Size Polymorphism in Alleles of the Myoglobin Gene from Biomphalaria Mollusks

Abstract
:1. Introduction
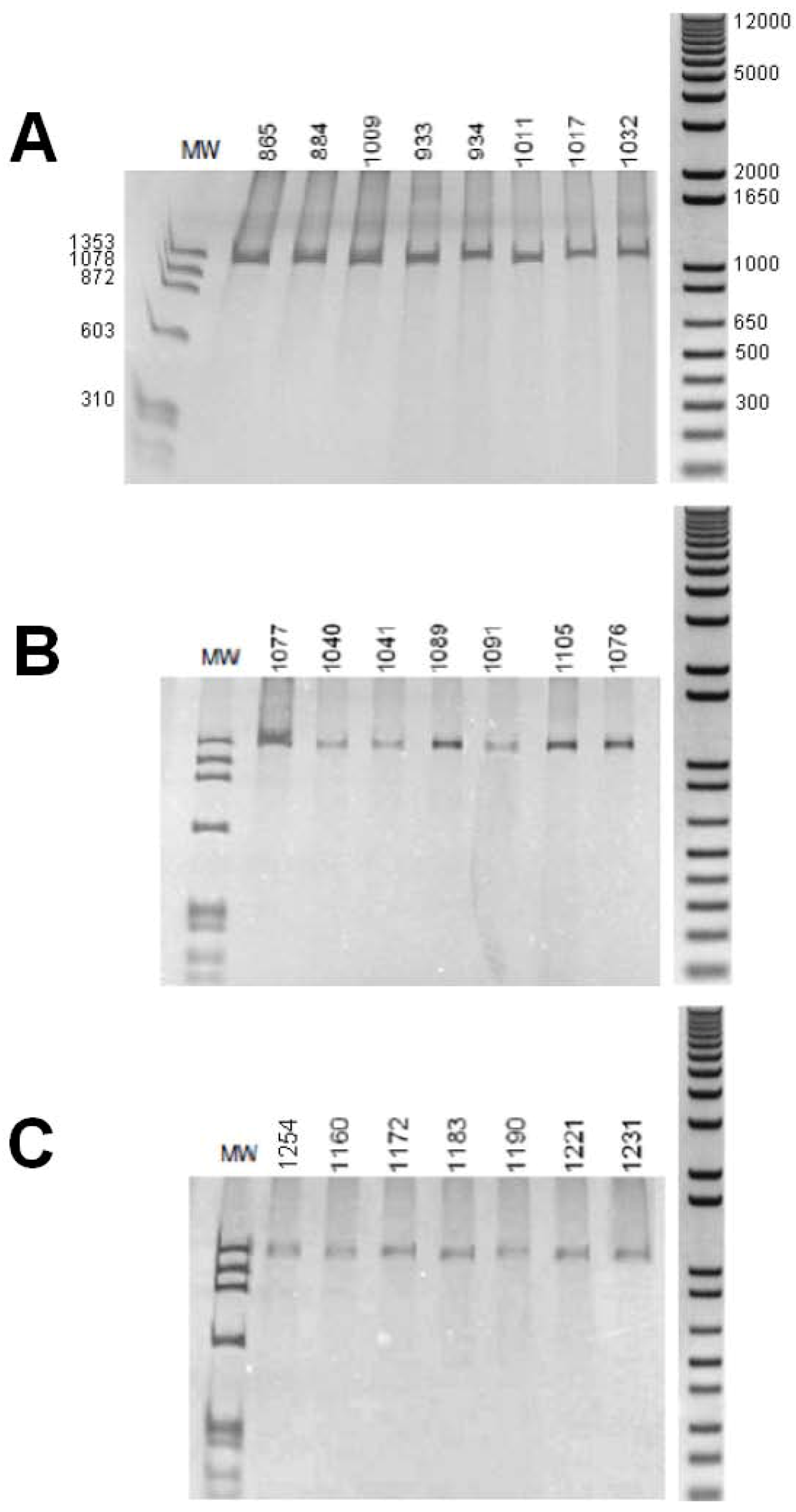
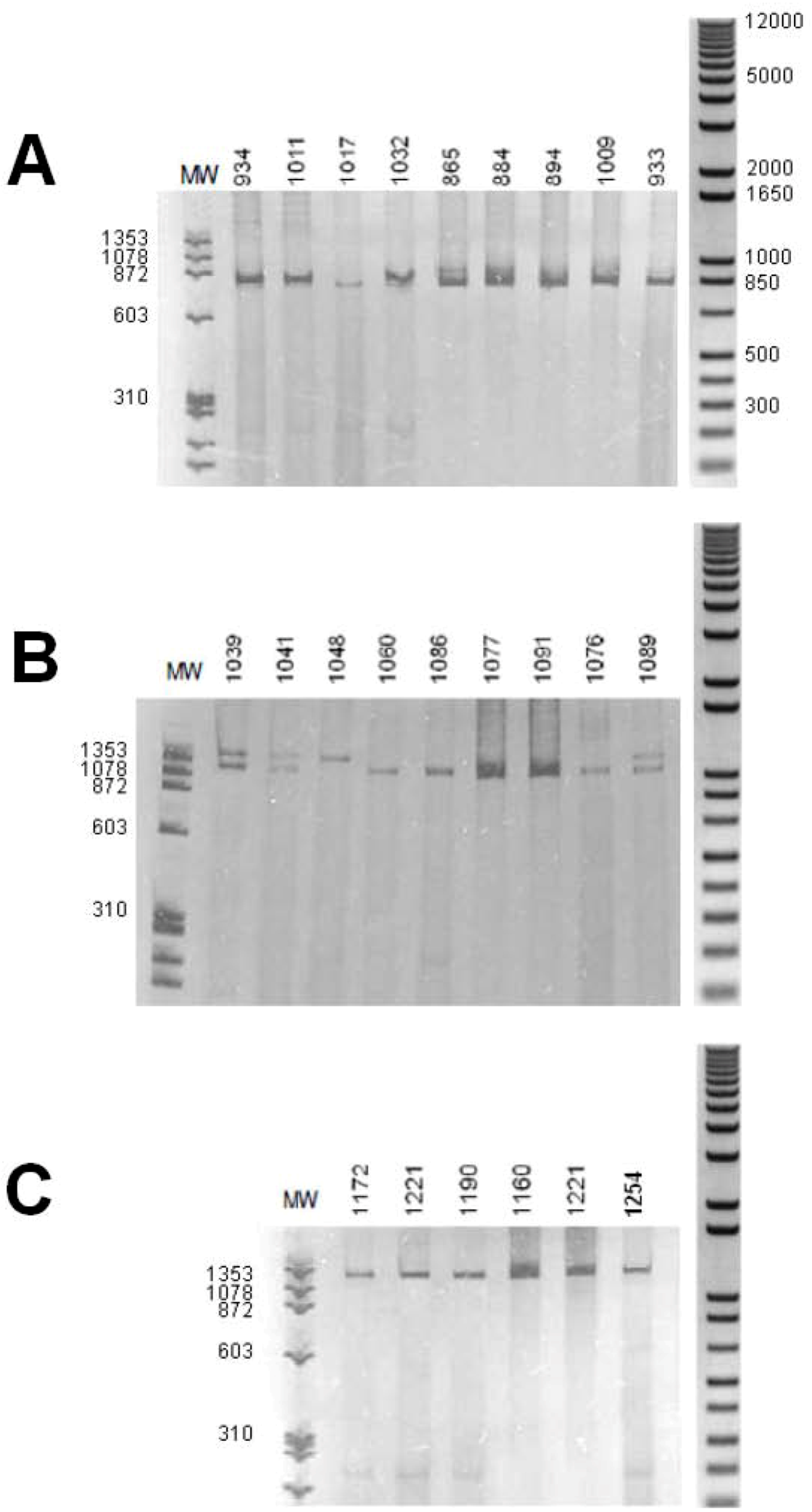
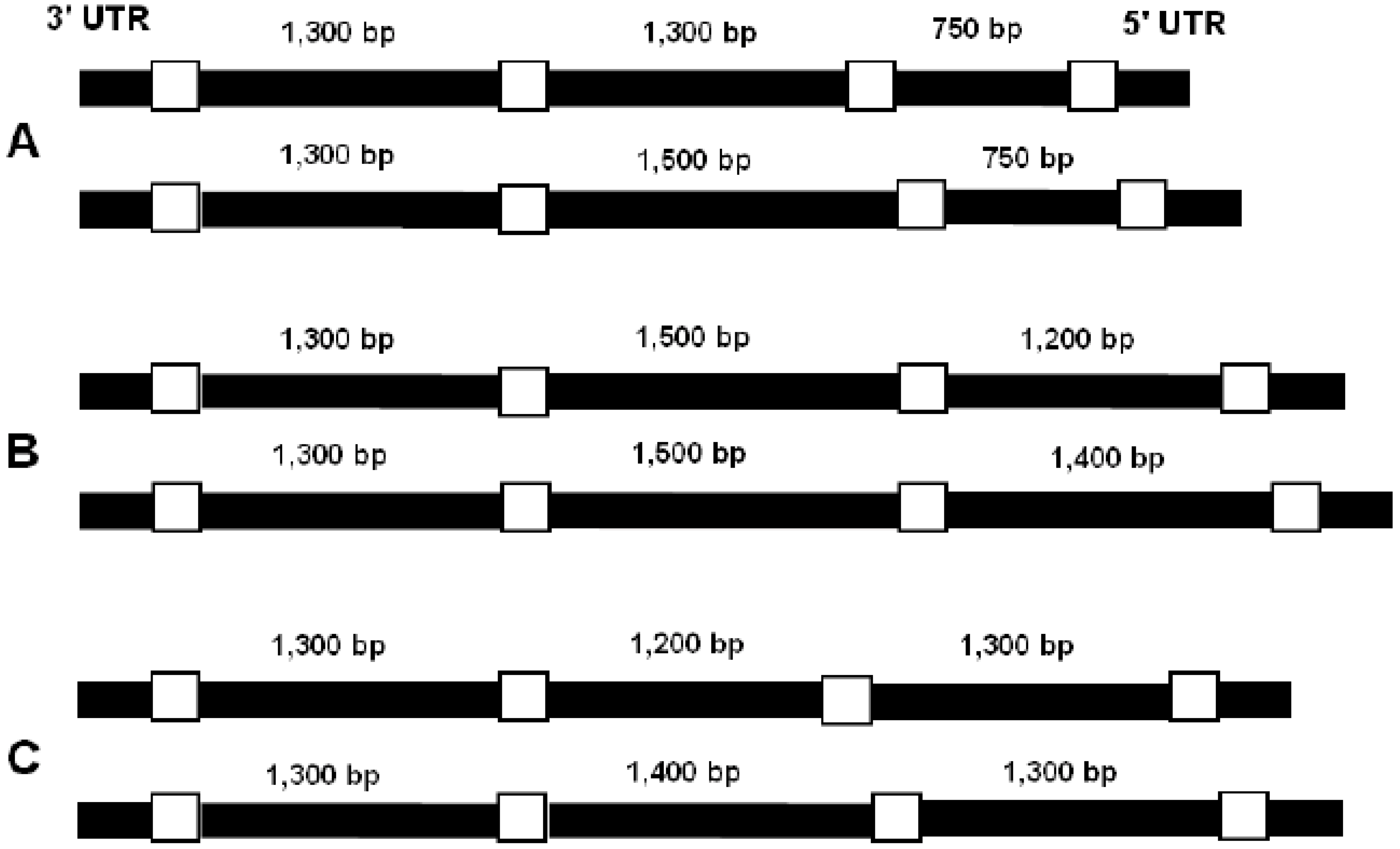
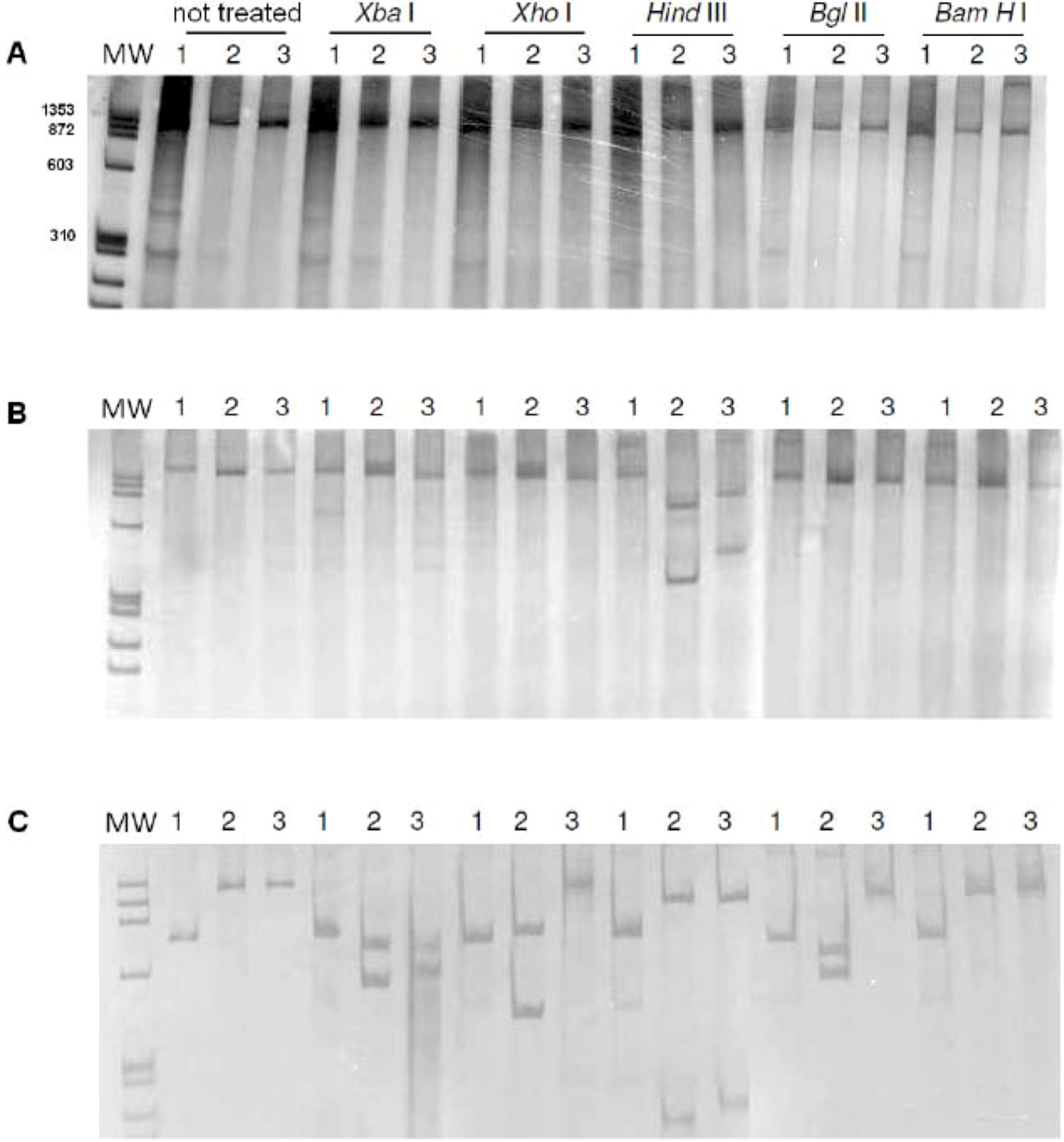
2. Results, Discussion





3. Experimental Section
3.1. Snails

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Individuals a | Province from Brazil |
|---|---|---|
| B. glabrata | 865 | South East |
| 884, 1009, 1011, 1017 | North East | |
| 933, 934, 1032 | North | |
| B. straminea | 1040, 1041, | North |
| 1076, 1077, 1091 | North East | |
| 1089, 1105 | South East | |
| B. tenagophila | 1160, 1183, 1231 | South East |
| 1172, 1190 | South | |
| 1221 | North East | |
| 1254 | Central West |
3.2. Genomic DNA Extraction of Snails
3.3. Amplification of the Introns from the Biomphalaria Myoglobin Gene
3.4. PCR-RFLP
4. Conclusion

References and Notes
- Berget, S.M.; Moore, C.; Sharp, P.A. Spliced segments at the 5 terminus of adenovirus 2 late mRNA. Proc. Natl. Acad. Sci. U. S. A. 1977, 74, 3171–3175. [Google Scholar] [CrossRef] [PubMed]
- Chow, L.T.; Gelinas, R.E.; Broker, J.R.; Roberts, R.J. An amazing sequence arrangement at the 5 ends of adenovirus 2 messenger RNA. Cell 1977, 12, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Jeffreys, A.J.; Flavell, R.A. The rabbit beta-globin gene contains a large insert in the coding sequence. Cell 1977, 12, 1097–1108. [Google Scholar] [CrossRef] [PubMed]
