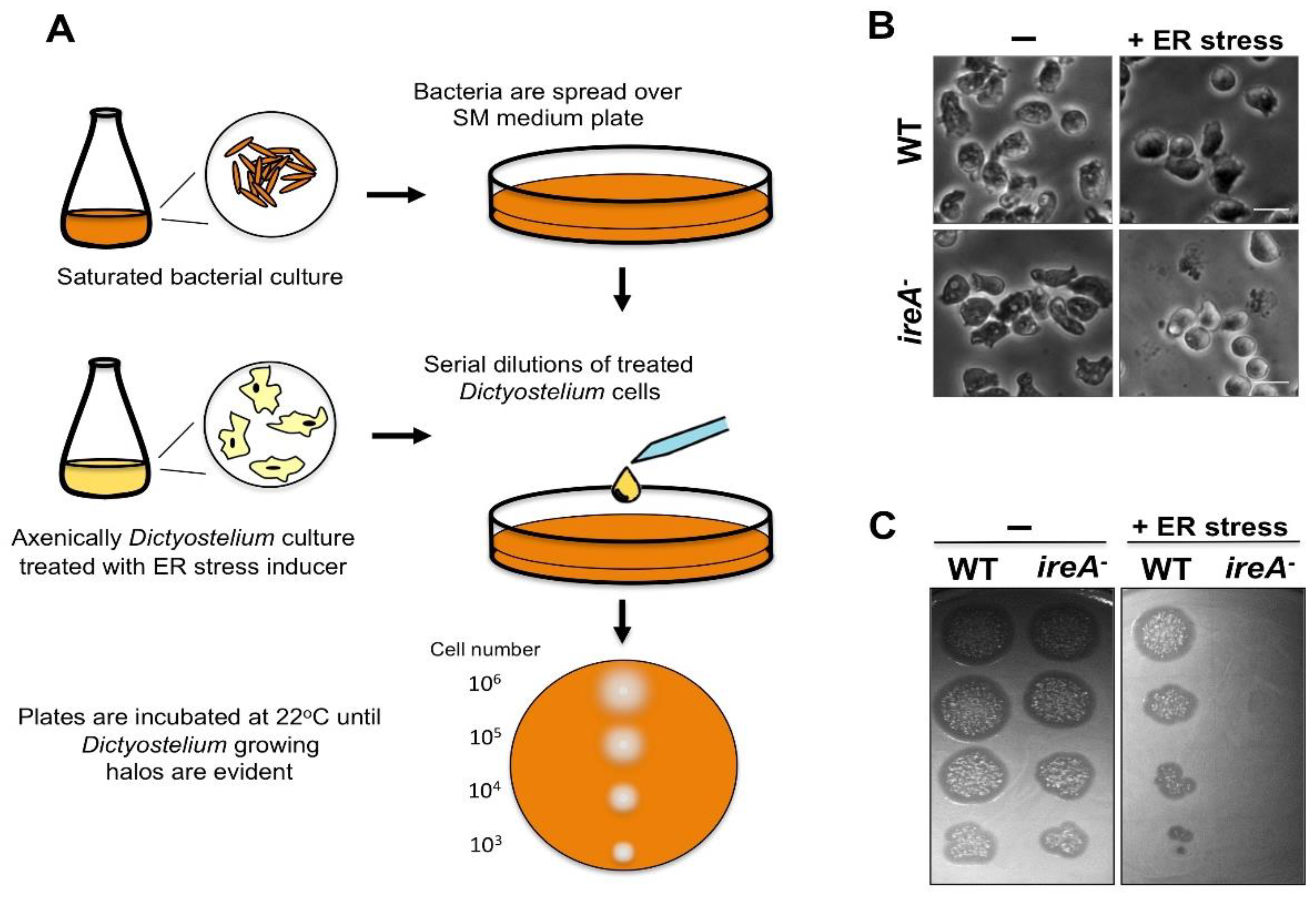
5. Assessing ER Stress in Dictyostelium
6. Concluding Remarks
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Sheikh, S.; Thulin, M.; Cavender, J.C.; Escalante, R.; Kawakami, S.; Lado, C.; Landolt, J.C.; Nanjundiah, V.; Queller, D.C.; Strassmann, J.E.; et al. A New Classification of the Dictyostelids. Protist 2018, 169, 1–28. [Google Scholar] [CrossRef] [PubMed]
- Annesley, S.; Fisher, P. Dictyostelium discoideum—A model for many reasons. Mol. Cell. Biochem. 2009, 329, 73–91. [Google Scholar] [CrossRef] [PubMed]
- Muñoz-Braceras, S.; Mesquita, A.; Escalante, R. Dictyostelium discoideum as a model in biomedical research. In Dictyostelids. Evolution, Genomics and Cell Biology; Romeralo, M., Baldauf, S., Escalante, R., Eds.; Springer: Berlin/Heiderberg, Germany, 2013; pp. 1–34. ISBN 978-3-642-38486-8. [Google Scholar]
- Loomis, W.F. Cell signaling during development of Dictyostelium. Dev. Biol. 2014, 391, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Eichinger, L.; Pachebat, J.A.; Glockner, G.; Rajandream, M.-A.; Sucgang, R.; Berriman, M.; Song, J.; Olsen, R.; Szafranski, K.; Xu, Q.; et al. The genome of the social amoeba Dictyostelium discoideum. Nature 2005, 435, 43–57. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Williams, J.G. Dictyostelium finds new roles to model. Genetics 2010, 185, 717–726. [Google Scholar] [CrossRef] [PubMed]
- Domínguez-Martín, E.; Ongay-Larios, L.; Kawasaki, L.; Vincent, O.; Coello, G.; Coria, R.; Escalante, R. IreA controls endoplasmic reticulum stress-induced autophagy and survival through homeostasis recovery. Mol. Cell. Biol. 2018, MCB.00054-18. [Google Scholar] [CrossRef]
- Roussel, B.D.; Kruppa, A.J.; Miranda, E.; Crowther, D.C.; Lomas, D.A.; Marciniak, J.S. Endoplasmic reticulum dysfunction in neurological disease. Lancet Neurol. 2013, 12, 105–118. [Google Scholar] [CrossRef] [Green Version]
- Ozcan, L.; Tabas, I. Role of Endoplasmic Reticulum Stress in Metabolic Disease and Other Disorders. Annu. Rev. Med. 2012, 63, 317–328. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Annesley, S.J.; Chen, S.; Francione, L.M.; Sanislav, O.; Chavan, A.J.; Farah, C.; De Piazza, S.W.; Storey, C. L.; Ilievska, J.; Fernando, S.G.; et al. Dictyostelium, a microbial model for brain disease. Biochim. Biophys. Acta Gen. Subj. 2014, 1840, 1413–1432. [Google Scholar] [CrossRef] [PubMed]
- Huber, R.J. Using the social amoeba Dictyostelium to study the functions of proteins linked to neuronal ceroid lipofuscinosis. J. Biomed. Sci. 2016, 23, 83. [Google Scholar] [CrossRef] [PubMed]
- Myre, M.A.; Huber, R.J.; Day, D.H.O. Functional Analysis of Proteins Involved in Neurodegeneration Using the Model Organism Dictyostelium : Alzheimer’s, Huntington’s, and Batten Disease; Elsevier Inc.: New York, NY, USA, 2018; ISBN 9780128040782. [Google Scholar]
- Malinovska, L.; Palm, S.; Gibson, K.; Verbavatz, J.-M.; Alberti, S. Dictyostelium discoideum has a highly Q/N-rich proteome and shows an unusual resilience to protein aggregation. Proc. Natl. Acad. Sci. USA 2015, 112, E2620–E2629. [Google Scholar] [CrossRef] [PubMed]
- Santarriaga, S.; Petersen, A.; Ndukwe, K.; Brandt, A.; Gerges, N.; Scaglione, J.B.; Scaglione, K.M. The social amoeba Dictyostelium discoideum is highly resistant to polyglutamine aggregation. J. Biol. Chem. 2015, 290, 25571–25578. [Google Scholar] [CrossRef] [PubMed]
- Westrate, L.M.; Lee, J.E.; Prinz, W.A.; Voeltz, G.K. Form Follows Function: The Importance of Endoplasmic Reticulum Shape. Annu. Rev. Biochem. 2015, 84, 791–811. [Google Scholar] [CrossRef] [PubMed]
- Nixon-Abell, J.; Obara, C.J.; Weigel, A.V.; Li, D.; Legant, W.R.; Xu, C.S.; Pasolli, H.A.; Harvey, K.; Hess, H.F.; Betzig, E.; et al. Increased spatiotemporal resolution reveals highly dynamic dense tubular matrices in the peripheral ER. Science 2016, 354, aaf3928. [Google Scholar] [CrossRef] [PubMed]
- Fernández-Busnadiego, R.; Saheki, Y.; de Camilli, P. Three-dimensional architecture of extended synaptotagmin-mediated endoplasmic reticulum—Plasma membrane contact sites. Proc. Natl. Acad. Sci. USA 2015, 112, E2004–E2013. [Google Scholar] [CrossRef] [PubMed]
- Hetzer, M.W. The nuclear envelope. Cold Spring Harb. Perspect. Biol. 2010. [Google Scholar] [CrossRef] [PubMed]
- Terasaki, M.; Shemesh, T.; Kasthuri, N.; Klemm, R.W.; Schalek, R.; Hayworth, K.J.; Hand, A.R.; Yankova, M.; Huber, G.; Lichtman, J.W.; et al. Stacked endoplasmic reticulum sheets are connected by helicoidal membrane motifs. Cell 2013, 154, 285–296. [Google Scholar] [CrossRef] [PubMed]
- Zurek, N.; Sparks, L.; Voeltz, G. Reticulon short hairpin transmembrane domains are used to shape ER tubules. Traffic 2011, 12, 28–41. [Google Scholar] [CrossRef] [PubMed]
