Synaptonemal Complex Length Variation in Wild-Type Male Mice

Abstract
:1. Introduction
2. Results
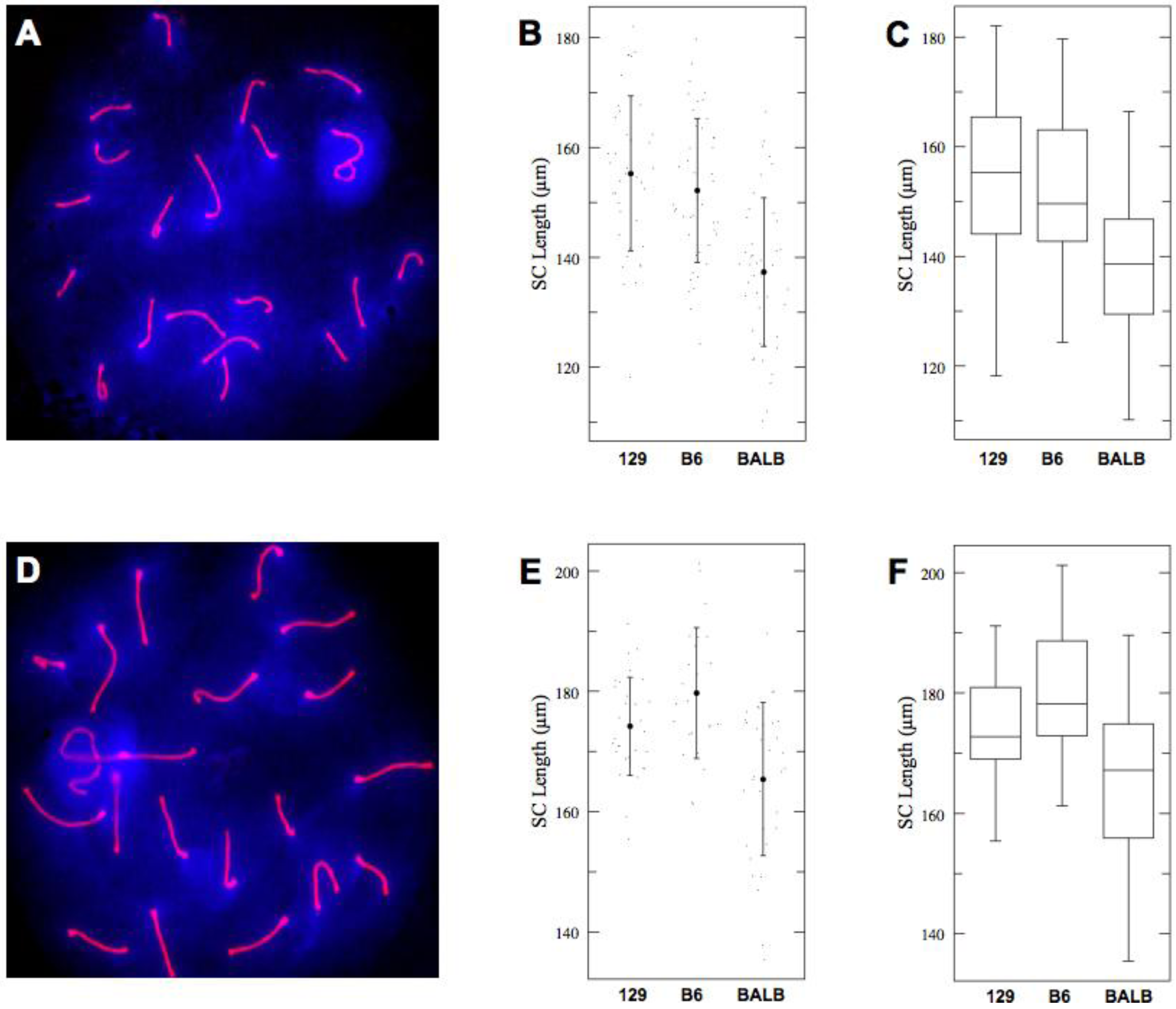
2.1. Short SCs in BALB/c Spermatocytes


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain | n | Mean | SD | 95% CI |
|---|---|---|---|---|
| 129S4 | 69 | 163.48 | 15.1 | 159.3–167.7 |
| C57BL6J | 68 | 161.89 | 18.1 | 157.7–166.1 |
| BALBc | 75 | 149.69 | 19.2 | 145.7–153.7 |

| Strain | n | Mean | SD | 95% CI |
|---|---|---|---|---|
| 129S4 (early) | 39 | 155.25 | 14.1 | 151.0–159.5 |
| C57BL6J (early) | 44 | 152.17 | 13.1 | 148.1–156.2 |
| BALBc (early) | 42 | 137.53 | 13.4 | 133.4–141.7 |
| 129S4 (late) | 30 | 174.18 | 8.15 | 170.3–178.1 |
| C57BL6J (late) | 24 | 179.71 | 10.9 | 175.3–184.1 |
| BALBc (late) | 33 | 165.41 | 12.7 | 161.7–169.2 |
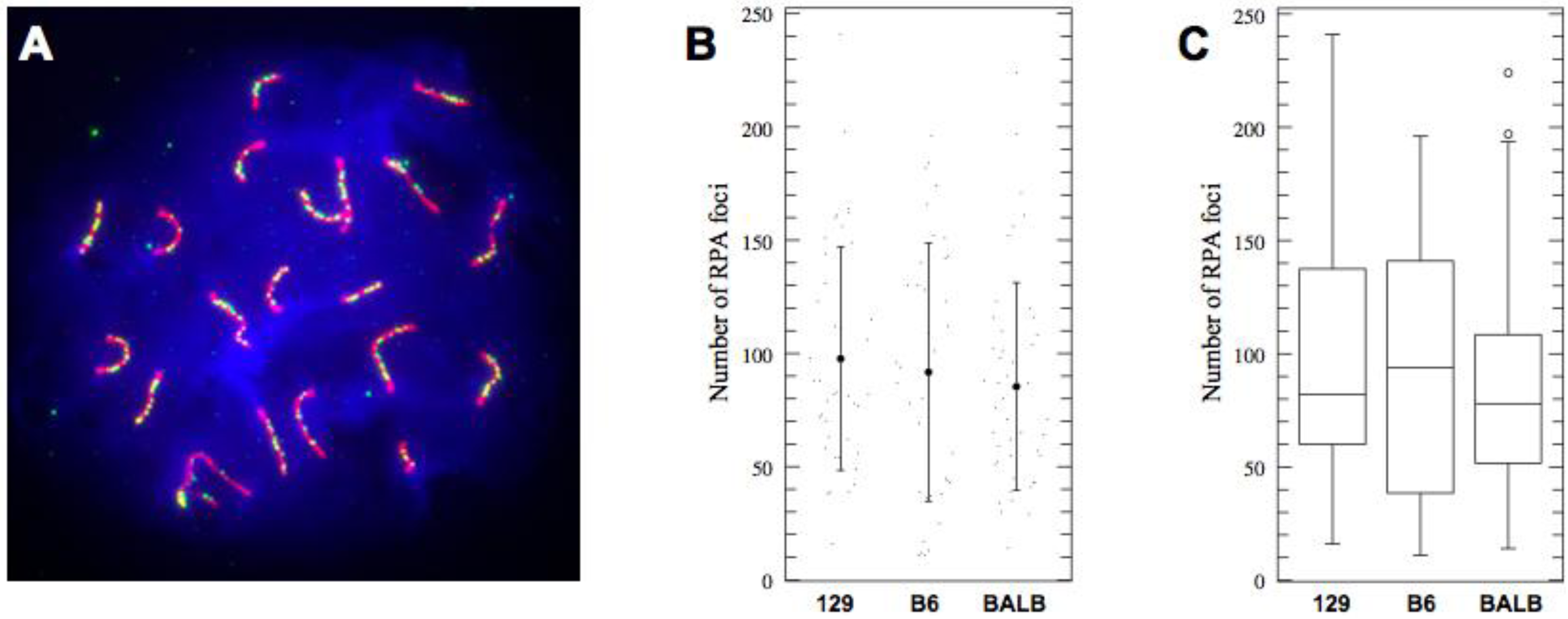
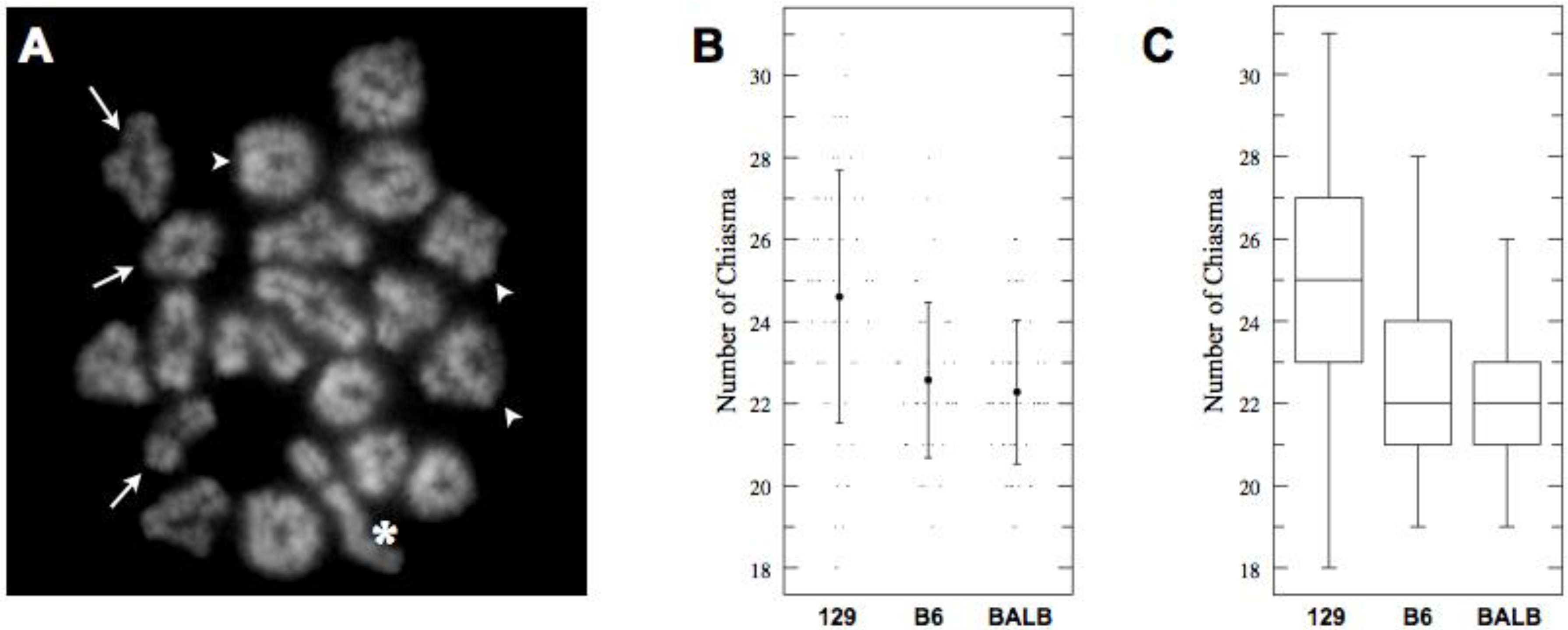
2.2. No Increase in COs despite Longer SCs in C57BL/6J Mice

| Strain | n | Mean | SD | 95% CI |
|---|---|---|---|---|
| 129S4 | 41 | 97.68 | 49.3 | 82.06–113.3 |
| C57BL6J | 37 | 91.73 | 57.0 | 75.29–108.2 |
| BALBc | 46 | 85.39 | 45.9 | 70.65–100.1 |

| Strain | n | Mean | SD | 95% CI |
|---|---|---|---|---|
| 129S4 | 53 | 24.60 | 3.08 | 23.97–25.24 |
| C57BL6J | 40 | 22.56 | 1.89 | 21.84–23.31 |
| BALBc | 56 | 22.29 | 1.76 | 21.67–22.90 |
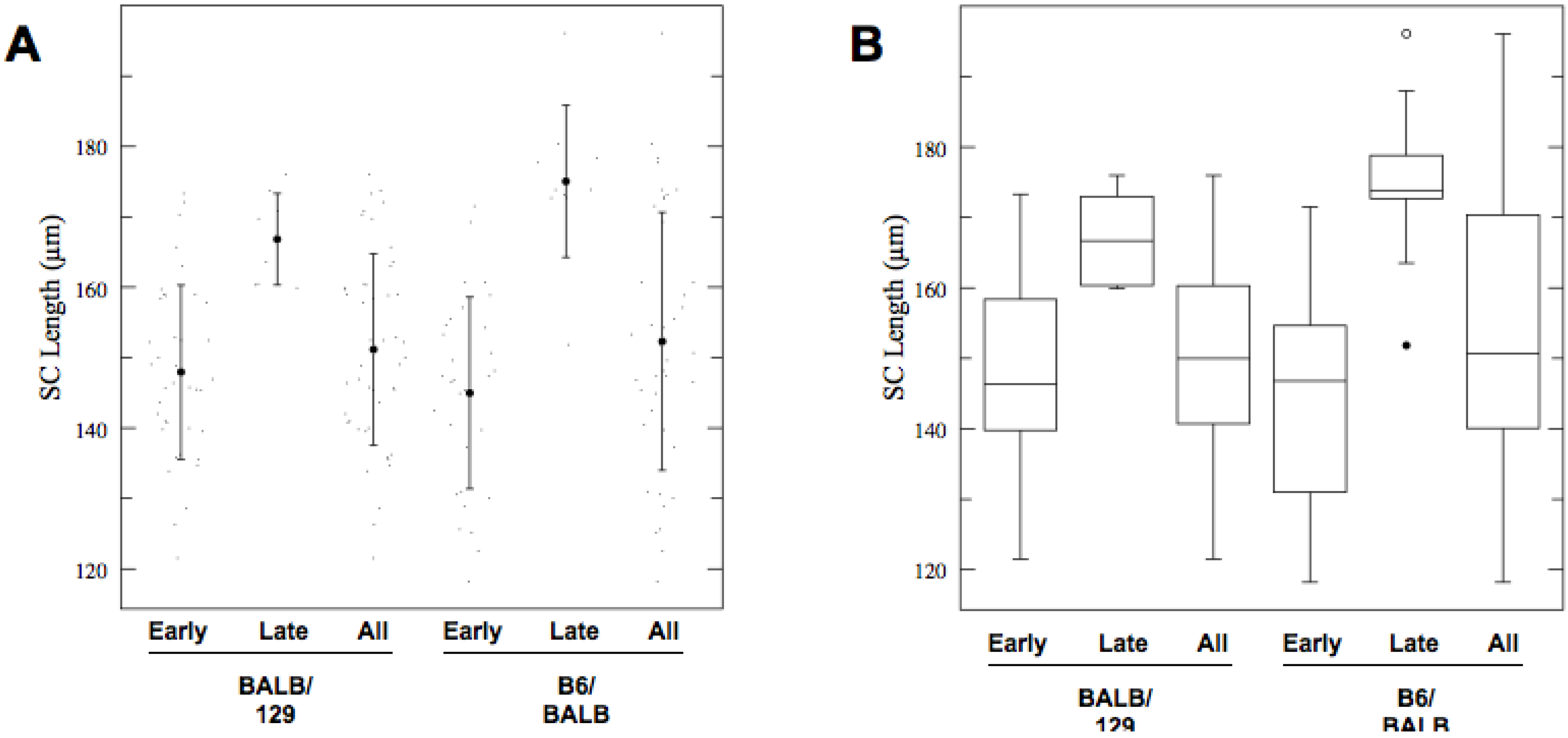
2.3. The Length of Meiotic Axes Is Potentially Determined Genetically

| Strain | n | Mean | SD | 95% CI |
|---|---|---|---|---|
| BALBc/129S4 (early) | 39 | 147.95 | 12.4 | 143.4–152.5 |
| BALBc/129S4 (late) | 8 | 166.82 | 6.51 | 156.9–176.8 |
| BALBc/129S4(all) | 47 | 151.16 | 13.6 | 147–155.3 |
