The Little Fly that Could: Wizardry and Artistry of Drosophila Genomics

1
VIB Center for the Biology of Disease, VIB, 3000 Leuven, Belgium
2
Center for Human Genetics, University of Leuven School of Medicine, 3000 Leuven, Belgium
*
Authors to whom correspondence should be addressed.
Genes 2014, 5(2), 385-414; https://doi.org/10.3390/genes5020385
Submission received: 3 March 2014
/
Revised: 16 April 2014
/
Accepted: 21 April 2014
/
Published: 13 May 2014
(This article belongs to the Special Issue Grand Celebration: 10th Anniversary of the Human Genome Project)
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:For more than 100 years now, the fruit fly Drosophila melanogaster has been at the forefront of our endeavors to unlock the secrets of the genome. From the pioneering studies of chromosomes and heredity by Morgan and his colleagues, to the generation of fly models for human disease, Drosophila research has been at the forefront of genetics and genomics. We present a broad overview of some of the most powerful genomics tools that keep Drosophila research at the cutting edge of modern biomedical research.
1. Introduction
The Human Genome Project, on its way to producing an assembled genome of Homo sapiens, has gone through several test runs yielding sequenced genomes of other organisms of high relevance for research into human development and disease. The first published genome of a free-living organism was that of the proteobaterium Haemophilus influenzae [1], followed by sequencing of the genome of Saccharomyces cerevisiae yeast, the first eukaryotic genome sequenced [2], and the genome of Caenorhabditis elegans, the first genome of a multicellular organism and the first animal genome [3]. The second animal genome sequenced was that of the fruit fly Drosophila melanogaster [4]. In this review, we discuss the significance of the sequencing of the Drosophila genome as well as the technical advances and new research avenues that have accompanied it.
2. Drosophila as a Model
2.1. In Development
The fruit fly has been studied for over a century and the lessons learned from fly research makes it almost impossible to enumerate but a few of the most notable cases. The pioneering studies that identified genes involved in Drosophila embryo segmentation [5,6] and establishment of segment polarity [6] were seminal for understanding conserved developmental strategies in the animal kingdom. The discovery of homeotic genes is one of the best-known examples of genes discovered in the fruit fly, and these were found to be conserved and play analogous roles in humans [7,8,9]. Drosophila has played a seminal role in sensory organ development research. The discovery of the eyeless gene [10], a fly homolog of human and mouse PAX6 [11,12], and determination of its targets [13] shed light on vertebrate eye development and led to discovery of novel disease related genes in humans [14]. The proneural gene atonal plays a crucial role in the development of Drosophila photoreceptor neurons [15] and chordotonal organs [16]. Its function is conserved in mammals, where its homologs Math5 and Math1 were shown to be involved in regulating formation of retinal ganglion cells [17] and inner ear mechanosensory hair cells [18].
2.2. In Signaling
Drosophila has been extensively used for studies of signaling pathways. In Hedgehog signaling, both the Hedgehog ligand itself [6,19,20] and its receptor Patched [6,21,22] were first identified in the fly, though the link between the two was first established in mammals [23,24]. The ligand of the Wnt signaling pathway turned out to be a well-known Drosophila segment polarity protein, i.e., Wingless. The Wnt receptor, Frizzled [25], and several other signal transduction cascade members were identified in the fly as members of the Wnt pathway [26,27,28]. The planar cell polarity (PCP) pathway is yet another example of a signaling cascade in which key players and mechanisms of action have been, to a large extend, identified in Drosophila [29,30]. The Notch signaling pathway, associated with cell fate control, lateral inhibition, and signal integration during development, has been discovered and extensively studied in fruit flies [31,32,33]. Finally, major components and mechanisms of action of the Hippo signaling pathway have been described in Drosophila [34,35,36]. All these pathways play major roles in human development and disease.
2.3. In Disease
Over the past two decades the fruit fly became an increasingly popular model organism for the study of human disease, with focus on neurodegenerative [37] and neuromuscular [38] diseases as well as cancer [39]. Neurological diseases that have been modeled in Drosophila include trinucleotide repeat disorders [40,41,42], Alzheimer’s disease [43,44,45,46], Parkinson’s disease [47,48], amyotrophic lateral sclerosis [49,50], and dystrophy [51]. Other examples that include use of the fruit fly model are studies of alcohol abuse [52,53], cocaine addiction [54], obesity [55] and diabetes [56], cardiac diseases [57], and asthma [58]. Drosophila has been demonstrated to be a great model to identify tumor suppressor genes [59] or genes involved in metastasis [60]. Thanks to the conservation of major signaling pathways, tumor suppressors and oncogenes, various fly cancer models have been established. Understanding how signal transduction pathways like Hippo, Notch, Dpp or JAK-STAT affect tumor formation was aided by research in fruit flies [61,62,63]. Drosophila has been used as a model for tumor invasion and metastasis [64], and as a platform to identify novel therapeutic targets [65].
3. Meet the Drosophila Genome
The Drosophila genome is estimated to be approximately 200 Mb, with one third of it forming pericentric heterochromatin [66]. It is organized on three autosomes (numbered 2, 3 and 4) and sex chromosomes, X (also referred to as the first chromosome) and Y. The initial assembly of the fruit fly genome was published in March 2000, after almost a year of whole genome shotgun sequencing. The first published assembly, referred to as Release 1 of the genome, included 13,991 genes encoding for 14,080 peptides. Over two thirds of annotated genes were assigned gene ontology (GO) terms upon annotation. The initial assembly contained ~1300 gaps in mapped sequences [4] that were filled with subsequent releases.
The third release of the genome was the first that included pericentric heterochromatin sequences [67]. The mutations indicated in the sequenced strain’s genotype, as well as several other identified mutations, have been corrected with wild-type sequence [68]. With that release, a comprehensive set of resources were published, including a library of full-length cDNAs for 40% of genes [69] and an atlas of gene expression patterns during embryogenesis [70]. Sequence analysis provided insights into transposable elements within the genome [71], core promoter structures [72], and largely improved annotation of gene models [68].
The current, fifth assembly of the genome has closed all but 9 gaps in the main assembly. The sequenced genome covers over 120 Mb of euchromatin, and over 9 Mb of mapped and over 10 Mb of unmapped heterochromatin. The current annotation revision contains 13,942 protein coding genes and over 2354 non-coding RNA genes, including ribosomal (rRNAs), transport (tRNAs), micro- (miRNAs), and small nuclear (snRNA) and small nucleolar (snoRNA) RNAs [73]. Through genome analysis, fruit flies have been found to contain complex gene structures. Approximately 7.5% of all genes, including non-coding RNAs, are located within the introns of other genes. Messenger RNAs for about 15% of genes overlap with mRNAs of genes on the opposite strands. Over 30 genes have been identified as dicistronic, i.e., producing single mRNA encoding for two separate protein products through independent translation initiation events [68]. Over 30% of Drosophila melanogaster genes were found to be alternatively spliced [74], yielding a diverse set of almost 30,000 protein-coding transcripts [73]. The next release of the genome assembly (Release 6) is expected this year (2014).
Improved assembly and annotation of the fruit fly genome was possible not only due to new sequencing data, but also thanks to advances in bioinformatics tools. An integrated computational pipeline and a tailored database schema have been developed to facilitate genomic data storage and automated sequence annotation [75]. Computed annotations have been manually curated by experts and to aid in this task, a dedicated annotation editor was developed [76]. Finally, automated genome annotation in general requires the use of computational tools, some of which were first applied in the Drosophila genome project [72,77].
4. Genomes by the Dozen
Analysis of coding parts of the genome can be facilitated by comparison of genomic sequences with sequences of cDNAs originating from the same species. Most of the DNA in the majority of species, however, is non-coding. One approach to identify functional non-coding DNA segments, such as cis-regulatory elements, relies on finding conserved regions or motifs across related species. This naturally requires having more than one genome sequenced and was a driving force behind sequencing of the genomes of Schizosaccharomyces pombe [78] and Caenorhabditis briggsae [79] in the yeast and worm research communities, respectively. In the Drosophila genus, the comparative genomics era began with sequencing of the Drosophila pseudoobscura genome [80]. The two genomes were found to be very similar, despite 25–55 million years of evolutionary divergence. Synteny is preserved in blocks containing 10.7 genes on average, which corresponds to ~83 kb. The vast majority of synteny breaks were caused by intrachromosomal rearrangements. On average, ~48% of the base pairs are conserved between these two species.
The next advance in Drosophila comparative genomics came with sequencing of ten further species, Drosophila sechellia, simulans, yakuba, erecta, ananassae, persimilis, willistoni, mojavensis, virilis, and grimshawi. These species span a broad spectrum of morphologies, ecologies, and behaviors, yet have identical body plans and very similar life cycles [81]. Furthermore, these species share approximately 70% of their genes. Genome sizes estimated by flow cytometry vary between 130 Mb in D. mojavensis to 364 Mb in D. virilis [66]. The synteny conservation between sequenced species varies with an average of 122 genes per block between D. melanogaster and simulans down to 8 genes per block between D. melanogaster and grimshawi. Overall genome size, number of genes, distribution of transposable element classes, and patterns of codon usage are all very similar across the 12 sequenced genomes. At a finer scale, however, the number of structural changes and rearrangements is larger, including rearrangements of genes within the Hox cluster or highly dynamic sizes and content of multigene families [81].
Together, the 12 Drosophila genomes provide a solid platform for annotation and analysis of both coding and non-coding DNA. This unprecedented dataset enabled the use of evolutionary signatures—specific patterns of change in DNA elements upon selection—for de novo prediction and correction of previously annotated protein-coding gene models [82], non-coding RNAs, and transcription factor (TF) binding sites [83]. Identification of TF binding motifs has traditionally been based on DNA alignments. Alignment-based methods can also be used for the identification of cis-regulatory modules (CRMs), which are comprised of a number of TF binding motifs [84]. In many cases, however, the number and order of individual motifs varies between species, especially when these are distant, while preserving regulatory outcome. To address such cases, alignment-free approaches have also been developed [85,86].
5. Genomes by Population
Drosophila provides an unmatched set of resources for studying quantitative traits [87]. In the post genomic era, genome-wide association studies (GWAS) have become a preferred method for analyzing complex traits. The GWAS methodology is now routinely and successfully applied in the identification of human disease-associated genes [88]. Two fruit fly resources, the Drosophila Genetic Reference Panel (DGRP) [89] and the Drosophila Synthetic Population Resource (DSPR) [90], offer large sets of sequenced and mapped fly lines tailored for GWAS and quantitative trait loci (QTL) mapping.
The DGRP is a collection of more than 200 fully sequenced recombinant inbred lines (RILs) that were established from mated females collected from a market in Raleigh, North Carolina, USA. The genomic sequences of these lines contain over 4.5 million single nucleotide polymorphisms (SNPs), over one hundred thousand polymorphic microsatellites, and over 36 thousand transposable elements [89]. The DGRP has been extensively characterized and in addition to detailed genomic sequence analysis includes microarray [91] and RNA-seq [92] datasets for selected lines. To date, numerous genome-wide association studies have been published on various traits using DGRP, including oxidative stress [93], mitochondrial function [94], viral infection resistance [95], and sleep [96].
The Drosophila Synthetic Population Resource uses a different approach. Over 1,700 DSPR RILs were established from 15 isogenic founder lines created from geographically distinct Drosophila populations. The founder lines were split in two groups of eight (with one line in both groups) and mixed for 50 generations to create two synthetic populations, from which two sets of RILs were established. The founder lines were fully sequenced and each RIL was mapped using restriction-site associated DNA (RAD) markers onto the founders’ sequence with 17 kb median resolution. The number of SNPs in the founder lines exceeds 1.6 million [90]. The DSPR is complementary to DGRP and both resources can be used together for cross-validation and to increase the mapping power [97].
6. Decoding the Genome’s Secrets
6.1. The modENCODE Project
The information encoded by genomes goes far beyond a simple trinucleotide code used to translate nucleic acid sequence into protein. A plethora of information is hidden within introns, UTRs, non-coding RNAs, cis-regulatory elements, and chromatin marks. These elements are known to regulate where and when a gene product is expressed. The human ENCODE (ENCyclopedia Of DNA Elements) project [98] aims to identify and understand the information carried by the human genome. The modENCODE project is the model organism counterpart of ENCODE with focus on two species C. elegans [99] and D. melanogaster [100]. The fruit fly modENCODE data includes high-throughput transcriptome sequencing (RNA-seq), chromatin immunoprecipitation followed by sequencing (ChIP–seq) for transcription factor binding sites and histone modifications, DNA replication patterns, and nucleosome occupancy. The samples have been collected from 12–30 developmental time points of the sequenced D. melanogaster strain and from several cell lines [100].
6.2. The Transcriptome
Comprehensive transcriptomics data has redefined gene models for 75% of fly genes by adding new exons or splice variants. The majority of annotation changes were supported by direct cDNA evidence. Analysis of transcription start sites (TSSs) for over half of Drosophila genes resulted in identification of over 1500 novel promoters. The structural analysis of RNA-seq-identified transcripts that did not seem to encode proteins revealed that a majority of them has no thermodynamically stable secondary structure, suggesting structure-independent functions. Among structural non-coding RNAs, several hundred novel small regulatory RNAs (miRNAs, siRNAs, and piRNA) have been identified. Additionally, transcription start sites for both protein coding and non-coding RNAs have been derived from the presence of chromatin marks characteristic of transcriptionally active regions, such as H3K4me3 enrichment, H3K9ac, and presence of RNA polymerase II in TSS-proximal regions [100].
6.3. Chromatin Landscape
Eukaryotic genomes are organized into large domains that exhibit distinct chromatin properties [100]. Analysis of large-scale organization of the chromatin landscape has revealed unexpected complexity and plasticity among different cell types. Some regions in the usually silent pericentric heterochromatin exhibited surprisingly high gene expression activity. Conversely, large regions of normally transcriptionally active euchromatin harbored histone marks (H3K9me2) typical for heterochromatin [100,101]. Chromatin signatures characteristic of various functional elements have been identified by ChIP-chip for 18 histone modifications (both activating, such as H3K4me, H3K9/18/27ac, H2B ubiquitination and repressive, such as H3K9me2/3 or Polycomb associated H3K27me3) and variants (H1, H4) from several cell lines and developmental stages. Correlating chromatin signatures with transcriptome and protein binding data (replication factors, insulator-binding proteins, and transcription factors) helped identify marks specific for promoters, actively transcribed regions, introns, insulators, and origins of replication [100]. The presence of specific chromatin marks was found to correlate with the physical properties of chromatin, where transcriptionally active chromatin exhibited high solubility and high nucleosome-turnover rates [100]. Computational analysis of combinatorial patterns of histone modifications revealed distinct chromatin states associated with active TSSs, exons, introns, and other open chromatin as well as closed chromatin states [100,102,103].