- di Sano, F.; Bernardoni, P.; Piacentini, M. The reticulons: Guardians of the structure and function of the endoplasmic reticulum. Exp. Cell Res. 2012, 318, 1201–1207. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shibata, Y.; Voss, C.; Rist, J.M.; Hu, J.; Rapoport, T.A.; Prinz, W.A.; Voeltz, G.K. The reticulon and Dp1/Yop1p proteins form immobile oligomers in the tubular endoplasmic reticulum. J. Biol. Chem. 2008, 283, 18892–18904. [Google Scholar] [CrossRef] [PubMed]
- Oertle, T.; Klinger, M.; Stuermer, C.A.O.; Schwab, M.E. A reticular rhapsody: Phylogenic evolution and nomenclature of the RTN/Nogo gene family. FASEB J. 2003, 17, 1238–1247. [Google Scholar] [CrossRef] [PubMed]
- Chiurchiù, V.; Maccarrone, M.; Orlacchio, A. The role of reticulons in neurodegenerative diseases. NeuroMol. Med. 2014, 16, 3–15. [Google Scholar] [CrossRef] [PubMed]
- Schwarz, D.S.; Blower, M.D. The endoplasmic reticulum: Structure, function and response to cellular signaling. Cell. Mol. Life Sci. 2016, 73, 79–94. [Google Scholar] [CrossRef] [PubMed]
- Bola, B.; Allan, V. How and why does the endoplasmic reticulum move? Biochem. Soc. Trans. 2009, 37 Pt 5, 961–965. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fehrenbacher, K.L.; Davis, D.; Wu, M.; Boldogh, I.; Pon, L.A. Endoplasmic reticulum dynamics, inheritance, and cytoskeletal interactions in budding yeast. Mol. Biol. Cell 2002, 13, 854–865. [Google Scholar] [CrossRef] [PubMed]
- Boevink, P.; Oparka, K.; Cruz, S.S.; Martin, B.; Betteridge, A.; Hawes, C. Stacks on tracks: The plant Golgi apparatus traffics on an actin/ER network. Plant J. 1998, 15, 441–447. [Google Scholar] [CrossRef] [PubMed]
- Liu, T.Y.; Bian, X.; Sun, S.; Hu, X.; Klemm, R.W.; Prinz, W.A.; Rapoport, T.A.; Hu, J. Lipid interaction of the C terminus and association of the transmembrane segments facilitate atlastin-mediated homotypic endoplasmic reticulum fusion. Proc. Natl. Acad. Sci. USA 2012, 109, E2146–E2154. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McNew, J.A.; Sondermann, H.; Lee, T.; Stern, M.; Brandizzi, F. GTP-Dependent Membrane Fusion. Annu. Rev. Cell Dev. Biol. 2013, 29, 529–550. [Google Scholar] [CrossRef] [PubMed]
- Bian, X.; Klemm, R.W.; Liu, T.Y.; Zhang, M.; Sun, S.; Sui, X.; Liu, X.; Rapoport, T.A.; Hu, J. Structures of the atlastin GTPase provide insight into homotypic fusion of endoplasmic reticulum membranes. Proc. Natl. Acad. Sci. USA 2011, 108, 3976–3981. [Google Scholar] [CrossRef] [PubMed]
- Byrnes, L.J.; Sondermann, H. Structural basis for the nucleotide-dependent dimerization of the large G protein atlastin-1/SPG3A. Proc. Natl. Acad. Sci. USA 2011, 108, 2216–2221. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Lockwood, S.K.; Hoeltzel, M.F.; Schiefelbein, J.W. The Root Hair Defective3 gene encodes an evolutionarily conserved protein with GTP-binding motifs and is required for regulated cell enlargement in Arabidopsis. Genes Dev. 1997, 11, 799–811. [Google Scholar] [CrossRef] [PubMed]
- Hu, J.; Shibata, Y.; Zhu, P.P.; Voss, C.; Rismanchi, N.; Prinz, W.A.; Rapoport, T.A.; Blackstone, C. A Class of Dynamin-like GTPases Involved in the Generation of the Tubular ER Network. Cell 2009, 138, 549–561. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Steiner, B.; Swart, A.L.; Welin, A.; Weber, S.; Personnic, N.; Kaech, A.; Freyre, C.; Ziegler, U.; Klemm, R. W.; Hilbi, H. ER remodeling by the large GTPase atlastin promotes vacuolar growth of Legionella pneumophila. EMBO Rep. 2017, 18, 1–20. [Google Scholar] [CrossRef] [PubMed]
- Phillips, M.J.; Voeltz, G.K. Structure and function of ER membrane contact sites with other organelles. Nat. Rev. Mol. Cell Biol. 2016, 17, 69–82. [Google Scholar] [CrossRef] [PubMed]
- Wideman, J.G.; Muñoz-Gómez, S.A. The evolution of ERMIONE in mitochondrial biogenesis and lipid homeostasis: An evolutionary view from comparative cell biology. Biochim. Biophys. Acta Mol. Cell Biol. Lipids 2016, 1861, 900–912. [Google Scholar] [CrossRef] [PubMed]
- Kornmann, B.; Currie, E.; Collins, S.R.; Schuldiner, M.; Nunnari, J.; Weissman, J.S.; Walter, P. An ER-mitochondria tethering complex revealed by a synthetic biology screen. Science 2009, 325, 477–481. [Google Scholar] [CrossRef] [PubMed]
- Tabara, L.C.; Escalante, R. VMP1 establishes ER-microdomains that regulate membrane contact sites and autophagy. PLoS ONE 2016, 11, 18892–18904. [Google Scholar] [CrossRef] [PubMed]
- Herrera-Cruz, M.S.; Simmen, T. Of yeast, mice and men: MAMs come in two flavors. Biol. Direct 2017, 12, 1–21. [Google Scholar] [CrossRef] [PubMed]
- AhYoung, A.P.; Lu, B.; Cascio, D.; Egea, P.F. Crystal structure of Mdm12 and combinatorial reconstitution of Mdm12/Mmm1 ERMES complexes for structural studies. Biochem. Biophys. Res. Commun. 2017, 488, 129–135. [Google Scholar] [CrossRef] [PubMed]
- Fey, P.; Gaudet, P.; Pilcher, K.E.; Franke, J.; Chisholm, R.L. dictyBase and the Dicty Stock Center. Methods Mol. Biol. 2006, 346, 51–74. [Google Scholar] [PubMed]