- Lynch, M.; Richardson, A.O. The evolution of spliceosomal introns. Curr. Opin. Genet. Dev. 2002, 12, 701–710. [Google Scholar] [CrossRef] [PubMed]
- Fedorova, L.; Fedorov, A. Introns in gene evolution. Genetica 2003, 118, 123–131. [Google Scholar] [CrossRef] [PubMed]
- Cavalier-Smith, T. Selfish DNA, the origin of introns. Nature 1985, 315, 283–284. [Google Scholar] [CrossRef] [PubMed]
- Dibb, N.J. Why do genes have introns? FEBS Lett. 1993, 325, 135–139. [Google Scholar] [CrossRef] [PubMed]
- Bertone, P.; Stolc, V.; Royce, T.E.; Rozowsky, J.S.; Urban, A.E.; Zhu, X.; Rinn, J.L.; Tongprasit, W.; Samanta, M.; Weissman, S.; Gerstein, M.; Snyder, M. Global identification of human transcribed sequences with genome tiling arrays. Science 2004, 306, 2242–2246. [Google Scholar] [CrossRef] [PubMed]
- Guigó, R.; Flicek, P.; Abril, J.F.; Reymond, A.; Lagarde, J.; Denoeud, F.; Antonarakis, S.; Ashburner, M.; Bajic, V.B.; Birney, E.; Castelo, R.; Eyras, E.; Ucla, C.; Gingeras, T.R.; Harrow, J.; Hubbard, T.; Lewis, S.E.; Reese, M.G. EGASP: The human ENCODE Genome Annotation Assessment Project. Genome Biol. 2006, 7, 1–31. [Google Scholar] [CrossRef] [PubMed]
