
Reappearance from Obscurity: Mammalian Rad52 in Homologous Recombination

{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
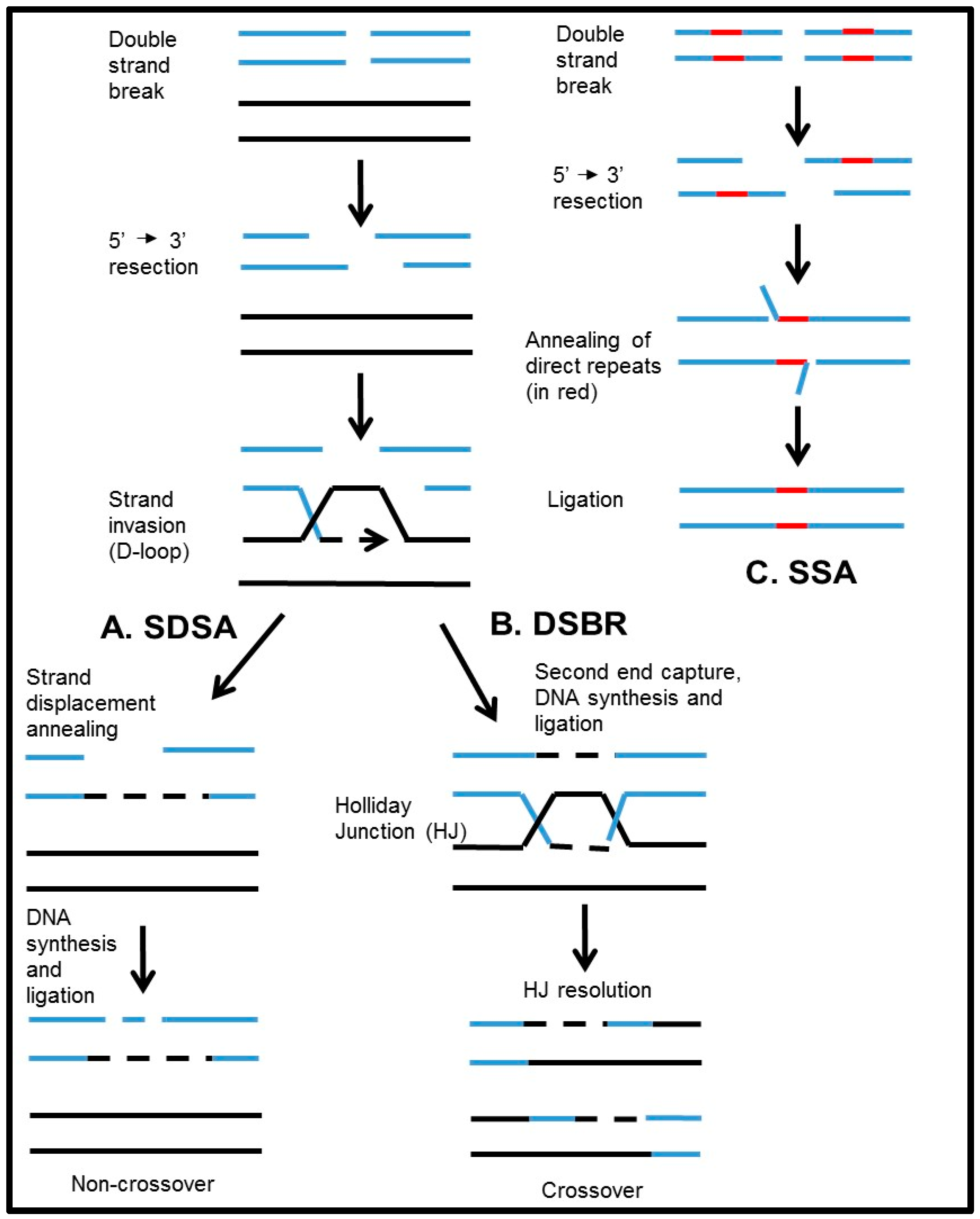
2. The Role of RAD52 in HR in Mammals
3. Activities of Rad52 Protein
3.1. ssDNA Annealing Activity
3.2. Stimulation of Rad51
3.3. DNA Invasion or Strand Exchange
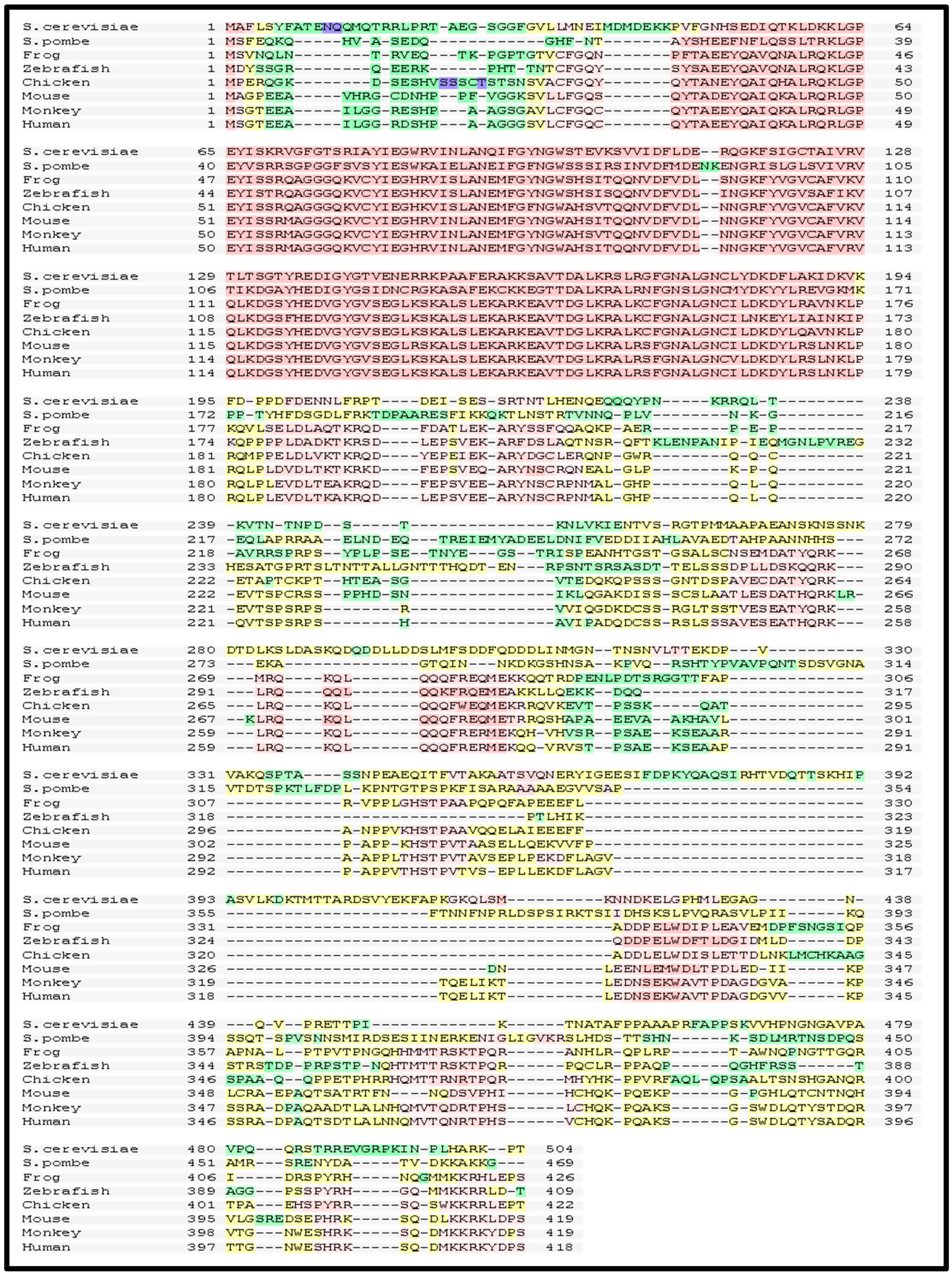
4. Role of the Structural Domains of Rad52
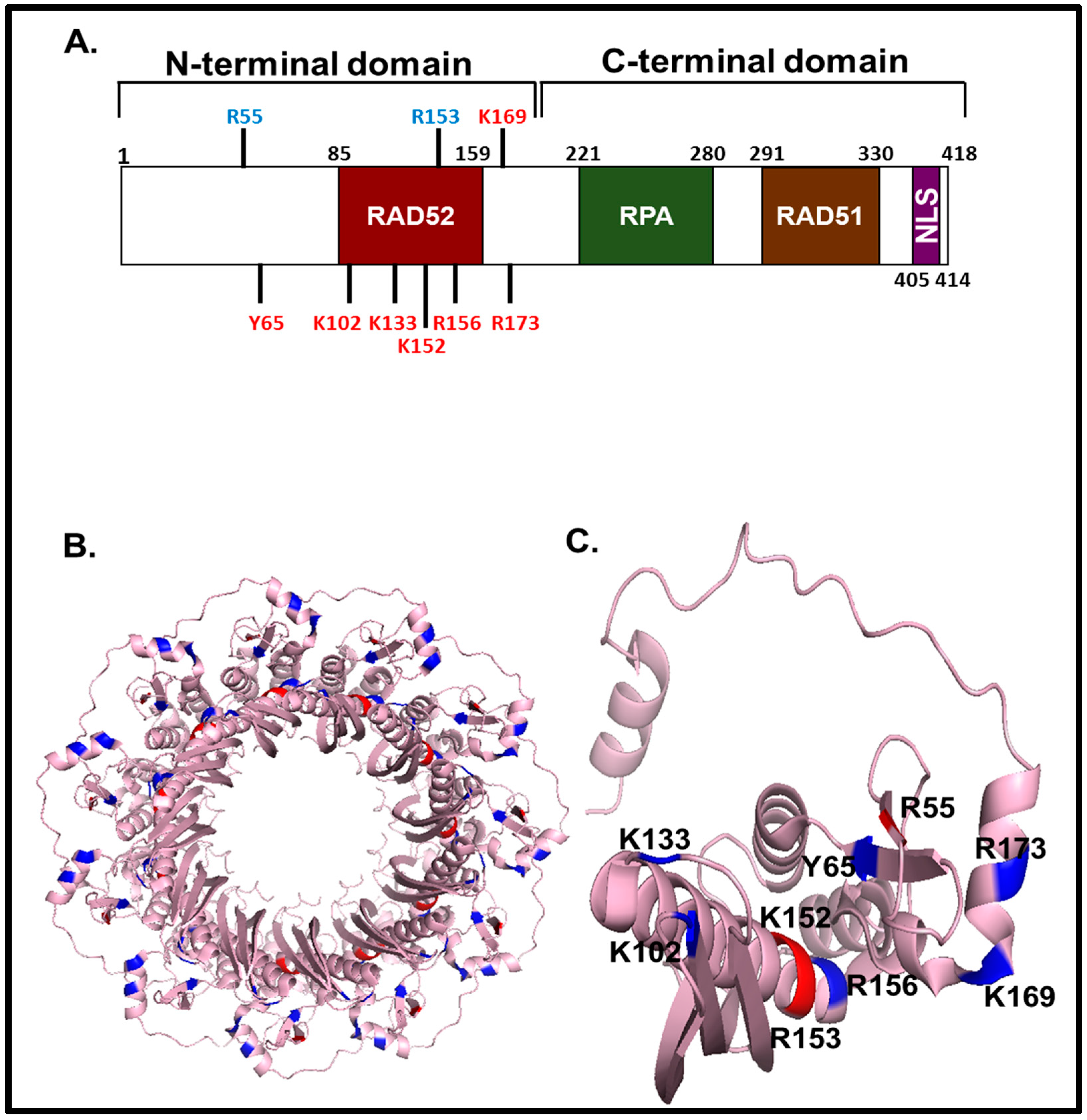
4.1. DNA Binding
4.2. RAD52 Multimerization
4.3. Interactions of RAD52 with RPA, RAD51 and MUS81
4.4. Nuclear Localization Signal: Cellular Localization of RAD52 and its Truncated Isomers
5. Regulation of RAD52 in the Cell
5.1. Cell Cycle Regulation of RAD52 Protein
5.2. Post-Translational Modification of RAD52
6. RAD52 in RNA-directed DNA Repair
7. RAD52 as a Therapeutic Target
8. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interests
References
- San Filippo, J.; Sung, P.; Klein, H. Mechanism of eukaryotic homologous recombination. Ann. Rev. Biochem. 2008, 77, 229–257. [Google Scholar] [CrossRef] [PubMed]
- Pâques, F.; Haber, J.E. Multiple pathways of recombination induced by double-strand breaks in Saccharomyces cerevisiae. Microbiol. Mol. Biol. Rev. 1999, 63, 349–404. [Google Scholar] [PubMed]
- Allers, T.; Lichten, M. Differential timing and control of noncrossover and crossover recombination during meiosis. Cell 2001, 106, 47–57. [Google Scholar] [CrossRef]
- Wu, L.; Hickson, I.D. The bloom’s syndrome helicase suppresses crossing over during homologous recombination. Nature 2003, 426, 870–874. [Google Scholar] [CrossRef] [PubMed]
- Bugreev, D.V.; Hanaoka, F.; Mazin, A.V. Rad54 dissociates homologous recombination intermediates by branch migration. Nat. Struct. Mol. Biol. 2007, 14, 746–753. [Google Scholar] [CrossRef] [PubMed]
- Llorente, B.; Smith, C.E.; Symington, L.S. Break-induced replication: What is it and what is it for? Cell Cycle 2008, 7, 859–864. [Google Scholar] [CrossRef] [PubMed]
- McEachern, M.J.; Haber, J.E. Break-induced replication and recombinational telomere elongation in yeast. Ann. Rev. Biochem. 2006, 75, 111–135. [Google Scholar] [CrossRef] [PubMed]
- Game, J.C.; Mortimer, R.K. A genetic study of x-ray sensitive mutants in yeast. Mutat. Res. 1974, 24, 281. [Google Scholar] [CrossRef]
- Symington, L.S. Role of Rad52 epistasis group genes in homologous recombination and double-strand break repair. Microbiol. Mol. Biol. Rev. 2002, 66, 630–670. [Google Scholar] [CrossRef] [PubMed]
- Malone, R.E.; Esposito, R.E. The RAD52 gene is required for homothallic interconversion of mating types and spontaneous mitotic recombination in yeast. Proc. Natl. Acad. Sci. USA 1980, 77, 503–507. [Google Scholar] [CrossRef] [PubMed]
- Schiestl, R.H.; Dominska, M.; Petes, T.D. Transformation of Saccharomyces cerevisiae with nonhomologous DNA: Illegitimate integration of transforming DNA into yeast chromosomes and in vivo ligation of transforming DNA to mitochondrial DNA sequences. Mol. Cell. Biol. 1993, 13, 2697–2705. [Google Scholar] [CrossRef] [PubMed]
- Rijkers, T.; van den Ouweland, J.; Morolli, B.; Rolink, A.G.; Baarends, W.M.; van Sloun, P.P.; Lohman, P.H.; Pastink, A. Targeted inactivation of mouse RAD52 reduces homologous recombination but not resistance to ionizing radiation. Mol. Cell. Biol. 1998, 18, 6423–6429. [Google Scholar] [CrossRef] [PubMed]