6.4. Transcriptional Regulation
The modENCODE project has identified binding sites for almost 40 transcription factors through both ChIP-chip and ChIP-seq. The analysis has revealed that out of nearly 40,000 identified unique binding sites found, 5% are bound by 8 different transcription factors or more and are considered High Occupancy Target (HOT) regions. Furthermore, almost 40% of the sites can be bound by more than two factors [103]. The HOT regions exhibit decreased nucleosome density, increased nucleosome turnover and often colocalize with TSS and ORC (origin recognition complex) binding sites, suggesting interplay between chromatin regulation, TF binding, and DNA replication [100,103]. In total, modENCODE ChIP experiments revealed over 500 silencers, 2300 new promoters, over 14 candidate CBP-bound cis-regulatory elements, and over 7500 putative insulators [103]. Pairwise analysis of binding site co-occurrence has revealed over 800 known and putative transcription factor co-binding interactions. Binding sites for transcription factors regulating biologically opposing roles exhibited negative associations. The modENCODE TF binding data sets combined with external data were used to construct a network covering over 80 transcription factors and characterizing over 800, largely novel, regulatory interactions. Binding site co-occurrence among various analyzed promoters corresponded to temporal co-expression of the respective target genes, supporting the existence of combinatorial transcription factor codes [103].
7. The Fruit Fly Toolkit
7.1. Getting Constructs in
Drosophila is famous for its extensive range of forward and reverse genetics tools. The powerful toolkit was primed by the discovery of P transposable element-based germline transformation [104]. This revolutionary development allowed, for the first time, efficient delivery of foreign DNA into the genome. Development of the vast majority of Drosophila tools required at some stage use of P-element or other transposon systems. P-elements were used for gene cloning [105], genetic rescue [106], and as potent mutagens by their insertion [107] or excision [108]. P-element insertions have enabled the creation of enhancer traps, thus allowing visualization of gene expression patterns using genetically encoded reporters [109]. The Drosophila Gene Disruption Project used P-elements to create single transposon insertions in over 30% of fly genes [110]. The remaining genes, due to target sequence bias of P-elements, are currently targeted using other transposons [111]. The catalog of transposons that can be used for fly transformation has been expanded over the years and includes mariner [112,113], Minos [114,115], and piggyBac [116,117]. Each of these transposable elements, except for Minos that seems to insert randomly, has its hot and cold spots, but failure to target a certain region can often be addressed by using a different transposon [111].
While random, transposon-mediated transgenesis is desirable for gene disruption or genomic targeting, but integration of reporter or rescue constructs calls for more control over the locus where these integrate, thus reducing the chance of position effects that can strongly influence gene expression [118,119]. Early attempts to repeatedly target a specific locus in the fruit fly genome were based on transposon homing [120]. Short regulatory sequences from Polycomb target genes or from the linotte locus included in the transposon were shown to increase the likelihood of such transposon landing in the vicinity of genomic regions bearing these sequences [120,121]. The low resolution (30 bp) and efficiency (20% of insertions) of this homing technique prompted further developments in the field. The introduction of an irreversible, site-specific recombinase from the phiC31 phage ushered in a new era in fly transgenesis. The phiC31 integrase catalyzes unidirectional recombination between two attachment sites, attP and attB, leading to the formation of attL and attR sites [122]. A circular construct harboring an attB site can be efficiently and specifically integrated into an attP site located on the genome [123]. The phiC31 integrase system has, for the first time, enabled transformation of flies with BAC-sized constructs [124]. The integrase can be expressed from mRNA co-injected with the construct [123] or from the genome under a germ-line specific promoter, the latter method being more efficient [125]. Several dozens of attP landing lines have been created and tested [123,124,125,126], creating unlimited possibilities to combine transgenes.
7.2. Express What You Want, Where You Want, and When You Want
P-element transgenesis has enabled the creation of a plethora of other Drosophila tools, of which the Gal4/UAS system is the most notable example. The system is based on yeast transcription factor gene GAL4 fused to a minimal promoter. This construct is randomly inserted into the genome, for instance by means of P-element transgenesis, hitchhiking nearby enhancers and creating an enhancer trap. Alternatively, enhancer sequences can be cloned upstream of the minimal promoter and other arrangements, with enhancers cloned elsewhere, for example within introns, are also possible. The second component of the system is the Gal4 binding site, known as the upstream activation sequence (UAS), driving expression of the target gene. The combination of a GAL4 enhancer trap with a UAS-driven target enables expression of the gene of interest in the desired tissues or cell types [127]. Several collections of GAL4 enhancer traps have been created using P-element [128,129] and piggyBac [110,130] insertions. The enhancer trap resources have recently been supplemented by large collections of cloned enhancers driving expression of Gal4 [131,132]. The Gal4 expression pattern can be refined spatially [133] or temporarily [134] using the Gal80 repressor [135]. Further control over Gal4-driven expression can be obtained using variants requiring drugs for activation [136,137,138,139]. Today, the Gal4-UAS system is one of several binary expression systems available in Drosophila. Other examples include the LexA transactivator that binds LexOp sites [140] and the Q system with QF transactivator, QUAS binding sites and the QS repressor whose activity can additionally be drug controlled [141]. The existing binary systems can be combined to provide fine control over target expression pattern or for simultaneous targeting of different cellular populations [142].
7.3. Mutant Tissue on Demand
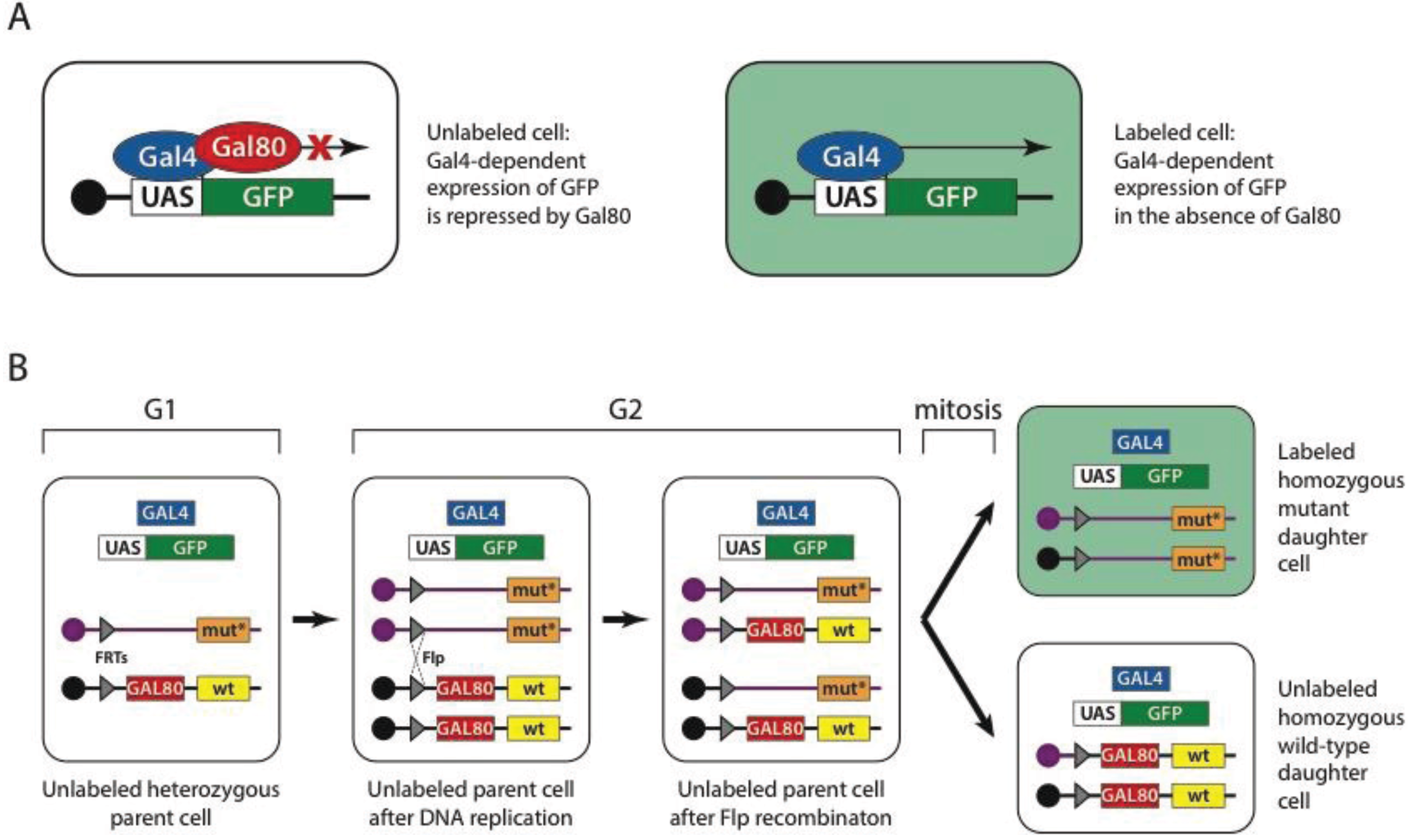
The Gal4-UAS system is an important component of yet another powerful fruit fly tool, the mosaic analysis with a repressible cell marker or MARCM. Induction of mosaicism in Drosophila is used either for studying an otherwise lethal phenotype within a tissue of interest [143] or for marking a clone of cells within a tissue of interest [144]. Mosaics can be created using flippase (Flp) mediated mitotic recombination between homologous chromosomes [145]. In this technique, homologous chromosomes carry an insertion (usually P-element mediated) of a flippase recognition target (FRT) site. One of the chromosomes carries a wild type and the other a mutant allele of the gene of interest. In the presence of flippase, recombination events between homologous chromosomes can occur during cell division, leading to the generation of homozygous mutant cells from heterozygous precursors. The MARCM technique (Figure 1) enhances Flp-mediated mitotic recombination by uniquely labeling mutant cells using a genetically encoded marker. The mutant clone is marked with a UAS-GFP (green fluorescent protein) construct, driven by ubiquitously expressed GAL4. These two transgenes are usually inserted together on any chromosome, except the wild type chromosome that carries the Gal4 repressor—GAL80 under the control of a ubiquitous promoter. The presence of the Gal4 repressor on the wild-type chromosome prevents GFP expression in both heterozygous and homozygous wild type cells [135]. The MARCM technique was later extended to label wild type cells as well [146].
MARCM, among other mitotic-recombination-based approaches, has enabled the creation of tissue specific mutant cells for genes where a mutant exists. This, however, is not yet [111,147] the case for all fruit fly genes. Post-transcriptional gene silencing by double-stranded RNA (dsRNA) [148], commonly known as RNA interference (RNAi), allows the silencing of virtually any transcript encoded by the genome [149]. Drosophila is not only one of the first organisms where RNAi has been used to silence genes [150,151], but it has also played an important role in studying the mechanism of dsRNA dependent gene silencing [152]. Injections of dsRNA into the Drosophila embryos were used to pioneer RNAi in the fruit fly. However, this mode of delivery has limited use for studying gene function in the late stages of development or in a tissue specific manner. A combination of genetically encoded hairpin-loop RNAs with a Gal4/UAS system has been introduced to address these issues and place Drosophila RNAi under spatio-temporal control [153]. Efficient transformation techniques developed for Drosophila Schneider (S2) cells [154] combined with Drosophila dsRNA libraries allowed RNAi screens in cell culture on a genome-wide scale [155,156,157,158]. With genome-wide libraries of fly lines carrying UAS-driven hairpin RNAs, tissue-specific RNAi screens in the whole animals became possible [159,160]. While the first library used P-element for transgenesis, thus leading to variability of hairpin RNA expression levels in different lines, the next generation of libraries followed, using phiC31-mediated insertions into a defined locus [161,162]. RNAi in flies has proven very effective and allowed for a number of large scale screens to be performed, including ones targeting muscle development [162], heart function [163], obesity [164], pain [165], glial function [166], or piRNA pathways [167]. The off-target effect, a well-known pitfall of RNA interference, has been addressed in flies by specificity control using either cross-species rescue [168,169] or engineered RNAi-refractory transgenes [170].
7.4. Bright Rescue
Modern classical and reverse-genetic approaches often call for reliable sources of transgenes, both to induce new and rescue induced phenotypes. Classically, clones from cDNA libraries [69] combined with the Gal4/UAS system [127] have been used to specifically express a gene of interest in target tissue. These constructs could be used either to ectopically express a gene of interest [127], rescue a mutant phenotype [171], or by using a fusion of cDNA with a fluorescent protein coding sequence to visualize the localization of a protein of interest [172]. These approaches, however, do not allow simultaneous modification, such as introduction of point mutations, truncation, tagging, and expression of a protein of interest under native or nearly native control. This usually requires a larger genomic context.
