
Chromosomal Evolution in Lower Vertebrates: Sex Chromosomes in Neotropical Fishes
by
Marcelo de Bello Cioffi
1,*,†, 
Cassia Fernanda Yano
1,†,
Alexandr Sember
2 and
Luiz Antônio Carlos Bertollo
1 1
Departamento de Genética e Evolução, Universidade Federal de São Carlos, São Carlos, SP CEP 13565-905, Brazil
2
Laboratory of Fish Genetics, Institute of Animal Physiology and Genetics, Czech Academy of Sciences, Rumburská 89, Liběchov 277 21, Czech Republic
*
Author to whom correspondence should be addressed.
†
These authors have contributed equally to this work.
Genes 2017, 8(10), 258; https://doi.org/10.3390/genes8100258
Submission received: 25 August 2017
/
Revised: 27 September 2017
/
Accepted: 29 September 2017
/
Published: 5 October 2017
(This article belongs to the Special Issue Chromosomal Evolution)
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Fishes exhibit the greatest diversity of species among vertebrates, offering a number of relevant models for genetic and evolutionary studies. The investigation of sex chromosome differentiation is a very active and striking research area of fish cytogenetics, as fishes represent one of the most vital model groups. Neotropical fish species show an amazing variety of sex chromosome systems, where different stages of differentiation can be found, ranging from homomorphic to highly differentiated sex chromosomes. Here, we draw attention on the impact of recent developments in molecular cytogenetic analyses that helped to elucidate many unknown questions about fish sex chromosome evolution, using excellent characiform models occurring in the Neotropical region, namely the Erythrinidae family and the Triportheus genus. While in Erythrinidae distinct XY and/or multiple XY-derived sex chromosome systems have independently evolved at least four different times, representatives of Triportheus show an opposite scenario, i.e., highly conserved ZZ/ZW system with a monophyletic origin. In both cases, recent molecular approaches, such as mapping of repetitive DNA classes, comparative genomic hybridization (CGH), and whole chromosome painting (WCP), allowed us to unmask several new features linked to the molecular composition and differentiation processes of sex chromosomes in fishes.
1. Introduction
For cytologically distinguishable sex chromosomes to emerge, one of the homologues needs to acquire at least two linked alleles that are advantageous to one sex only and possibly detrimental to the other [1,2]. The next required step is the suppression of recombination between the homologues of the proto-sex chromosomes via chromosomal rearrangements (most commonly inversions or translocations) and/or accumulation of repetitive DNA sequences. In fact, this is a precondition for further gradual genetic and morphological differentiation of the sex pair, through the spreading of the non-recombining region facilitated by additional heterochromatin/repetitive DNA accumulation, genetic degeneration, and, in the long run, size modification of the sex-specific chromosome [1,2,3,4,5,6].
Unlike well-established sex chromosome systems in mammals, snakes, and birds [5,7], fish sex chromosomes often do not progress through the entire set of the aforementioned evolutionary steps [7,8,9,10]. In contrast, many of them are rather of cryptic nature, reflecting perhaps their relatively young evolutionary age [7,11]. Such characteristics often allow fish sex chromosomes to escape from detection under conventional cytogenetic analysis. Consequently, only about 10% of fish species surveyed so far display distinguishable sex chromosomes, with about half of them belonging to the Neotropical region [9,10,12,13]. Such ichthyofauna with approximately 6000 freshwater fish species, represent the world’s richest one [14].
Remarkably, even considering the small number of known cases, several different sex chromosome systems have been described for diverse fish taxa [13,15,16], demonstrating their high evolutionary diversity and plasticity. Overall, at least nine distinct sex chromosome systems have already been recognized among fishes, including the primary ♀XX/♂XY, ♂ZZ/♀ZW and the derived ♀XX/♂X0, ♂ZZ/♀Z0 systems, the standard ♀X1X1X2X2/♂X1X2Y, ♀XX/♂XY1Y2, ♂ZZ/♀ZW1W2 multiple ones [12,17,18], along with some unique types such as ♂Z1Z1Z2Z2/♀Z1Z2W1W2 and ♀X1X1X2X2/♂X1Y1X2Y2 [16,19,20]. The same extent of variability amounts also for the Neotropical ichthyofauna [17,18,21].
Sex chromosomes often emerged independently and at various times in many fish lineages, following distinct patterns of differentiation even in closely related species [7,10,11,12,22,23,24,25]. In fact, only few exceptions are known from this general view, as in the whole genus Triportheus [26,27] and some Leporinus [28,29], and Characidium species [30,31]. Similarly to what occurs among amphibians and reptiles [7,32], sex chromosome turnover is also a common trait in fishes, which may be achieved by (i) the recruitment of a new sex-determining gene on an autosome, (ii) the transposition of a sex-determining locus to an autosome, or (iii) fusion events among autosomes and cryptically differentiated sex chromosomes, giving rise to the so-called neo-sex chromosome systems [7,11,15,32,33]. Meiotic drive and sex-specific selection pressures are also probably evolutionary forces facilitating such turnover [15,32,34,35,36]. In addition, the high lability of fish sex chromosome systems and, in a broader scale, the sex determination mechanisms themselves, may be also related to the high plasticity of teleost genomes after whole-genome duplications [11] or even to the need for adaptation to varying environmental conditions (e.g., to counterbalance sex ratio distortions after the colonization of a new biotope) [37]. Finally, the stable conditions for biochemical reactions in warm-blooded organisms might favor their conservative sex chromosome constitution, unlike the situation in cold-blooded animals, and thus also in fishes [37].
The above-mentioned features qualify some fish species as excellent models to examine sex chromosome evolution, calling for more in-depth cytogenetic and genomic studies. While the identity, structure, and expression of particular genes are still rather poorly explored in fish sex determination and differentiation [35,37,38,39], relevant advances have been achieved concerning the molecular composition and differentiation process of sex chromosomes employing molecular cytogenetic procedures, such as chromosomal mapping of repetitive DNAs, comparative genomic hybridization (CGH), and whole chromosome painting (WCP) (e.g., [25,40,41,42]).
In this sense, our present review emphasizes two alternative models to investigate the evolution of sex chromosomes among Neotropical fishes, exemplified by species from the Erythrinidae family and the Triportheus genus from the Triportheidae family. These fish groups have been our investigation systems for years, for which a considerable amount of conventional and molecular cytogenetic data show differential pathways on the differentiation process of the sex chromosomes.
2. The Erythrinidae Family: A Broad Scenario on Fish Sex Chromosomes Evolution
Erythrinidae is a small characiform family, widely distributed in the Neotropical region, with only three genera: Hoplias (Gill, 1903), Hoplerythrinus (Gill, 1895), and Erythrinus (Scopoli, 1977), providing a rare opportunity to gain insights into the evolutionary forces that drive the origin of nascent sex chromosomes, the evolution of the sex pair and speciation processes. In fact, the lack of heteromorphic sex chromosomes, as well as the occurrence of sex chromosome systems with distinct evolutionary stages of differentiation, can be found among different populations of the wolf fish Hoplias malabaricus and/or the red wolf fish Erythrinus erythrinus [24,43]. Noteworthy, these fishes are also unique in the way that males are always the heterogametic sex—an uncommon trait among fishes (reviewed in [9]).
Based on a multitude of chromosomal and molecular data, especially within the last 20 years, it has become increasingly apparent that we are dealing with a group of species instead of a single taxonomic unit, both for H. malabaricus and E. erythrinus [24,44]. From the cytogenetic standpoint, seven distinct karyotype forms or karyomorphs (A–G) in H. malabaricus and four (A–D) in E. erythrinus have already been identified, respectively, based on major differences in diploid chromosome numbers (2n), chromosome morphology, and sex chromosomes [44]. Studies employing WCP, CGH, and repetitive DNA distribution patterns have highlighted the differentiation plasticity of the erythrinid sex chromosome systems [25,45,46,47]. More specifically, it was demonstrated that sex chromosomes could emerge via independent pathways, following distinct patterns of differentiation even within the same type of system and among closely related karyomorphs (Figure 1). These features suggest that sex chromosome turnover might play an important role in the speciation process of these fishes (Figure 2).
Within H. malabaricus, karyomorph B (2n = 42, both sexes) exhibits the most differentiated ♀XX/♂XY sex chromosome system, where the subtelocentric X chromosome is clearly distinguished from the small-sized metacentric Y by a conspicuous heterochromatic block on its long (q) arms [48,49]. Several complementary studies performing mapping of repetitive DNAs, such as 18S rDNA and various microsatellite motifs, reported the unusual enrichment of the X-restricted heterochromatin by such sequences [49,50,51]. These general data, in addition to WCP, indicate that the XY chromosomes of karyomorph B are likely derived from the chromosome pair No. 21 found in karyomorph A [25,52]. Noteworthy, a male-specific region is distally confined to the q arms of Y chromosome, corresponding to a C-positive heterochromatic block occupying the same chromosomal region [47]. Indeed, the location of this region stands out given that a large nucleolar organizer region (NOR) site in the corresponding region of the X chromosome is responsible for the big size difference between both sex chromosomes [48].
Karyomorph C (2n = 40, both sexes), is characterized by morphologically undifferentiated XX/XY sex chromosome system evidenced by a small, but considerable, heterochromatin accumulation on the exclusively proximal region of the X chromosome [53]. CGH results documented the presence of distinct male-specific sequences in this likely nascent sex-determining region [47]). As reported for karyomorph B, the preferential accumulation of repetitive DNAs on the X chromosome is also a particular feature for the nascent XY sex system of this karyomorph. Karyomorph D has 2n = 40 chromosomes in females and 2n = 39 in males, resulting from multiple sex chromosomes of X1X1X2X2/X1X2Y type [54]. Stabilized sex trivalents are found during male meiosis, as well as asynapsis in a presumed sequence divergence region [55], thus pointing to a putative sex-specific region. Rosa et al. [56] reported marked alterations in the location of constitutive heterochromatin and 18S rDNA sites on the Y chromosome, indicating that pericentric inversions probably have also taken place in the early process of sex-specific chromosome differentiation. However, CGH data did not reveal any unique Y-specific region [47]. Therefore, it is possible that yet insufficient sequence divergence within the sex-specific region prevents its detection by CGH, similarly to that recently reported in invertebrates [57].
Karyomorph F (2n = 40, for both sexes) also exhibits a nascent sex chromosome system of XX/XY type [45]. The male-specific content was shown as a prominent interstitial heterochromatic block on the short (p) arms of the large metacentric Y chromosome, also accumulated with several microsatellite motifs and retrotransposons (RTEs), that are absent on the X chromosome [45]. By contrast, karyomorph G (2n = 40 in females/ 41 in males) presents an XX/XY1Y2 neo-sex chromosomes, where the unusual acrocentric Y1 chromosome carries the male-specific region that is also enriched by several types of repetitive DNAs [47].