- Lok, B.H.; Carley, A.C.; Tchang, B.; Powell, S.N. RAD52 inactivation is synthetically lethal with deficiencies in BRCA1 and PALB2 in addition to BRCA2 through RAD51-mediated homologous recombination. Oncogene 2013, 32, 3552–3558. [Google Scholar] [CrossRef] [PubMed]
- Lok, B.H.; Powell, S.N. Molecular pathways: Understanding the role of RAD52 in homologous recombination for therapeutic advancement. Clin. Cancer Res. 2012, 18, 6400–6406. [Google Scholar] [CrossRef] [PubMed]
- Feng, Z.; Scott, S.P.; Bussen, W.; Sharma, G.G.; Guo, G.; Pandita, T.K.; Powell, S.N. RAD52 inactivation is synthetically lethal with BRCA2 deficiency. Proc. Natl. Acad. Sci. USA 2011, 108, 686–691. [Google Scholar] [CrossRef] [PubMed]
- Chandramouly, G.; McDevitt, S.; Sullivan, K.; Kent, T.; Luz, A.; Glickman, J.F.; Andrake, M.; Skorski, T.; Pomerantz, R.T. Small-molecule disruption of RAD52 rings as a mechanism for precision medicine in BRCA-deficient cancers. Chem. Biol. 2015, 22, 1491–1504. [Google Scholar] [CrossRef] [PubMed]
- Huang, F.; Goyal, N.; Sullivan, K.; Hanamshet, K.; Patel, M.; Mazina, O.M.; Wang, C.X.; An, W.F.; Spoonamore, J.; Metkar, S.; et al. Targeting BRCA1- and BRCA2-deficient cells with RAD52 small molecule inhibitors. Nucleic Acids Res. 2016. [Google Scholar] [CrossRef] [PubMed]
- Sullivan, K.; Cramer-Morales, K.; McElroy, D.L.; Ostrov, D.A.; Haas, K.; Childers, W.; Hromas, R.; Skorski, T. Identification of a small molecule inhibitor of RAD52 by structure-based selection. PLoS ONE 2016, 11, e0147230. [Google Scholar] [CrossRef] [PubMed]
- Cramer-Morales, K.; Nieborowska-Skorska, M.; Scheibner, K.; Padget, M.; Irvine, D.A.; Sliwinski, T.; Haas, K.; Lee, J.; Geng, H.; Roy, D.; et al. Personalized synthetic lethality induced by targeting RAD52 in leukemias identified by gene mutation and expression profile. Blood 2013, 122, 1293–1304. [Google Scholar] [CrossRef] [PubMed]
- Keskin, H.; Shen, Y.; Huang, F.; Patel, M.; Yang, T.; Ashley, K.; Mazin, A.V.; Storici, F. Transcript-RNA-templated DNA recombination and repair. Nature 2014, 515, 436–439. [Google Scholar] [CrossRef] [PubMed]
- Petrini, J.H.; Bressan, D.A.; Yao, M.S. The RAD52 epistasis group in mammalian double strand break repair. Semin. Immunol. 1997, 9, 181–188. [Google Scholar] [CrossRef] [PubMed]
- Sung, P.; Klein, H. Mechanism of homologous recombination: Mediators and helicases take on regulatory functions. Nat. Rev. Mol. Cell Biol. 2006, 7, 739–750. [Google Scholar] [CrossRef] [PubMed]
- Yamaguchi-Iwai, Y.; Sonoda, E.; Buerstedde, J.M.; Bezzubova, O.; Morrison, C.; Takata, M.; Shinohara, A.; Takeda, S. Homologous recombination, but not DNA repair, is reduced in vertebrate cells deficient in RAD52. Mol. Cell. Biol. 1998, 18, 6430–6435. [Google Scholar] [CrossRef] [PubMed]
- Park, M.S. Expression of human RAD52 confers resistance to ionizing radiation in mammalian cells. J. Biol. Chem. 1995, 270, 15467–15470. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Li, M.; Lee, E.Y.; Maizels, N. Localization and dynamic relocalization of mammalian RAD52 during the cell cycle and in response to DNA damage. Curr. Biol. 1999, 9, 975–978. [Google Scholar] [CrossRef]