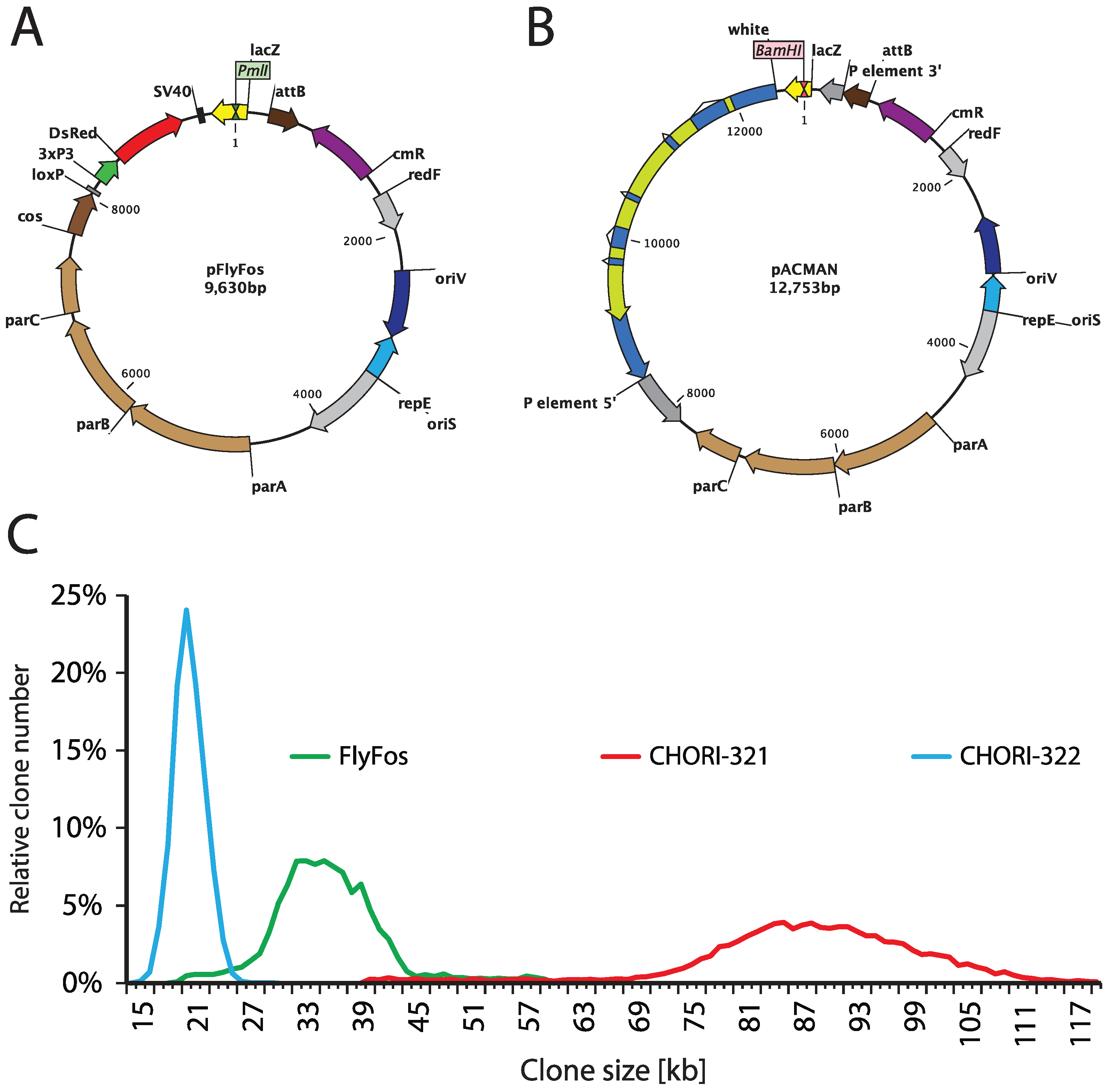
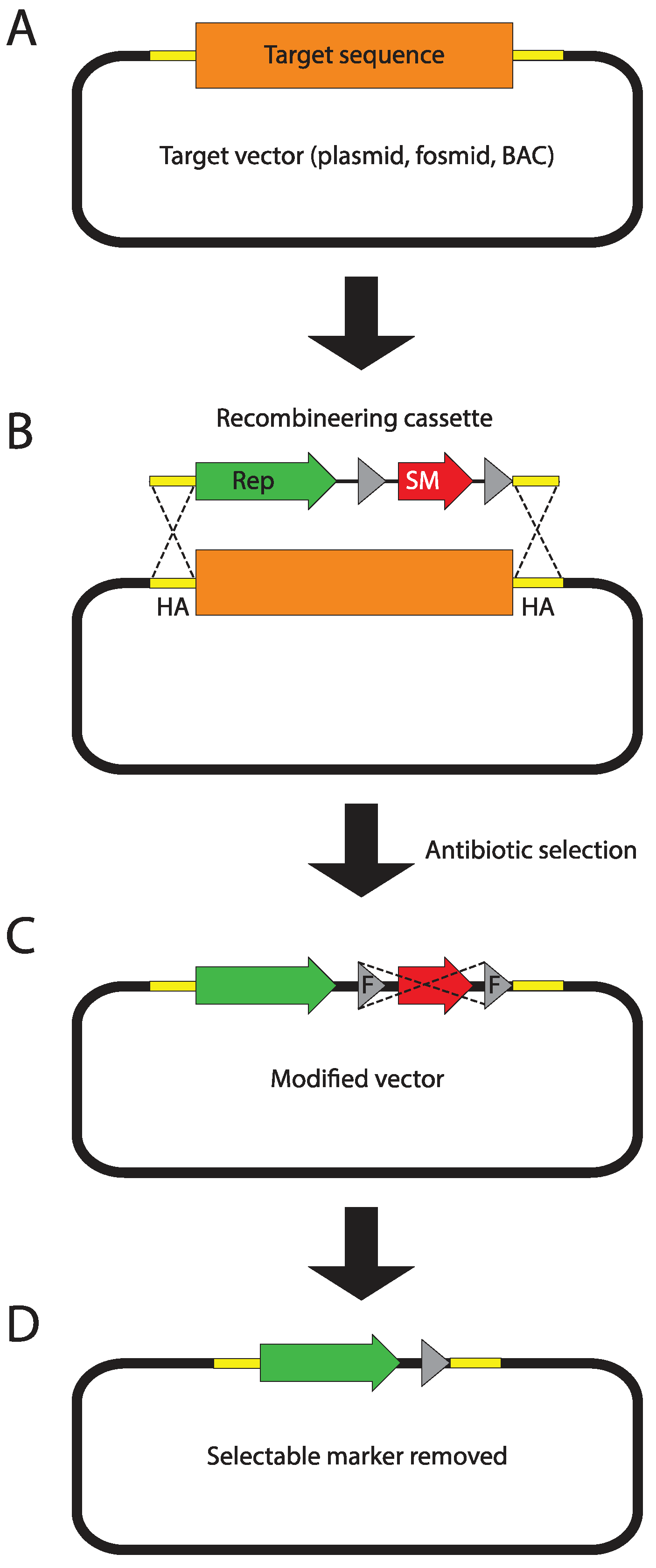
Genome-wide libraries of fruit fly genomic DNA cloned in bacterial artificial chromosomes (BACs) or fosmids, spanning between 20 and over 100 kb, have been constructed for the purpose of genome sequencing [4]. The p[ACMAN] system (Figure 2B,C) has enabled turning them into reliable sources of modifiable genomic inserts, tailored for fly transgenesis. The centerpiece of the system is a single copy vector harboring a second, inducible medium copy origin of replication (oriV), a fly selectable marker (white), and attachment site (attB) for phiC31-mediated transgenesis [124]. Site-specific-recombinase-based transformation enables the insertion of constructs over 100 kb in size. Genomic inserts are subcloned into the backbone using Red/ET homologous recombination (Figure 3), also known as recombineering [173,174,175,176]. The ability to arbitrarily modify and transform large genomic constructs has fostered the development of transformation ready genomic libraries of Drosophila melanogaster and other fly species. Two such resources have been created so far, the p[ACMAN] [177] and FlyFos [178]. The p[ACMAN] features BAC libraries with average insert sizes of 21 and 83 kb. The vector used is similar to the one in the p[ACMAN] subcloning kit. The FlyFos system (Figure 2A,C) features 36 kb fosmid libraries for Drosophila melanogaster and pseudoobscura [169,178]. The library vector also includes an inducible oriV, attB site, and a dominant fluorescent marker, selectable in diverse insect species [179]. The liquid culture recombineering pipeline [180] introduced in the system enables high-throughput gene tagging with a variety of tags in 96-well format [178].
Figure 1.
Mosaic analysis with a repressible cell marker (MARCM). (A) Gal4 transcription factor (blue oval) drives expression of green fluorescent protein (GFP) gene (green box) by binding the upstream activation sequence (UAS) (white box). This expression is repressed when Gal80 (red oval) is present. As a consequence cells that do not carry a gene encoding Gal80 but carry genes encoding Gal4 and UAS-GFP are marked green. (B) In MARCM, the GAL80 repressor gene (red box) is carried on a chromosome that bears the wild-type allele of a gene (yellow box) of interest and a flippase recognition target (FRT) site (grey triangle) placed pericentricaly. The homologous chromosome carries a FRT site in exactly the same position and a mutant allele (orange box), but does not carry the GAL80 gene. Cells also carry the GAL4 gene and UAS-GFP on the other chromosomes. During G2 phase (after DNA replication), flippase mediates recombination between two FRT sites of homologous chromosomes, thus generating sister chromatids; one of which carries the wild-type allele and GAL80 repressor and the other the mutant allele. During mitosis, sister chromatids are distributed to daughter cells, generating cells that are homozygous wild-type or homozygous mutant. Cells that are homozygous mutant are the only cells lacking the GAL80 gene and thus are labeled with GFP. Reproduced with permission from MacMillan: Nature Protocols ©2007 [181].
Figure 1.
Mosaic analysis with a repressible cell marker (MARCM). (A) Gal4 transcription factor (blue oval) drives expression of green fluorescent protein (GFP) gene (green box) by binding the upstream activation sequence (UAS) (white box). This expression is repressed when Gal80 (red oval) is present. As a consequence cells that do not carry a gene encoding Gal80 but carry genes encoding Gal4 and UAS-GFP are marked green. (B) In MARCM, the GAL80 repressor gene (red box) is carried on a chromosome that bears the wild-type allele of a gene (yellow box) of interest and a flippase recognition target (FRT) site (grey triangle) placed pericentricaly. The homologous chromosome carries a FRT site in exactly the same position and a mutant allele (orange box), but does not carry the GAL80 gene. Cells also carry the GAL4 gene and UAS-GFP on the other chromosomes. During G2 phase (after DNA replication), flippase mediates recombination between two FRT sites of homologous chromosomes, thus generating sister chromatids; one of which carries the wild-type allele and GAL80 repressor and the other the mutant allele. During mitosis, sister chromatids are distributed to daughter cells, generating cells that are homozygous wild-type or homozygous mutant. Cells that are homozygous mutant are the only cells lacking the GAL80 gene and thus are labeled with GFP. Reproduced with permission from MacMillan: Nature Protocols ©2007 [181].

Figure 2.
FlyFos and p[ACMAN] genomic libraries. (A) FlyFos library is cloned in a fosmid vector, pFlyFos. Genomic inserts were cloned into the PmlI site. pFlyFos features an inducible origin of replication (oriS for single copy and oriV for arabinose-inducible moderate copy maintenance), attB site for fly transgenesis, and 3xP3-dsRed as a fly-selectable marker. (B) p[ACMAN] libraries are cloned into the BamHI site of a p[ACMAN] bacterial artificial chromosome (BAC) vector. This vector also features inducible oriS/oriV and attB site, but uses white as fly selectable marker. In addition to phiC31-mediated transgenesis, p[ACMAN] vector carrying small inserts can theoretically be used for P-element transformation. (C) Size distribution of FlyFos and p[ACMAN] D. melanogaster libraries. There are two p[ACMAN] libraries: CHORI-321 with average clone size of 83.3 kb and CHORI-322 with average clone size of 21 kb. The FlyFos library has an average clone size of 36 kb.
Figure 2.
FlyFos and p[ACMAN] genomic libraries. (A) FlyFos library is cloned in a fosmid vector, pFlyFos. Genomic inserts were cloned into the PmlI site. pFlyFos features an inducible origin of replication (oriS for single copy and oriV for arabinose-inducible moderate copy maintenance), attB site for fly transgenesis, and 3xP3-dsRed as a fly-selectable marker. (B) p[ACMAN] libraries are cloned into the BamHI site of a p[ACMAN] bacterial artificial chromosome (BAC) vector. This vector also features inducible oriS/oriV and attB site, but uses white as fly selectable marker. In addition to phiC31-mediated transgenesis, p[ACMAN] vector carrying small inserts can theoretically be used for P-element transformation. (C) Size distribution of FlyFos and p[ACMAN] D. melanogaster libraries. There are two p[ACMAN] libraries: CHORI-321 with average clone size of 83.3 kb and CHORI-322 with average clone size of 21 kb. The FlyFos library has an average clone size of 36 kb.

Figure 3.
Recombineering principles. (A) The target sequence (orange box) is carried on a single copy fosmid or bacterial artificial chromosome (BAC) vector. The 50 bp fragments flanking the target sequence, called homology arms (HA), are depicted as yellow boxes. In this example, the target sequence will be replaced with the recombineering cassette. However, when homology arms are designed to directly follow each other, the cassette can be simply inserted into the target vector. (B) The PCR-amplified recombineering cassette harboring homology arms (introduced as primer overhangs) on its termini is electroporated into bacteria carrying the target vector. In the depicted example, the cassette contains a reporter (green arrow) and a flippase recognition target (FRT)-flanked (grey triangles) bacterial selectable marker (red arrow). Homologous recombinase, transiently expressed in bacteria mediates recombination between homology arms replacing the target sequence with the recombineering cassette. (C) Recombinant bacterial cells are selected using the selectable marker encoded in the recombineering cassette. If the selectable marker is flanked by FRT sites, it can now be removed (flipped-out) through transient expression of flippase. (D) The final recombineering product contains the desired sequence and a 34 bp FRT scar flanked by the homology arms.
Figure 3.
Recombineering principles. (A) The target sequence (orange box) is carried on a single copy fosmid or bacterial artificial chromosome (BAC) vector. The 50 bp fragments flanking the target sequence, called homology arms (HA), are depicted as yellow boxes. In this example, the target sequence will be replaced with the recombineering cassette. However, when homology arms are designed to directly follow each other, the cassette can be simply inserted into the target vector. (B) The PCR-amplified recombineering cassette harboring homology arms (introduced as primer overhangs) on its termini is electroporated into bacteria carrying the target vector. In the depicted example, the cassette contains a reporter (green arrow) and a flippase recognition target (FRT)-flanked (grey triangles) bacterial selectable marker (red arrow). Homologous recombinase, transiently expressed in bacteria mediates recombination between homology arms replacing the target sequence with the recombineering cassette. (C) Recombinant bacterial cells are selected using the selectable marker encoded in the recombineering cassette. If the selectable marker is flanked by FRT sites, it can now be removed (flipped-out) through transient expression of flippase. (D) The final recombineering product contains the desired sequence and a 34 bp FRT scar flanked by the homology arms.

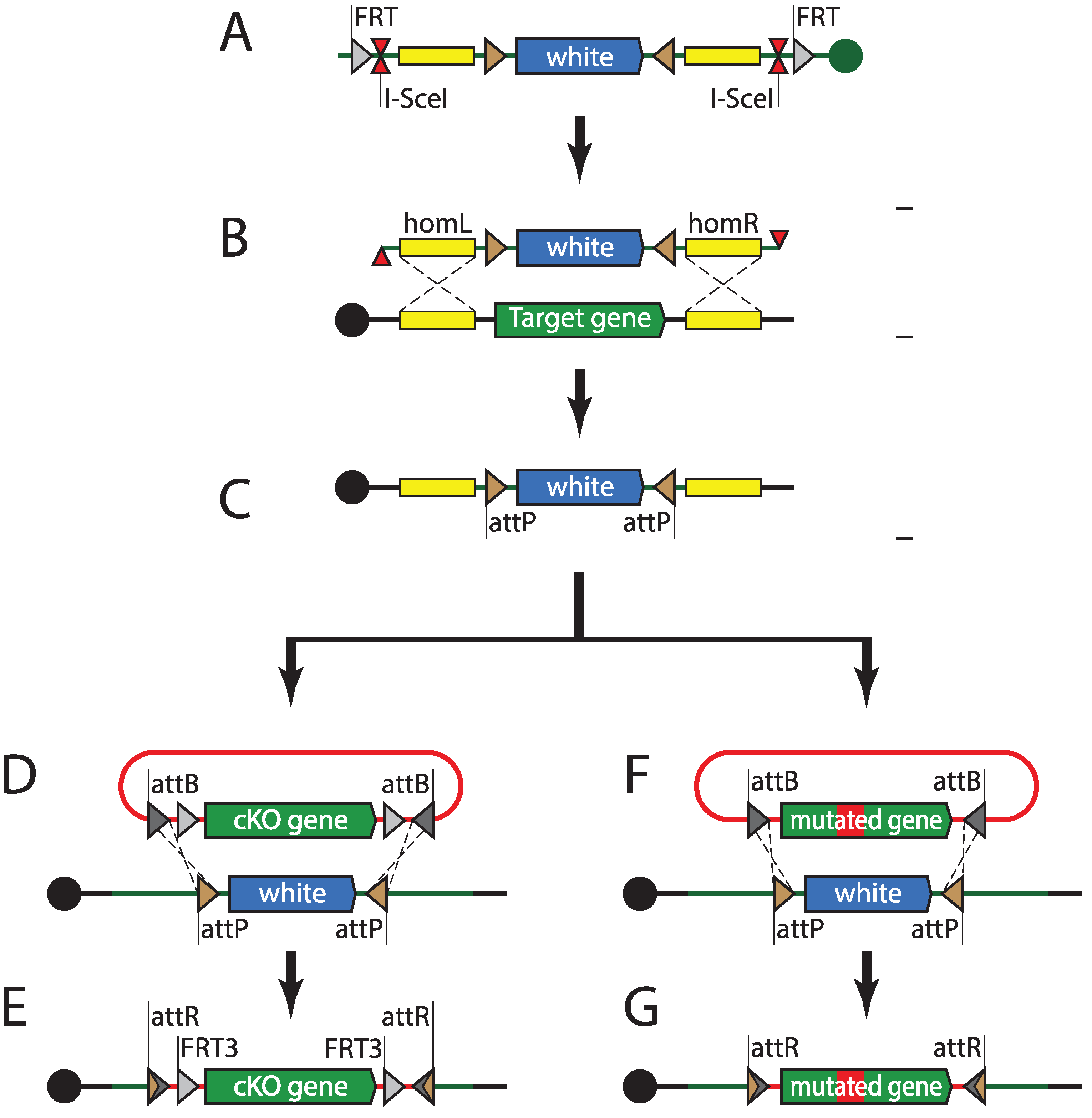
BAC and fosmid-based recombineering has enabled the introduction of modified “third alleles” of genes of interest. The powerful fruit fly genetics toolkit also allows for modifications of genes in situ, in their native loci. The first in situ genomic targeting in Drosophila was performed using the ends-in technique [182]. Ends-in genomic targeting relies on double strand break (DSB) repair through homologous recombination. The targeting construct contains homology arms, one of which is antiparallel to the genomic sequence, and leads to duplication of the targeted locus upon recombination. Initial targeting attempts involving linear DNA injection into the germline were unsuccessful. Inserting the targeting construct into a random locus first, via P-element transgenesis, has solved the issue. FRT sites present on the flanks of the construct were used to mobilize the targeting construct from the genome before generating DSB using I-SceI nuclease [182]. Ends-out targeting uses very similar basic logic, but relies on homology arms that are both parallel to the genomic locus, therefore leading to a clean insertion or replacement [183]. Both ends-in and ends-out have provided reliable means to target genomic loci; however, at a cost of relatively low efficiency. This has made targeting the same locus with different cassettes a labor-intensive task. The integrase-mediated approach for gene knock-out (IMAGO) technique (Figure 4) combines ends-out targeting with phiC31-mediated recombinase-mediated cassette exchange (RMCE) [184]. IMAGO uses ends-out to replace the targeted locus with an attP-flanked selectable marker, which can subsequently be replaced with any desirable construct, thus enabling in situ gene tagging, conditional knock-outs, or functional analysis of orthologs. An alternative strategy uses a single attP site and a loxP-flanked selectable marker as the knock-out cassette [185]. Rescue constructs can then be integrated into the target locus using phiC31-mediated transgenesis, just like into any other landing site.