- Doolittle, W.F. Gene in pieces: Were they ever together. Nature 1978, 272, 581–582. [Google Scholar] [CrossRef]
- Darnell Jr., J.E. On the origin of prokaryotes. Science 1978, 202, 1257–1260. [Google Scholar] [PubMed]
- Palmer, J.D.; Logsdon Jr. , J.M. The recent origins of introns . Curr. Opin. Genet. Dev. 1991, 1, 470–477. [Google Scholar] [CrossRef]
- Logsdon Jr., J.M.; Stoltzfus, A.; Doolittle, W.F. Recent cases of spliceosomal intron gain? Curr. Biol. 1998, 8, R560–R563. [Google Scholar] [CrossRef] [PubMed]
- Logsdon Jr., J.M.; Tyshenko, M.G.; Dixon, C.; Jafari, J.D.; Walker, V.K.; Palmer, J.D. Seven newly discovered intron positions in the triose-phosphate isomerase gene: Evidence for the introns-late theory. Proc. Natl. Acad. Sci. U. S. A. 1995, 92, 8507–8511. [Google Scholar] [CrossRef] [PubMed]
- Rzhetsky, A.; Ayala, F.J.; Hsu, L.C.; Ghang, C.; Yoshida, A. Exon/intron structure of aldehyde dehydrogenase genes supports the “introns-late” theory. Proc. Natl. Acad. Sci. U. S. A. 1997, 94, 6820–6825. [Google Scholar] [CrossRef] [PubMed]
- Frugoli, J.A.; McPeek, M.A.; Thomas, T.L.; Mcclung, C.R. Intron loss, gain during evolution of the catalase gene family in angiosperms. Genetics 1998, 149, 355–365. [Google Scholar] [PubMed]
- Tarrio, R.; Rodriguez-Trelles, F.; Ayala, F.J. New Drosophila introns originate by duplication. Proc. Natl. Acad. Sci. U. S. A. 1998, 95, 1658–1662. [Google Scholar] [CrossRef] [PubMed]