- Calvo-garrido, J.; Carilla-latorre, S.; La, F.; Egea, G.; Escalante, R. Vacuole Membrane Protein 1 Is an Endoplasmic Reticulum Protein Required for Organelle Biogenesis, Protein Secretion, and Development. Mol. Biol. Cell. 2008, 19, 3442–3453. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Calvo-Garrido, J.; King, J.S.; Muñoz-Braceras, S.; Escalante, R. Vmp1 regulates PtdIns3P signaling during autophagosome formation in Dictyostelium discoideum. Traffic 2014, 11, 1235–1246. [Google Scholar] [CrossRef] [PubMed]
- Fagone, P.; Jackowski, S. Membrane phospholipid synthesis and endoplasmic reticulum function. J. Lipid Res. 2009, 50, S311–S316. [Google Scholar] [CrossRef] [PubMed]
- Appenzeller-Herzog, C. The ER-Golgi intermediate compartment (ERGIC): In search of its identity and function. J. Cell Sci. 2006, 119, 2173–2183. [Google Scholar] [CrossRef] [PubMed]
- Davidoff, F.; Korn, E.D. Fatty acid and phospholipid composition of the cellular slime mold, Dictyostelium discoideum. J. Biol. Chem. 1963, 238, 3199–3209. [Google Scholar] [PubMed]
- Weeks, G.; Herring, F.G. The lipid composition and membrane fluidity of Dictyostelium discoideum plasma membranes at various stages during differentiation. J. Lipid Res. 1980, 21, 681–686. [Google Scholar] [PubMed]
- Clark, J.; Kay, R.R.; Kielkowska, A.; Niewczas, I.; Fets, L.; Oxley, D.; Stephens, L.R.; Hawkins, P.T. Dictyostelium uses ether-linked inositol phospholipids for intracellular signalling. EMBO J. 2014, 33, 2188–2200. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Du, X.; Barisch, C.; Paschke, P.; Herrfurth, C.; Bertinetti, O.; Pawolleck, N.; Otto, H.; Rühling, H.; Feussner, I.; Herberg, F.W.; et al. Dictyostelium lipid droplets host novel proteins. Eukaryot. Cell 2013, 12, 1517–1529. [Google Scholar] [CrossRef] [PubMed]
- Lu, X.; Gruia-Gray, J.; Copeland, N.G.; Gilbert, D.J.; Jenkins, N.A.; Londos, C.; Kimmel, A.R. The murine perilipin gene: The lipid droplet-associated perilipins derive from tissue-specific, mRNA splice variants and define a gene family of ancient origin. Mamm. Gen. 2001, 12, 741–749. [Google Scholar] [CrossRef]
- Du, X.; Herrfurth, C.; Gottlieb, T.; Kawelke, S.; Feussner, K.; Rühling, H.; Feussner, I.; Maniak, M. Dictyostelium discoideum Dgat2 can substitute for the essential function of Dgat1 in triglyceride production but not in ether lipid synthesis. Eukaryot. Cell 2014, 13, 517–526. [Google Scholar] [CrossRef] [PubMed]
- Xu, C.; Ng, T.W.D. Glycosylation-directed quality control of protein folding. Nat. Rev. Mol. Cell Biol. 2015, 16, 742–752. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kelleher, D.J.; Gilmore, R. An evolving view of the eukaryotic oligosaccharyltransferase. Glycobiology 2006, 16, 47R–62R. [Google Scholar] [CrossRef] [PubMed]
- Braakman, I.; Hebert, D.N. Protein folding in the endoplasmic reticulum. Cold Spring Harb. Perspect. Biol. 2013, 5, a013201. [Google Scholar] [CrossRef] [PubMed]
- Halperin, L.; Jung, J.; Michalak, M. The many functions of the endoplasmic reticulum chaperones and folding enzymes. IUBMB Life 2014, 66, 318–326. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gidalevitz, T.; Stevens, F.; Argon, Y. Orchestration of secretory protein folding by ER chaperones. Biochim. Biophys. Acta Mol. Cell Res. 2013, 1833, 2410–2424. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Müller-Taubenberger, A.; Lupas, N.; Li, H.; Ecke, M.; Simmeth, E.; Gerisch, G. Calreticulin and calnexin in the endoplasmic reticulum are important for phagocytosis. EMBO J. 2001, 20, 6772–6782. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morita, T.; Saitoh, K.; Takagi, T.; Maeda, Y. Involvement of the glucose-regulated protein 94 (Dd-GRP94) in starvation response of Dictyostelium discoideum cells. Biochem. Biophys. Res. Commun. 2016, 274, 323–331. [Google Scholar] [CrossRef] [PubMed]
- Braakman, I.; Bulleid, N.J. Protein Folding and Modification in the Mammalian Endoplasmic Reticulum. Annu. Rev. Biochem. 2011, 80, 71–99. [Google Scholar] [CrossRef] [PubMed]
- Bulleid, N.J. Disulfide Bond Formation in the Mammalian Endoplasmic Reticulum. Cold Spring Harb. Perspect. Biol. 2012, 4, a013219. [Google Scholar] [CrossRef] [PubMed]
- Monnat, J.; Hacker, U.; Geissler, H.; Rauchenberger, R.; Neuhaus, E.M.; Maniak, M.; Soldati, T. Dictyostelium discoideum protein disulfide isomerase, an endoplasmic reticulum resident enzyme lacking a KDEL-type retrieval signal. FEBS Lett. 1997, 418, 357–362. [Google Scholar] [CrossRef] [Green Version]
- Monnat, J.; Neuhaus, E.M.; Pop, M.S.; Ferrari, D.M.; Kramer, B.; Soldati, T. Identification of a Novel Saturable Endoplasmic Reticulum Localization Mechanism Mediated by the C-Terminus of a Dictyostelium Protein Disulfide Isomerase. Mol. Biol. Cell 2000, 11, 3469–3484. [Google Scholar] [CrossRef] [PubMed]
- Oslowski, C.M.; Urano, F. Measuring ER Stress and the Unfolded Protein Response Using Mammalian Tissue Culture System, 1st ed.; Elsevier Inc.: New York, NY, USA, 2011; Volume 490. [Google Scholar]