Similarly to H. malabaricus, the species E. erythrinus exhibits extensive karyotype diversity among populations, with four karyomorphs (A–D) currently recognized [58]. While karyomorph A is the only one lacking differentiated sex chromosomes, karyomorphs B, C, and D share a multiple X1X1X2X2/X1X2Y sex chromosome system, though they still differ in 2n and karyotype composition. Chromosomal mapping of repetitive DNA by means of FISH (fluorescence in situ hybridization) demonstrated the nature of chromosomal rearrangements and genomic modifications leading to establishment of this X1X2Y sex system. More specifically, the neo-Y owns its origin to a centric fusion event. In support of this, FISH with telomeric probe highlighted the presence of interstitial telomeric sequences (ITSs) in the centromeric region of the neo-Y chromosome, the only metacentric element in the karyotype [59].
The repeated occurrence of XY and XY-derived sex chromosome systems in Erythrinidae fishes suggested, at first, a close relationship or even a common origin among them. However, chromosomal painting experiments pointed to at least four independent origins and differentiation processes of the sex chromosomes (Figure 2). A set of whole chromosome probes, including (i) the X chromosome of H. malabaricus karyomorph B (HMB-X), (ii) the X1 chromosome of H. malabaricus karyomorph D (HMD-X1), (iii) the Y chromosome of H. malabaricus karyomorph F (HMF-Y,) and (iv) the Y chromosome of E. erythrinus karyomorph D (ERY-Y), were used in WCP experiments, providing the pathways of the sex chromosomes differentiation and important hints about interspecific and inter-karyomorph relationships (Figure 3).
The lack of recognizable signals on the H. malabaricus karyomorphs B, C, and F sex chromosomes after WCP experiments showed that their XY chromosomes have evolved independently from different autosomal pairs [45,52]. The same approach also identified an autosomal pair from karyomorph A as homeologous to the well-differentiated XY sex chromosomes of karyomorph B and showed that karyomorphs C and D display a close evolutionary proximity. Indeed, in the latter case, the neo-Y chromosome of karyomorph D was proven to arise from a tandem fusion between the nascent Y chromosome and one homologue of the autosomal pair no. 20 present in karyomorph C [53].
Karyomorphs E, F, and G are evidently closely related, thus forming a separate evolutionary clade [43]. The origin of the sex chromosomes in karyomorphs F and G likely resulted from tandem fusion events between acrocentric and submetacentric chromosomes, as found in karyomorph E. In fact, this proposition was strengthened by recent results indicating that in karyomorph F, such rearrangements are found in a homozygous condition, i.e., the large-sized metacentric XY chromosomes, while in karyomorph G, just one homologue per each chromosomal pair had fused and thus giving rise to the unpaired large-sized neo-X chromosome, in addition to the remaining unfused a and sm chromosomes designated as Y1 and Y2, respectively [46]. Based on these findings, we proposed the parallel evolution of sex chromosome systems of karyomorphs F and G from an E-like karyomorph through different evolutionary scenarios. Mapping of the HMF-Y probe to chromosome complements of other H. malabaricus karyomorphs confirmed that sex chromosomes of karyomorph F evolved from different autosome pairs compared to the sex chromosomes of karyomorphs B, C and D [46].
In contrast to H. malabaricus, cross-FISH experiments pointed to a common origin of the multiple X1X2Y sex chromosomes present in E. erythrinus karyomorphs, where the centric fusion that gave rise to the neo-Y chromosome probably has occurred before the karyomorphs divergence [61]. However, the same X1X2Y sex system present in H. malabaricus karyomorph D has evolved independently, in which different autosomal pairs were converted to sex chromosomes.
In summary, the representatives of Erythrinidae family serve as a very useful model for studying the emergence and differentiation of the sex chromosomes among fishes. In fact, it highlights a variety of specific scenarios, ranging from emergent to well differentiated sex systems, with a common or independent origins among its clades.
3. The Triportheus Genus: A Particular Pathway on Fish Sex Chromosomes Evolution
The Triportheus genus (Cope, 1872) comprises 19 extant species popularly known as freshwater sardines [14]. They belong to the monophyletic Triportheidae family, which is also composed of four other genera: Lignobrycon, Engraulisoma, Clupeacharax, and Agoniates [62,63]. As to the genus Triportheus, its origin dates back to 26.2 ± 6.5 Myr, where Triportheus auritus phylogenetically represents the oldest lineage (20.7 ± 6.5 Myr; [63]).
Similarly to H. malabaricus and E. erythrinus, this characiform taxon also exhibits a wide distribution throughout South American hydrographic basins [62,64,65]. However, unlike such groups, Triportheus exhibits a striking difference of the evolutionary trends dealing with the sex chromosomes. All Tiportheus species studied so far share the same sex chromosome system (ZZ/ZW), with evident signs of monophyletic origin (Figure 4). This feature is unique among fishes surveyed to date and it sharply contrasts with the overall picture provided by fish sex chromosome evolution, in which repeated and independent origins, along with frequent turnovers, represent the most common situation documented so far [7,11].
To date, 11 out of the 19 Triportheus species had been cytogenetically analyzed and all of them share an identical karyotype macrostructure, with 2n = 52 both in males and females [26,27,42,66,67,68,69,70,71,72,73,74,75]. They also share a ZZ/ZW sex chromosome system, with the common metacentric Z chromosome, the largest element of the karyotype. By contrast, the smaller female W chromosome shows interspecific variability in size, morphology, and the content of C-positive heterochromatin [26,67]. The W and Z chromosomes show almost equal size in the most phylogenetically basal species, T. auritus, while in the more derived ones, the W chromosome displays a size reduction [42]. Nonetheless, there is no direct correlation between the phylogenetic position and the degree of W degeneration [26,42], nor the most derived taxon in the available recent phylogeny [62] bears the smallest W chromosome. (Figure 5).
Besides the reduction in size, the W chromosome of Triportheus has undergone similarly progressive evolutionary dynamics as the Y and W chromosomes of higher vertebrates, with a large accumulation of heterochromatin and repetitive DNA sequences. In this sense, such chromosome is typically almost entirely heterochromatic, while the Z chromosome shows C-positive blocks restricted only to the centromeric and telomeric regions [27,67,68]. In addition, all Triportheus species possess characteristically a W-specific 18S rDNA cluster located in the terminal region of the q arms (Wq) [68,72,73,74]. In fact, it has been proposed that the amplification of the rDNA sites on the W chromosome had led to the recombination reduction between the ZW chromosomes [69,75]. Similarly, Reed & Phillips [77] have already suggested a functional importance of NOR on the putative sex chromosome of the Arctic char as it might be able to decrease considerably the recombination rate in the adjacent region (via interference) and thus to facilitate the formation of a possible sex-specific locus there. The location of rDNA sequences within the sex-specific region was already confirmed by FISH and CGH in T. auritus [42] and in studies performed in turtles [78,79], while it was alternatively found located adjacent to such region, e.g., in frogs [80]. The maintenance of the 18S rDNA cluster on the W chromosome throughout the evolutionary history of all Triportheus species reflects the probable role of these sequences in the evolution of the ZZ/ZW sex chromosome system of this genus [42].
Additionally, the differentiation of the W chromosome in Triportheus has been also associated with other repetitive DNA sequences. The W chromosome of Triportheus albus also possess another multigene family, the U2 small nuclear RNA (snRNA). Until now, only a few cases are known to occur in fish sex chromosomes (as reviewed in [75]). Microsatellites have also been tightly linked with the W differentiation of Triportheus due to their intrinsic ability to rapidly expand in the non-recombinant chromosomal regions [26,74,81,82].
The WCP using both W- and Z-derived probes confirmed the hypothesis about the origin of the ZW sex chromosome system within the Triportheus genus. In fact, the Z-probe painted the whole length of the Z chromosome equally in all species, showing its conserved nature in both size and genetic content [42,71] (Figure 6). In turn, the W chromosome of all species was strongly painted using the W-probe from the phylogenetically basal species T. auritus, demonstrating that the ZW system in Triportheus had a common origin within the genus [42]. Moreover, recent findings showing a morphologically similar ZZ/ZW sex system in Lignobrycon myersi (Triportheidae) [83], as well as the occurrence of female heterogamety in species from a sister family (Gasterpelecidae) [84], open new perspectives for investigations of the evolutionary history of the sex chromosomes among these groups. (Figure 4). Thus, the presence of a highly conserved ZZ/ZW system with a monophyletic origin place Triportheus as a unique model providing a particular scenario of the evolution of sex chromosomes among the Neotropical fishes.
4. Concluding Remarks
In the light of recent cytogenetic progresses, we aimed to illustrate here two contrasting scenarios of sex chromosome evolution in Neotropical fishes: (i) representatives of the Erythrinidae family show high variability and multiple origins of their sex chromosome systems, and (ii) representatives of the genus Triportheus highlight a case of monophyletic and stable sex chromosome system, but with different stages of degeneration the sex-specific chromosome. Nonetheless, both models share some common properties, such as repetitive DNA sequences playing a substantial role in the differentiation processes of heteromorphic sex chromosomes. In contrast, the multiple neo-sex chromosomes, arising through the structural rearrangements, do not undergo progressive heterochromatinization [9]—a feature also repeatedly documented in other fish groups [8,20,85,86] that presumably has a large impact on the stabilization of the sex multivalents during meiosis [86,87]. In this sense, it appears to be evident that the highest occurrence of XY-derived multiple sex systems reflects the early stage of differentiation of the XX/XY sex system found in most fishes examined so far. In contrast, the more rapid heterochromatinization and repetitive DNA accumulation observed in the otherwise very frequent ZZ/ZW systems would probably hinder the successful formation of stable ZZ/ZW-derived neo-sex systems. In conclusion, the two alternative models illustrated by the Erythrinidae and Triportheus species represent only a fraction of possible evolutionary scenarios within the spectacular fish sex chromosome diversity among lower vertebrates.
As illustrated above, the recruitment of WCP and its cross-species application together with CGH-based experiments represent powerful toolbox for deciphering homologies between distinct sex chromosomes on the interspecific or interpopulational level. However, what accounts for such rapid evolution of sex chromosomes among fishes is yet not fully understood. To obtain a more informed and comprehensive picture, next generation sequencing (NGS) data could be harnessed as a valuable option for studying the genetic basis of sex determination, providing opportunities to identify homologies between sex chromosomes across populations and species in a high throughput manner. Besides, searching for genome-specific satellite DNAs and their subsequent FISH mapping might add another layer of investigation and bring a more complex view about the process underlying fish sex chromosomes evolution.