Genomic targeting techniques using DSBs induced in the targeting construct have proven to be robust tools. However, these approaches have a quite high price tag, because of their low efficiency. Homologous recombination with genomic loci is known to be much more effective if DSBs are introduced in the chromosome [186]. Induction of chromosomal DSBs in specific genomic loci requires designer nucleases that can target a sequence of choice. Currently three custom nuclease systems are in broad use: zinc finger nucleases (ZFNs) [187], transcription activator-like effector nucleases (TALENs), and the bacterial clustered regularly interspaced short palindromic repeat (CRISPR) system and its RNA-driven Cas9 nuclease. Double strand breaks are repaired using one of two cellular mechanisms: non-homologous end joining (NHEJ) and homologous recombination (HR). NHEJ involves processing and ligation of broken strands and usually leads to insertions and deletions [188]. However; it has also been shown to mediate efficient knock-ins in zebrafish [189]. HR requires a sequence homologous to the locus in which DSB has occurred, either from a sister chromatid, paralogous locus; or provided linear or plasmid DNA, and can, therefore, be exploited to insert or replace a genomic sequence with custom constructs [190].
Figure 4.
Integrase-mediated approach for gene knock-out (IMAGO). (A) A targeting construct harboring white gene flanked by attP sites, 1 kb–5 kb homology arms, I-SceI meganuclease site, and flippase recognition target (FRT) sites is inserted into the fly genome using transposition or site-specific integration. (B) The targeting cassette is mobilized into the circular episome by flippase and subsequently linearized by meganuclease. This linear fragment induces cellular double strand break repair mechanisms and (with certain frequency) replaces the genomic locus flanked by homology arms. (C) Recombinant progeny are selected for white dominant marker. The attP sites flanking the white gene can be used for recombinase-mediated cassette exchange. (D, F) A plasmid containing the attB-flanked construct (cKO or a mutant allele) is injected into phiC31-expressing fly embryos and exchanges the attP-flanked white gene. (E, G) Recombinant progeny carrying modified alleles are selected for loss of the white dominant marker.
Figure 4.
Integrase-mediated approach for gene knock-out (IMAGO). (A) A targeting construct harboring white gene flanked by attP sites, 1 kb–5 kb homology arms, I-SceI meganuclease site, and flippase recognition target (FRT) sites is inserted into the fly genome using transposition or site-specific integration. (B) The targeting cassette is mobilized into the circular episome by flippase and subsequently linearized by meganuclease. This linear fragment induces cellular double strand break repair mechanisms and (with certain frequency) replaces the genomic locus flanked by homology arms. (C) Recombinant progeny are selected for white dominant marker. The attP sites flanking the white gene can be used for recombinase-mediated cassette exchange. (D, F) A plasmid containing the attB-flanked construct (cKO or a mutant allele) is injected into phiC31-expressing fly embryos and exchanges the attP-flanked white gene. (E, G) Recombinant progeny carrying modified alleles are selected for loss of the white dominant marker.

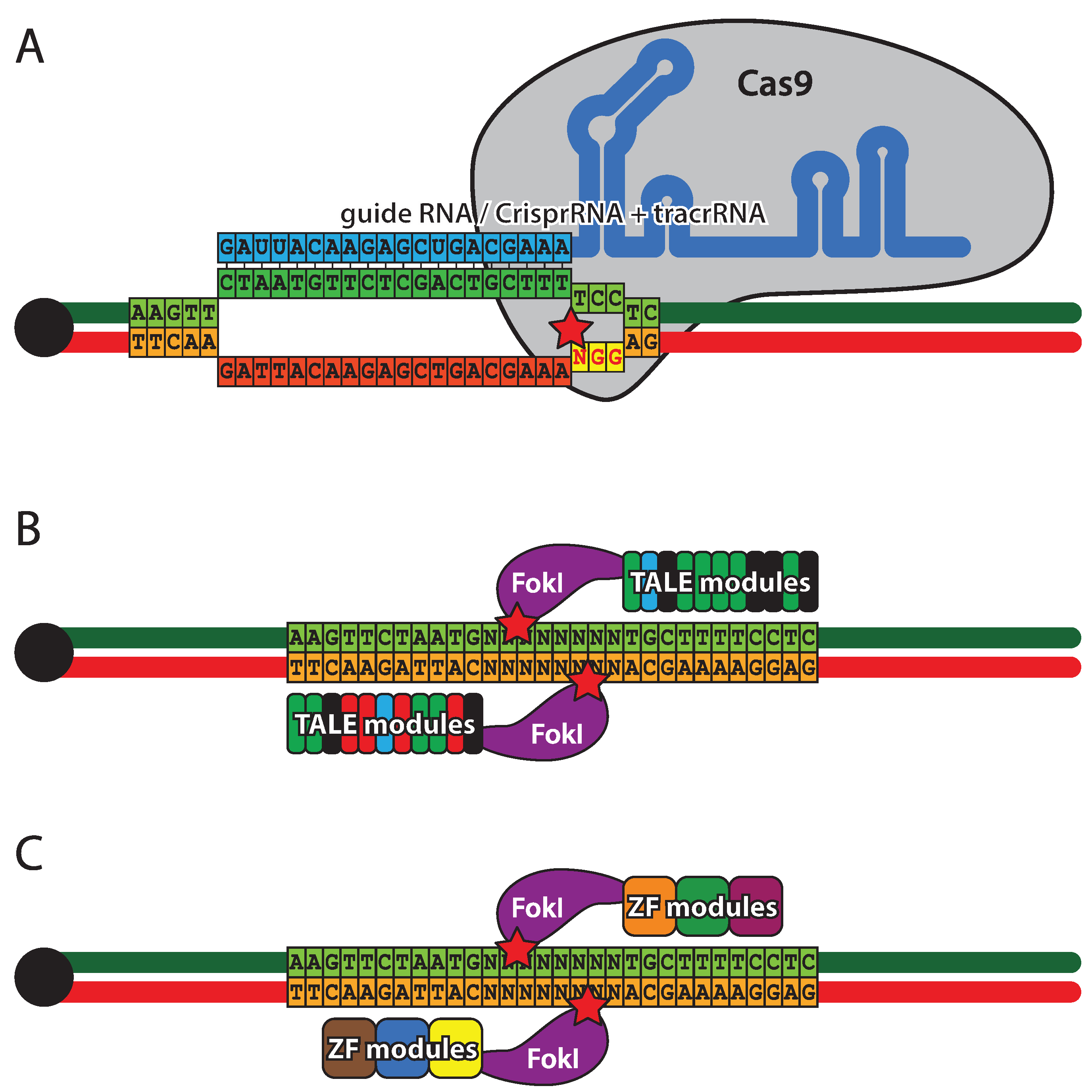
ZFNs were the first designer nuclease system (Figure 5A) to be introduced in flies [188]. They are comprised of three to four zinc-finger DNA binding modules, each recognizing three base pairs, and a FokI endonuclease. Since FokI needs to dimerize for activity, a pair of ZFNs is required for DNA cleavage [187]. The specificity and affinity of zinc-finger modules is context dependent, therefore, several strategies have been developed to achieve assembly of optimal DNA binding domains [191,192,193,194]. TALENs (Figure 5B), similar to ZFNs, are hybrids of DNA binding domains derived from transcription factors and FokI endonuclease and, as a consequence, two TALENs are required to form a functional nuclease [195,196,197]. The TALE (transcription activator-like effector) domains contain a tandem array of 15.5–19.5 repeats, each made of 34 residues, two of which provide DNA-binding specificity against a single nucleotide [198]. Due to the highly repetitive coding sequence of the TALE domain, special approaches have been developed for its efficient assembly using type IIs endonucleases [197]. CRISPR (Figure 5C), a defensive nuclease system from Streptococcus pyogenes, takes a completely different approach to DNA cleavage. The specificity is provided by a crRNA pairing with a 20 nt complimentary sequence within the DNA target. The cleavage is performed by Cas9 nuclease that requires trans-activating CRISPR RNA (tracrRNA) in addition to crRNA for activity. The complimentary region of the DNA target must be followed by a 3 bp PAM (protospacer adjacent motif) [199,200]. A pair of crRNA and tracrRNA can be replaced by a single hybrid guide RNA (sgRNA), thus reducing the system to two components [201]. To date, several implementations of the CRISPR system have been created in Drosophila [202,203,204], including transgenic flies with genomically encoded sources of Cas9 [205,206,207] and tracrRNA/sgRNA [208]. The CRISPR system has been combined with classical ends-out targeting and site-specific integrase approaches, resulting in a versatile toolkit for genome engineering [209]. It should be stated that at this stage the efficiency and specificity of all designer-nuclease-based approaches in vivo remains to be fully established, although CRISPR is showing great promise.
8. Conclusions
Drosophila occupies a paramount position among model organisms, largely due to the variety of genetic tools unique to the fruit fly, its short generation time, and ease of transformation. The Drosophila classic Gal4/UAS two-component expression system and its counterparts, LexA and Q, can be combined with one another and with site-specific recombination systems like Flp/FRT, Cre/LoxP or phiC31, yielding novel combinatorial systems for even tighter spatio-temporal gene expression control, clonal analysis, and lineage tracing [210]. The fruit fly genome is easily accessible using a broad range of genome engineering tools, including those based on classic transposition, site-specific recombinases and fosmid/BAC recombineering [211], as well as the emerging field of genome editing using designer nucleases [212]. Availability of an almost complete genomic sequence for over 12 species from genus Drosophila and dozens of various D. melanogaster strain genomes make fruit flies an excellent model for comparative genomics and population genetics. A large number of human disease-related genes that have homologs in the fruit fly [213,214] connected with powerful resources for QTL mapping and GWAS [89,90] make Drosophila an attractive model for studying the genetic basis of human disease.
Rapid development of genome engineering techniques, especially those introducing synthetic approaches using designer DNA binding domains of TALEs [215,216] and the CRISPR system [217], will undeniably affect the Drosophila field in the next years. Completing the genome and transcriptome sequencing effort for additional fly species [218] will aid in further functional annotation of the Drosophila genome and fuel the evolutionary developmental biology field.
Figure 5.
Designer nucleases. (A) Zink finger nucleases (ZFNs) combine a zinc finger DNA binding domain with a FokI nickase. Each zinc finger recognizes a triplet of bases and usually three to six zinc fingers are present in the targeting domain. Cleavage occurs outside the target sequence and requires a pair of ZFNs, each binding one DNA strand. (B) Transcription activator-like effector nucleases (TALEN), similarly to ZFNs have two domains: a DNA binding domain and a FokI nickase domain. The targeting domain is composed of 33–35 amino acid repeats, each binding a single nucleotide. The cleavage mechanism of TALENs is identical to ZFNs. (C) Clustered regularly interspaced short palindromic repeats (CRISPR) is a RNA driven double-stranded DNA endonuclease system. Cleavage specificity is provided by crRNA (cyan) that hybridizes with the target sequence (green). Cleavage is performed by the Cas9 protein that, in addition to crRNA, requires tracrRNA for activity. The cleavage site (star) is located between the target sequence and NGG protospacer adjacent motif, complimentary to the sequence immediately downstream of the target. crRNA and tracrRNA can be fused to form guide RNA of similar activity.
Figure 5.
Designer nucleases. (A) Zink finger nucleases (ZFNs) combine a zinc finger DNA binding domain with a FokI nickase. Each zinc finger recognizes a triplet of bases and usually three to six zinc fingers are present in the targeting domain. Cleavage occurs outside the target sequence and requires a pair of ZFNs, each binding one DNA strand. (B) Transcription activator-like effector nucleases (TALEN), similarly to ZFNs have two domains: a DNA binding domain and a FokI nickase domain. The targeting domain is composed of 33–35 amino acid repeats, each binding a single nucleotide. The cleavage mechanism of TALENs is identical to ZFNs. (C) Clustered regularly interspaced short palindromic repeats (CRISPR) is a RNA driven double-stranded DNA endonuclease system. Cleavage specificity is provided by crRNA (cyan) that hybridizes with the target sequence (green). Cleavage is performed by the Cas9 protein that, in addition to crRNA, requires tracrRNA for activity. The cleavage site (star) is located between the target sequence and NGG protospacer adjacent motif, complimentary to the sequence immediately downstream of the target. crRNA and tracrRNA can be fused to form guide RNA of similar activity.

Acknowledgments
Work in the Hassan lab is supported by VIB and grants form FWO, BELSPO, and KU Leuven. Radoslaw K. Ejsmont was supported by an EMBO ALTF 1056-2011 long-term fellowship and currently by a Marie Curie Actions co-funded Omics@VIB postdoctoral fellowship.