- Bakthavatsalam, D.; Gomer, R.H. The secreted proteome profile of developing Dictyostelium discoideum cells. Proteomics 2010, 10, 2556–2559. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lam, T.Y.; Siu, C.-H. Inhibition of cell differentiation and cell cohesion by tunicamycin in Dictyostelium discoideum. Dev. Biol. 1982, 92, 398–407. [Google Scholar] [CrossRef]
- Yamada, H.; Takatsuki, A.; Hirano, T.; Miyazaki, T.; Tamura, G. Effects of Tunicamycin on Cell Adhesion and Biosynthesis of of Glycoproteins in Aggregation-Competent Cells of Dictyostelium discoideum. J Biochem. 1982, 92, 399–406. [Google Scholar] [CrossRef] [PubMed]
- Ochiai, H.; Stadler, J.; Westphal, M.; Wagle, G.; Merkl, R.; Gerisch, G. Monoclonal antibodies against contact sites A of Dictyostelium discoideum: Detection of modifications of the glycoprotein in tunicamycin-treated cells. EMBO J. 1982, 1, 1011–1016. [Google Scholar] [PubMed]
- Hirano, T.; Yamada, H.; Miyazaki, T. Inhibition of cell adhesion in Dictyostelium discoideum by tunicamycin is prevented by leupeptin. J. Biochem. 1983, 93, 1249–1257. [Google Scholar] [CrossRef] [PubMed]
- McDonald, C.J.; Sampson, J. The effects of inhibition of protein glycosylation on the aggregation of Dictyostelium discoideum. J. Embryol. Exp. Morphol. 1983, 78, 229–248. [Google Scholar] [PubMed]
- Sadeghi, H.; Klein, C. Inhibition of N-linked glycosylation in Dictyostetium discoideum: Effects of aggregate formation. Differentiation 1988, 38, 99–103. [Google Scholar] [CrossRef] [PubMed]
- Frank, L.C.-L.; Chen, G.; Nicole, W.A.; Shaulsky, G. Altered N-glycosylation modulates TgrB1/TgrC1-mediated development but not allorecognition in Dictyostelium. J. Cell Sci. 2015, 128, 3990–3996. [Google Scholar] [CrossRef]
- Huber, R.J.; Myre, M.A.; Cotman, S.L. Aberrant adhesion impacts early development in a Dictyostelium model for juvenile neuronal ceroid lipofuscinosis. Cell Adhes. Migr. 2017, 11, 399–418. [Google Scholar] [CrossRef] [PubMed]
- Browning, D.D.; O’Day, D.H. Concanavalin A and wheat germ agglutinin binding glycoproteins associated with cell fusion and zygote differentiation in Dictyostelium discoideum: Effects of calcium ions and tunicamycin on glycoprotein profiles. Biochem. Cell Biol. 1991, 69, 282–290. [Google Scholar] [CrossRef] [PubMed]
- Kuwayama, H.; Ecke, M.; Gerisch, G.; VanHaastert, P.J.M. Protection against osmotic stress by cDGMP-mediated myosin phosphorylation. Science 1996, 271, 207–209. [Google Scholar] [CrossRef] [PubMed]
- Serafimidis, I.; Bloomfield, G.; Skelton, J.; Ivens, A.; Kay, R.R. A new environmentally resistant cell type from Dictyostelium. Microbiology 2007, 153, 619–630. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Travers, K.J.; Patil, C.K.; Wodicka, L.; Lockhart, D.J.; Weissman, J.S.; Walter, P. Functional and genomic analyses reveal an essential coordination between the unfolded protein response and ER-associated degradation. Cell 2000, 101, 249–258. [Google Scholar] [CrossRef]
- Hollien, J.; Weissman, J.S. Decay of Endoplasmic Reticulum-Localized mRNAs During the Unfolded Protein Response. Science 2006, 313, 104–107. [Google Scholar] [CrossRef] [PubMed]
- Hollien, J.; Lin, J.H.; Li, H.; Stevens, N.; Walter, P.; Weissman, J.S. Regulated Ire1-dependent decay of messenger RNAs in mammalian cells. J. Cell Biol. 2009, 186, 323–331. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, K.P.K.; Dey, M.; Neculai, D.; Cao, C.; Dever, T.E.; Sicheri, F. Structure of the Dual Enzyme Ire1 Reveals the Basis for Catalysis and Regulation in Nonconventional RNA Splicing. Cell 2008, 132, 89–100. [Google Scholar] [CrossRef] [PubMed]
- Shamu, C.; Cox, J.; Walter, P. The unfolded-protein-response pathway in yeast. Trends Cell Biol. 1994, 4, 56–60. [Google Scholar] [CrossRef]
- Bertolotti, A.; Wang, X.Z.; Novoa, I.; Jungreis, R.; Schlessinger, K.; Cho, J.H.; West, A.B.; Ron, D. Increased sensitivity to dextran sodium sulfate colitis in IRE1β-deficient mice. J. Clin. Investig. 2001, 107, 585–593. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martino, M.B.; Jones, L.; Brighton, B.; Ehre, C.; Abdulah, L.; Davis, C.W.; Ron, D.; O’Neal, W.K.; Ribeiro, C.M.P. The ER stress transducer IRE1β is required for airway epithelial mucin production. Mucosal Immunol. 2013, 6, 639–654. [Google Scholar] [CrossRef] [PubMed]
- Fanata, W.I.D.; Lee, S.Y.; Lee, K.O. The unfolded protein response in plants: A fundamental adaptive cellular response to internal and external stresses. Proteomics. J. 2013, 93, 356–368. [Google Scholar] [CrossRef] [PubMed]
- Okamura, K.; Kimata, Y.; Higashio, H.; Tsuru, A.; Kohno, K. Dissociation of Kar2p/BiP from an ER Sensory Molecule, Ire1p, Triggers the Unfolded Protein Response in Yeast. Biochem. Biophys. Res. Commun. 2000, 279, 445–450. [Google Scholar] [CrossRef] [PubMed]
- Kimata, Y.; Ishiwata-Kimata, Y.; Ito, T.; Hirata, A.; Suzuki, T.; Oikawa, D.; Takeuchi, M.; Kohno, K. Two regulatory steps of ER-stress sensor Ire1 involving its cluster formation and interaction with unfolded proteins. J. Cell Biol. 2007, 179, 75–86. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gardner, B.M.; Walter, P. Unfolded proteins are Ire1-activating ligands that directly induce the unfolded protein response. Science 2011, 333, 1891–1894. [Google Scholar] [CrossRef] [PubMed]