Acknowledgments
This study was supported by Conselho Nacional de Desenvolvimento Científico e Tecnológico—CNPq (Proc. nos 304992/2015-1, 306896/2014-1 and 152105/2016-6), Fundação de Amparo à Pesquisa do Estado de São Paulo-FAPESP (Proc. No 2016/22196-2) and the project EXCELLENCE CZ.02.1.01/0.0/0.0/15_003/0000460 OP RDE.
Author Contributions
M.d.B.C. and C.F.Y. designed the study and drafted the manuscript. A.S. and L.A.C.B. drafted and revised the manuscript. All authors read and approved the final version of the manuscript.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Ohno, S. Sex Chromosomes and Sex-Linked Genes; Springer-Verlag: New York, NY, USA, 1967; p. 185. ISBN 978-3-642-88180-0. [Google Scholar]
- Charlesworth, D.; Charlesworth, B.; Marais, G. Steps in the evolution of heteromorphic sex chromosomes. Heredity 2005, 95, 118–128. [Google Scholar] [CrossRef] [PubMed]
- Steinemann, S.; Steinemann, M. Retroelements: Tools for sex chromosome evolution. Cytogenet. Genome Res. 2005, 110, 134–143. [Google Scholar] [CrossRef] [PubMed]
- Bachtrog, D. A dynamic view of sex chromosome evolution. Curr. Opin. Genet. Dev. 2006, 16, 578–585. [Google Scholar] [CrossRef] [PubMed]
- Graves, J.A.M. Weird animal genomes and the evolution of vertebrate sex and sex chromosomes. Annu. Rev. Genet. 2008, 42, 565–586. [Google Scholar] [CrossRef] [PubMed]
- Wright, A.E.; Dean, R.; Zimmer, F.; Mank, J.E. How to make a sex chromosome. Nat. Commun. 2016, 7. [Google Scholar] [CrossRef] [PubMed]
- Schartl, M.; Schmid, M.; Nanda, I. Dynamics of vertebrate sex chromosome evolution: From equal size to giants and dwarfs. Chromosoma 2016, 125, 553–571. [Google Scholar] [CrossRef] [PubMed]
- Almeida-Toledo, L.F.; Foresti, F. Morphologically differentiated sex chromosomes in neotropical freshwater fish. Genetica 2001, 111, 91–100. [Google Scholar] [CrossRef] [PubMed]
- Cioffi, M.B.; Camacho, J.P.M.; Bertollo, L.A.C. Repetitive DNAs and differentiation of sex chromosomes in neotropical fishes. Cytogenet. Genome Res. 2011, 132, 188–194. [Google Scholar] [CrossRef] [PubMed]
- Cioffi, M.B.; Moreira-Filho, O.; Almeida-Toledo, L.F.; Bertollo, L.A.C. The contrasting role of heterochromatin in the differentiation of sex chromosomes: An overview from Neotropical fishes. J. Fish Biol. 2012, 80, 2125–2139. [Google Scholar] [CrossRef] [PubMed]
- Mank, J.E.; Avise, J.C. Evolutionary diversity and turn-over of sex determination in teleost fishes. Sex. Dev. 2009, 3, 60–67. [Google Scholar] [CrossRef] [PubMed]
- Devlin, R.H.; Nagahama, Y. Sex determination and sex differentiation in fish: An overview of genetic, physiological, and environmental influences. Aquaculture 2002, 208, 191–364. [Google Scholar] [CrossRef]
- Oliveira, C.; Almeida-Toledo, L.F.; Foresti, F. Karyotypic evolution in Neotropical fishes. In Fish Cytogenetics; Pisano, E., Ozouf-Costaz, C., Foresti, F., Kapoor, B.G., Eds.; Science Publishers: Enfield, NH, USA, 2007; pp. 111–164. ISBN 978-1-57808-330-5. [Google Scholar]
- Froese, R.; Pauly, D. (Eds.) FishBase. World Wide Web Electronic Publication. Available online: www.fishbase.org (accessed on 2 February 2017).
- Kitano, J.; Peichel, C.L. Turnover of sex chromosomes and speciation in fishes. Environ. Biol. Fishes 2012, 94, 549–558. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, M.; Garcia, C.; Matoso, D.A.; de Jesus, I.S.; Feldberg, E. A new multiple sex chromosome system X1X1X2X2/X1Y1X2Y2 in Siluriformes: Cytogenetic characterization of Bunocephalus coracoideus (Aspredinidae). Genetica 2016, 144, 591–599. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, C.; Foresti, F.; Hilsdorf, A.W.S. Genetics of neotropical fish: From chromosomes to populations. Fish Physiol. Biochem. 2009, 35, 81–100. [Google Scholar] [CrossRef] [PubMed]
- Arai, R. Fish Karyotypes: A Check List, 1st ed.; Springer: Tokyo, Japan, 2011; p. 340. ISBN 978-4-431-53877. [Google Scholar]
- Parise-Maltempi, P.P.; Martins, C.; Oliveira, C.; Foresti, F. Identification of a new repetitive element in the sex chromosomes of Leporinus elongatus (Teleostei: Characiformes: Anostomidae): New insights into the sex chromosomes of Leporinus Cytogenet. Genome Res. 2007, 116, 218–223. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, R.R.; Feldberg, E.; dos Anjos, M.B.; Zuanon, J. Occurrence of multiple sexual chromosomes (XX/XY1Y2 and Z1Z1Z2Z2/Z1Z2W1W2) in catfishes of the genus Ancistrus (Siluriformes: Loricariidae) from the Amazon basin. Genetica 2008, 134, 243–249. [Google Scholar] [CrossRef] [PubMed]
- Araya-Jaime, C.; Mateussi, N.T.B.; Utsunomia, R.; Costa-Silva, G.J.; Oliveira, C.; Foresti, F. ZZ/Z0: The new system of sex chromosomes in Eigenmannia aff. trilineata (Teleostei: Gymnotiformes: Sternopygidae) characterized by molecular cytogenetics and DNA barcoding. Zebrafish 2017. [Google Scholar] [CrossRef] [PubMed]
- Woram, R.A.; Gharbi, K.; Sakamoto, T.; Hoyheim, B.; Holm, L.-E.; Naish, K.; McGowan, C.; Ferguson, M.M.; Phillips, R.B.; Stein, J.; et al. Comparative genome analysis of the primary sex-determining locus in salmonid fishes. Genome Res. 2003, 13, 272–280. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mank, J.E.; Promislow, D.E.L.; Avise, J.C. Evolution of alternative sex-determining mechanisms in teleost fishes. Biol. J. Linn. Soc. 2006, 87, 83–93. [Google Scholar] [CrossRef]
- Cioffi, M.B.; Molina, W.F.; Artoni, R.F.; Bertollo, L.A.C. Chromosomes as tools for discovering biodiversity—The case of Erythrinidae fish family. In Recent Trends in Cytogenetic Studies—Methodologies and Applications; Tirunilai, P., Ed.; InTech Publisher: Rijeka, Croatia, 2012; ISBN 978-953-51-0178-9. [Google Scholar]
- Cioffi, M.B.; Liehr, T.; Trifonov, V.; Molina, W.F.; Bertollo, L.A.C. Independent sex chromosome evolution in lower vertebrates: A molecular cytogenetic overview in the Erythrinidae fish family. Cytogenet. Genome Res. 2013, 141, 186–194. [Google Scholar] [CrossRef] [PubMed]
- Artoni, R.F.; Falcão, J.N.; Moreira-Filho, O.; Bertollo, L.A.C. An uncommon condition for a sex chromosome system in Characidae fish. Distribution and differentiation of the ZZ/ZW system in Triportheus. Chromosome Res. 2001, 9, 449–456. [Google Scholar] [CrossRef] [PubMed]
- Yano, C.F.; Bertollo, L.A.C.; Liehr, T.; Troy, W.P.; Cioffi, M.B. W Chromosome dynamics in Triportheus species (Characiformes, Triportheidae): An ongoing process narrated by repetitive sequences. J. Hered. 2016, 107, 342–348. [Google Scholar] [CrossRef] [PubMed]
- Galetti, P.M., Jr.; Lima, N.R.W.; Venere, P.C. A monophyletic ZW sex chromosome system in Leporinus (Anostomidae, Characiformes). Cytologia 1995, 60, 375–382. [Google Scholar] [CrossRef]
- Vicente, V.E.; Bertollo, L.A.C.; Valentini, S.R.; Moreira-Filho, O. Origin and differentiation of sex chromosome system in Parodon hilarii (Pisces, Parodontidae). Satellite DNA, G and C-banding. Genetica 2003, 119, 115–120. [Google Scholar] [CrossRef] [PubMed]
- Manchado, M.; Zuasti, E.; Cross, I.; Merlo, A.; Infante, C.; Rebordinos, L. Molecular characterization and chromosomal mapping of the 5S rRNA gene in Solea senegalensis: A new linkage to the U1, U2, and U5 small nuclear RNA genes. Genome 2006, 49, 79–86. [Google Scholar] [CrossRef] [PubMed]
- Pansonato-Alves, J.C.; Serrano, É.A.; Utsunomia, R.; Scacchetti, P.C.; Oliveira, C.; Foresti, F. Single origin of sex chromosomes and multiple origins of B chromosomes in fish genus Characidium. PLoS ONE 2014, 9, e107169. [Google Scholar] [CrossRef] [PubMed]
- Pennell, M.W.; Kirkpatrick, M.; Otto, S.P.; Vamosi, J.C.; Peichel, C.L.; Valenzuela, N.; Kitano, J. Y fuse? Sex chromosome fusions in fishes and reptiles. PLoS Genet. 2015, 11, e1005237. [Google Scholar] [CrossRef] [PubMed]