- Liu, Y.; Maizels, N. Coordinated response of mammalian RAD51 and RAD52 to DNA damage. EMBO Rep. 2000, 1, 85–90. [Google Scholar] [CrossRef] [PubMed]
- Maser, R.S.; Monsen, K.J.; Nelms, B.E.; Petrini, J.H. hMre11 and hRad50 nuclear foci are induced during the normal cellular response to DNA double-strand breaks. Mol. Cell. Biol. 1997, 17, 6087–6096. [Google Scholar] [CrossRef] [PubMed]
- Chun, J.; Buechelmaier, E.S.; Powell, S.N. Rad51 paralog complexes BCDX2 and CX3 act at different stages in the BRCA1-BRCA2-dependent homologous recombination pathway. Mol. Cell. Biol. 2013, 33, 387–395. [Google Scholar] [CrossRef] [PubMed]
- Fujimori, A.; Tachiiri, S.; Sonoda, E.; Thompson, L.H.; Dhar, P.K.; Hiraoka, M.; Takeda, S.; Zhang, Y.; Reth, M.; Takata, M. Rad52 partially substitutes for the Rad51 paralog XRCC3 in maintaining chromosomal integrity in vertebrate cells. EMBO J. 2001, 20, 5513–5520. [Google Scholar] [CrossRef] [PubMed]
- Mortensen, U.H.; Bendixen, C.; Sunjevaric, I.; Rothstein, R. DNA strand annealing is promoted by the yeast Rad52 protein. Proc. Natl. Acad. Sci. USA 1996, 93, 10729–10734. [Google Scholar] [CrossRef] [PubMed]
- Sugiyama, T.; New, J.H.; Kowalczykowski, S.C. DNA annealing by Rad52 protein is stimulated by specific interaction with the complex of replication protein A and single-stranded DNA. Proc. Natl. Acad. Sci. USA 1998, 95, 6049–6054. [Google Scholar] [CrossRef] [PubMed]
- Reddy, G.; Golub, E.I.; Radding, C.M. Human RAD52 protein promotes single-strand DNA annealing followed by branch migration. Mutat. Res. 1997, 377, 53–59. [Google Scholar] [CrossRef]
- Kagawa, W.; Kagawa, A.; Saito, K.; Ikawa, S.; Shibata, T.; Kurumizaka, H.; Yokoyama, S. Identification of a second DNA binding site in the human RAD52 protein. J. Biol. Chem. 2008, 283, 24264–24273. [Google Scholar] [CrossRef] [PubMed]
- Bi, B.; Rybalchenko, N.; Golub, E.I.; Radding, C.M. Human and yeast Rad52 proteins promote DNA strand exchange. Proc. Natl. Acad. Sci. USA 2004, 101, 9568–9572. [Google Scholar] [CrossRef] [PubMed]
- New, J.H.; Sugiyama, T.; Zaitseva, E.; Kowalczykowski, S.C. Rad52 protein stimulates DNA strand exchange by Rad51 and replication protein a. Nature 1998, 391, 407–410. [Google Scholar] [PubMed]
- Sung, P. Function of yeast Rad52 protein as a mediator between replication protein A and the Rad51 recombinase. J. Biol. Chem. 1997, 272, 28194–28197. [Google Scholar] [CrossRef] [PubMed]
- Shinohara, A.; Ogawa, T. Stimulation by rad52 of yeast rad51-mediated recombination. Nature 1998, 391, 404–407. [Google Scholar] [CrossRef] [PubMed]
- Jensen, R.B.; Carreira, A.; Kowalczykowski, S.C. Purified human BRCA2 stimulates RAD51-mediated recombination. Nature 2010, 467, 678–683. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Heyer, W.D. Who’s who in human recombination: BRCA2 and RAD52. Proc. Natl. Acad. Sci. USA 2011, 108, 441–442. [Google Scholar] [CrossRef] [PubMed]
- Brosh, R.M., Jr. DNA helicases involved in DNA repair and their roles in cancer. Nat. Rev. Cancer 2013, 13, 542–558. [Google Scholar] [CrossRef] [PubMed]
- Wold, M.S. Replication protein A: A heterotrimeric, single-stranded DNA-binding protein required for eukaryotic DNA metabolism. Ann. Rev. Biochem. 1997, 66, 61–92. [Google Scholar] [CrossRef] [PubMed]