- Bowring, C.E.; Llewellyn, D.H. Differences in HAC1 mRNA processing and translation between yeast and mammalian cells indicate divergence of the eukaryotic ER stress response. Biochem. Biophys. Res. Commun. 2001, 287, 789–800. [Google Scholar] [CrossRef] [PubMed]
- Hooks, K.B.; Griffiths-Jones, S. Conserved RNA structures in the non-canonical Hac1/Xbp1 intron. RNA Biol. 2011, 8, 552–556. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, H.; Ng, B.S.H.; Thibault, G. Endoplasmic reticulum stress response in yeast and humans. Biosci. Rep. 2014, 34, 321–330. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kimmig, P.; Diaz, M.; Zheng, J.; Williams, C.C.; Lang, A.; Aragón, T.; Li, H.; Walter, P. The unfolded protein response in fission yeast modulates stability of select mRNAs to maintain protein homeostasis. Elife 2012, 1, e00048. [Google Scholar] [CrossRef] [PubMed]
- Tirasophon, W.; Lee, K.; Callaghan, B.; Welihinda, A.; Kaufman, R.J. The endoribonuclease activity of mammalian IRE1 autoregulates its mRNA and is required for the unfolded protein response. Genes Dev. 2000, 14, 2725–2736. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rubio, C.; Pincus, D.; Korennykh, A.; Schuck, S.; El-Samad, H.; Walter, P. Homeostatic adaptation to endoplasmic reticulum stress depends on Ire1 kinase activity. J. Cell Biol. 2011, 193, 171–184. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- van Anken, E.; Pincus, D.; Coyle, S.; Aragón, T.; Osman, C.; Lari, F.; Gómez Puerta, S.; Korennykh, A.V.; Walter, P. Specificity in endoplasmic reticulum-stress signaling in yeast entails a step-wise engagement of HAC1 mRNA to clusters of the stress sensor Ire1. Elife 2014, 3, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Aragón, T.; van Anken, E.; Pincus, D.; Serafimova, I.M.; Korennykh, A.V.; Rubio, C.A.; Walter, P. Messenger RNA targeting to endoplasmic reticulum stress signalling sites. Nature 2009, 457, 736–740. [Google Scholar] [CrossRef] [PubMed]
- Coelho, D.S.; Cairrão, F.; Zeng, X.; Pires, E.; Coelho, A.V.; Ron, D.; Ryoo, H.D.; Domingos, P.M. Xbp1-Independent Ire1 Signaling Is Required for Photoreceptor Differentiation and Rhabdomere Morphogenesis in Drosophila. Cell Rep. 2013, 5, 791–801. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Brandizzi, F. IRE1: ER stress sensor and cell fate executor. Trends Cell Biol. 2013, 23, 547–555. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miyazaki, T.; Nakayama, H.; Nagayoshi, Y.; Kakeya, H.; Kohno, S. Dissection of Ire1 functions reveals stress response mechanisms uniquely evolved in Candida glabrata. PLoS Pathog. 2013, 9, e1003160. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.-X.; Srivastava, R.; Che, P.; Howell, S.H. An endoplasmic reticulum stress response in Arabidopsis is mediated by proteolytic processing and nuclear relocation of a membrane-associated transcription factor, bZIP28. Plant Cell 2007, 19, 4111–4119. [Google Scholar] [CrossRef] [PubMed]
- Che, P.; Bussell, J.D.; Zhou, W.; Estavillo, G.M.; Pogson, B.J.; Smith, S.M. Signaling from the endoplasmic reticulum activates brassinosteroid signaling and promotes acclimation to stress in Arabidopsis. Sci. Signal. 2010, 3, ra69. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Shen, J.; Prywes, R. The luminal domain of ATF6 senses endoplasmic reticulum (ER) stress and causes translocation of ATF6 from the er to the Golgi. J. Biol. Chem. 2002, 277, 13045–13052. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, K.; Yoshida, H.; Kokame, K.; Kaufman, R.J.; Mori, K. Differential contributions of ATF6 and XBP1 to the activation of endoplasmic reticulum stress-responsive cis-acting elements ERSE, UPRE and ERSE-II. J. Biochem. 2004, 136, 343–350. [Google Scholar] [CrossRef] [PubMed]
- Okada, T.; Yoshida, H.; Akazawa, R.; Negishi, M.; Mori, K. Distinct roles of activating transcription factor 6 (ATF6) and double-stranded RNA-activated protein kinase-like endoplasmic reticulum kinase (PERK) in transcription during the mammalian unfolded protein response. Biochem. J. 2002, 366 Pt 2, 585–594. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Druelle, C.; Drullion, C.; Deslé, J.; Martin, N.; Saas, L.; Cormenier, J.; Malaquin, N.; Huot, L.; Slomianny, C.; Bouali, F.; et al. ATF6α regulates morphological changes associated with senescence in human fibroblasts. Oncotarget 2016, 7, 67699–67715. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Skorczyk-Werner, A.; Chiang, W.C.; Wawrocka, A.; Wicher, K.; Jarmuz-Szymczak, M.; Kostrzewska-Poczekaj, M.; Jamsheer, A.; Płoski, R.; Rydzanicz, M.; Pojda-Wilczek, D.; et al. Autosomal recessive cone-rod dystrophy can be caused by mutations in the ATF6 gene. Eur. J. Hum. Genet. 2017, 25, 1210–1216. [Google Scholar] [CrossRef] [PubMed]