- Abbott, J.K.; Nordén, A.K.; Hansson, B. Sex chromosome evolution: Historical insights and future perspectives. Proc. Biol. Sci. 2017, 284. [Google Scholar] [CrossRef] [PubMed]
- Van Doorn, G.S.; Kirkpatrick, M. Turnover of sex chromosomes induced by sexual conflict. Nature 2007, 449, 909–912. [Google Scholar] [CrossRef] [PubMed]
- Kikuchi, K.; Hamaguchi, S. Novel sex-determining genes in fish and sex chromosome evolution. Dev. Dyn. 2013, 242, 339–353. [Google Scholar] [CrossRef] [PubMed]
- Pokorná, M.; Altmanová, M.; Kratochvíl, L. Multiple sex chromosomes in the light of female meiotic drive in amniote vertebrates. Chromosome Res. 2014, 22, 35–44. [Google Scholar] [CrossRef] [PubMed]
- Brykov, V.A. Mechanisms of sex determination in fish: Evolutionary and practical aspects. Russ. J. Mar. Biol. 2014, 40, 407–417. [Google Scholar] [CrossRef]
- Bachtrog, D.; Mank, J.E.; Peichel, C.L.; Kirkpatrick, M.; Otto, S.P.; Ashman, T.L.; Hahn, M.W.; Kitano, J.; Mayrose, I.; Ming, R.; et al. Sex determination: Why so many ways of doing it? PLoS Biol. 2014, 12, e1001899. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martínez, P.; Viñas, A.M.; Sánchez, L.; Díaz, N.; Ribas, L.; Piferrer, F. Genetic architecture of sex determination in fish: Applications to sex ratio control in aquaculture. Front. Genet. 2014, 5, 340. [Google Scholar] [CrossRef] [PubMed]
- Henning, F.; Trifonov, V.; Ferguson-Smith, M.A.; Almeida-Toledo, L.F. Non-homologous sex chromosomes in two species of the genus Eigenmannia (Teleostei: Gymnotiformes). Cytogenet. Genome Res. 2008, 121, 55–58. [Google Scholar] [CrossRef] [PubMed]
- Pazian, M.F.; Shimabukuro-Dias, C.K.; Pansonato-Alves, J.C.; Oliveira, C.; Foresti, F. Chromosome painting of Z and W sex chromosomes in Characidium (Characiformes, Crenuchidae). Genetica 2013, 141, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Yano, C.F.; Bertollo, L.A.C.; Ezaz, T.; Trifonov, V.; Sember, A.; Liehr, T.; Cioffi, M.B. Highly conserved Z and molecularly diverged W chromosomes in the fish genus Triportheus (Characiformes, Triportheidae). Heredity 2017, 118, 276–283. [Google Scholar] [CrossRef] [PubMed]
- Bertollo, L.A.C.; Born, G.G.; Dergam, J.A.; Fenocchio, A.S.; Moreira-Filho, O. A biodiversity approach in the neotropical Erythrinidae fish, Hoplias malabaricus. Karyotypic survey, geographic distribution of cytotypes and cytotaxonomic considerations. Chromosome Res. 2000, 8, 603–613. [Google Scholar] [CrossRef] [PubMed]
- Bertollo, L.A.C. Chromosome evolution in the Neotropical Erythrinidae fish family: An overview. In Fish Cytogenetics; Pizano, E., Ozouf-Costaz, C., Foresti, F., Kapoor, B.G., Eds.; Science Publishers: Enfield, NH, USA, 2007; pp. 195–211. [Google Scholar]
- Freitas, N.L.; Al-Rikabi, A.B.H.; Bertollo, L.A.C.; Ezaz, T.; Yano, C.F.; Oliveira, E.A.; Hatanaka, T.; Cioffi, M.B. Early stages of XY sex chromosomes differentiation in the fish Hoplias malabaricus (Characiformes, Erythrinidae) revealed by DNA repeats accumulation. Curr. Genomics 2017, 18. [Google Scholar] [CrossRef]
- Oliveira, E.A.; Sember, A.; Bertollo, L.A.C.; Yano, C.F.; Ezaz, T.; Moreira-Filho, O.; Hatanaka, T.; Trifonov, V.; Liehr, T.; Ráb, P.; et al. Tracking the evolutionary pathway of sex chromosomes among fishes: Characterizing the unique XX/XY1Y2 system in Hoplias malabaricus (Teleostei, Characiformes). Chromosoma 2017. Submitted. [Google Scholar]
- Sember, A.; Bertollo, L.A.C.; Yano, C.F.; Hatanaka, T.; Ráb, P.; de Oliveira, E.A.; Cioffi, M.B. Sex chromosome evolution and genomic divergence in the fish Hoplias malabaricus (Characiformes, Erythrinidae). Front. Genet. 2017. Submitted. [Google Scholar]
- Born, G.G.; Bertollo, L.A.C. An XX/XY sex chromosome system in a fish species, Hoplias malabaricus, with a polymorphic NOR-bearing X chromosome. Chromosome Res. 2000, 8, 111–118. [Google Scholar] [CrossRef] [PubMed]
- Cioffi, M.B.; Martins, C.; Vicari, M.R.; Rebordinos, L.; Bertollo, L.A.C. Differentiation of the XY sex chromosomes in the fish Hoplias malabaricus (Characiformes, Erythrinidae). Unusual accumulation of repetitive sequences on the X chromosome. Sex. Dev. 2010, 4, 176–185. [Google Scholar] [CrossRef] [PubMed]
- Cioffi, M.B.; Martins, C.; Bertollo, L.A.C. Comparative chromosome mapping of repetitive sequences. Implications for genomic evolution in the fish, Hoplias malabaricus. BMC Genet. 2009, 10, 34. [Google Scholar] [CrossRef] [PubMed]
- Cioffi, M.B.; Kejnovský, E.; Bertollo, L.A.C. The chromosomal distribution of microsatellite repeats in the genome of the wolf fish Hoplias malabaricus, focusing on the sex chromosomes. Cytogenet. Genome Res. 2011, 132, 289–296. [Google Scholar] [CrossRef] [PubMed]
- Cioffi, M.B.; Sánchez, A.; Marchal, J.A.; Kosyakova, N.; Liehr, T.; Trifonov, V.; Bertollo, L.A.C. Whole chromosome painting reveals independent origin of sex chromosomes in closely related forms of a fish species. Genetica 2011, 139, 1065–1072. [Google Scholar] [CrossRef] [PubMed]
- Cioffi, M.B.; Bertollo, L.A.C. Initial steps in XY chromosome differentiation in Hoplias. malabaricus and the origin of an X1X2Y sex chromosome system in this fish group. Heredity 2010, 105, 554–561. [Google Scholar] [CrossRef] [PubMed]
- Bertollo, L.A.C.; Fontes, M.S.; Fenocchio, A.S.; Cano, J. The X1X2Y sex chromosome system in the fish Hoplias. malabaricus. I. G-, C- and chromosome replication banding. Chromosome Res. 1997, 5, 493–499. [Google Scholar] [CrossRef] [PubMed]
- Bertollo, L.A.C.; Mestriner, C.A. The X1X2Y sex chromosome system in the fish Hoplias. malabaricus (Pisces, Erythrinidae). II. Meiotic analyses. Chromosome Res. 1998, 6, 141–147. [Google Scholar] [CrossRef] [PubMed]
- Rosa, R.; Laforga Vanzela, A.L.; Rubert, M.; Martins-Santos, I.C.; Giuliano-Caetano, L. Differentiation of Y chromosome in the X1X1X2X2/X1X2Y sex chromosome system of Hoplias malabaricus (Characiformes, Erythrinidae). Cytogenet. Genome Res. 2009, 127, 54–60. [Google Scholar] [CrossRef] [PubMed]
- Green, J.E.; Dalíková, M.; Sahara, K.; Marec, F.; Akam, M. XX/XY system of sex determination in the geophilomorph centipede Strigamia maritima. PLoS ONE 2016, 11, e0150292. [Google Scholar] [CrossRef] [PubMed]
- Bertollo, L.A.C.; Oliveira, C.; Molina, W.F.; Margarido, V.P.; Fontes, M.S.; Pastori, M.C.; Falcão, J.N.; Fenocchio, A.S. Chromosome evolution in the erythrinid fish, Erythrinus erythrinus (Teleostei: Characiformes). Heredity 2004, 93, 228–233. [Google Scholar] [CrossRef] [PubMed]
- Cioffi, M.B.; Martins, C.; Bertollo, L.A.C. Chromosome spreading of associated transposable elements and ribosomal DNA in the fish Erythrinus erythrinus. Implications for genome change and karyoevolution in fish. BMC Evol. Biol. 2010, 10, 271. [Google Scholar] [CrossRef] [PubMed]
- Martins, N.F.; Bertollo, L.A.C.; Troy, W.P.; Feldberg, E.; Valentin, F.C.S.; Cioffi, M.B. Differentiation and evolutionary relationships in Erythrinus erythrinus (Characiformes, Erythrinidae): Comparative chromosome mapping of repetitive sequences. Rev. Fish Biol. Fish. 2013, 23, 261–269. [Google Scholar] [CrossRef]
- Cioffi, M.B.; Sánchez, A.; Marchal, J.A.; Kosyakova, N.; Liehr, T.; Trifonov, V.; Bertollo, L.A.C. Cross-species chromosome painting tracks the independent origin of multiple sex chromosomes in two cofamiliar Erythrinidae fishes. BMC Evol. Biol. 2011, 11, 186. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, C.; Avelino, G.S.; Abe, K.T.; Mariguela, T.C.; Benine, R.C.; Ortí, G.; Vari, R.P.; Corrêa e Castro, R.M. Phylogenetic relationships within the speciose family Characidae (Teleostei: Ostariophysi: Characiformes) based on multilocus analysis and extensive ingroup sampling. BMC Evol. Biol. 2011, 11, 275. [Google Scholar] [CrossRef] [PubMed]