Author Contributions
Wrote the manuscript: Radoslaw K. Ejsmont and Bassem A. Hassan.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Fleischmann, R.D.; Adams, M.D.; White, O.; Clayton, R.A.; Kirkness, E.F.; Kerlavage, A.R.; Bult, C.J.; Tomb, J.F.; Dougherty, B.A.; Merrick, J.M.; et al. Whole-genome random sequencing and assembly of haemophilus influenzae Rd. Science 1995, 269, 496–512. [Google Scholar]
- Clayton, R.A.; White, O.; Ketchum, K.A.; Venter, J.C. The first genome from the third domain of life. Nature 1997, 387, 459–462. [Google Scholar] [CrossRef]
- C. elegans Sequencing Consortium. Genome sequence of the nematode C. elegans: A platform for investigating biology. Science 1998, 282, 2012–2018. [CrossRef]
- Adams, M.D.; Celniker, S.E.; Holt, R.A.; Evans, C.A.; Gocayne, J.D.; Amanatides, P.G.; Scherer, S.E.; Li, P.W.; Hoskins, R.A.; Galle, R.F.; et al. The genome sequence of drosophila melanogaster. Science 2000, 287, 2185–2195. [Google Scholar] [CrossRef]
- Lewis, E.B. A gene complex controlling segmentation in drosophila. Nature 1978, 276, 565–570. [Google Scholar] [CrossRef]
- Nusslein-Volhard, C.; Wieschaus, E. Mutations affecting segment number and polarity in drosophila. Nature 1980, 287, 795–801. [Google Scholar] [CrossRef]
- Gehring, W.J.; Hiromi, Y. Homeotic genes and the homeobox. Annu. Rev. Genet. 1986, 20, 147–173. [Google Scholar] [CrossRef]
- Lawrence, P.A.; Morata, G. Homeobox genes: Their function in drosophila segmentation and pattern formation. Cell 1994, 78, 181–189. [Google Scholar] [CrossRef]
- Krumlauf, R. Hox genes in vertebrate development. Cell 1994, 78, 191–201. [Google Scholar] [CrossRef]
- Quiring, R.; Walldorf, U.; Kloter, U.; Gehring, W.J. Homology of the eyeless gene of drosophila to the small eye gene in mice and aniridia in humans. Science 1994, 265, 785–789. [Google Scholar]
- Ton, C.C.; Hirvonen, H.; Miwa, H.; Weil, M.M.; Monaghan, P.; Jordan, T.; van Heyningen, V.; Hastie, N.D.; Meijers-Heijboer, H.; Drechsler, M.; et al. Positional cloning and characterization of a paired box- and homeobox-containing gene from the aniridia region. Cell 1991, 67, 1059–1074. [Google Scholar] [CrossRef]
- Hill, R.E.; Favor, J.; Hogan, B.L.; Ton, C.C.; Saunders, G.F.; Hanson, I.M.; Prosser, J.; Jordan, T.; Hastie, N.D.; van Heyningen, V. Mouse small eye results from mutations in a paired-like homeobox-containing gene. Nature 1991, 354, 522–525. [Google Scholar] [CrossRef]
- Halder, G.; Callaerts, P.; Flister, S.; Walldorf, U.; Kloter, U.; Gehring, W.J. Eyeless initiates the expression of both sine oculis and eyes absent during drosophila compound eye development. Development 1998, 125, 2181–2191. [Google Scholar]
- Kumar, J.P. The sine oculis homeobox (six) family of transcription factors as regulators of development and disease. Cell. Mol. Life Sci. 2009, 66, 565–583. [Google Scholar] [CrossRef]
- Jarman, A.P.; Grell, E.H.; Ackerman, L.; Jan, L.Y.; Jan, Y.N. Atonal is the proneural gene for drosophila photoreceptors. Nature 1994, 369, 398–400. [Google Scholar]
- Jarman, A.P.; Grau, Y.; Jan, L.Y.; Jan, Y.N. Atonal is a proneural gene that directs chordotonal organ formation in the drosophila peripheral nervous system. Cell 1993, 73, 1307–1321. [Google Scholar] [CrossRef]
- Brown, N.L.; Patel, S.; Brzezinski, J.; Glaser, T. Math5 is required for retinal ganglion cell and optic nerve formation. Development 2001, 128, 2497–2508. [Google Scholar]
- Bermingham, N.A.; Hassan, B.A.; Price, S.D.; Vollrath, M.A.; Ben-Arie, N.; Eatock, R.A.; Bellen, H.J.; Lysakowski, A.; Zoghbi, H.Y. Math1: An essential gene for the generation of inner ear hair cells. Science 1999, 284, 1837–1841. [Google Scholar] [CrossRef]
- Tabata, T.; Eaton, S.; Kornberg, T.B. The drosophila hedgehog gene is expressed specifically in posterior compartment cells and is a target of engrailed regulation. Genes Dev. 1992, 6, 2635–2645. [Google Scholar] [CrossRef]
- Lee, J.J.; von Kessler, D.P.; Parks, S.; Beachy, P.A. Secretion and localized transcription suggest a role in positional signaling for products of the segmentation gene hedgehog. Cell 1992, 71, 33–50. [Google Scholar] [CrossRef]
- Hooper, J.E.; Scott, M.P. The drosophila patched gene encodes a putative membrane protein required for segmental patterning. Cell 1989, 59, 751–765. [Google Scholar] [CrossRef]
- Nakano, Y.; Guerrero, I.; Hidalgo, A.; Taylor, A.; Whittle, J.R.; Ingham, P.W. A protein with several possible membrane-spanning domains encoded by the drosophila segment polarity gene patched. Nature 1989, 341, 508–513. [Google Scholar]
- Stone, D.M.; Hynes, M.; Armanini, M.; Swanson, T.A.; Gu, Q.; Johnson, R.L.; Scott, M.P.; Pennica, D.; Goddard, A.; Phillips, H.; et al. The tumour-suppressor gene patched encodes a candidate receptor for sonic hedgehog. Nature 1996, 384, 129–134. [Google Scholar] [CrossRef]
- Marigo, V.; Davey, R.A.; Zuo, Y.; Cunningham, J.M.; Tabin, C.J. Biochemical evidence that patched is the hedgehog receptor. Nature 1996, 384, 176–179. [Google Scholar] [CrossRef]
- Bhanot, P.; Brink, M.; Samos, C.H.; Hsieh, J.C.; Wang, Y.; Macke, J.P.; Andrew, D.; Nathans, J.; Nusse, R. A new member of the frizzled family from drosophila functions as a wingless receptor. Nature 1996, 382, 225–230. [Google Scholar] [CrossRef]
- Siegfried, E.; Chou, T.B.; Perrimon, N. Wingless signaling acts through zeste-white 3, the drosophila homolog of glycogen synthase kinase-3, to regulate engrailed and establish cell fate. Cell 1992, 71, 1167–1179. [Google Scholar] [CrossRef]
- Wehrli, M.; Dougan, S.T.; Caldwell, K.; O'Keefe, L.; Schwartz, S.; Vaizel-Ohayon, D.; Schejter, E.; Tomlinson, A.; DiNardo, S. Arrow encodes an ldl-receptor-related protein essential for wingless signalling. Nature 2000, 407, 527–530. [Google Scholar] [CrossRef]
- Brunner, E.; Peter, O.; Schweizer, L.; Basler, K. Pangolin encodes a lef-1 homologue that acts downstream of armadillo to transduce the wingless signal in drosophila. Nature 1997, 385, 829–833. [Google Scholar] [CrossRef]
- Adler, P.N. The genetic control of tissue polarity in drosophila. BioEssays 1992, 14, 735–741. [Google Scholar]
- Eaton, S.; Julicher, F. Cell flow and tissue polarity patterns. Curr. Opin. Genet. Dev. 2011, 21, 747–752. [Google Scholar] [CrossRef]
- Wharton, K.A.; Johansen, K.M.; Xu, T.; Artavanis-Tsakonas, S. Nucleotide sequence from the neurogenic locus notch implies a gene product that shares homology with proteins containing egf-like repeats. Cell 1985, 43, 567–581. [Google Scholar]
- Artavanis-Tsakonas, S.; Matsuno, K.; Fortini, M.E. Notch signaling. Science 1995, 268, 225–232. [Google Scholar]
- Artavanis-Tsakonas, S.; Rand, M.D.; Lake, R.J. Notch signaling: Cell fate control and signal integration in development. Science 1999, 284, 770–776. [Google Scholar] [CrossRef]
- Wu, S.; Huang, J.; Dong, J.; Pan, D. Hippo encodes a STE-20 family protein kinase that restricts cell proliferation and promotes apoptosis in conjunction with salvador and warts. Cell 2003, 114, 445–456. [Google Scholar]
- Harvey, K.F.; Pfleger, C.M.; Hariharan, I.K. The drosophila mst ortholog, hippo, restricts growth and cell proliferation and promotes apoptosis. Cell 2003, 114, 457–467. [Google Scholar]
- Udan, R.S.; Kango-Singh, M.; Nolo, R.; Tao, C.; Halder, G. Hippo promotes proliferation arrest and apoptosis in the salvador/warts pathway. Nat. Cell Biol. 2003, 5, 914–920. [Google Scholar] [CrossRef]
- Fortini, M.E.; Bonini, N.M. Modeling human neurodegenerative diseases in drosophila: On a wing and a prayer. Trends Genet. 2000, 16, 161–167. [Google Scholar] [CrossRef]
- Lloyd, T.E.; Taylor, J.P. Flightless flies: Drosophila models of neuromuscular disease. Ann. N. Y. Acad. Sci. 2010, 1184, e1–e20. [Google Scholar]
- Potter, C.J.; Turenchalk, G.S.; Xu, T. Drosophila in cancer research. An expanding role. Trends Genet. 2000, 16, 33–39. [Google Scholar] [CrossRef]
- Jackson, G.R.; Salecker, I.; Dong, X.; Yao, X.; Arnheim, N.; Faber, P.W.; MacDonald, M.E.; Zipursky, S.L. Polyglutamine-expanded human huntingtin transgenes induce degeneration of drosophila photoreceptor neurons. Neuron 1998, 21, 633–642. [Google Scholar] [CrossRef]
- Warrick, J.M.; Paulson, H.L.; Gray-Board, G.L.; Bui, Q.T.; Fischbeck, K.H.; Pittman, R.N.; Bonini, N.M. Expanded polyglutamine protein forms nuclear inclusions and causes neural degeneration in drosophila. Cell 1998, 93, 939–949. [Google Scholar]
- Morales, J.; Hiesinger, P.R.; Schroeder, A.J.; Kume, K.; Verstreken, P.; Jackson, F.R.; Nelson, D.L.; Hassan, B.A. Drosophila fragile x protein, dfxr, regulates neuronal morphology and function in the brain. Neuron 2002, 34, 961–972. [Google Scholar] [CrossRef]
- Luo, L.Q.; Martin-Morris, L.E.; White, K. Identification, secretion, and neural expression of appl, a drosophila protein similar to human amyloid protein precursor. J. Neurosci. 1990, 10, 3849–3861. [Google Scholar]
- Luo, L.; Tully, T.; White, K. Human amyloid precursor protein ameliorates behavioral deficit of flies deleted for appl gene. Neuron 1992, 9, 595–605. [Google Scholar] [CrossRef]
- Ye, Y.; Fortini, M.E. Apoptotic activities of wild-type and alzheimer’s disease-related mutant presenilins in drosophila melanogaster. J. Cell Biol. 1999, 146, 1351–1364. [Google Scholar] [CrossRef]
- Wittmann, C.W.; Wszolek, M.F.; Shulman, J.M.; Salvaterra, P.M.; Lewis, J.; Hutton, M.; Feany, M.B. Tauopathy in drosophila: Neurodegeneration without neurofibrillary tangles. Science 2001, 293, 711–714. [Google Scholar] [CrossRef]
- Feany, M.B.; Bender, W.W. A drosophila model of parkinson’s disease. Nature 2000, 404, 394–398. [Google Scholar] [CrossRef]
- Whitworth, A.J.; Theodore, D.A.; Greene, J.C.; Benes, H.; Wes, P.D.; Pallanck, L.J. Increased glutathione s-transferase activity rescues dopaminergic neuron loss in a drosophila model of parkinson’s disease. Proc. Natl. Acad. Sci. USA 2005, 102, 8024–8029. [Google Scholar]
- Parkes, T.L.; Elia, A.J.; Dickinson, D.; Hilliker, A.J.; Phillips, J.P.; Boulianne, G.L. Extension of drosophila lifespan by overexpression of human sod1 in motorneurons. Nat. Genet. 1998, 19, 171–174. [Google Scholar]
- Shahidullah, M.; Le Marchand, S.J.; Fei, H.; Zhang, J.; Pandey, U.B.; Dalva, M.B.; Pasinelli, P.; Levitan, I.B. Defects in synapse structure and function precede motor neuron degeneration in drosophila models of fus-related als. J. Neurosci. 2013, 33, 19590–19598. [Google Scholar] [CrossRef]
- Van der Plas, M.C.; Pilgram, G.S.; Plomp, J.J.; de Jong, A.; Fradkin, L.G.; Noordermeer, J.N. Dystrophin is required for appropriate retrograde control of neurotransmitter release at the drosophila neuromuscular junction. J. Neurosci. 2006, 26, 333–344. [Google Scholar]
- Moore, M.S.; DeZazzo, J.; Luk, A.Y.; Tully, T.; Singh, C.M.; Heberlein, U. Ethanol intoxication in drosophila: Genetic and pharmacological evidence for regulation by the camp signaling pathway. Cell 1998, 93, 997–1007. [Google Scholar] [CrossRef]
- Kaun, K.R.; Azanchi, R.; Maung, Z.; Hirsh, J.; Heberlein, U. A drosophila model for alcohol reward. Nat. Neurosci. 2011, 14, 612–619. [Google Scholar]
- McClung, C.; Hirsh, J. Stereotypic behavioral responses to free-base cocaine and the development of behavioral sensitization in drosophila. Curr. Biol. 1998, 8, 109–112. [Google Scholar] [CrossRef]
- Al-Anzi, B.; Sapin, V.; Waters, C.; Zinn, K.; Wyman, R.J.; Benzer, S. Obesity-blocking neurons in drosophila. Neuron 2009, 63, 329–341. [Google Scholar] [CrossRef]
- Baker, K.D.; Thummel, C.S. Diabetic larvae and obese flies-emerging studies of metabolism in drosophila. Cell Metab. 2007, 6, 257–266. [Google Scholar] [CrossRef]