- Chiang, W.-C.; Chan, P.; Wissinger, B.; Vincent, A.; Skorczyk-Werner, A.; Krawczyński, M.R.; Kaufman, R.J.; Tsang, S.H.; Héon, E.; Kohl, S.; et al. Achromatopsia mutations target sequential steps of ATF6 activation. Proc. Natl. Acad. Sci. USA 2017, 114, 400–405. [Google Scholar] [CrossRef] [PubMed]
- Marciniak, S.J.; Garcia-Bonilla, L.; Hu, J.; Harding, H.P.; Ron, D. Activation-dependent substrate recruitment by the eukaryotic translation initiation factor 2 kinase PERK. J. Cell Biol. 2006, 172, 201–209. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jindrich, K.; Degnan, B.M. The diversification of the basic leucine zipper family in eukaryotes correlates with the evolution of multicellularity Genome evolution and evolutionary systems biology. BMC EBiol. 2016, 16, 1–12. [Google Scholar]
- Nagashima, Y.; Mishiba, K.; Suzuki, E.; Shimada, Y.; Iwata, Y.; Koizumi, N. Arabidopsis IRE1 catalyses unconventional splicing of bZIP60 mRNA to produce the active transcription factor. Sci. Rep. 2011, 1, 29. [Google Scholar] [CrossRef] [PubMed]
- Ruggiano, A.; Foresti, O.; Carvalho, P. ER-associated degradation: Protein quality control and beyond. J. Cell Biol. 2014, 204, 869–879. [Google Scholar] [CrossRef] [PubMed]
- Tsai, B.; Ye, Y.; Rapoport, T.A. Retro-translocation of proteins from the endoplasmic reticulum into the cytosol. Nat. Rev. Mol. Cell Biol. 2002, 3, 246–255. [Google Scholar] [CrossRef] [PubMed]
- Klionsky, D.J.; Emr, S.D. Autophagy as a regulated pathway of cellular degradation. Science 2000, 290, 1717–1721. [Google Scholar] [CrossRef] [PubMed]
- Walter, P.; Ron, D. The unfolded protein response: From stress pathway to homeostatic regulation. Science 2011, 334, 1081–1086. [Google Scholar] [CrossRef] [PubMed]
- Urano, F.; Wang, X.-Z.; Bertolotti, A.; Zhang, Y.; Chung, P.; Harding, H.P.; Ron, D. Coupling of Stress in the Endoplasmic Reticulum to Activation of JNK Protein Kinases by Transmembrane Protein Kinase IRE1. Science 2000, 287, 664–666. [Google Scholar] [CrossRef] [PubMed]
- Ogata, M.; Hino, S.; Saito, A.; Morikawa, K.; Kondo, S.; Kanemoto, S.; Murakami, T.; Taniguchi, M.; Tanii, I.; Yoshinaga, K.; et al. Autophagy is activated for cell survival after endoplasmic reticulum stress. Mol. Cell. Biol. 2006, 26, 9220–9231. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Burgos, J.S.; Deng, Y.; Srivastava, R.; Howell, S.H.; Bassham, D.C. Degradation of the endoplasmic reticulum by autophagy during endoplasmic reticulum stress in Arabidopsis. Plant Cell 2012, 24, 4635–4651. [Google Scholar] [CrossRef] [PubMed]
- Goldberg, J.M.; Manning, G.; Liu, A.; Fey, P.; Pilcher, K.E.; Xu, Y.; Smith, J.L. The dictyostelium kinome—Analysis of the protein kinases from a simple model organism. PLoS Genet. 2006, 2, e38. [Google Scholar] [CrossRef] [PubMed]
- Caffrey, D.R.; O’Neill, L.A.; Shields, D.C. The evolution of the MAP kinase pathways: Coduplication of interacting proteins leads to new signaling cascades. J. Mol. Evol. 1999, 49, 567–582. [Google Scholar] [CrossRef] [PubMed]
- Giusti, C.; Tresse, E.; Luciani, M.F.; Golstein, P. Autophagic cell death: Analysis in Dictyostelium. Biochim. Biophys. Acta Mol. Cell Res. 2009, 1793, 1422–1431. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kroemer, G.; Galluzzi, L.; Vandenabeele, P.; Abrams, J.; Alnemri, E.S.; Baehrecke, E.H.; Blagosklonny, M.V.; El-Deiry, W.S.; Golstein, P.; Green, D.R.; et al. Classification of cell death: Recommendations of the Nomenclature Committee on Cell Death 2009. Cell Death Differ. 2009, 16, 3–11. [Google Scholar] [CrossRef] [PubMed]
- Baehrecke, E.H. Autophagic programmed cell death in Drosophila. Cell Death Differ. 2003, 10, 940–945. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hofius, D.; Schultz-Larsen, T.; Joensen, J.; Tsitsigiannis, D.I.; Petersen, N.H.T.; Mattsson, O.; Jørgensen, L. B.; Jones, J.D.G.; Mundy, J.; Petersen, M. Autophagic Components Contribute to Hypersensitive Cell Death in Arabidopsis. Cell 2009, 137, 773–783. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Levine, B. Autosis and autophagic cell death: The dark side of autophagy. Cell Death and Differentiation 2015, 22, 367–376. [Google Scholar] [CrossRef] [PubMed]
- Whittingham, W.F.; Raper, K.B. Non-Viability of Stalk Cells in Dictyostelium. Proc. Natl. Acad. Sci. USA 1960, 46, 642–649. [Google Scholar] [CrossRef] [PubMed]
- Levraud, J.-P.; Adam, M.; Luciani, M.-F.; de Chastellier, C.; Blanton, R.L.; Golstein, P. Dictyostelium cell death: Early emergence and demise of highly polarized paddle cells. J. Cell Biol. 2003, 160, 1105–1114. [Google Scholar] [CrossRef] [PubMed]
- Lam, D.; Golstein, P. A specific pathway inducing autophagic cell death is marked by an IP3R mutation. Autophagy 2008, 8627, 349–350. [Google Scholar] [CrossRef]
- Wolf, D.H.; Stolz, A. The Cdc48 machine in endoplasmic reticulum associated protein degradation. Biochim. Biophys. Acta Mol. Cell Res. 2012, 1823, 117–124. [Google Scholar] [CrossRef] [PubMed]