- Singleton, M.R.; Wentzell, L.M.; Liu, Y.; West, S.C.; Wigley, D.B. Structure of the single-strand annealing domain of human RAD52 protein. Proc. Natl. Acad. Sci. USA 2002, 99, 13492–13497. [Google Scholar] [CrossRef] [PubMed]
- Grimme, J.M.; Honda, M.; Wright, R.; Okuno, Y.; Rothenberg, E.; Mazin, A.V.; Ha, T.; Spies, M. Human rad52 binds and wraps single-stranded DNA and mediates annealing via two hRAD52-ssdna complexes. Nucleic Acids Res. 2010, 38, 2917–2930. [Google Scholar] [CrossRef] [PubMed]
- Khade, N.V.; Sugiyama, T. Roles of C-terminal region of yeast and human Rad52 in Rad51-nucleoprotein filament formation and ssdna annealing. PLoS ONE 2016, 11, e0158436. [Google Scholar] [CrossRef] [PubMed]
- Stark, J.M.; Pierce, A.J.; Oh, J.; Pastink, A.; Jasin, M. Genetic steps of mammalian homologous repair with distinct mutagenic consequences. Mol. Cell. Biol. 2004, 24, 9305–9316. [Google Scholar] [CrossRef] [PubMed]
- Shinohara, A.; Shinohara, M.; Ohta, T.; Matsuda, S.; Ogawa, T. Rad52 forms ring structures and co-operates with rpa in single-strand DNA annealing. Genes Cells 1998, 3, 145–156. [Google Scholar] [CrossRef] [PubMed]
- Fishman-Lobell, J.; Rudin, N.; Haber, J.E. Two alternative pathways of double-strand break repair that are kinetically separable and independently modulated. Mol. Cell. Biol. 1992, 12, 1292–1303. [Google Scholar] [CrossRef] [PubMed]
- Ivanov, E.L.; Sugawara, N.; Fishman-Lobell, J.; Haber, J.E. Genetic requirements for the single-strand annealing pathway of double- strand break repair in Saccharomyces cerevisiae. Genetics 1996, 142, 693–704. [Google Scholar] [PubMed]
- Sugawara, N.; Haber, J.E. Characterization of double-strand break-induced recombination: Homology requirements and single-stranded DNA formation. Mol. Cell. Biol. 1992, 12, 563–575. [Google Scholar] [CrossRef] [PubMed]
- Cruz-Garcia, A.; Lopez-Saavedra, A.; Huertas, P. BRCA1 accelerates ctip-mediated DNA-end resection. Cell Rep. 2014, 9, 451–459. [Google Scholar] [CrossRef] [PubMed]
- Sugiyama, T.; Kantake, N.; Wu, Y.; Kowalczykowski, S.C. Rad52-mediated DNA annealing after Rad51-mediated DNA strand exchange promotes second ssdna capture. EMBO J. 2006, 25, 5539–5548. [Google Scholar] [CrossRef] [PubMed]
- McIlwraith, M.J.; West, S.C. DNA repair synthesis facilitates Rad52-mediated second-end capture during dsb repair. Mol. Cell 2008, 29, 510–516. [Google Scholar] [CrossRef] [PubMed]
- Benson, F.E.; Baumann, P.; West, S.C. Synergistic actions of Rad51 and Rad52 in recombination and DNA repair. Nature 1998, 391, 401–404. [Google Scholar] [PubMed]
- Sugiyama, T.; Kowalczykowski, S.C. Rad52 protein associates with replication protein A (RPA)-single-stranded DNA to accelerate Rad51-mediated displacement of RPA and presynaptic complex formation. J. Biol. Chem. 2002, 277, 31663–31672. [Google Scholar] [CrossRef] [PubMed]
- New, J.H.; Kowalczykowski, S.C. Rad52 protein has a second stimulatory role in DNA strand exchange that complements replication protein-a function. J. Biol. Chem. 2002, 277, 26171–26176. [Google Scholar] [CrossRef] [PubMed]
- Kowalczykowski, S.C. Structural biology: Snapshots of DNA repair. Nature 2008, 453, 463–466. [Google Scholar] [CrossRef] [PubMed]