- Mariguela, T.C.; Roxo, F.F.; Foresti, F.; Oliveira, C. Phylogeny and biogeography of Triportheidae (Teleostei: Characiformes) based on molecular data. Mol. Phylogenet. Evol. 2016, 96, 130–139. [Google Scholar] [CrossRef] [PubMed]
- Malabarba, M.C.S.L. Revision of the Neotropical genus Triportheus. Cope, 1872 (Characiformes: Characidae). Neotrop. Ichthyol. 2004, 2, 167–204. [Google Scholar] [CrossRef]
- Prestes, L.; Mota Soares, M.G.; Silva, F.R.; Bittencourt, M.M. Dynamic population from Triportheus albus, T. angulatus and T. auritus (Characiformes: Characidae) in Amazonian Central lakes. Biota Neotrop. 2010, 10, 177–181. [Google Scholar] [CrossRef]
- Falcão, J.N. Caracterização Cariotípica em Peixes do Gênero Triportheus (Teleostei, Characiformes, Characidae). Ph.D. Thesis, Universidade de São Paulo, Ribeirão Preto, SP, Brazil, 1988. [Google Scholar]
- Bertollo, L.A.C.; Cavallaro, Z.I.A. Highly differentiated ZZ/ZW sex chromosome system in a Characidae fish, Triportheus guentheri. Cytogenet. Genome Res. 1992, 60, 60–63. [Google Scholar] [CrossRef]
- Sanchez, S.; Jorge, L.C. A new report of the ZZ/ZW sex chromosome system in the genus Triportheus. (Pisces, Triportheinae). Cytologia 1999, 64, 395–400. [Google Scholar] [CrossRef]
- Artoni, R.F.; Bertollo, L.A.C. Evolutionary aspects of the ZZ/ZW sex chromosome system in the Characidae fish, genus Triportheus. A monophyletic state and NOR location on the W chromosome. Heredity 2002, 89, 15–19. [Google Scholar] [CrossRef] [PubMed]
- Nirchio, M.; Oliveira, C.; Ferreira, I.A.; Granado, A.; Ron, E. Extensive polymorphism and chromosomal characteristics of ribosomal DNA in the characid fish Triportheus venezuelensis (Characiformes, Characidae). Genet. Mol. Biol. 2007, 30, 25–30. [Google Scholar] [CrossRef]
- Diniz, D.; Laudicina, A.; Cioffi, M.B.; Bertollo, L.A.C. Microdissection and whole chromosome painting. Improving sex chromosome analysis in Triportheus (Teleostei, Characiformes). Cytogenet. Genome Res. 2008, 122, 163–168. [Google Scholar] [CrossRef] [PubMed]
- Diniz, D.; Moreira-Filho, O.; Bertollo, L.A.C. Molecular cytogenetics and characterization of a ZZ/ZW sex chromosome system in Triportheus nematurus (Characiformes, Characidae). Genetica 2008, 133, 85–91. [Google Scholar] [CrossRef] [PubMed]
- Diniz, D.; Laudicina, A.; Bertollo, L.A.C. Chromosomal location of 18S and 5S rDNA sites in Triportheus fish species (Characiformes, Characidae). Genet. Mol. Biol. 2009, 32, 37–41. [Google Scholar] [CrossRef] [PubMed]
- Yano, C.F.; Poltronieri, J.; Bertollo, L.A.C.; Artoni, R.F.; Liehr, T.; Cioffi, M.B. Chromosomal mapping of repetitive DNAs in Triportheus trifurcatus (Characidae, Characiformes): Insights into the differentiation of the Z and W chromosomes. PLoS ONE 2014, 9, e90946. [Google Scholar] [CrossRef] [PubMed]
- Yano, C.F.; Bertollo, L.A.C.; Rebordinos, L.; Merlo, M.A.; Liehr, T.; Portela-Bens, S. Evolutionary dynamics of rDNAs and U2 small nuclear DNAs in Triportheus (Characiformes, Triportheidae): High variability and particular syntenic organization. Zebrafish 2017, 14, 146–154. [Google Scholar] [CrossRef] [PubMed]
- Marquioni, V.; Bertollo, L.A.C.; Diniz, D.; Cioffi, M.B. Comparative chromosomal mapping in Triportheus fish species. Analysis of synteny between ribosomal genes. Micron 2013, 45, 129–135. [Google Scholar] [CrossRef] [PubMed]
- Reed, K.M.; Phillips, R.B. Polymorphism of the nucleolar organizer region (NOR) on the putative sex chromosomes of Arctic char (Salvelinus alpinus) is not sex related. Chromosome Res. 1997, 5, 221–227. [Google Scholar] [CrossRef] [PubMed]
- Kawai, A.; Nishida-Umehara, C.; Ishijima, J.; Tsuda, Y.; Ota, H.; Matsuda, Y. Different origins of bird and reptile sex chromosomes inferred from comparative mapping of chicken Z-linked genes. Cytogenet. Genome Res. 2007, 117, 92–102. [Google Scholar] [CrossRef] [PubMed]
- Badenhorst, D.; Stanyon, R.; Engstrom, T.; Valenzuela, N. A ZZ/ZW microchromosome system in the spiny softshell turtle, Apalone spinifera, reveals an intriguing sex chromosome conservation in Trionychidae. Chromosome Res. 2013, 21, 137–147. [Google Scholar] [CrossRef] [PubMed]
- Gatto, K.P.; Busin, C.S.; Lourenço, L.B. Unraveling the sex chromosome heteromorphism of the paradoxical frog Pseudis tocantins. PLoS ONE 2016, 11, e0156176. [Google Scholar] [CrossRef] [PubMed]
- Kejnovský, E.; Michalovová, M.; Šteflová, P.; Kejnovská, I.; Manzano, S.; Hobza, R.; Kubát, Z.; Kovařík, J.; Jamilena, M.; Vyskot, B. Expansion of microsatellites on evolutionary young Y chromosome. PLoS ONE 2013, 8, e45519. [Google Scholar] [CrossRef] [PubMed]
- Cioffi, M.B.; Kejnovský, E.; Marquioni, V.; Poltronieri, J.; Molina, W.F.; Diniz, D.; Bertollo, L.A.C. The key role of repeated DNAs in sex chromosome evolution in two fish species with ZW sex chromosome system. Mol. Cytogenet. 2012, 5, 28. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues, A.S.; Aline Souza Medrado, A.S.; Diniz, D.; Oliveira, C.; Affonso, P.R.A.M. ZZ/ZW sex chromosome system in the endangered fish Lignobrycon myersi Miranda-Ribeiro, 1956 (Teleostei, Characiformes, Triportheidae). Comp. Cytogenet. 2016, 10, 245–254. [Google Scholar] [CrossRef]
- Terencio, M.L.; Schneider, C.H.; Gross, M.C.; Silva, A.M.; Feldberg, E.; Porto, J.I.R. Comparative cytogenetics of Carnegiella marthae and Carnegiella strigata (Characiformes, Gasteropelecidae) and description of a ZZ/ZW sex chromosome system. Genet. Mol. Biol. 2008, 31, 231–234. [Google Scholar] [CrossRef]
- Soares, R.X.; Bertollo, L.A.C.; Cioffi, M.B.; Costa, G.W.W.F.; Molina, W.F. Chromosomal distribution of two multigene families and the unusual occurrence of an X1X1X2X2/X1X2Y sex chromosome system in the dolphinfish (Coryphaenidae): An evolutionary perspective. Genet. Mol. Res. 2014, 13, 2470–2479. [Google Scholar] [CrossRef] [PubMed]
- Bitencourt, J.A.; Sampaio, I.; Ramos, R.T.C.; Vicari, M.R.; Affonso, P.R.A.M. First report of sex chromosomes in Achiridae (Teleostei: Pleuronectiformes) with inferences about the origin of the multiple X1X1X2X2/X1X2Y system and dispersal of ribosomal genes in Achirus achirus. Zebrafish 2016, 14, 90–95. [Google Scholar] [CrossRef] [PubMed]
- Moreira-Filho, O.; Bertollo, L.A.C.; Galetti, P.M., Jr. Distribution of sex chromosome mechanisms in neotropical fish and description of a ZZ/ZW system in Parodon hilarii (Parodontidae). Caryologia 1993, 46, 115–125. [Google Scholar] [CrossRef]
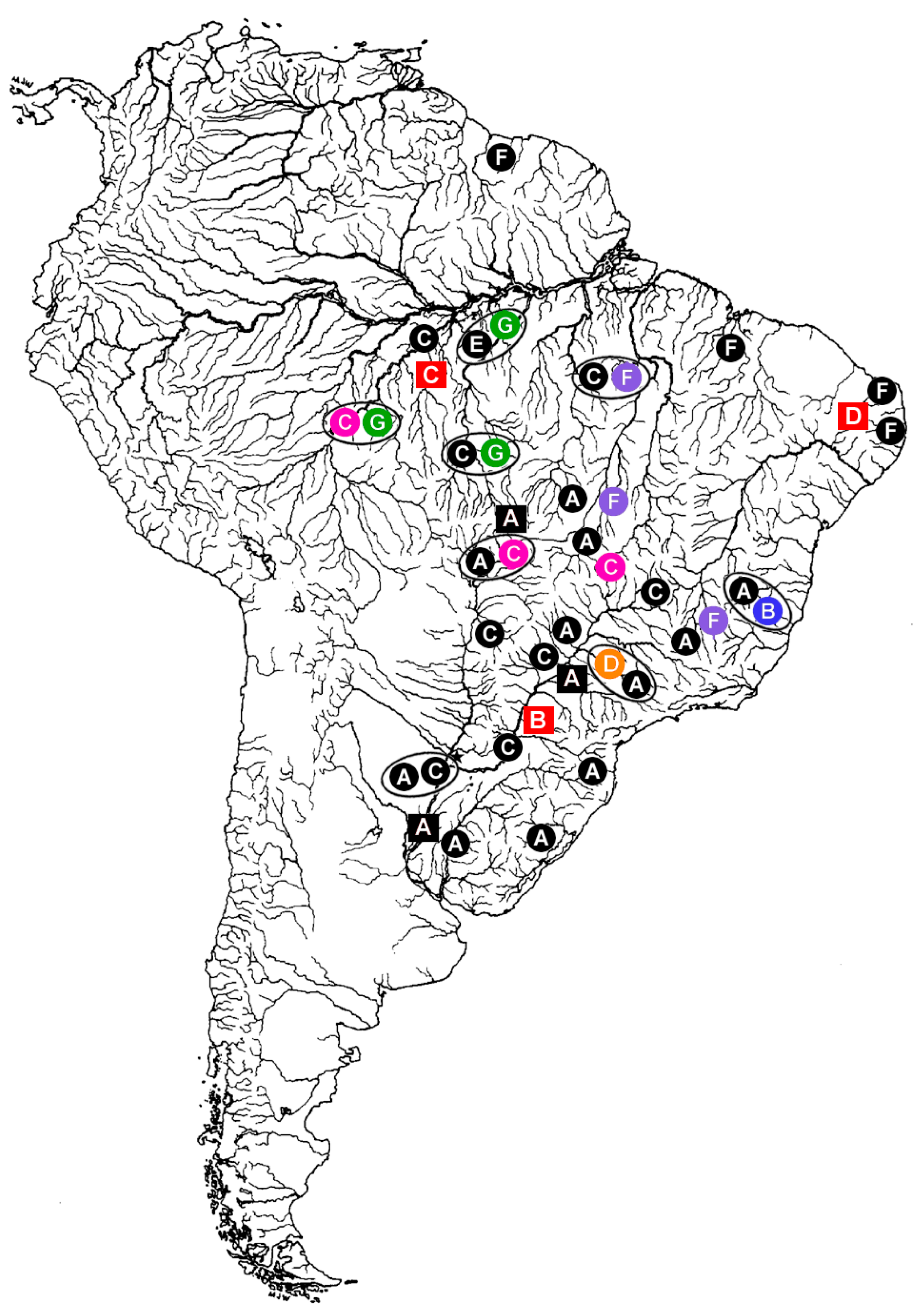
Figure 1.
Distribution of Hoplias malabaricus (HMA) karyomorphs A–G (circles) and Erythrinus erythrinus (ERY) karyomorphs A–D (squares) in the South America. The large open circles indicate some of the sympatric conditions already detected among distinct H. malabaricus karyomorphs. The sex chromosome systems until now identified among the distinct karyomoprhs are highlighted in colors, as follows: Black: karyomorphs with homomorphic and/or unidentified sex chromosomes; Blue: the XY sex system of HMA karyomorph B; Pink: the XY sex system of HMA karyomorph C; Orange: the X1X2Y sex system of karyomorph D; Purple: the XY system of HMA karyomorph F; Green: The XY1Y2 sex system of karyomorph G; Red: the X1X2Y sex system of ERY karyomorphs B, C, and D. Note that the occurrence of XY sex systems in HMA karyomorphs C and F are highlighted only for populations where these systems were already investigated. Modified from [24].