| C57BL6J/BALBc (early) | 31 | 144.98 | 13.6 | 139.9–150.0 |
| C57BL6J/BALBc (late) | 10 | 175.05 | 10.8 | 166.1–184.0 |
| C57BL6J/BALBc (all) | 41 | 152.31 | 18.3 | 147.9–156.7 |
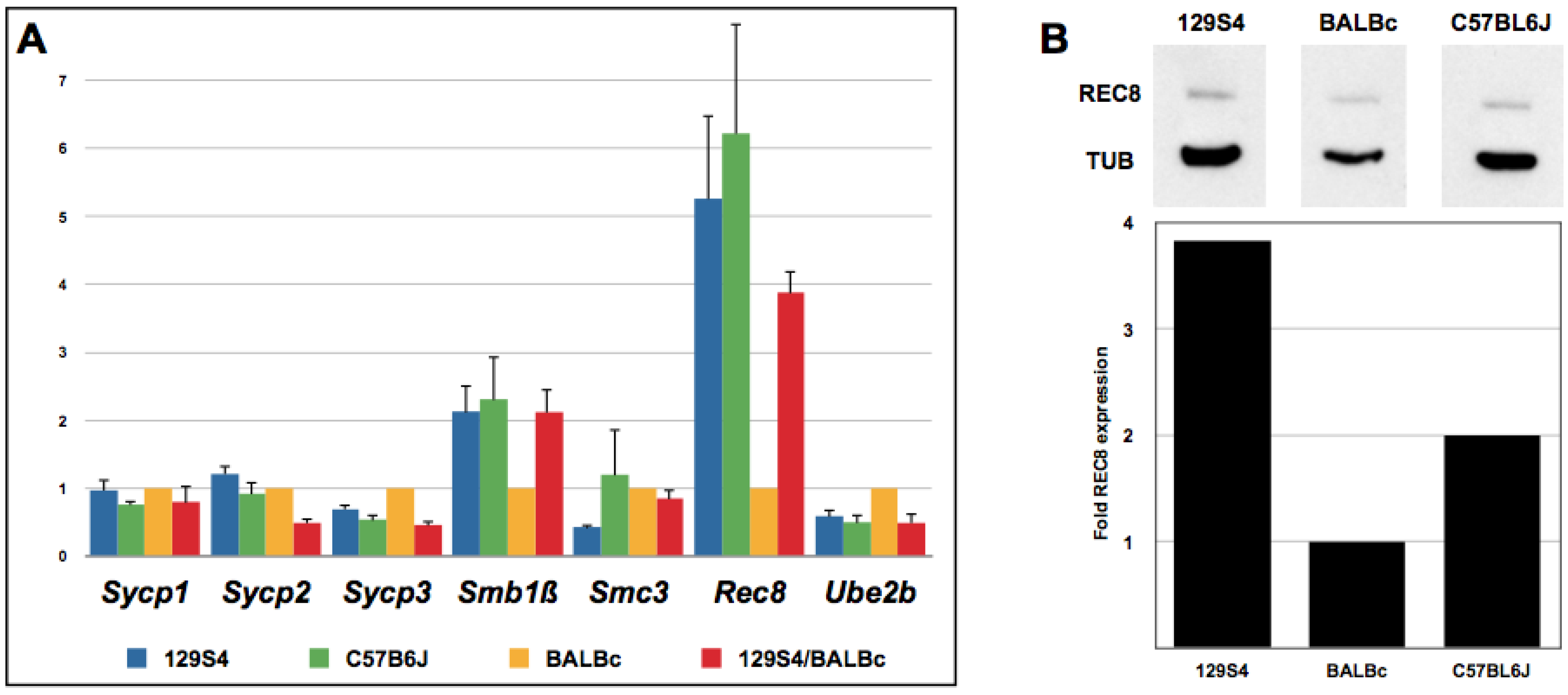
2.4. Rec8 Expression Is Reduced in BALB/c Mice

3. Discussion
4. Experimental Section
4.1. Preparation of Slides, Immunofluorescence Staining and Antibodies
4.2. Imaging and Measurements
4.3. RNA Expression Analysis
4.4. Western Blot Analysis
5. Conclusions
Acknowledgements
References
- Costa, Y.; Cooke, H.J. Dissecting the mammalian synaptonemal complex using targeted mutations. Chromosome Res. 2007, 15, 579–589. [Google Scholar]
- Moses, M.J. Structure and function of the synaptonemal complex. Genetics 1969, 61, 41–51. [Google Scholar]
- Moses, M.J.; Dresser, M.E.; Poorman, P.A. Composition and role of the synaptonemal complex. Symp. Soc. Exp. Biol. 1984, 38, 245–270. [Google Scholar]
- Page, S.L.; Hawley, R.S. The genetics and molecular biology of the synaptonemal complex. Annu. Rev. Cell Dev. Biol. 2004, 20, 525–558. [Google Scholar] [CrossRef]
- Zickler, D.; Kleckner, N. Meiotic chromosomes: integrating structure and function. Annu. Rev. Genet. 1999, 33, 603–754. [Google Scholar] [CrossRef]
- Kleckner, N. Chiasma formation: chromatin/axis interplay and the role(s) of the synaptonemal complex. Chromosoma 2006, 115, 175–194. [Google Scholar] [CrossRef]
- Fox, D.P. The control of chiasma distribution in the locust, Schistocerca gregaria (Forskal). Chromosoma 1973, 43, 289–328. [Google Scholar] [CrossRef]