- Sugiyama, T.; Zaitseva, E.M.; Kowalczykowski, S.C. A single-stranded DNA-binding protein is needed for efficient presynaptic complex formation by the Saccharomyces cerevisiae Rad51 protein. J. Biol. Chem. 1997, 272, 7940–7945. [Google Scholar] [CrossRef] [PubMed]
- Song, B.; Sung, P. Functional interactions among yeast Rad51 recombinase, Rad52 mediator, and replication protein A in DNA strand exchange. J. Biol. Chem. 2000, 275, 15895–15904. [Google Scholar] [CrossRef] [PubMed]
- Gibb, B.; Ye, L.F.; Kwon, Y.; Niu, H.; Sung, P.; Greene, E.C. Protein dynamics during presynaptic-complex assembly on individual single-stranded DNA molecules. Nat. Struct. Mol. Biol. 2014, 21, 893–900. [Google Scholar] [CrossRef] [PubMed]
- Hays, S.L.; Firmenich, A.A.; Massey, P.; Banerjee, R.; Berg, P. Studies of the interaction between Rad52 protein and the yeast single-stranded DNA binding protein rpa. Mol. Cell. Biol. 1998, 18, 4400–4406. [Google Scholar] [CrossRef] [PubMed]
- Milne, G.T.; Weaver, D.T. Dominant negative alleles of RAD52 reveal a DNA repair/recombination complex including RAD51 and RAD52. Genes Dev. 1993, 7, 1755–1765. [Google Scholar] [CrossRef] [PubMed]
- Shinohara, A.; Ogawa, H.; Ogawa, T. Rad51 protein involved in repair and recombination in s. Cerevisiae is a reca-like protein. Cell 1992, 69, 457–470. [Google Scholar] [CrossRef]
- Shen, Z.; Cloud, K.G.; Chen, D.J.; Park, M.S. Specific interactions between the human RAD51 and RAD52 proteins. J. Biol. Chem. 1996, 271, 148–152. [Google Scholar] [PubMed]
- Park, M.S.; Ludwig, D.L.; Stigger, E.; Lee, S.H. Physical interaction between human RAD52 and RPA is required for homologous recombination in mammalian cells. J. Biol. Chem. 1996, 271, 18996–19000. [Google Scholar] [CrossRef] [PubMed]
- Davies, A.A.; Masson, J.Y.; McIlwraith, M.J.; Stasiak, A.Z.; Stasiak, A.; Venkitaraman, A.R.; West, S.C. Role of BRCA2 in control of the RAD51 recombination and DNA repair protein. Mol. Cell 2001, 7, 273–282. [Google Scholar] [CrossRef]
- Kagawa, W.; Kurumizaka, H.; Ikawa, S.; Yokoyama, S.; Shibata, T. Homologous pairing promoted by the human RAD52 protein. J. Biol. Chem. 2001, 276, 35201–35208. [Google Scholar] [CrossRef] [PubMed]
- Iyer, L.M.; Koonin, E.V.; Aravind, L. Classification and evolutionary history of the single-strand annealing proteins, Rect, Redbeta, ERF and RAD52. BMC Genomics 2002, 3, 8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Noirot, P.; Gupta, R.C.; Radding, C.M.; Kolodner, R.D. Hallmarks of homology recognition by RecA-like recombinases are exhibited by the unrelated Escherichia coli RecT protein. EMBO J. 2003, 22, 324–334. [Google Scholar] [CrossRef] [PubMed]
- Pezza, R.J.; Voloshin, O.N.; Volodin, A.A.; Boateng, K.A.; Bellani, M.A.; Mazin, A.V.; Camerini-Otero, R.D. The dual role of HOP2 in mammalian meiotic homologous recombination. Nucleic Acids Res. 2014, 42, 2346–2357. [Google Scholar] [CrossRef] [PubMed]
- Stasiak, A.Z.; Larquet, E.; Stasiak, A.; Muller, S.; Engel, A.; Van Dyck, E.; West, S.C.; Egelman, E.H. The human RAD52 protein exists as a heptameric ring. Curr. Biol. 2000, 10, 337–340. [Google Scholar] [CrossRef]
- Mazin, A.V.; Kowalczykowski, S.C. The function of the secondary DNA-binding site of RecA protein during DNA strand exchange. EMBO J. 1998, 17, 1161–1168. [Google Scholar] [CrossRef] [PubMed]