- Wolf, M.J.; Amrein, H.; Izatt, J.A.; Choma, M.A.; Reedy, M.C.; Rockman, H.A. Drosophila as a model for the identification of genes causing adult human heart disease. Proc. Natl. Acad. Sci. USA 2006, 103, 1394–1399. [Google Scholar] [CrossRef]
- Roeder, T.; Isermann, K.; Kallsen, K.; Uliczka, K.; Wagner, C. A drosophila asthma model—What the fly tells us about inflammatory diseases of the lung. Adv. Exp. Med. Biol. 2012, 710, 37–47. [Google Scholar] [CrossRef]
- Xu, T.; Wang, W.; Zhang, S.; Stewart, R.A.; Yu, W. Identifying tumor suppressors in genetic mosaics: The drosophila lats gene encodes a putative protein kinase. Development 1995, 121, 1053–1063. [Google Scholar]
- Pagliarini, R.A.; Xu, T. A genetic screen in drosophila for metastatic behavior. Science 2003, 302, 1227–1231. [Google Scholar] [CrossRef]
- Januschke, J.; Gonzalez, C. Drosophila asymmetric division, polarity and cancer. Oncogene 2008, 27, 6994–7002. [Google Scholar] [CrossRef]
- Brumby, A.M.; Richardson, H.E. Using drosophila melanogaster to map human cancer pathways. Nat. Rev. Cancer 2005, 5, 626–639. [Google Scholar] [CrossRef]
- Vidal, M.; Cagan, R.L. Drosophila models for cancer research. Curr. Opin. Genet. Dev. 2006, 16, 10–16. [Google Scholar] [CrossRef]
- Miles, W.O.; Dyson, N.J.; Walker, J.A. Modeling tumor invasion and metastasis in drosophila. Dis. Models Mech. 2011, 4, 753–761. [Google Scholar] [CrossRef]
- Gonzalez, C. Drosophila melanogaster: A model and a tool to investigate malignancy and identify new therapeutics. Nat. Rev. Cancer 2013, 13, 172–183. [Google Scholar] [CrossRef]
- Bosco, G.; Campbell, P.; Leiva-Neto, J.T.; Markow, T.A. Analysis of drosophila species genome size and satellite DNA content reveals significant differences among strains as well as between species. Genetics 2007, 177, 1277–1290. [Google Scholar] [CrossRef]
- Hoskins, R.A.; Smith, C.D.; Carlson, J.W.; Carvalho, A.B.; Halpern, A.; Kaminker, J.S.; Kennedy, C.; Mungall, C.J.; Sullivan, B.A.; Sutton, G.G.; et al. Heterochromatic sequences in a drosophila whole-genome shotgun assembly. Genome Biol. 2002. [Google Scholar] [CrossRef] [Green Version]
- Misra, S.; Crosby, M.A.; Mungall, C.J.; Matthews, B.B.; Campbell, K.S.; Hradecky, P.; Huang, Y.; Kaminker, J.S.; Millburn, G.H.; Prochnik, S.E.; et al. Annotation of the drosophila melanogaster euchromatic genome: A systematic review. Genome Biol. 2002. [Google Scholar] [CrossRef] [Green Version]
- Stapleton, M.; Carlson, J.; Brokstein, P.; Yu, C.; Champe, M.; George, R.; Guarin, H.; Kronmiller, B.; Pacleb, J.; Park, S.; et al. A drosophila full-length cdna resource. Genome Biol. 2002. [Google Scholar] [CrossRef]
- Tomancak, P.; Beaton, A.; Weiszmann, R.; Kwan, E.; Shu, S.; Lewis, S.E.; Richards, S.; Ashburner, M.; Hartenstein, V.; Celniker, S.E.; et al. Systematic determination of patterns of gene expression during drosophila embryogenesis. Genome Biol. 2002. [Google Scholar] [CrossRef] [Green Version]
- Kaminker, J.S.; Bergman, C.M.; Kronmiller, B.; Carlson, J.; Svirskas, R.; Patel, S.; Frise, E.; Wheeler, D.A.; Lewis, S.E.; Rubin, G.M.; et al. The transposable elements of the drosophila melanogaster euchromatin: A genomics perspective. Genome Biol. 2002. [Google Scholar] [CrossRef] [Green Version]
- Ohler, U.; Liao, G.C.; Niemann, H.; Rubin, G.M. Computational analysis of core promoters in the drosophila genome. Genome Biol. 2002. [Google Scholar] [CrossRef]
- Marygold, S.J.; Leyland, P.C.; Seal, R.L.; Goodman, J.L.; Thurmond, J.; Strelets, V.B.; Wilson, R.J.; the FlyBase consortium. Flybase: Improvements to the bibliography. Nucleic Acids Res. 2013, 41, D751–D757. [Google Scholar] [CrossRef]
- Daines, B.; Wang, H.; Wang, L.; Li, Y.; Han, Y.; Emmert, D.; Gelbart, W.; Wang, X.; Li, W.; Gibbs, R.; et al. The drosophila melanogaster transcriptome by paired-end RNA sequencing. Genome Res. 2011, 21, 315–324. [Google Scholar] [CrossRef]
- Mungall, C.J.; Misra, S.; Berman, B.P.; Carlson, J.; Frise, E.; Harris, N.; Marshall, B.; Shu, S.; Kaminker, J.S.; Prochnik, S.E.; et al. An integrated computational pipeline and database to support whole-genome sequence annotation. Genome Biol. 2002. [Google Scholar] [CrossRef]
- Lewis, S.E.; Searle, S.M.; Harris, N.; Gibson, M.; Lyer, V.; Richter, J.; Wiel, C.; Bayraktaroglir, L.; Birney, E.; Crosby, M.A.; et al. Apollo: A sequence annotation editor. Genome Biol. 2002. [Google Scholar] [CrossRef]
- Reese, M.G.; Kulp, D.; Tammana, H.; Haussler, D. Genie—Gene finding in drosophila melanogaster. Genome Res. 2000, 10, 529–538. [Google Scholar] [CrossRef]
- Wood, V.; Gwilliam, R.; Rajandream, M.A.; Lyne, M.; Lyne, R.; Stewart, A.; Sgouros, J.; Peat, N.; Hayles, J.; Baker, S.; et al. The genome sequence of schizosaccharomyces pombe. Nature 2002, 415, 871–880. [Google Scholar] [CrossRef] [Green Version]
- Stein, L.D.; Bao, Z.; Blasiar, D.; Blumenthal, T.; Brent, M.R.; Chen, N.; Chinwalla, A.; Clarke, L.; Clee, C.; Coghlan, A.; et al. The genome sequence of caenorhabditis briggsae: A platform for comparative genomics. PLoS Biol. 2003, 1, e45. [Google Scholar]
- Richards, S.; Liu, Y.; Bettencourt, B.R.; Hradecky, P.; Letovsky, S.; Nielsen, R.; Thornton, K.; Hubisz, M.J.; Chen, R.; Meisel, R.P.; et al. Comparative genome sequencing of drosophila pseudoobscura: Chromosomal, gene, and cis-element evolution. Genome Res. 2005, 15, 1–18. [Google Scholar] [CrossRef]
- Drosophila 12 Genomes Consortium; Clark, A.G.; Eisen, M.B.; Smith, D.R.; Bergman, C.M.; Oliver, B.; Markow, T.A.; Kaufman, T.C.; Kellis, M.; Gelbart, W.; et al. Evolution of genes and genomes on the drosophila phylogeny. Nature 2007, 450, 203–218. [Google Scholar] [CrossRef] [Green Version]
- Lin, M.F.; Carlson, J.W.; Crosby, M.A.; Matthews, B.B.; Yu, C.; Park, S.; Wan, K.H.; Schroeder, A.J.; Gramates, L.S.; St Pierre, S.E.; et al. Revisiting the protein-coding gene catalog of drosophila melanogaster using 12 fly genomes. Genome Res. 2007, 17, 1823–1836. [Google Scholar] [CrossRef]
- Stark, A.; Lin, M.F.; Kheradpour, P.; Pedersen, J.S.; Parts, L.; Carlson, J.W.; Crosby, M.A.; Rasmussen, M.D.; Roy, S.; Deoras, A.N.; et al. Discovery of functional elements in 12 drosophila genomes using evolutionary signatures. Nature 2007, 450, 219–232. [Google Scholar] [CrossRef]
- Majoros, W.H.; Ohler, U. Modeling the evolution of regulatory elements by simultaneous detection and alignment with phylogenetic pair hmms. PLoS Comput. Biol. 2010, 6, e1001037. [Google Scholar] [CrossRef]
- Arunachalam, M.; Jayasurya, K.; Tomancak, P.; Ohler, U. An alignment-free method to identify candidate orthologous enhancers in multiple drosophila genomes. Bioinformatics 2010, 26, 2109–2115. [Google Scholar] [CrossRef]
- Aerts, S.; Quan, X.J.; Claeys, A.; Naval Sanchez, M.; Tate, P.; Yan, J.; Hassan, B.A. Robust target gene discovery through transcriptome perturbations and genome-wide enhancer predictions in drosophila uncovers a regulatory basis for sensory specification. PLoS Biol. 2010, 8, e1000435. [Google Scholar] [CrossRef]
- Mackay, T.F. Quantitative trait loci in drosophila. Nat. Rev. Genet. 2001, 2, 11–20. [Google Scholar] [CrossRef]
- Hindorff, L.A.; Sethupathy, P.; Junkins, H.A.; Ramos, E.M.; Mehta, J.P.; Collins, F.S.; Manolio, T.A. Potential etiologic and functional implications of genome-wide association loci for human diseases and traits. Proc. Natl. Acad. Sci. USA 2009, 106, 9362–9367. [Google Scholar] [CrossRef]
- Mackay, T.F.; Richards, S.; Stone, E.A.; Barbadilla, A.; Ayroles, J.F.; Zhu, D.; Casillas, S.; Han, Y.; Magwire, M.M.; Cridland, J.M.; et al. The drosophila melanogaster genetic reference panel. Nature 2012, 482, 173–178. [Google Scholar]
- King, E.G.; Merkes, C.M.; McNeil, C.L.; Hoofer, S.R.; Sen, S.; Broman, K.W.; Long, A.D.; Macdonald, S.J. Genetic dissection of a model complex trait using the drosophila synthetic population resource. Genome Res. 2012, 22, 1558–1566. [Google Scholar] [CrossRef]
- Ayroles, J.F.; Carbone, M.A.; Stone, E.A.; Jordan, K.W.; Lyman, R.F.; Magwire, M.M.; Rollmann, S.M.; Duncan, L.H.; Lawrence, F.; Anholt, R.R.; et al. Systems genetics of complex traits in drosophila melanogaster. Nat. Genet. 2009, 41, 299–307. [Google Scholar] [CrossRef]
- Massouras, A.; Waszak, S.M.; Albarca-Aguilera, M.; Hens, K.; Holcombe, W.; Ayroles, J.F.; Dermitzakis, E.T.; Stone, E.A.; Jensen, J.D.; Mackay, T.F.; et al. Genomic variation and its impact on gene expression in drosophila melanogaster. PLoS Genet. 2012, 8, e1003055. [Google Scholar] [CrossRef]
- Jordan, K.W.; Craver, K.L.; Magwire, M.M.; Cubilla, C.E.; Mackay, T.F.; Anholt, R.R. Genome-wide association for sensitivity to chronic oxidative stress in drosophila melanogaster. PLoS One 2012, 7, e38722. [Google Scholar]
- Jumbo-Lucioni, P.; Bu, S.; Harbison, S.T.; Slaughter, J.C.; Mackay, T.F.; Moellering, D.R.; de Luca, M. Nuclear genomic control of naturally occurring variation in mitochondrial function in drosophila melanogaster. BMC Genomics 2002. [Google Scholar] [CrossRef]
- Magwire, M.M.; Fabian, D.K.; Schweyen, H.; Cao, C.; Longdon, B.; Bayer, F.; Jiggins, F.M. Genome-wide association studies reveal a simple genetic basis of resistance to naturally coevolving viruses in drosophila melanogaster. PLoS Genet. 2012, 8, e1003057. [Google Scholar] [CrossRef]
- Harbison, S.T.; McCoy, L.J.; Mackay, T.F. Genome-wide association study of sleep in drosophila melanogaster. BMC Genomics 2013. [Google Scholar] [CrossRef]
- King, E.G.; Macdonald, S.J.; Long, A.D. Properties and power of the drosophila synthetic population resource for the routine dissection of complex traits. Genetics 2012, 191, 935–949. [Google Scholar] [CrossRef]
- ENCODE Project Consortium. The encode (encyclopedia of DNA elements) project. Science 2004, 306, 636–640. [Google Scholar] [CrossRef]
- Gerstein, M.B.; Lu, Z.J.; van Nostrand, E.L.; Cheng, C.; Arshinoff, B.I.; Liu, T.; Yip, K.Y.; Robilotto, R.; Rechtsteiner, A.; Ikegami, K.; et al. Integrative analysis of the caenorhabditis elegans genome by the modencode project. Science 2010, 330, 1775–1787. [Google Scholar] [CrossRef]
- modENCODE Consortium; Roy, S.; Ernst, J.; Kharchenko, P.V.; Kheradpour, P.; Negre, N.; Eaton, M.L.; Landolin, J.M.; Bristow, C.A.; Ma, L.; et al. Identification of functional elements and regulatory circuits by drosophila modencode. Science 2010, 330, 1787–1797. [Google Scholar] [CrossRef]
- Riddle, N.C.; Minoda, A.; Kharchenko, P.V.; Alekseyenko, A.A.; Schwartz, Y.B.; Tolstorukov, M.Y.; Gorchakov, A.A.; Jaffe, J.D.; Kennedy, C.; Linder-Basso, D.; et al. Plasticity in patterns of histone modifications and chromosomal proteins in drosophila heterochromatin. Genome Res. 2011, 21, 147–163. [Google Scholar] [CrossRef]
- Kharchenko, P.V.; Alekseyenko, A.A.; Schwartz, Y.B.; Minoda, A.; Riddle, N.C.; Ernst, J.; Sabo, P.J.; Larschan, E.; Gorchakov, A.A.; Gu, T.; et al. Comprehensive analysis of the chromatin landscape in drosophila melanogaster. Nature 2011, 471, 480–485. [Google Scholar] [CrossRef]
- Nègre, N.; Brown, C.D.; Ma, L.; Bristow, C.A.; Miller, S.W.; Wagner, U.; Kheradpour, P.; Eaton, M.L.; Loriaux, P.; Sealfon, R.; et al. A cis-regulatory map of the drosophila genome. Nature 2011, 471, 527–531. [Google Scholar] [CrossRef] [Green Version]
- Spradling, A.C.; Rubin, G.M. Transposition of cloned p elements into drosophila germ line chromosomes. Science 1982, 218, 341–347. [Google Scholar]