- Froenicke, L.; Anderson, L.K.; Wienberg, J.; Ashley, T. Male mouse recombination maps for each autosome identified by chromosome painting. Am. J. Hum. Genet. 2002, 71, 1353–1368. [Google Scholar] [CrossRef]
- Berchowitz, L.E.; Copenhaver, G.P. Genetic intereference: Don’t stand close to me. Curr. Genomics 2010, 11, 91–102. [Google Scholar] [CrossRef]
- de Boer, E.; Dietrich, A.J.; Hoog, C.; Stam, P.; Heyting, C. Meiotic interference among MLH1 foci requires neither an intact axial element structure nor full synapsis. J. Cell Sci. 2007, 120, 731–736. [Google Scholar] [CrossRef]
- de Boer, E.; Stam, P.; Dietrich, A.J.; Pastink, A.; Heyting, C. Two levels of interference in mouse meiotic recombination. Proc. Natl. Acad. Sci. USA 2006, 103, 9607–9612. [Google Scholar] [CrossRef]
- Revenkova, E.; Jessberger, R. Keeping sister chromatids together: cohesins in meiosis. Reproduction 2005, 130, 783–790. [Google Scholar]
- Eijpe, M.; Heyting, C.; Gross, B.; Jessberger, R. Association of mammalian SMC1 and SMC3 proteins with meiotic chromosomes and synaptonemal complexes. J. Cell Sci. 2000, 113, 673–682. [Google Scholar]
- Revenkova, E.; Eijpe, M.; Heyting, C.; Gross, B.; Jessberger, R. Novel meiosis-specific isoform of mammalian SMC1. Mol. Cell Biol. 2001, 21, 6984–6998. [Google Scholar] [CrossRef]
- Revenkova, E.; Eijpe, M.; Heyting, C.; Hodges, C.A.; Hunt, P.A.; Liebe, B.; Scherthan, H.; Jessberger, R. Cohesin SMC1 beta is required for meiotic chromosome dynamics, sister chromatid cohesion and DNA recombination. Nat. Cell Biol. 2004, 6, 555–562. [Google Scholar] [CrossRef]
- Eijpe, M.; Offenberg, H.; Jessberger, R.; Revenkova, E.; Heyting, C. Meiotic cohesin REC8 marks the axial elements of rat synaptonemal complexes before cohesins SMC1beta and SMC3. J. Cell Biol. 2003, 160, 657–670. [Google Scholar]