- Van Dyck, E.; Hajibagheri, N.M.; Stasiak, A.; West, S.C. Visualisation of human RAD52 protein and its complexes with hRAD51 and DNA. J. Mol. Biol. 1998, 284, 1027–1038. [Google Scholar] [CrossRef] [PubMed]
- Chow, S.A.; Radding, C.M. Ionic inhibition of formation of reca nucleoprotein networks blocks homologous pairing. Proc. Natl. Acad. Sci. USA 1985, 82, 5646–5650. [Google Scholar] [CrossRef] [PubMed]
- Tsang, S.S.; Chow, S.A.; Radding, C.M. Networks of DNA and RecA protein are intermediates in homologous pairing. Biochemistry 1985, 24, 3226–3232. [Google Scholar] [CrossRef] [PubMed]
- Kagawa, W.; Kurumizaka, H.; Ishitani, R.; Fukai, S.; Nureki, O.; Shibata, T.; Yokoyama, S. Crystal structure of the homologous-pairing domain from the human RAD52 recombinase in the undecameric form. Mol. Cell 2002, 10, 359–371. [Google Scholar] [CrossRef]
- Arai, N.; Kagawa, W.; Saito, K.; Shingu, Y.; Mikawa, T.; Kurumizaka, H.; Shibata, T. Vital roles of the second DNA-binding site of Rad52 protein in yeast homologous recombination. J. Biol. Chem. 2011, 286, 17607–17617. [Google Scholar] [CrossRef] [PubMed]
- Lloyd, J.A.; McGrew, D.A.; Knight, K.L. Identification of residues important for DNA binding in the full-length human RAD52 protein. J. Mol. Biol. 2005, 345, 239–249. [Google Scholar] [CrossRef] [PubMed]
- Adzuma, K.; Ogawa, T.; Ogawa, H. Primary structure of the RAD52 gene in saccharomyces cerevisiae. Mol. Cell. Biol. 1984, 4, 2735–2744. [Google Scholar] [CrossRef] [PubMed]
- Bezzubova, O.Y.; Schmidt, H.; Ostermann, K.; Heyer, W.D.; Buerstedde, J.M. Identification of a chicken RAD52 homologue suggests conservation of the RAD52 recombination pathway throughout the evolution of higher eukaryotes. Nucleic Acids Res. 1993, 21, 5945–5949. [Google Scholar] [CrossRef] [PubMed]
- Shen, Z.; Denison, K.; Lobb, R.; Gatewood, J.M.; Chen, D.J. The human and mouse homologs of the yeast Rad52 gene: cDNA cloning, sequence analysis, assignment to human chromosome 12p12.2-p13, and mRNA expression in mouse tissues. Genomics 1995, 25, 199–206. [Google Scholar] [CrossRef]
- Takahashi, N.; Dawid, I.B. Characterization of zebrafish Rad52 and replication protein A for oligonucleotide-mediated mutagenesis. Nucleic Acids Res. 2005, 33, e120. [Google Scholar] [CrossRef] [PubMed]
- Lloyd, J.A.; Forget, A.L.; Knight, K.L. Correlation of biochemical properties with the oligomeric state of human RAD52 protein. J. Biol. Chem. 2002, 277, 46172–46178. [Google Scholar] [CrossRef] [PubMed]
- Di Tommaso, P.; Moretti, S.; Xenarios, I.; Orobitg, M.; Montanyola, A.; Chang, J.M.; Taly, J.F.; Notredame, C. T-coffee: A web server for the multiple sequence alignment of protein and RNA sequences using structural information and homology extension. Nucleic Acids Res. 2011, 39, W13–W17. [Google Scholar] [CrossRef] [PubMed]
- Navadgi, V.M.; Dutta, A.; Rao, B.J. Human RAD52 facilitates a three-stranded pairing that follows no strand exchange: A novel pairing function of the protein. Biochemistry 2003, 42, 15237–15251. [Google Scholar] [CrossRef] [PubMed]
- Mazina, O.M.; Mazin, A.V.; Drexel University College of Medicine, Philadelphia, PA, USA. Unpublished work. 2016.