- Müller-Taubenberger, A. Application of fluorescent protein tags as reporters in live-cell imaging studies. Meth. Mol. Biol. 2006, 346, 229–246. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Function/Features | Dictyostelium a | Human a | Saccharomyces cerevisiae a | Arabidopsis thaliana a |
|---|---|---|---|---|
| ER structure | ||||
| Transmembrane protein that promotes membrane curvature, and participates in maintenance of tubular ER morphology | Reticulon-like group C (Rtnlc)/ Q54CA6 | Reticulons-1 to 4 (RTN1 to RTN4)/ Q16799, O75298, O95197, Q9JK11 | Reticulon-like proteins 1 (RTN1) and 2 (RTN2)/ A0A250W951, Q12443 | Reticulon-like proteins B1 to 18, and 21 to 23 (RTNLB1 to 18 and RTNLB21 to 23)/ Q9SUR3, Q9SUT9, Q9SH59, Q9FFS0, O82352, Q6DBN4, Q9M145, Q9SS37, Q9LJQ5, Q6NPD8, Q9LT71, Q9M392, O64837, A2RVT6, Q9ZU43, Q8GYH6, Q6DR04, Q8LDS3, Q56X72, Q8GWH5, P0C941 |
| Dynamin-like GTPase that mediates homotypic ER fusion | Sey1/ Q54W90 | Atlastin-1 (ATL1)/ Q8WXF7 | Sey1/ Q99287 | Root hair defective 3 (RHD3) and root hair defective 3 homolog 2 (RHD3-2)/ P93042, Q9FKE9 |
| ER contact sites Components of the mitochondria encounter sites (ERMES), which are involved in the tether between the ER and the mitochondria to promote inter-organellar calcium and phospholipid exchange | Maintenance of mitochondrial morphology-1 (Mmm1)/ Q54MI5 | ND | Maintenance of mitochondrial morphology protein 1 (Mmm1)/ P41800 | ND |
| Mitochondrial distribution and morphology-10 (Mdm10)/ Q54XQ5 | ND | Mitochondrial distribution and morphology 10 (Mdm10)/ P18409 | ND | |
| Mitochondrial distribution and morphology 34 (Mdm34)/ Q869R5 | ND | Mitochondrial distribution and morphology protein 34 (Mdm34)/ P53083 | ND | |
| Transmembrane protein required to regulate ER contact sites, essential for autophagy and proper ER homeostasis | Vacuole membrane protein 1 (Vmp1)/ Q54NL4 | Vacuole membrane protein 1 (VMP1)/ Q96GC9 | ND | Vacuole membrane proteins 1 (KMS1) and 2 (KMS2)/ Q5XF36, F4I8Q7 |
| Lipid metabolism | ||||
| Protein that associates to the lipid droplet surface | Perilipin (PlnA)/ Q54WC4 | Perilipin proteins 1 to 5 (PLIN1 to 5)/ O60240, Q99541, O60664, Q96Q06, Q00G26 | ND | ND |
| Catalyze the conversion of acyl coenzyme A (CoA) and 1,2-diacylglycerol to CoA and triacylglycerol. | Diacylglycerol O-acyltransferases 1 (Dgat1) and 2 (Dgat2)/ Q55BH9, Q54GC1 | Diacylglycerol O-acyltransferases 1 (DGAT1), 2-acylglycerol O-acyltransferase 1 (MOGAT1)/ O75907, Q96PD6 | Sterol O-acyltransferases 1 (Are1) and 2 (Are2)/ P25628, P53629 | Diacylglycerol O-acyltransferase 1 (DGAT1)/ Q9SLD2 |
| Protein folding and modification | ||||
| Subunits of the oligosaccharyl transferase complex, which catalyzes asparagine-linked glycosylation of newly synthesized proteins in the ER lumen | Oligosaccharyl transferase-1 (Ost1)/ Q54C27 | Dolichyl-diphosphooligosaccharide-protein glycosyltransferase subunit 1 (RPN1)/ P04843 | Dolichyl-diphosphooligosaccharide-protein glycosyltransferase subunit 1 (Ost1)/ P41543 | Dolichyl-diphosphooligosaccharide-protein glycosyltransferase subunits 1A (OST1A) and 1B (OST1B)/ Q9SFX3, Q9ZUA0 |
| Oligosaccharyl transferase-2 (Ost2)/ Q54FB6 | Dolichyl-diphosphooligosaccharide-protein glycosyltransferase subunit (DAD1)/ P61803 | Dolichyl-diphosphooligosaccharide-protein glycosyltransferase subunit (OST2)/ P46964 | Dolichyl-diphosphooligosaccharide-protein glycosyltransferase subunits 1 (DAD1) and 2 (DAD2)/ Q39080, O22622 | |
| Oligosaccharyl transferase-3 (Ost3)/ Q54N33 | ND | Dolichyl-diphosphooligosaccharide-protein glycosyltransferase subunit 3 (Ost3)/ P48439 | Dolichyl-diphosphooligosaccharide-protein glycosyltransferase subunits 3A (OST3A) and 3B (OST3B)/ F4I8X8, Q9SYB5 | |
| Oligosaccharyl transferase-4 (Ost4)/ Q54V54 | Dolichyl-diphosphooligosaccharide-protein glycosyltransferase subunit 4 (OST4)/ P0C6T2 | Dolichyl-diphosphooligosaccharide-protein glycosyltransferase subunit 4 (Ost4)/ Q99380 | ND | |
| Oligosaccharyl transferase complex subunit C (Ostc)/ Q54X66 | Oligosaccharyltransferase complex subunit OSTC (OSTC)/ Q9NRP0 | ND | Oligosaccharyl transferase complex/magnesium transporter family protein (At4g29870)/ Q9SZQ8 | |
| Wheat germ agglutinin-binding protein (Wbp1)/ Q54E62 | Dolichyl-diphosphooligosaccharide-protein glycosyltransferase 48 kDa subunit (DDOST)/ P39656 | Dolichyl-diphosphooligosaccharide-protein glycosyltransferase subunit (Wbp1)/ P33767 | Dolichyl-diphosphooligosaccharide-protein glycosyltransferase 48 kDa subunit (OST48)/ Q944K2 | |
| Suppressor of a WBP1 mutation (Swp1)/ Q54HG9 | Dolichyl-diphosphooligosaccharide-protein glycosyltransferase subunit 2 (RPN2)/ P04844 | Dolichyl-diphosphooligosaccharide-protein glycosyltransferase subunit (Swp1)/ Q02795 | Dolichyl-diphosphooligosaccharide-protein glycosyltransferase subunit 2 (RPN2)/ Q93Z16 | |