- Prieto, I.; Suja, J.A.; Pezzi, N.; Kremer, L.; Martinez, A.C.; Rufas, J.S.; Barbero, J.L. Mammalian STAG3 is a cohesin specific to sister chromatid arms in meiosis I. Nat. Cell Biol. 2001, 3, 761–766. [Google Scholar] [CrossRef]
- Kouznetsova, A.; Novak, I.; Jessberger, R.; Hoog, C. SYCP2 and SYCP3 are required for cohesin core integrity at diplotene but not for centromere cohesion at the first meiotic division. J. Cell Sci. 2005, 118, 2271–2278. [Google Scholar] [CrossRef]
- Yang, F.; de La Fuente, R.; Leu, N.A.; Baumann, C.; McLaughlin, K.J.; Wang, P.J. Mouse SYCP2 is required for synaptonemal complex assembly and chromosomal synapsis during male meiosis. J. Cell Biol. 2006, 173, 497–507. [Google Scholar] [CrossRef]
- Yuan, L.; Liu, J.G.; Zhao, J.; Brundell, E.; Daneholt, B.; Hoog, C. The murine SCP3 gene is required for synaptonemal complex assembly, chromosome synapsis, and male fertility. Mol. Cell 2000, 5, 73–83. [Google Scholar] [CrossRef]
- Liu, J.G.; Yuan, L.; Brundell, E.; Bjorkroth, B.; Daneholt, B.; Hoog, C. Localization of the N-terminus of SCP1 to the central element of the synaptonemal complex and evidence for direct interactions between the N-termini of SCP1 molecules organized head-to-head. Exp. Cell Res. 1996, 226, 11–19. [Google Scholar] [CrossRef]
- Schmekel, K.; Meuwissen, R.L.; Dietrich, A.J.; Vink, A.C.; van Marle, J.; van Veen, H.; Heyting, C. Organization of SCP1 protein molecules within synaptonemal complexes of the rat. Exp. Cell Res. 1996, 226, 20–30. [Google Scholar] [CrossRef]