Figure 1.
Distribution of Hoplias malabaricus (HMA) karyomorphs A–G (circles) and Erythrinus erythrinus (ERY) karyomorphs A–D (squares) in the South America. The large open circles indicate some of the sympatric conditions already detected among distinct H. malabaricus karyomorphs. The sex chromosome systems until now identified among the distinct karyomoprhs are highlighted in colors, as follows: Black: karyomorphs with homomorphic and/or unidentified sex chromosomes; Blue: the XY sex system of HMA karyomorph B; Pink: the XY sex system of HMA karyomorph C; Orange: the X1X2Y sex system of karyomorph D; Purple: the XY system of HMA karyomorph F; Green: The XY1Y2 sex system of karyomorph G; Red: the X1X2Y sex system of ERY karyomorphs B, C, and D. Note that the occurrence of XY sex systems in HMA karyomorphs C and F are highlighted only for populations where these systems were already investigated. Modified from [24].
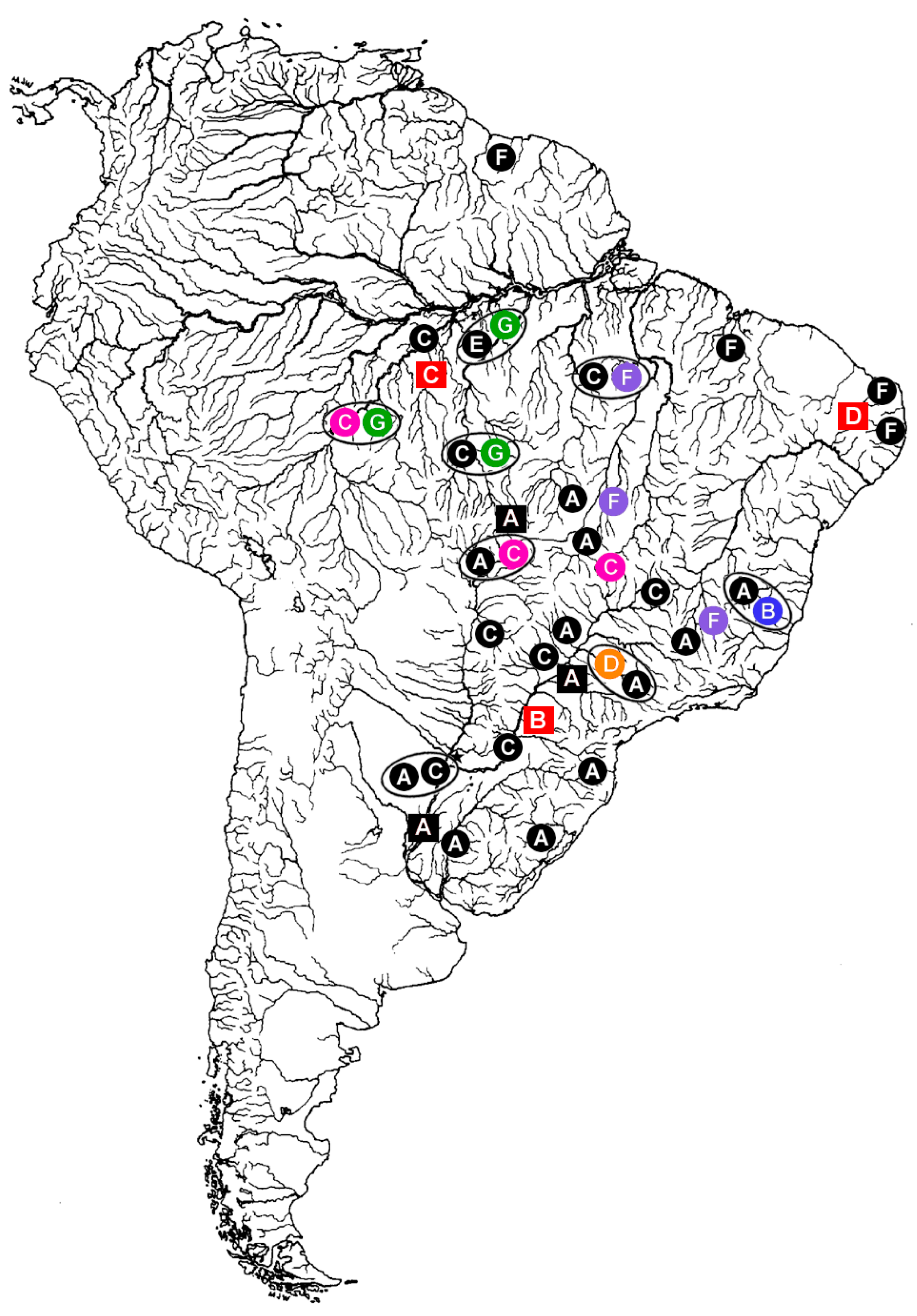
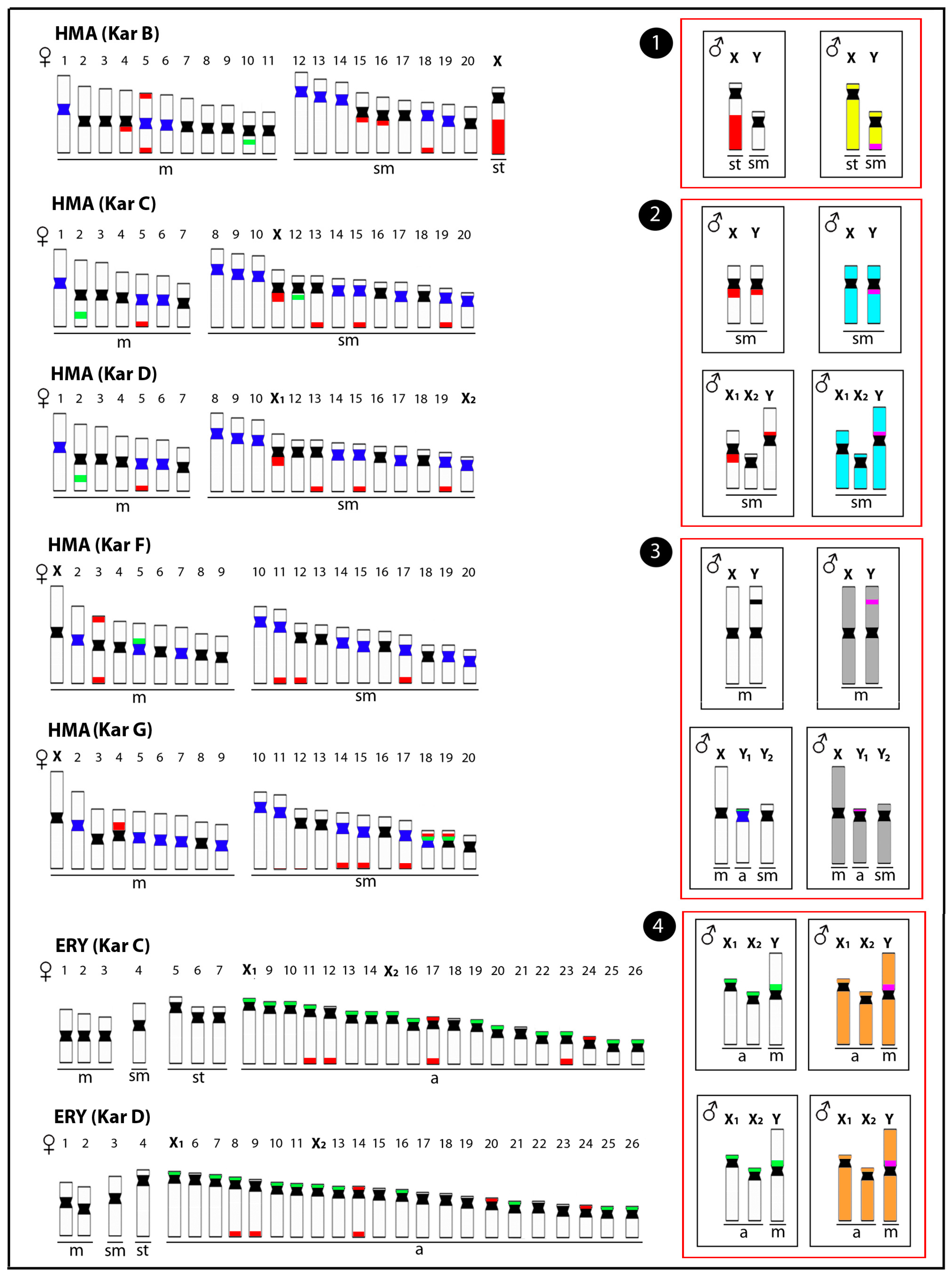
Figure 2.
Representative idiograms of Erythrinidae species Hoplias malabaricus (HMA) and Erythrinus erythrinus (ERY) with differentiated sex chromosomes, highlighting the distribution of different classes of repetitive DNAs and the male-specific regions. The locations of the satellite 5SHindIII-DNA, 18S rDNA, and 5S rDNA sites on the chromosomes are indicated in dark blue, red, and green, respectively. The male-specific regions identified by CGH are highlighted in pink. The sex chromosomes of these species have resulted from at least four independent evolutionary events (red boxes), including (1) the XY chromosomes of HMA karyomorph B (yellow), (2) the XY chromosomes of HMA karyomorph C and their derived X1X2Y chromosomes of karyomorph D (light blue), (3) the XY chromosomes of HMA karyomorph F and the XY1Y2 chromosomes of karyomorph G (grey), and (4) the X1X2Y chromosomes of ERY karyomorphs C and D (orange). m = metacentric, sm = submetacentric, st = subtelocentric and a = acrocentric chromosomes. Data from [25,45,50,59,60].
Figure 2.
Representative idiograms of Erythrinidae species Hoplias malabaricus (HMA) and Erythrinus erythrinus (ERY) with differentiated sex chromosomes, highlighting the distribution of different classes of repetitive DNAs and the male-specific regions. The locations of the satellite 5SHindIII-DNA, 18S rDNA, and 5S rDNA sites on the chromosomes are indicated in dark blue, red, and green, respectively. The male-specific regions identified by CGH are highlighted in pink. The sex chromosomes of these species have resulted from at least four independent evolutionary events (red boxes), including (1) the XY chromosomes of HMA karyomorph B (yellow), (2) the XY chromosomes of HMA karyomorph C and their derived X1X2Y chromosomes of karyomorph D (light blue), (3) the XY chromosomes of HMA karyomorph F and the XY1Y2 chromosomes of karyomorph G (grey), and (4) the X1X2Y chromosomes of ERY karyomorphs C and D (orange). m = metacentric, sm = submetacentric, st = subtelocentric and a = acrocentric chromosomes. Data from [25,45,50,59,60].