- Venkatesh, B.; Ning, Y.; Brenner, S. Late changes in spliceosomal introns define clades in vertebrate evolution. Proc. Natl. Acad. Sci. U. S. A. 1999, 96, 10267–10271. [Google Scholar] [CrossRef] [PubMed]
- Rogosin, I.B.; Wolf, Y.L.; Sorokin, A.V.; Mirkin, B.G.; Koonin, E.V. Remarkable interkingdom conservation of intron positions, massive, lineage-specific intron loss, gain in eukaryotic evoltuion. Curr. Biol. 2003, 13, 1512–1517. [Google Scholar] [CrossRef] [PubMed]
- van der Peer, Y.; Neefs, J.; de Rijk, P.; de Wachter, R. Evolution of eukaryotes as deduced from small ribosomal subunit RNA sequences. Biochem. Syst. Ecol. 1993, 21, 43–55. [Google Scholar] [CrossRef]
- Rogers, J.H. The role of introns in evolution. FEBS 1990, 268, 339–343. [Google Scholar] [CrossRef]
- Dewilde, S.; Winnepenninckx, B.; Arndt, N.H.L.; Nascimento, D.G.; Santoro, M.M.; Knight, M.; Miller, A.N.; Kerlavage, A.R.; Geoghagen, N.; Van Marck, E.; Liu, L.X.; Weber, R.E.; Moens, L. Characterization of the myoblobin, its coding gene of the mollusk Biomphalaria glabrata. J. Biol. Chem. 1998, 273, 13583–13592. [Google Scholar] [CrossRef] [PubMed]
- Jellie, A.M.; Tate, W.P.; Trotman, C.N.A. Evolutionary history of introns in a multidomain globin gene. J. Mol. Evol. 1996, 42, 641–647. [Google Scholar] [CrossRef] [PubMed]