- Hamer, G.; Wang, H.; Bolcun-Filas, E.; Cooke, H.J.; Benavente, R.; Hoog, C. Progression of meiotic recombination requires structural maturation of the central element of the synaptonemal complex. J. Cell Sci. 2008, 121, 2445–2451. [Google Scholar] [CrossRef]
- Bolcun-Filas, E.; Costa, Y.; Speed, R.; Taggart, M.; Benavente, R.; de Rooij, D.G.; Cooke, H.J. SYCE2 is required for synaptonemal complex assembly, double strand break repair, and homologous recombination. J. Cell Biol. 2007, 176, 741–747. [Google Scholar] [CrossRef]
- Costa, Y.; Speed, R.; Ollinger, R.; Alsheimer, M.; Semple, C.A.; Gautier, P.; Maratou, K.; Novak, I.; Hoog, C.; Benavente, R.; Cooke, H.J. Two novel proteins recruited by synaptonemal complex protein 1 (SYCP1) are at the centre of meiosis. J. Cell Sci. 2005, 118, 2755–2762. [Google Scholar] [CrossRef]
- Blat, Y.; Protacio, R.U.; Hunter, N.; Kleckner, N. Physical and functional interactions among basic chromosome organizational features govern early steps of meiotic chiasma formation. Cell 2002, 111, 791–802. [Google Scholar] [CrossRef]
- Moens, P.B.; Pearlman, R.E. Chromatin organization at meiosis. Bioessays 1988, 9, 151–153. [Google Scholar] [CrossRef]
- Moens, P.B.; Pearlman, R.E.; Heng, H.H.; Traut, W. Chromosome cores and chromatin at meiotic prophase. Curr. Top Dev. Biol. 1998, 37, 241–262. [Google Scholar]
- Moens, P.B.; Pearlman, R.E. Visualization of DNA sequences in meiotic chromosomes. Methods Cell Biol. 1991, 35, 101–108. [Google Scholar] [CrossRef]