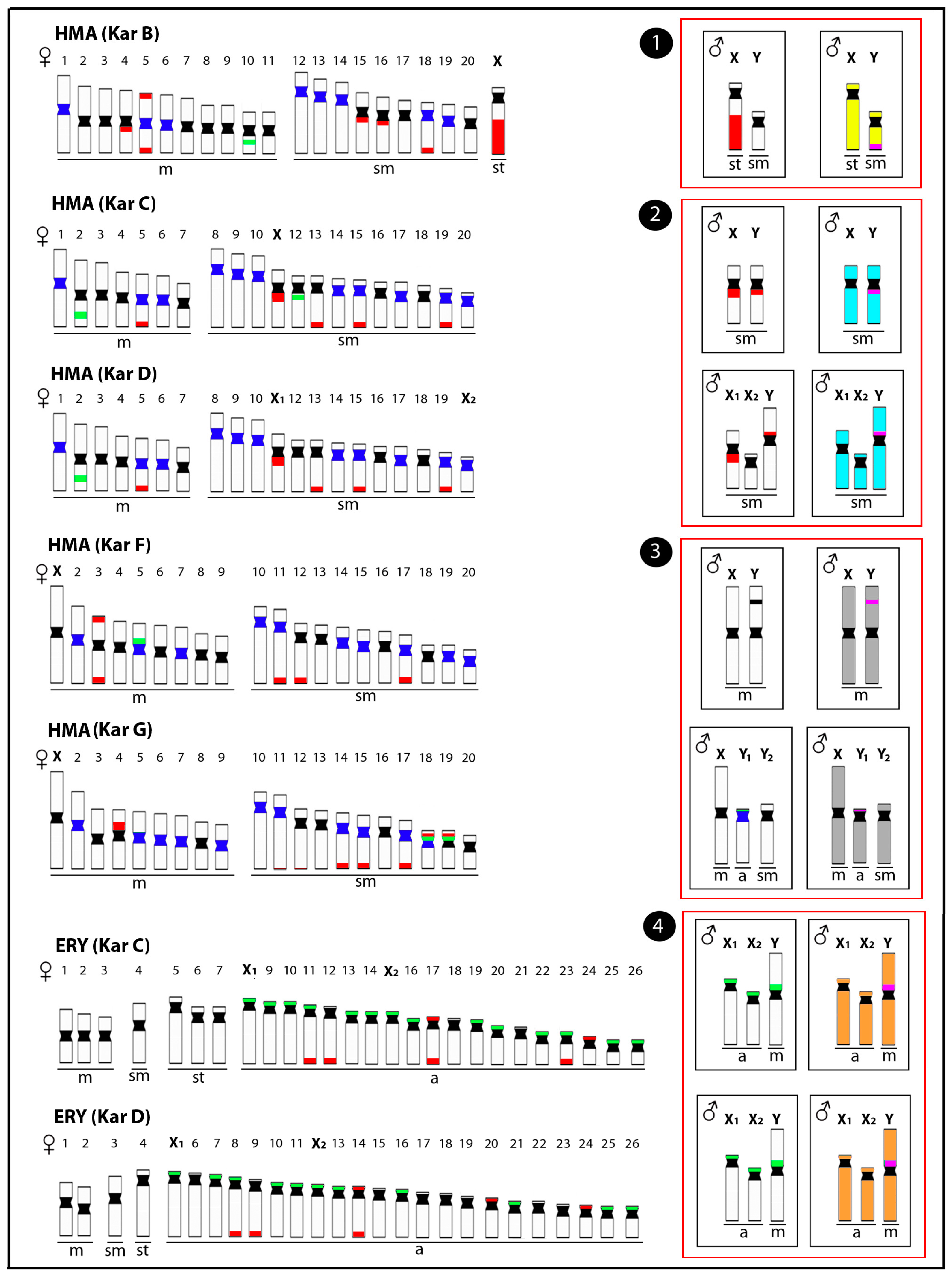
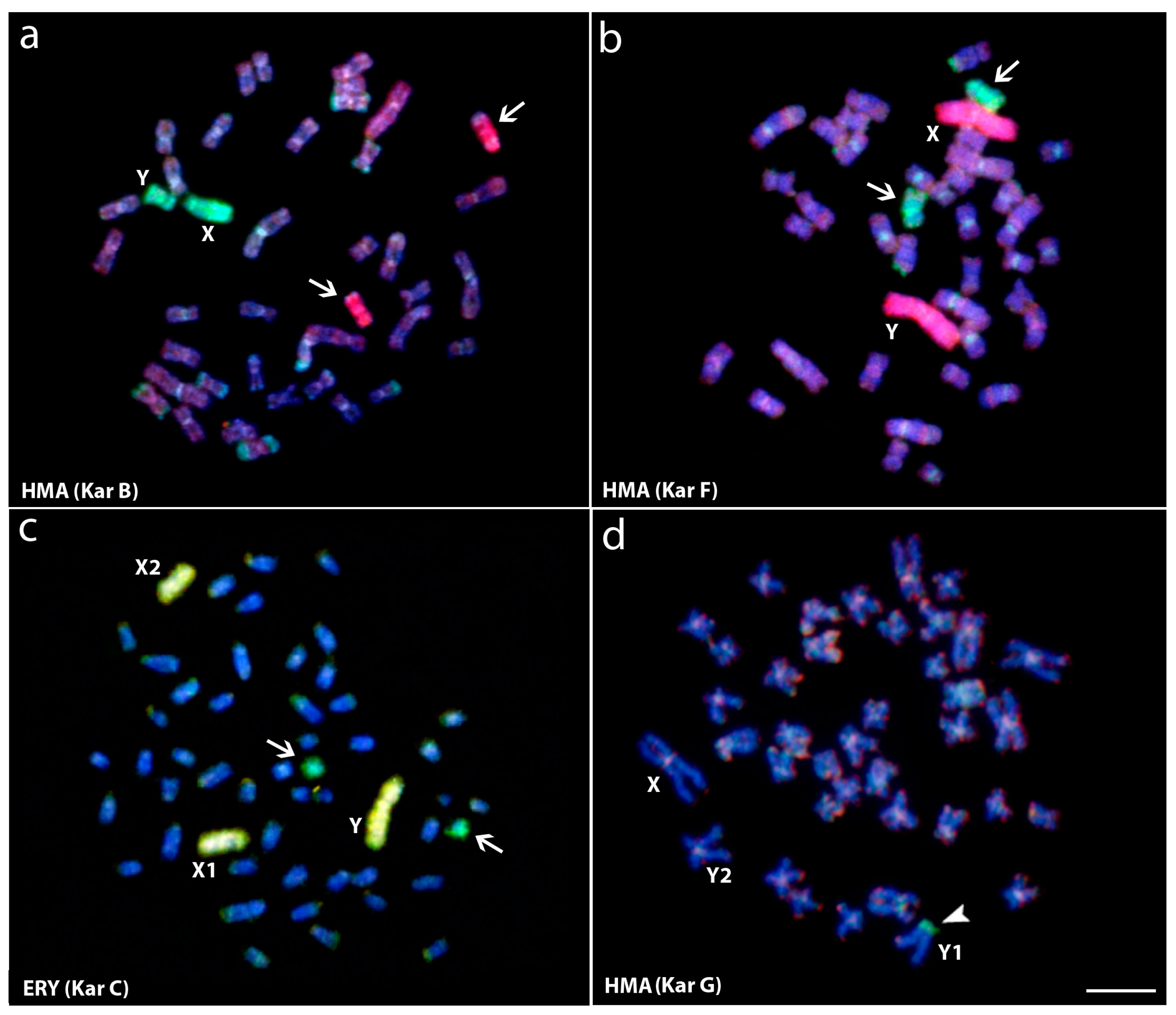
Figure 3.
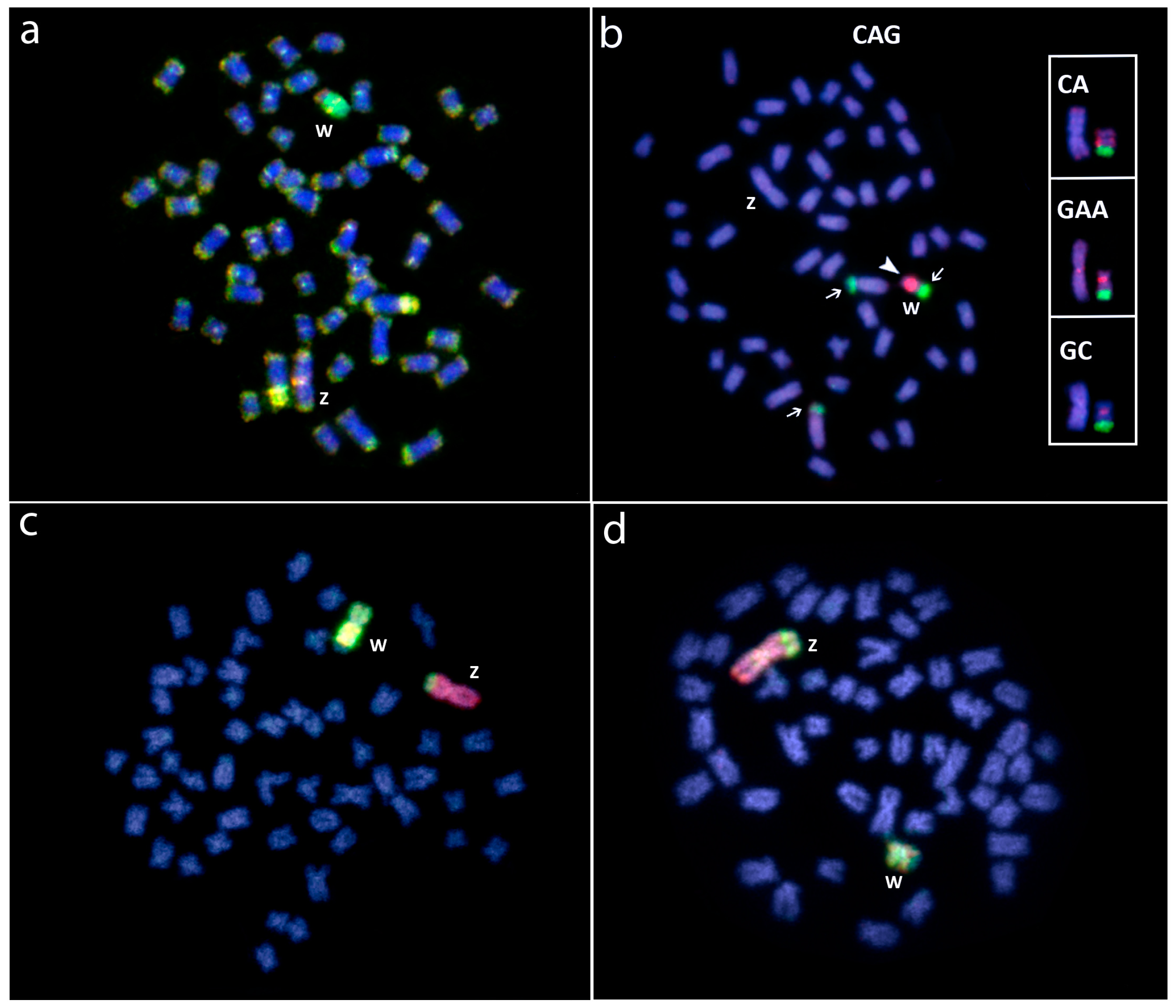
Whole chromosome painting (WCP) and comparative genomic hybridization (CGH) experiments to Hoplias malabaricus (HMA) and Erythrinus erythrinus (ERY) metaphase chromosomes. (a) Male HMA karyomorph B mapped with the X (green) and X1 (red) probes from karyomorphs B and D, respectively; (b) Male of HMA karyomorph F mapped with the X (green) and the Y (red) probes from karyomorphs B and F, respectively; (c) Male metaphase of ERY karyomorph D mapped with the X (green) and the Y (yellow) probes, from HMA karyomorph B and ERY karyomorph D, respectively. The arrows indicate the non-homologous chromosomes painted with the sex-specific probes; (d) Male of HMA karyomorph G after CGH experiments showing the hybridization pattern of the male-derived probe (green) and female-derived probe (red). Arrowhead points to the male-specific region on the Y1 chromosome (d). Bar = 10 μm. Data from [25,46].