- Ranatunga, W.; Jackson, D.; Lloyd, J.A.; Forget, A.L.; Knight, K.L.; Borgstahl, G.E. Human RAD52 exhibits two modes of self-association. J. Biol. Chem. 2001, 276, 15876–15880. [Google Scholar] [CrossRef] [PubMed]
- Seong, C.; Sehorn, M.G.; Plate, I.; Shi, I.; Song, B.; Chi, P.; Mortensen, U.; Sung, P.; Krejci, L. Molecular anatomy of the recombination mediator function of saccharomyces cerevisiae Rad52. J. Biol. Chem. 2008, 283, 12166–12174. [Google Scholar] [CrossRef] [PubMed]
- Parsons, C.A.; Baumann, P.; Van Dyck, E.; West, S.C. Precise binding of single-stranded DNA termini by human RAD52 protein. EMBO J. 2000, 19, 4175–4181. [Google Scholar] [CrossRef] [PubMed]
- Van Dyck, E.; Stasiak, A.Z.; Stasiak, A.; West, S.C. Binding of double-strand breaks in DNA by human RAD52 protein. Nature 1999, 398, 728–731. [Google Scholar] [CrossRef] [PubMed]
- Boundy-Mills, K.L.; Livingston, D.M. A saccharomyces cerevisiae RAD52 allele expressing a C-terminal truncation protein: Activities and intragenic complementation of missense mutations. Genetics 1993, 133, 39–49. [Google Scholar] [PubMed]
- Krejci, L.; Song, B.; Bussen, W.; Rothstein, R.; Mortensen, U.H.; Sung, P. Interaction with Rad51 is indispensable for recombination mediator function of Rad52. J. Biol. Chem. 2002, 277, 40132–40141. [Google Scholar] [CrossRef] [PubMed]
- Ciccia, A.; Bredemeyer, A.L.; Sowa, M.E.; Terret, M.E.; Jallepalli, P.V.; Harper, J.W.; Elledge, S.J. The SIOD disorder protein SMARCAL1 is an RPA-interacting protein involved in replication fork restart. Genes Dev. 2009, 23, 2415–2425. [Google Scholar] [CrossRef] [PubMed]
- Bochkareva, E.; Korolev, S.; Lees-Miller, S.P.; Bochkarev, A. Structure of the RPA trimerization core and its role in the multistep DNA-binding mechanism of rpa. EMBO J. 2002, 21, 1855–1863. [Google Scholar] [CrossRef] [PubMed]
- Jackson, D.; Dhar, K.; Wahl, J.K.; Wold, M.S.; Borgstahl, G.E. Analysis of the human replication protein a:Rad52 complex: Evidence for crosstalk between RPA32, RPA70, RAD52 and DNA. J. Mol. Biol. 2002, 321, 133–148. [Google Scholar] [CrossRef]
- Ciccia, A.; Constantinou, A.; West, S.C. Identification and characterization of the human Mus81-Eme1 endonuclease. J. Biol. Chem. 2003, 278, 25172–25178. [Google Scholar] [CrossRef] [PubMed]
- Osman, F.; Whitby, M.C. Exploring the roles of Mus81-Eme1/mms4 at perturbed replication forks. DNA Repair 2007, 6, 1004–1017. [Google Scholar] [CrossRef] [PubMed]
- Murfuni, I.; Basile, G.; Subramanyam, S.; Malacaria, E.; Bignami, M.; Spies, M.; Franchitto, A.; Pichierri, P. Survival of the replication checkpoint deficient cells requires Mus81-RAD52 function. PLoS Genet 2013, 9, e1003910. [Google Scholar] [CrossRef] [PubMed]
- Vejrup-Hansen, R.; Mizuno, K.; Miyabe, I.; Fleck, O.; Holmberg, C.; Murray, J.M.; Carr, A.M.; Nielsen, O. Schizosaccharomyces pombe mms1 channels repair of perturbed replication into Rhp51 independent homologous recombination. DNA Repair 2011, 10, 283–295. [Google Scholar] [CrossRef] [PubMed]
- Doe, C.L.; Osman, F.; Dixon, J.; Whitby, M.C. DNA repair by a Rad22-Mus81-dependent pathway that is independent of Rhp51. Nucleic Acids Res. 2004, 32, 5570–5581. [Google Scholar] [CrossRef] [PubMed]
- Koike, M.; Yutoku, Y.; Koike, A. The C-terminal region of RAD52 is essential for RAD52 nuclear and nucleolar localization, and accumulation at DNA damage sites immediately after irradiation. Biochem. Biophys. Res. Commun. 2013, 435, 260–266. [Google Scholar] [CrossRef] [PubMed]
- Kito, K.; Wada, H.; Yeh, E.T.; Kamitani, T. Identification of novel isoforms of human RAD52. Biochim. Biophys. Acta. 1999, 1489, 303–314. [Google Scholar] [CrossRef]
- Thorpe, P.H.; Marrero, V.A.; Savitzky, M.H.; Sunjevaric, I.; Freeman, T.C.; Rothstein, R. Cells expressing murine RAD52 splice variants favor sister chromatid repair. Mol. Cell. Biol. 2006, 26, 3752–3763. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Sugiyama, T.; Kowalczykowski, S.C. DNA annealing mediated by Rad52 and Rad59 proteins. J. Biol. Chem. 2006, 281, 15441–15449. [Google Scholar] [CrossRef] [PubMed]
- Feng, Q.; During, L.; de Mayolo, A.A.; Lettier, G.; Lisby, M.; Erdeniz, N.; Mortensen, U.H.; Rothstein, R. Rad52 and Rad59 exhibit both overlapping and distinct functions. DNA Repair 2007, 6, 27–37. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Davis, A.P.; Symington, L.S. The yeast recombinational repair protein Rad59 interacts with Rad52 and stimulates single-strand annealing. Genetics 2001, 159, 515–525. [Google Scholar] [PubMed]
- Petukhova, G.; Stratton, S.A.; Sung, P. Single strand DNA binding and annealing activities in the yeast recombination factor Rad59. J. Biol. Chem. 1999, 274, 33839–33842. [Google Scholar] [CrossRef] [PubMed]
- Barlow, J.H.; Rothstein, R. Timing is everything: Cell cycle control of Rad52. Cell Div. 2010, 5, 7. [Google Scholar] [CrossRef] [PubMed]