- Hernandez-Hernandez, A.; Rincon-Arano, H.; Recillas-Targa, F.; Ortiz, R.; Valdes-Quezada, C.; Echeverria, O.M.; Benavente, R.; Vazquez-Nin, G.H. Differential distribution and association of repeat DNA sequences in the lateral element of the synaptonemal complex in rat spermatocytes. Chromosoma 2008, 117, 77–87. [Google Scholar] [CrossRef]
- Moens, P.B.; Pearlman, R.E. Telomere and centromere DNA are associated with the cores of meiotic prophase chromosomes. Chromosoma 1990, 100, 8–14. [Google Scholar] [CrossRef]
- Pearlman, R.E.; Tsao, N.; Moens, P.B. Synaptonemal complexes from DNase-treated rat pachytene chromosomes contain (GT)n and LINE/SINE sequences. Genetics 1992, 130, 865–872. [Google Scholar]
- Kleckner, N.; Storlazzi, A.; Zickler, D. Coordinate variation in meiotic pachytene SC length and total crossover/chiasma frequency under conditions of constant DNA length. Trends Genet. 2003, 19, 623–628. [Google Scholar] [CrossRef]
- Lynn, A.; Koehler, K.E.; Judis, L.; Chan, E.R.; Cherry, J.P.; Schwartz, S.; Seftel, A.; Hunt, P.A.; Hassold, T.J. Covariation of synaptonemal complex length and mammalian meiotic exchange rates. Science 2002, 296, 2222–2225. [Google Scholar] [CrossRef]
- Tres, L.L. Extensive pairing of the XY bivalent in mouse spermatocytes as visualized by whole-mount electron microscopy. J. Cell Sci. 1977, 25, 1–15. [Google Scholar]
- Wold, M.S. Replication protein A: a heterotrimeric, single-stranded DNA-binding protein required for eukaryotic DNA metabolism. Annu. Rev. Biochem. 1997, 66, 61–92. [Google Scholar] [CrossRef]
- Plug, A.W.; Peters, A.H.; Xu, Y.; Keegan, K.S.; Hoekstra, M.F.; Baltimore, D.; de Boer, P.; Ashley, T. ATM and RPA in meiotic chromosome synapsis and recombination. Nat. Genet. 1997, 17, 457–461. [Google Scholar] [CrossRef]