- Searles, L.L.; Jokerst, R.S.; Bingham, P.M.; Voelker, R.A.; Greenleaf, A.L. Molecular cloning of sequences from a drosophila RNA polymerase ii locus by p element transposon tagging. Cell 1982, 31, 585–592. [Google Scholar] [CrossRef]
- Rubin, G.M.; Spradling, A.C. Genetic transformation of drosophila with transposable element vectors. Science 1982, 218, 348–353. [Google Scholar]
- Cooley, L.; Kelley, R.; Spradling, A. Insertional mutagenesis of the drosophila genome with single P elements. Science 1988, 239, 1121–1128. [Google Scholar]
- Cooley, L.; Thompson, D.; Spradling, A.C. Constructing deletions with defined endpoints in drosophila. Proc. Natl. Acad. Sci. USA 1990, 87, 3170–3173. [Google Scholar] [CrossRef]
- O'Kane, C.J.; Gehring, W.J. Detection in situ of genomic regulatory elements in drosophila. Proc. Natl. Acad. Sci. USA 1987, 84, 9123–9127. [Google Scholar] [CrossRef]
- Bellen, H.J.; Levis, R.W.; Liao, G.; He, Y.; Carlson, J.W.; Tsang, G.; Evans-Holm, M.; Hiesinger, P.R.; Schulze, K.L.; Rubin, G.M.; et al. The bdgp gene disruption project: Single transposon insertions associated with 40% of drosophila genes. Genetics 2004, 167, 761–781. [Google Scholar] [CrossRef]
- Bellen, H.J.; Levis, R.W.; He, Y.; Carlson, J.W.; Evans-Holm, M.; Bae, E.; Kim, J.; Metaxakis, A.; Savakis, C.; Schulze, K.L.; et al. The drosophila gene disruption project: Progress using transposons with distinctive site specificities. Genetics 2011, 188, 731–743. [Google Scholar] [CrossRef]
- Jacobson, J.W.; Medhora, M.M.; Hartl, D.L. Molecular structure of a somatically unstable transposable element in drosophila. Proc. Natl. Acad. Sci. USA 1986, 83, 8684–8688. [Google Scholar] [CrossRef]
- Garza, D.; Medhora, M.; Koga, A.; Hartl, D.L. Introduction of the transposable element mariner into the germline of drosophila melanogaster. Genetics 1991, 128, 303–310. [Google Scholar]
- Franz, G.; Savakis, C. Minos, a new transposable element from drosophila hydei, is a member of the tc1-like family of transposons. Nucleic Acids Res. 1991, 19, 6646. [Google Scholar] [CrossRef]
- Loukeris, T.G.; Arca, B.; Livadaras, I.; Dialektaki, G.; Savakis, C. Introduction of the transposable element minos into the germ line of drosophila melanogaster. Proc. Natl. Acad. Sci. USA 1995, 92, 9485–9489. [Google Scholar] [CrossRef]
- Handler, A.M.; McCombs, S.D.; Fraser, M.J.; Saul, S.H. The lepidopteran transposon vector, piggybac, mediates germ-line transformation in the mediterranean fruit fly. Proc. Natl. Acad. Sci. USA 1998, 95, 7520–7525. [Google Scholar] [CrossRef]
- Handler, A.M.; Harrell, R.A., 2nd. Germline transformation of drosophila melanogaster with the piggybac transposon vector. Insect Mol. Biol. 1999, 8, 449–457. [Google Scholar] [CrossRef]
- Spradling, A.C.; Rubin, G.M. The effect of chromosomal position on the expression of the drosophila xanthine dehydrogenase gene. Cell 1983, 34, 47–57. [Google Scholar] [CrossRef]
- Levis, R.; Hazelrigg, T.; Rubin, G.M. Effects of genomic position on the expression of transduced copies of the white gene of drosophila. Science 1985, 229, 558–561. [Google Scholar]
- Taillebourg, E.; Dura, J.M. A novel mechanism for p element homing in drosophila. Proc. Natl. Acad. Sci. USA 1999, 96, 6856–6861. [Google Scholar] [CrossRef]
- Cheng, Y.; Kwon, D.Y.; Arai, A.L.; Mucci, D.; Kassis, J.A. P-element homing is facilitated by engrailed polycomb-group response elements in drosophila melanogaster. PLoS One 2012, 7, e30437. [Google Scholar]
- Thorpe, H.M.; Smith, M.C. In vitro site-specific integration of bacteriophage DNA catalyzed by a recombinase of the resolvase/invertase family. Proc. Natl. Acad. Sci. USA 1998, 95, 5505–5510. [Google Scholar] [CrossRef]
- Groth, A.C.; Fish, M.; Nusse, R.; Calos, M.P. Construction of transgenic drosophila by using the site-specific integrase from phage phic31. Genetics 2004, 166, 1775–1782. [Google Scholar]
- Venken, K.J.; He, Y.; Hoskins, R.A.; Bellen, H.J. P[acman]: A bac transgenic platform for targeted insertion of large DNA fragments in d. Melanogaster. Science 2006, 314, 1747–1751. [Google Scholar] [CrossRef]
- Bischof, J.; Maeda, R.K.; Hediger, M.; Karch, F.; Basler, K. An optimized transgenesis system for drosophila using germ-line-specific phic31 integrases. Proc. Natl. Acad. Sci. USA 2007, 104, 3312–3317. [Google Scholar] [CrossRef]
- Markstein, M.; Pitsouli, C.; Villalta, C.; Celniker, S.E.; Perrimon, N. Exploiting position effects and the gypsy retrovirus insulator to engineer precisely expressed transgenes. Nat. Genet. 2008, 40, 476–483. [Google Scholar] [CrossRef]
- Brand, A.H.; Perrimon, N. Targeted gene expression as a means of altering cell fates and generating dominant phenotypes. Development 1993, 118, 401–415. [Google Scholar]
- Manseau, L.; Baradaran, A.; Brower, D.; Budhu, A.; Elefant, F.; Phan, H.; Philp, A.V.; Yang, M.; Glover, D.; Kaiser, K.; et al. Gal4 enhancer traps expressed in the embryo, larval brain, imaginal discs, and ovary of drosophila. Dev. Dyn. 1997, 209, 310–322. [Google Scholar] [CrossRef]
- Hayashi, S.; Ito, K.; Sado, Y.; Taniguchi, M.; Akimoto, A.; Takeuchi, H.; Aigaki, T.; Matsuzaki, F.; Nakagoshi, H.; Tanimura, T.; et al. Getdb, a database compiling expression patterns and molecular locations of a collection of gal4 enhancer traps. Genesis 2002, 34, 58–61. [Google Scholar] [CrossRef]
- Horn, C.; Offen, N.; Nystedt, S.; Hacker, U.; Wimmer, E.A. Piggybac-based insertional mutagenesis and enhancer detection as a tool for functional insect genomics. Genetics 2003, 163, 647–661. [Google Scholar]
- Jenett, A.; Rubin, G.M.; Ngo, T.T.; Shepherd, D.; Murphy, C.; Dionne, H.; Pfeiffer, B.D.; Cavallaro, A.; Hall, D.; Jeter, J.; et al. A gal4-driver line resource for drosophila neurobiology. Cell Rep. 2012, 2, 991–1001. [Google Scholar] [CrossRef]
- Stark, A.; Dickson, B.J. Vt (vienna tile) gal4 Driver Lines. Available online: http://stockcenter.vdrc.at/control/vtlibrary/ (accessed on 10 January 2014).
- Suster, M.L.; Seugnet, L.; Bate, M.; Sokolowski, M.B. Refining gal4-driven transgene expression in drosophila with a gal80 enhancer-trap. Genesis 2004, 39, 240–245. [Google Scholar] [CrossRef]
- McGuire, S.E.; Le, P.T.; Osborn, A.J.; Matsumoto, K.; Davis, R.L. Spatiotemporal rescue of memory dysfunction in drosophila. Science 2003, 302, 1765–1768. [Google Scholar] [CrossRef]
- Lee, T.; Luo, L. Mosaic analysis with a repressible cell marker for studies of gene function in neuronal morphogenesis. Neuron 1999, 22, 451–461. [Google Scholar] [CrossRef]
- Han, D.D.; Stein, D.; Stevens, L.M. Investigating the function of follicular subpopulations during drosophila oogenesis through hormone-dependent enhancer-targeted cell ablation. Development 2000, 127, 573–583. [Google Scholar]
- Osterwalder, T.; Yoon, K.S.; White, B.H.; Keshishian, H. A conditional tissue-specific transgene expression system using inducible gal4. Proc. Natl. Acad. Sci. USA 2001, 98, 12596–12601. [Google Scholar] [CrossRef]
- Roman, G.; Endo, K.; Zong, L.; Davis, R.L. P[switch], a system for spatial and temporal control of gene expression in drosophila melanogaster. Proc. Natl. Acad. Sci. USA 2001, 98, 12602–12607. [Google Scholar] [CrossRef]
- Stebbins, M.J.; Urlinger, S.; Byrne, G.; Bello, B.; Hillen, W.; Yin, J.C. Tetracycline-inducible systems for drosophila. Proc. Natl. Acad. Sci. USA 2001, 98, 10775–10780. [Google Scholar] [CrossRef]
- Lai, S.L.; Lee, T. Genetic mosaic with dual binary transcriptional systems in drosophila. Nat. Neurosci. 2006, 9, 703–709. [Google Scholar] [CrossRef]
- Potter, C.J.; Tasic, B.; Russler, E.V.; Liang, L.; Luo, L. The q system: A repressible binary system for transgene expression, lineage tracing, and mosaic analysis. Cell 2010, 141, 536–548. [Google Scholar] [CrossRef]
- Yagi, R.; Mayer, F.; Basler, K. Refined lexa transactivators and their use in combination with the drosophila gal4 system. Proc. Natl. Acad. Sci. USA 2010, 107, 16166–16171. [Google Scholar] [CrossRef]
- Dang, D.T.; Perrimon, N. Use of a yeast site-specific recombinase to generate embryonic mosaics in drosophila. Dev. Genet. 1992, 13, 367–375. [Google Scholar] [CrossRef]
- Harrison, D.A.; Perrimon, N. Simple and efficient generation of marked clones in drosophila. Curr. Biol. 1993, 3, 424–433. [Google Scholar] [CrossRef]
- Golic, K.G. Site-specific recombination between homologous chromosomes in drosophila. Science 1991, 252, 958–961. [Google Scholar]
- Yu, H.H.; Chen, C.H.; Shi, L.; Huang, Y.; Lee, T. Twin-spot marcm to reveal the developmental origin and identity of neurons. Nat. Neurosci. 2009, 12, 947–953. [Google Scholar] [CrossRef]
- Venken, K.J.; Schulze, K.L.; Haelterman, N.A.; Pan, H.; He, Y.; Evans-Holm, M.; Carlson, J.W.; Levis, R.W.; Spradling, A.C.; Hoskins, R.A.; et al. Mimic: A highly versatile transposon insertion resource for engineering drosophila melanogaster genes. Nat. Methods 2011, 8, 737–743. [Google Scholar] [CrossRef]
- Fire, A.; Xu, S.; Montgomery, M.K.; Kostas, S.A.; Driver, S.E.; Mello, C.C. Potent and specific genetic interference by double-stranded RNA in caenorhabditis elegans. Nature 1998, 391, 806–811. [Google Scholar] [CrossRef]
- Hannon, G.J. RNA interference. Nature 2002, 418, 244–251. [Google Scholar] [CrossRef]
- Kennerdell, J.R.; Carthew, R.W. Use of dsrna-mediated genetic interference to demonstrate that frizzled and frizzled 2 act in the wingless pathway. Cell 1998, 95, 1017–1026. [Google Scholar] [CrossRef]
- Misquitta, L.; Paterson, B.M. Targeted disruption of gene function in drosophila by RNA interference (RNA-i): A role for nautilus in embryonic somatic muscle formation. Proc. Natl. Acad. Sci. USA 1999, 96, 1451–1456. [Google Scholar] [CrossRef]
- Hammond, S.M.; Bernstein, E.; Beach, D.; Hannon, G.J. An RNA-directed nuclease mediates post-transcriptional gene silencing in drosophila cells. Nature 2000, 404, 293–296. [Google Scholar] [CrossRef]
- Kennerdell, J.R.; Carthew, R.W. Heritable gene silencing in drosophila using double-stranded RNA. Nat. Biotechnol. 2000, 18, 896–898. [Google Scholar] [CrossRef]
- Clemens, J.C.; Worby, C.A.; Simonson-Leff, N.; Muda, M.; Maehama, T.; Hemmings, B.A.; Dixon, J.E. Use of double-stranded RNA interference in drosophila cell lines to dissect signal transduction pathways. Proc. Natl. Acad. Sci. USA 2000, 97, 6499–6503. [Google Scholar]
- Kiger, A.A.; Baum, B.; Jones, S.; Jones, M.R.; Coulson, A.; Echeverri, C.; Perrimon, N. A functional genomic analysis of cell morphology using RNA interference. J. Biol. 2003. [Google Scholar] [CrossRef] [Green Version]
- Lum, L.; Yao, S.; Mozer, B.; Rovescalli, A.; von Kessler, D.; Nirenberg, M.; Beachy, P.A. Identification of hedgehog pathway components by RNAi in drosophila cultured cells. Science 2003, 299, 2039–2045. [Google Scholar] [CrossRef]
- Boutros, M.; Kiger, A.A.; Armknecht, S.; Kerr, K.; Hild, M.; Koch, B.; Haas, S.A.; Paro, R.; Perrimon, N.; Heidelberg Fly Array, C. Genome-wide RNAi analysis of growth and viability in drosophila cells. Science 2004, 303, 832–835. [Google Scholar] [CrossRef]
- Foley, E.; O’Farrell, P.H. Functional dissection of an innate immune response by a genome-wide RNAi screen. PLoS Biol. 2004, 2, E203. [Google Scholar] [CrossRef] [Green Version]
- Dietzl, G.; Chen, D.; Schnorrer, F.; Su, K.-C.; Barinova, Y.; Fellner, M.; Gasser, B.; Kinsey, K.; Oppel, S.; Scheiblauer, S.; et al. A genome-wide transgenic RNAi library for conditional gene inactivation in drosophila. Nature 2007, 448, 151–156. [Google Scholar] [CrossRef]
- Ueda, R. RNAi Fly—A Comprehensive RNAi-Mutant Fly Bank. Available online: http://www.shigen.nig.ac.jp/fly/nigfly/about/aboutrnai.jsp/ (accessed on 13 January 2014).