- Karanam, K.; Kafri, R.; Loewer, A.; Lahav, G. Quantitative live cell imaging reveals a gradual shift between DNA repair mechanisms and a maximal use of HR in mid Sphase. Mol. Cell 2012, 47, 320–329. [Google Scholar] [CrossRef] [PubMed]
- Barlow, J.H.; Rothstein, R. Rad52 recruitment is DNA replication independent and regulated by Cdc28 and the Mec1 kinase. EMBO J. 2009, 28, 1121–1130. [Google Scholar] [CrossRef] [PubMed]
- Antunez de Mayolo, A.; Lisby, M.; Erdeniz, N.; Thybo, T.; Mortensen, U.H.; Rothstein, R. Multiple start codons and phosphorylation result in discrete RAD52 protein species. Nucleic Acids Res. 2006, 34, 2587–2597. [Google Scholar] [CrossRef] [PubMed]
- Bellini, A.; Girard, P.M.; Lambert, S.; Tessier, L.; Sage, E.; Francesconi, S. Stress activated protein kinase pathway modulates homologous recombination in fission yeast. PLoS ONE 2012, 7, e47987. [Google Scholar] [CrossRef] [PubMed]
- Bellini, A.; Girard, P.M.; Tessier, L.; Sage, E.; Francesconi, S. Fission yeast Rad52 phosphorylation restrains error prone recombination pathways. PLoS ONE 2014, 9, e95788. [Google Scholar] [CrossRef] [PubMed]
- Kitao, H.; Yuan, Z.M. Regulation of ionizing radiation-induced RAD52 nuclear foci formation by c-ABL-mediated phosphorylation. J. Biol. Chem. 2002, 277, 48944–48948. [Google Scholar] [CrossRef] [PubMed]
- Honda, M.; Okuno, Y.; Yoo, J.; Ha, T.; Spies, M. Tyrosine phosphorylation enhances RAD52-mediated annealing by modulating its DNA binding. EMBO J. 2011, 30, 3368–3382. [Google Scholar] [CrossRef] [PubMed]
- Sacher, M.; Pfander, B.; Hoege, C.; Jentsch, S. Control of Rad52 recombination activity by double-strand break-induced sumo modification. Nat. Cell Biol. 2006, 8, 1284–1290. [Google Scholar] [CrossRef] [PubMed]
- Ohuchi, T.; Seki, M.; Branzei, D.; Maeda, D.; Ui, A.; Ogiwara, H.; Tada, S.; Enomoto, T. Rad52 sumoylation and its involvement in the efficient induction of homologous recombination. DNA Repair 2008, 7, 879–889. [Google Scholar] [CrossRef] [PubMed]
- Bergink, S.; Ammon, T.; Kern, M.; Schermelleh, L.; Leonhardt, H.; Jentsch, S. Role of Cdc48/p97 as a SUMO-targeted segregase curbing Rad51-Rad52 interaction. Nat. Cell Biol. 2013, 15, 526–532. [Google Scholar] [CrossRef] [PubMed]
- Torres-Rosell, J.; Sunjevaric, I.; De Piccoli, G.; Sacher, M.; Eckert-Boulet, N.; Reid, R.; Jentsch, S.; Rothstein, R.; Aragon, L.; Lisby, M. The Smc5-smc6 complex and SUMO modification of Rad52 regulates recombinational repair at the ribosomal gene locus. Nat. Cell Biol. 2007, 9, 923–931. [Google Scholar] [CrossRef] [PubMed]
- Saito, K.; Kagawa, W.; Suzuki, T.; Suzuki, H.; Yokoyama, S.; Saitoh, H.; Tashiro, S.; Dohmae, N.; Kurumizaka, H. The putative nuclear localization signal of the human RAD52 protein is a potential sumoylation site. J. Biochem. 2010, 147, 833–842. [Google Scholar] [CrossRef] [PubMed]
- Choi, B.H.; Chen, Y.; Dai, W. Chromatin PTEN is involved in DNA damage response partly through regulating Rad52 sumoylation. Cell Cycle 2013, 12, 3442–3447. [Google Scholar] [CrossRef] [PubMed]
- Storici, F.; Bebenek, K.; Kunkel, T.A.; Gordenin, D.A.; Resnick, M.A. RNA-templated DNA repair. Nature 2007, 447, 338–341. [Google Scholar] [CrossRef] [PubMed]
- Shen, Y.; Storici, F. Detection of RNA-templated double-strand break repair in yeast. Methods Mol. Biol. 2011, 745, 193–204. [Google Scholar] [PubMed]
- Wei, L.; Nakajima, S.; Bohm, S.; Bernstein, K.A.; Shen, Z.; Tsang, M.; Levine, A.S.; Lan, L. DNA damage during the G0/G1 phase triggers RNA-templated, Cockayne syndrome B-dependent homologous recombination. Proc. Natl. Acad. Sci. USA 2015, 112, E3495–E3504. [Google Scholar] [CrossRef] [PubMed]
- Gunn, A.; Stark, J.M. I-SceI-based assays to examine distinct repair outcomes of mammalian chromosomal double strand breaks. Methods Mol. Biol. 2012, 920, 379–391. [Google Scholar] [PubMed]
- Hengel, S.R.; Malacaria, E.; Folly da Silva Constantino, L.; Bain, F.E.; Diaz, A.; Koch, B.G.; Yu, L.; Wu, M.; Pichierri, P.; Spies, M.A.; et al. Small-molecule inhibitors identify the RAD52-ssdna interaction as critical for recovery from replication stress and for survival of BRCA2 deficient cells. Elife 2016, 5, e14740. [Google Scholar] [CrossRef] [PubMed]



© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hanamshet, K.; Mazina, O.M.; Mazin, A.V. Reappearance from Obscurity: Mammalian Rad52 in Homologous Recombination. Genes 2016, 7, 63. https://doi.org/10.3390/genes7090063
Hanamshet K, Mazina OM, Mazin AV. Reappearance from Obscurity: Mammalian Rad52 in Homologous Recombination. Genes. 2016; 7(9):63. https://doi.org/10.3390/genes7090063
Chicago/Turabian StyleHanamshet, Kritika, Olga M. Mazina, and Alexander V. Mazin. 2016. "Reappearance from Obscurity: Mammalian Rad52 in Homologous Recombination" Genes 7, no. 9: 63. https://doi.org/10.3390/genes7090063