- Moens, P.B.; Kolas, N.K.; Tarsounas, M.; Marcon, E.; Cohen, P.E.; Spyropoulos, B. The time course and chromosomal localization of recombination-related proteins at meiosis in the mouse are compatible with models that can resolve the early DNA-DNA interactions without reciprocal recombination. J. Cell Sci. 2002, 115, 1611–1622. [Google Scholar]
- Hillers, K.J. Crossover interference. Curr. Biol. 2004, 14, R1036–R1037. [Google Scholar] [CrossRef]
- Bannister, L.A.; Reinholdt, L.G.; Munroe, R.J.; Schimenti, J.C. Positional cloning and characterization of mouse mei8, a disrupted allelle of the meiotic cohesin Rec8. Genesis 2004, 40, 184–194. [Google Scholar] [CrossRef]
- Xu, H.; Beasley, M.D.; Warren, W.D.; van der Horst, G.T.; McKay, M.J. Absence of mouse REC8 cohesin promotes synapsis of sister chromatids in meiosis. Dev. Cell 2005, 8, 949–961. [Google Scholar] [CrossRef]
- Baarends, W.M.; Wassenaar, E.; Hoogerbrugge, J.W.; van Cappellen, G.; Roest, H.P.; Vreeburg, J.; Ooms, M.; Hoeijmakers, J.H.; Grootegoed, J.A. Loss of HR6B ubiquitin-conjugating activity results in damaged synaptonemal complex structure and increased crossing-over frequency during the male meiotic prophase. Mol. Cell Biol. 2003, 23, 1151–1162. [Google Scholar] [CrossRef]
- Brar, G.A.; Hochwagen, A.; Ee, L.S.; Amon, A. The multiple roles of cohesin in meiotic chromosome morphogenesis and pairing. Mol. Biol. Cell 2009, 20, 1030–1047. [Google Scholar]
- Klein, F.; Mahr, P.; Galova, M.; Buonomo, S.B.; Michaelis, C.; Nairz, K.; Nasmyth, K. A central role for cohesins in sister chromatid cohesion, formation of axial elements, and recombination during yeast meiosis. Cell 1999, 98, 91–103. [Google Scholar] [CrossRef]