- Ni, J.-Q.; Liu, L.-P.; Binari, R.; Hardy, R.; Shim, H.-S.; Cavallaro, A.; Booker, M.; Pfeiffer, B.D.; Markstein, M.; Wang, H.; et al. A drosophila resource of transgenic RNAi lines for neurogenetics. Genetics 2009, 182, 1089–1100. [Google Scholar] [CrossRef]
- Schnorrer, F.; Schonbauer, C.; Langer, C.C.; Dietzl, G.; Novatchkova, M.; Schernhuber, K.; Fellner, M.; Azaryan, A.; Radolf, M.; Stark, A.; et al. Systematic genetic analysis of muscle morphogenesis and function in drosophila. Nature 2010, 464, 287–291. [Google Scholar] [CrossRef]
- Neely, G.G.; Kuba, K.; Cammarato, A.; Isobe, K.; Amann, S.; Zhang, L.; Murata, M.; Elmen, L.; Gupta, V.; Arora, S.; et al. A global in vivo drosophila RNAi screen identifies not3 as a conserved regulator of heart function. Cell 2010, 141, 142–153. [Google Scholar] [CrossRef]
- Pospisilik, J.A.; Schramek, D.; Schnidar, H.; Cronin, S.J.; Nehme, N.T.; Zhang, X.; Knauf, C.; Cani, P.D.; Aumayr, K.; Todoric, J.; et al. Drosophila genome-wide obesity screen reveals hedgehog as a determinant of brown versus white adipose cell fate. Cell 2010, 140, 148–160. [Google Scholar] [CrossRef]
- Neely, G.G.; Hess, A.; Costigan, M.; Keene, A.C.; Goulas, S.; Langeslag, M.; Griffin, R.S.; Belfer, I.; Dai, F.; Smith, S.B.; et al. A genome-wide drosophila screen for heat nociception identifies alpha2delta3 as an evolutionarily conserved pain gene. Cell 2010, 143, 628–638. [Google Scholar]
- Ghosh, A.; Kling, T.; Snaidero, N.; Sampaio, J.L.; Shevchenko, A.; Gras, H.; Geurten, B.; Gopfert, M.C.; Schulz, J.B.; Voigt, A.; et al. A global in vivo drosophila RNAi screen identifies a key role of ceramide phosphoethanolamine for glial ensheathment of axons. PLoS Genet. 2013, 9, e1003980. [Google Scholar]
- Czech, B.; Preall, J.B.; McGinn, J.; Hannon, G.J. A transcriptome-wide RNAi screen in the drosophila ovary reveals factors of the germline piRNA pathway. Mol. Cell 2013, 50, 749–761. [Google Scholar] [CrossRef]
- Kondo, S.; Booker, M.; Perrimon, N. Cross-species RNAi rescue platform in drosophila melanogaster. Genetics 2009, 183, 1165–1173. [Google Scholar]
- Langer, C.C.H.; Ejsmont, R.K.; Schönbauer, C.; Schnorrer, F.; Tomancak, P. In vivo RNAi rescue in drosophila melanogaster with genomic transgenes from drosophila pseudoobscura. PLoS One 2010, 5, e8928. [Google Scholar]
- Schulz, J.G.; David, G.; Hassan, B.A. A novel method for tissue-specific RNAi rescue in drosophila. Nucleic Acids Res. 2009, 37, e93. [Google Scholar]
- Lawrence, P.A.; Bodmer, R.; Vincent, J.P. Segmental patterning of heart precursors in drosophila. Development 1995, 121, 4303–4308. [Google Scholar]
- Murray, M.J.; Merritt, D.J.; Brand, A.H.; Whitington, P.M. In vivo dynamics of axon pathfinding in the drosophilia CNS: A time-lapse study of an identified motorneuron. J. Neurobiol. 1998, 37, 607–621. [Google Scholar] [CrossRef]
- Murphy, K.C. Use of bacteriophage lambda recombination functions to promote gene replacement in escherichia coli. J. Bacteriol. 1998, 180, 2063–2071. [Google Scholar]
- Zhang, Y.; Buchholz, F.; Muyrers, J.P.; Stewart, A.F. A new logic for DNA engineering using recombination in escherichia coli. Nat. Genet. 1998, 20, 123–128. [Google Scholar]
- Muyrers, J.P.; Zhang, Y.; Testa, G.; Stewart, A.F. Rapid modification of bacterial artificial chromosomes by et-recombination. Nucleic Acids Res. 1999, 27, 1555–1557. [Google Scholar] [CrossRef]
- Yu, D.; Ellis, H.M.; Lee, E.C.; Jenkins, N.A.; Copeland, N.G.; Court, D.L. An efficient recombination system for chromosome engineering in Escherichia coli. Proc. Natl. Acad. Sci. USA 2000, 97, 5978–5983. [Google Scholar]
- Venken, K.J.T.; Carlson, J.W.; Schulze, K.L.; Pan, H.; He, Y.; Spokony, R.; Wan, K.H.; Koriabine, M.; de Jong, P.J.; White, K.P.; et al. Versatile P[acman] bac libraries for transgenesis studies in drosophila melanogaster. Nat. Methods 2009, 6, 431–434. [Google Scholar]
- Ejsmont, R.K.; Sarov, M.; Winkler, S.; Lipinski, K.A.; Tomancak, P. A toolkit for high-throughput, cross-species gene engineering in drosophila. Nat. Methods 2009, 6, 435–437. [Google Scholar]
- Horn, C.; Jaunich, B.; Wimmer, E.A. Highly sensitive, fluorescent transformation marker for drosophila transgenesis. Dev. Genes Evol. 2000, 210, 623–629. [Google Scholar] [CrossRef]
- Poser, I.; Sarov, M.; Hutchins, J.R.; Heriche, J.K.; Toyoda, Y.; Pozniakovsky, A.; Weigl, D.; Nitzsche, A.; Hegemann, B.; Bird, A.W.; et al. Bac transgeneomics: A high-throughput method for exploration of protein function in mammals. Nat. Methods 2008, 5, 409–415. [Google Scholar]
- Wu, J.S.; Luo, L. A protocol for mosaic analysis with a repressible cell marker (Marcm) in drosophila. Nat. Protoc. 2006, 1, 2583–2589. [Google Scholar]
- Rong, Y.S.; Golic, K.G. Gene targeting by homologous recombination in drosophila. Science 2000, 288, 2013–2018. [Google Scholar] [CrossRef]
- Gong, W.J.; Golic, K.G. Ends-out, or replacement, gene targeting in drosophila. Proc. Natl. Acad. Sci. USA 2003, 100, 2556–2561. [Google Scholar] [CrossRef]
- Choi, C.M.; Vilain, S.; Langen, M.; van Kelst, S.; de Geest, N.; Yan, J.; Verstreken, P.; Hassan, B.A. Conditional mutagenesis in drosophila. Science 2009. [Google Scholar] [CrossRef]
- Huang, J.; Zhou, W.; Dong, W.; Watson, A.M.; Hong, Y. From the cover: Directed, efficient, and versatile modifications of the drosophila genome by genomic engineering. Proc. Natl. Acad. Sci. USA 2009, 106, 8284–8289. [Google Scholar]
- Gloor, G.B.; Nassif, N.A.; Johnson-Schlitz, D.M.; Preston, C.R.; Engels, W.R. Targeted gene replacement in drosophila via p element-induced gap repair. Science 1991, 253, 1110–1117. [Google Scholar] [CrossRef]
- Kim, Y.G.; Cha, J.; Chandrasegaran, S. Hybrid restriction enzymes: Zinc finger fusions to Fok I cleavage domain. Proc. Natl. Acad. Sci. USA 1996, 93, 1156–1160. [Google Scholar] [CrossRef]
- Bibikova, M.; Golic, M.; Golic, K.G.; Carroll, D. Targeted chromosomal cleavage and mutagenesis in drosophila using zinc-finger nucleases. Genetics 2002, 161, 1169–1175. [Google Scholar]
- Auer, T.O.; Duroure, K.; de Cian, A.; Concordet, J.P.; Del Bene, F. Highly efficient crispr/cas9-mediated knock-in in zebrafish by homology-independent DNA repair. Genome Res. 2014, 24, 142–153. [Google Scholar] [CrossRef]
- Beumer, K.; Bhattacharyya, G.; Bibikova, M.; Trautman, J.K.; Carroll, D. Efficient gene targeting in drosophila with zinc-finger nucleases. Genetics 2006, 172, 2391–2403. [Google Scholar]
- Maeder, M.L.; Thibodeau-Beganny, S.; Osiak, A.; Wright, D.A.; Anthony, R.M.; Eichtinger, M.; Jiang, T.; Foley, J.E.; Winfrey, R.J.; Townsend, J.A.; et al. Rapid “open-source” engineering of customized zinc-finger nucleases for highly efficient gene modification. Mol. Cell 2008, 31, 294–301. [Google Scholar] [CrossRef]
- Sander, J.D.; Dahlborg, E.J.; Goodwin, M.J.; Cade, L.; Zhang, F.; Cifuentes, D.; Curtin, S.J.; Blackburn, J.S.; Thibodeau-Beganny, S.; Qi, Y.; et al. Selection-free zinc-finger-nuclease engineering by context-dependent assembly (coda). Nat. Methods 2011, 8, 67–69. [Google Scholar] [CrossRef]
- Kim, S.; Lee, M.J.; Kim, H.; Kang, M.; Kim, J.S. Preassembled zinc-finger arrays for rapid construction of zfns. Nat. Methods 2011. [Google Scholar] [CrossRef]
- Doyon, Y.; Vo, T.D.; Mendel, M.C.; Greenberg, S.G.; Wang, J.; Xia, D.F.; Miller, J.C.; Urnov, F.D.; Gregory, P.D.; Holmes, M.C. Enhancing zinc-finger-nuclease activity with improved obligate heterodimeric architectures. Nat. Methods 2011, 8, 74–79. [Google Scholar]
- Mahfouz, M.M.; Li, L.; Shamimuzzaman, M.; Wibowo, A.; Fang, X.; Zhu, J.K. De novo-engineered transcription activator-like effector (tale) hybrid nuclease with novel DNA binding specificity creates double-strand breaks. Proc. Natl. Acad. Sci. USA 2011, 108, 2623–2628. [Google Scholar]
- Miller, J.C.; Tan, S.; Qiao, G.; Barlow, K.A.; Wang, J.; Xia, D.F.; Meng, X.; Paschon, D.E.; Leung, E.; Hinkley, S.J.; et al. A tale nuclease architecture for efficient genome editing. Nat. Biotechnol. 2011, 29, 143–148. [Google Scholar] [CrossRef]
- Morbitzer, R.; Elsaesser, J.; Hausner, J.; Lahaye, T. Assembly of custom tale-type DNA binding domains by modular cloning. Nucleic Acids Res. 2011, 39, 5790–5799. [Google Scholar] [CrossRef]
- Moscou, M.J.; Bogdanove, A.J. A simple cipher governs DNA recognition by tal effectors. Science 2009. [Google Scholar] [CrossRef]
- Brouns, S.J.; Jore, M.M.; Lundgren, M.; Westra, E.R.; Slijkhuis, R.J.; Snijders, A.P.; Dickman, M.J.; Makarova, K.S.; Koonin, E.V.; van der Oost, J. Small crispr rnas guide antiviral defense in prokaryotes. Science 2008, 321, 960–964. [Google Scholar] [CrossRef]
- Gasiunas, G.; Barrangou, R.; Horvath, P.; Siksnys, V. Cas9-crrna ribonucleoprotein complex mediates specific DNA cleavage for adaptive immunity in bacteria. Proc. Natl. Acad. Sci. USA 2012, 109, E2579–E2586. [Google Scholar]
- Jinek, M.; Chylinski, K.; Fonfara, I.; Hauer, M.; Doudna, J.A.; Charpentier, E. A programmable dual-RNA-guided DNA endonuclease in adaptive bacterial immunity. Science 2012, 337, 816–821. [Google Scholar] [CrossRef]
- Bassett, A.R.; Tibbit, C.; Ponting, C.P.; Liu, J.L. Highly efficient targeted mutagenesis of drosophila with the crispr/cas9 system. Cell Rep. 2013, 4, 220–228. [Google Scholar] [CrossRef]
- Gratz, S.J.; Cummings, A.M.; Nguyen, J.N.; Hamm, D.C.; Donohue, L.K.; Harrison, M.M.; Wildonger, J.; O’Connor-Giles, K.M. Genome engineering of drosophila with the crispr RNA-guided cas9 nuclease. Genetics 2013, 194, 1029–1035. [Google Scholar] [CrossRef]
- Yu, Z.; Ren, M.; Wang, Z.; Zhang, B.; Rong, Y.S.; Jiao, R.; Gao, G. Highly efficient genome modifications mediated by crispr/cas9 in drosophila. Genetics 2013, 195, 289–291. [Google Scholar] [CrossRef]
- Sebo, Z.L.; Lee, H.B.; Peng, Y.; Guo, Y. A simplified and efficient germline-specific crispr/cas9 system for drosophila genomic engineering. Fly 2014, 8, 52–57. [Google Scholar]
- Ren, X.; Sun, J.; Housden, B.E.; Hu, Y.; Roesel, C.; Lin, S.; Liu, L.-P.; Yang, Z.; Mao, D.; Sun, L.; et al. Optimized gene editing technology for drosophila melanogaster using germ line-specific cas9. Proc. Natl. Acad. Sci. USA 2013, 110, 19012–19017. [Google Scholar] [CrossRef]
- Gratz, S.J.; Ukken, F.P.; Rubinstein, C.D.; Thiede, G.; Donohue, L.K.; Cummings, A.M.; O’Connor-Giles, K.M. Highly specific and efficient crispr/cas9-catalyzed homology-directed repair in drosophila. Genetics 2014. [Google Scholar] [CrossRef]
- Kondo, S.; Ueda, R. Highly improved gene targeting by germline-specific cas9 expression in drosophila. Genetics 2013, 195, 715–721. [Google Scholar] [CrossRef]
- Baena-Lopez, L.A.; Alexandre, C.; Mitchell, A.; Pasakarnis, L.; Vincent, J.P. Accelerated homologous recombination and subsequent genome modification in drosophila. Development 2013, 140, 4818–4825. [Google Scholar] [CrossRef]
- Del Valle Rodriguez, A.; Didiano, D.; Desplan, C. Power tools for gene expression and clonal analysis in drosophila. Nat. Methods 2012, 9, 47–55. [Google Scholar]
- Venken, K.J.; Bellen, H.J. Genome-wide manipulations of drosophila melanogaster with transposons, flp recombinase, and phic31 integrase. Methods Mol. Biol. 2012, 859, 203–228. [Google Scholar] [CrossRef]
- Gratz, S.J.; Wildonger, J.; Harrison, M.M.; O’Connor-Giles, K.M. Crispr/cas9-mediated genome engineering and the promise of designer flies on demand. Fly 2013. [Google Scholar] [CrossRef]
- Reiter, L.T.; Potocki, L.; Chien, S.; Gribskov, M.; Bier, E. A systematic analysis of human disease-associated gene sequences in drosophila melanogaster. Genome Res. 2001, 11, 1114–1125. [Google Scholar] [CrossRef]
- Chien, S.; Reiter, L.T.; Bier, E.; Gribskov, M. Homophila: Human disease gene cognates in drosophila. Nucleic Acids Res. 2002, 30, 149–151. [Google Scholar] [CrossRef]
- Richter, A.; Boch, J. Designer tales team up for highly efficient gene induction. Nat. Methods 2013, 10, 207–208. [Google Scholar] [CrossRef]
- Crocker, J.; Stern, D.L. Tale-mediated modulation of transcriptional enhancers in vivo. Nat. Methods 2013, 10, 762–767. [Google Scholar] [CrossRef]
- Mali, P.; Esvelt, K.M.; Church, G.M. Cas9 as a versatile tool for engineering biology. Nat. Methods 2013, 10, 957–963. [Google Scholar] [CrossRef]
- modENCODE Consortium Modencode Comparative Genomics Whitepaper. Available online: http://www.genome.gov/Pages/Research/Sequencing/SeqProposals/modENCODE_ComparativeGenomics_WhitePaper.pdf (accessed on 16 January 2014).
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
MDPI and ACS Style
Ejsmont, R.K.; Hassan, B.A. The Little Fly that Could: Wizardry and Artistry of Drosophila Genomics. Genes 2014, 5, 385-414. https://doi.org/10.3390/genes5020385
AMA Style
Ejsmont RK, Hassan BA. The Little Fly that Could: Wizardry and Artistry of Drosophila Genomics. Genes. 2014; 5(2):385-414. https://doi.org/10.3390/genes5020385
Chicago/Turabian StyleEjsmont, Radoslaw K., and Bassem A. Hassan. 2014. "The Little Fly that Could: Wizardry and Artistry of Drosophila Genomics" Genes 5, no. 2: 385-414. https://doi.org/10.3390/genes5020385