- Paraense, W.L. Estado atual da sistemática dos planorbídeos brasileiros. Arq. Mus. Nac. Rio de Janeiro 1975, 55, 105–128. [Google Scholar]
- Monis, P.T. The importance of systematics in parasitological research. Int. J. Parasitol. 1999, 29, 381–388. [Google Scholar] [CrossRef] [PubMed]
- Vidigal, T.H.A.; Dias Neto, E.; Simpson, A.J.; Carvalho, O.S. A low stringency polymerase chain reaction approach to the identification of Biomphalaria glabrata, B. tenagophila, intermediate snail hosts of Schistosoma mansoni in Brazil. Mem. Inst. Oswaldo Cruz 1996, 91, 739–744. [Google Scholar] [CrossRef] [PubMed]
- Knight, M.; Brindley, P.J.; Richards, C.S.; Lewis, F.A. Schistosoma mansoni: use of a cloned ribosomal RNA gene probe to detect restriction fragment length polymorphisms in the intermediate host Biomphalaria glabrata. Exp. Parasitol. 1991, 73, 285–294. [Google Scholar] [CrossRef] [PubMed]
- Sokolova, I.M.; Boulding, E.G. Length polymorphisms in an intron of aminopeptidase N provide a useful nuclear DNA marker for Littorina species (Caenogastropoda). J. Moll. Stud. 2004, 70, 165–172. [Google Scholar] [CrossRef]
- Suzuki, T.; Imai, K. Evolution of myoglobin. Cell. Mol. Life Sci. 1998, 54, 979–1004. [Google Scholar] [CrossRef] [PubMed]