- Storlazzi, A.; Tesse, S.; Ruprich-Robert, G.; Gargano, S.; Poggeler, S.; Kleckner, N.; Zickler, D. Coupling meiotic chromosome axis integrity to recombination. Genes Dev. 2008, 22, 796–809. [Google Scholar]
- Peters, A.H.; Plug, A.W.; van Vugt, M.J.; de Boer, P. A drying-down technique for the spreading of mammalian meiocytes from the male and female germline. Chromosome Res. 1997, 5, 66–68. [Google Scholar] [CrossRef]
- Baart, E.B.; de Rooij, D.G.; Keegan, K.S.; de Boer, P. Distribution of Atr protein in primary spermatocytes of a mouse chromosomal mutant: A comparison of preparation techniques. Chromosoma 2000, 109, 139–147. [Google Scholar] [CrossRef]
- Rosen, S.; Skaletsky, H.J. Primer3 on the WWW for general users and for biologist programmers. In Bioinformatics Methods and Protocols Methods in Molecular Biology; Krawets, S., Misener, S., Eds.; Humana Press: Totowa, NJ, USA, 2010; pp. 365–386. [Google Scholar]
© 2010 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Vranis, N.M.; Van der Heijden, G.W.; Malki, S.; Bortvin, A. Synaptonemal Complex Length Variation in Wild-Type Male Mice. Genes 2010, 1, 505-520. https://doi.org/10.3390/genes1030505
Vranis NM, Van der Heijden GW, Malki S, Bortvin A. Synaptonemal Complex Length Variation in Wild-Type Male Mice. Genes. 2010; 1(3):505-520. https://doi.org/10.3390/genes1030505
Chicago/Turabian StyleVranis, Neil M., Godfried W. Van der Heijden, Safia Malki, and Alex Bortvin. 2010. "Synaptonemal Complex Length Variation in Wild-Type Male Mice" Genes 1, no. 3: 505-520. https://doi.org/10.3390/genes1030505