| Staurosporine and temperature sensitivity (Stt3)/ Q54NM9 | Dolichyl-diphosphooligosaccharide-protein glycosyltransferase subunits A (STT3A) and B (STT3B)/ P46977, Q8TCJ2 | Dolichyl-diphosphooligosaccharide-protein glycosyltransferase subunit (Stt3)/ P39007 | Dolichyl-diphosphooligosaccharide-protein glycosyltransferase subunits A (STT3A) and B (STT3B)/ Q93ZY3, Q9FX21 | |
| Heat shock protein 70 (Hsp70)-family chaperone | 78 kDa Glucose-regulated protein (Grp78)/ Q8T869 | Binding immunoglobulin protein/78 kDa glucose-regulated protein (BiP/Grp78)/ P11021 | Binding immunoglobulin protein (BiP/Kar2)/ P16474 | Binding immunoglobulin protein 2 (BIP2)/ F4K007 |
| Hsp90-family chaperone | 94 kDa Glucose-regulated protein (Dd-grp94)/ Q9NKX1 | Endoplasmin (GRP94)/ P14625 | ATP-dependent molecular chaperone (Hsp82)/ P02829 | Endoplasmin homolog (HSP90-7)/ Q9STX5 |
| Calcium-binding proteins with chaperone activity | Calreticulin (CrtA)/ Q23858 | Calreticulin (CALR)/ P27797 | ND | Calreticulin-1 (CRT1) and 2 (CRT2)/ O04151, Q388587 |
| Calnexin (CnxA)/ Q55BA8 | Calnexin (CANX)/ P27824 | Calnexin homolog (Cne1)/ P27825 | Calnexin homolog 1 (CNX1) and 2 (CNX2)/ P29402, Q38798 | |
| ER luminal protein that catalyzes the formation and remodeling of protein disulfide bonds | Protein disulfide isomerases 1 (Pdi1) and 2 (Pdi2)/ Q86IA3, Q54EN4 | Protein disulfide isomerases (P4HB), A4 (PDIA4), A3 (PDIA3), and A6 (PDIA6)/ P07237, P13667, P30101, Q15084 | Protein disulfide isomerase (Pdi1)/ P17967 | Protein disulfide isomerase-like proteins 1-1 (PDIL1-1), 1-2 (PDIL1-2), 2-2 (PDIL2-2), and 2-3 (PDIL2-3)/ Q9XI01, Q9SRG3, O22263, O48773 |
| Unfolded Protein Response | ||||
| ER transmembrane serine and threonine kinase with ribonuclease activity that senses ER stress | Inositol-requiring enzyme A (IreA)/ Q55GJ2 | Inositol-requiring enzyme proteins 1α (IRE1 α or ERN1) and 1β (IRE1β or ERN2)/ O75460, Q76MJ5 | Inositol-requiring enzyme 1 (Ire1)/ P32361 | Inositol-requiring enzyme proteins 1a (IRE1a) and 1b (IRE1b)/ Q93VJ2, |
| Calcium channel Ion channel participates in calcium release from the ER, and is activated by inositol trisphosphate | Inositol 1,4,5-trisphosphate receptor (IplA)/ Q9NA13 | Inositol 1,4,5-trisphosphate receptors type 1 (ITPR1), type 2 (ITPR2), and type 3 (ITPR3)/ Q14643, Q14571, Q14573 | ND | ND |
| Gene ID | Name | Description | IreA-Dependent |
|---|---|---|---|
| DDB_G0276445 | Grp78 | Heat shock protein Hsp70 family protein. | no |
| DDB_G0274199 | DDB_G0274199 | Putative metallophosphoesterase. | no |
| DDB_G0278477 | sarB | ADP ribosylation factors/ Secretion-associated and Ras-related (ARF/SAR) superfamily protein. GTP-binding protein Sar1B involved in vesicular transport between the endoplasmic reticulum and the Golgi body. | no |
| DDB_G0283867 | cprC | Cysteine proteinase 3. | no |
| DDB_G0278371 | spc1 | Ortholog of the conserved microsomal signal peptidase 12 kDa subunit; the signal peptidase complex is a membrane-bound endo-proteinase that removes signal peptides from nascent proteins as they are translocated into the lumen of the endoplasmic reticulum. | no |
| DDB_G0281833 | DDB_G0281833 | Ubiquitin-conjugating enzyme E2. | no |
| DDB_G0283113 | eriA | RNA exonuclease. | no |
| DDB_G0290227 | npl4 | Ortholog of nuclear protein localization 4 (NPL4), which, together with ubiquitin fusion degradation protein 1 (Ufd1) and cell division cycle protein D (CdcD), is involved in recognition of polyubiquitinated proteins, and their presentation to the 26S proteasome for degradation. | no |
| DDB_G0287685 | cinC | Elongation factor 2. Translocates the peptidyl-tRNA from the aminoacyl site to the peptidyl site on the ribosome during protein synthesis; induced by cycloheximide; knockdown has significantly reduced ability for protein synthesis. | yes |
| DDB_G0269462 | DDB_G0269462 | Large protein containing two ubiquitin domains. | yes |
| DDB_G0291121 | cinB | Esterase/lipase/thioesterase domain-containing protein. | yes |
| DDB_G0285131 | derl2 | Derlin-2. component of endoplasmic reticulum-associated degradation (ERAD) for misfolded luminal proteins. | yes |
| DDB_G0270272 | uae1 | Ubiquitin activating enzyme E1. | yes |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Domínguez-Martín, E.; Hernández-Elvira, M.; Vincent, O.; Coria, R.; Escalante, R. Unfolding the Endoplasmic Reticulum of a Social Amoeba: Dictyostelium discoideum as a New Model for the Study of Endoplasmic Reticulum Stress. Cells 2018, 7, 56. https://doi.org/10.3390/cells7060056
Domínguez-Martín E, Hernández-Elvira M, Vincent O, Coria R, Escalante R. Unfolding the Endoplasmic Reticulum of a Social Amoeba: Dictyostelium discoideum as a New Model for the Study of Endoplasmic Reticulum Stress. Cells. 2018; 7(6):56. https://doi.org/10.3390/cells7060056
Chicago/Turabian StyleDomínguez-Martín, Eunice, Mariana Hernández-Elvira, Olivier Vincent, Roberto Coria, and Ricardo Escalante. 2018. "Unfolding the Endoplasmic Reticulum of a Social Amoeba: Dictyostelium discoideum as a New Model for the Study of Endoplasmic Reticulum Stress" Cells 7, no. 6: 56. https://doi.org/10.3390/cells7060056