- Dewilde, S.; Van Hauwaert, M.L.; Vinogradov, S.; Vierstraete, A.; Vanfleteren, J.; Moens, L. Protein, gene structure of a chlorocruorin chain of Eudistylia vancouverii. BBRC 2001, 281, 18–24. [Google Scholar] [PubMed]
- Lewin, R. Surprise finding with insect globin genes. Science 1984, 226, 328. [Google Scholar] [PubMed]
- Vinogradov, S.N.; Walz, D.A.; Pohajdak, B. Organization of non-vertebrate globin genes. Comp. Biochem. Physiol. 1992, 103, 759–773. [Google Scholar] [CrossRef]
© 2010 by the authors; licensee MDPI, Basel, Switzerland This is an open access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited.
Share and Cite
Teixeira, K.N.; Souza, K.N.; Vidigal, T.H.D.A.; Brito, C.A.; Santos, A.M.C.; Santoro, M.M. Size Polymorphism in Alleles of the Myoglobin Gene from Biomphalaria Mollusks. Genes 2010, 1, 357-370. https://doi.org/10.3390/genes1030357
Teixeira KN, Souza KN, Vidigal THDA, Brito CA, Santos AMC, Santoro MM. Size Polymorphism in Alleles of the Myoglobin Gene from Biomphalaria Mollusks. Genes. 2010; 1(3):357-370. https://doi.org/10.3390/genes1030357
Chicago/Turabian StyleTeixeira, Kádima N., Karyne N. Souza, Teofânia H.D.A. Vidigal, Cristiane A. Brito, Alexandre M.C. Santos, and Marcelo M. Santoro. 2010. "Size Polymorphism in Alleles of the Myoglobin Gene from Biomphalaria Mollusks" Genes 1, no. 3: 357-370. https://doi.org/10.3390/genes1030357