Figure 3.
Whole chromosome painting (WCP) and comparative genomic hybridization (CGH) experiments to Hoplias malabaricus (HMA) and Erythrinus erythrinus (ERY) metaphase chromosomes. (a) Male HMA karyomorph B mapped with the X (green) and X1 (red) probes from karyomorphs B and D, respectively; (b) Male of HMA karyomorph F mapped with the X (green) and the Y (red) probes from karyomorphs B and F, respectively; (c) Male metaphase of ERY karyomorph D mapped with the X (green) and the Y (yellow) probes, from HMA karyomorph B and ERY karyomorph D, respectively. The arrows indicate the non-homologous chromosomes painted with the sex-specific probes; (d) Male of HMA karyomorph G after CGH experiments showing the hybridization pattern of the male-derived probe (green) and female-derived probe (red). Arrowhead points to the male-specific region on the Y1 chromosome (d). Bar = 10 μm. Data from [25,46].
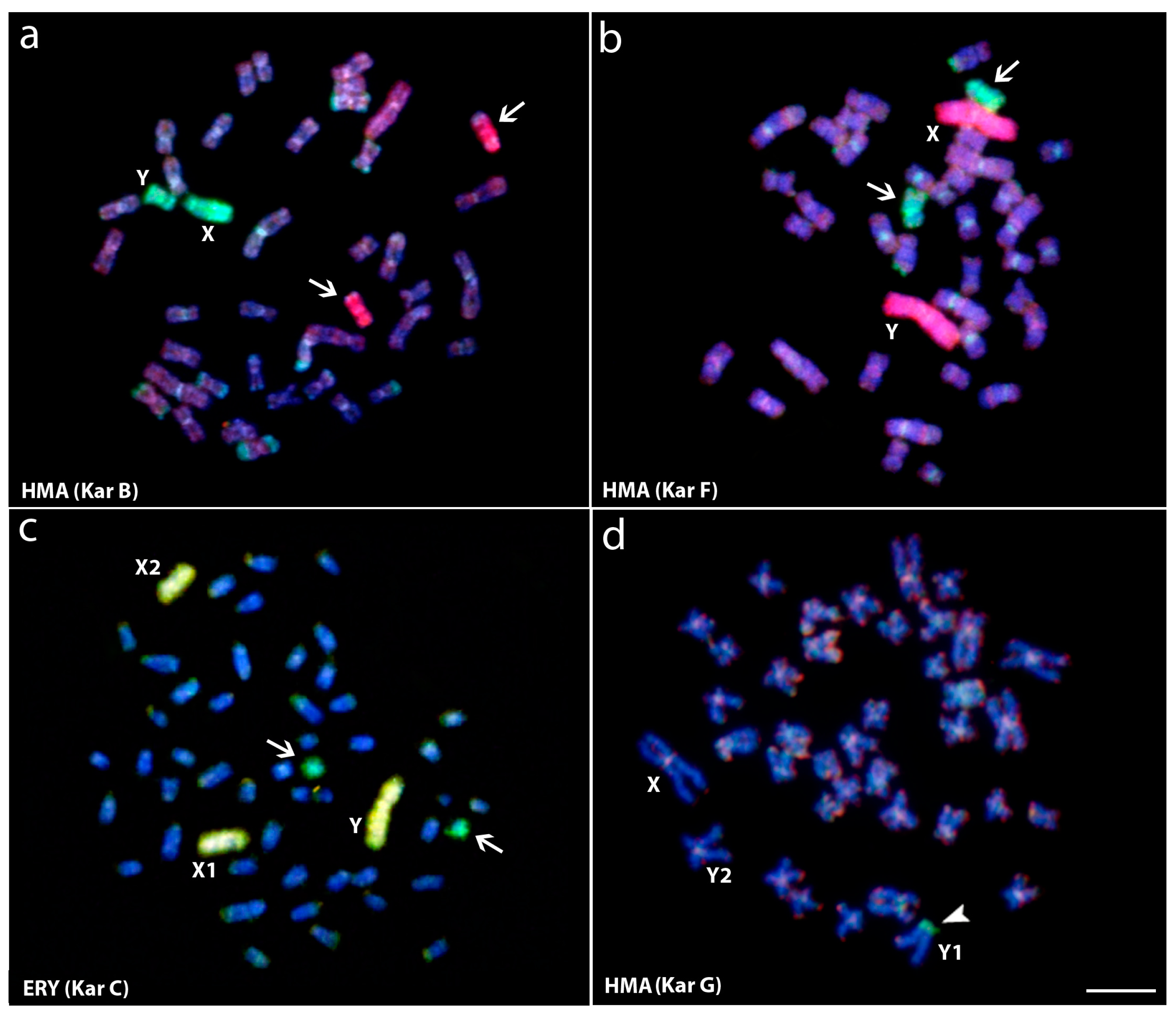
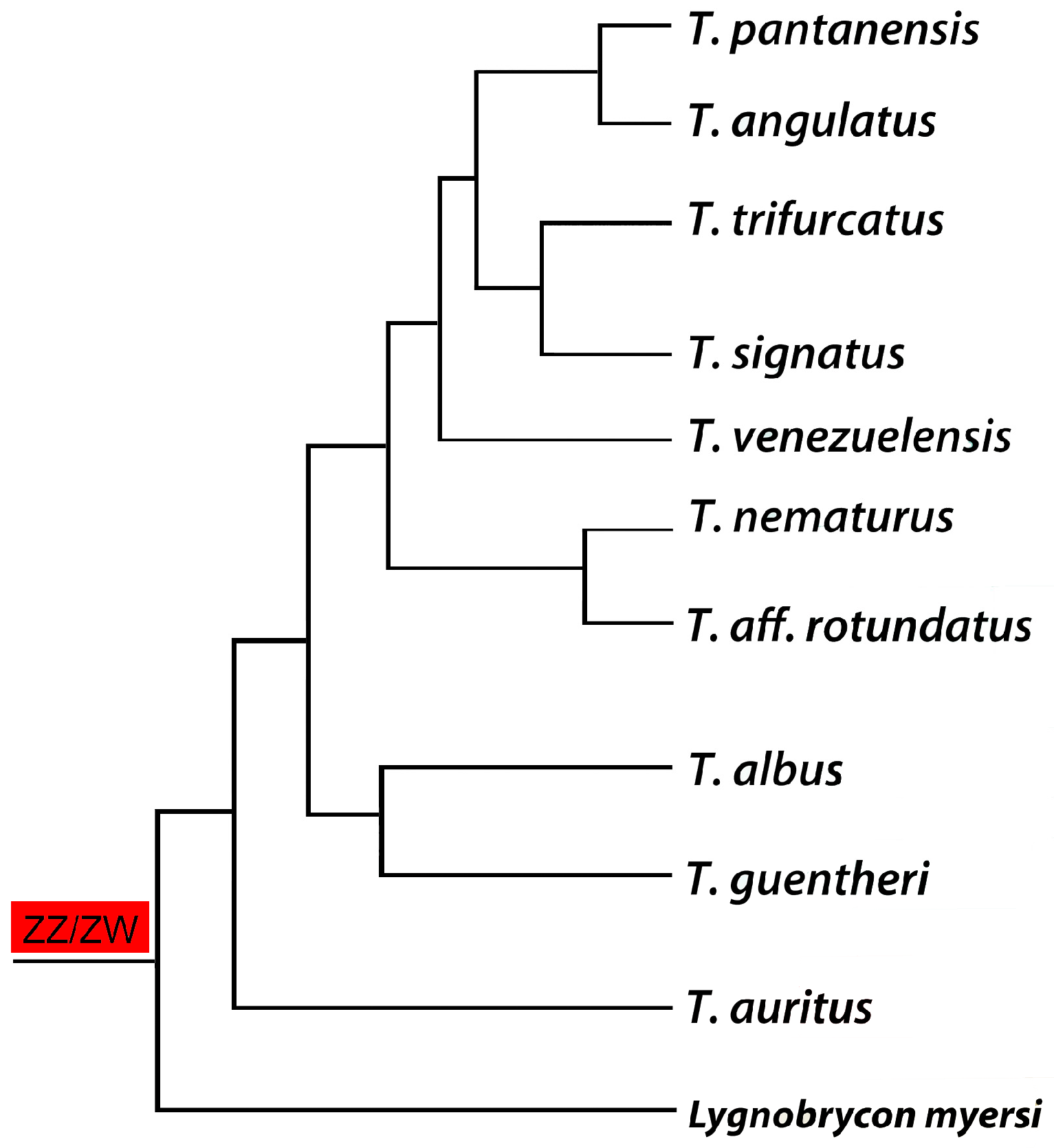
Figure 4.
Phylogenetic tree for the Triportheus genus (based on the data generated by [63]) highlighting the presence of a ZZ/ZW sex chromosome system in all analyzed species. Note that Lignobrycon myersi corresponds to the sister group of all other Triportheidae species and also possesses a similar ZW sex chromosome system, suggesting an early origin of this system in the family.
Figure 4.
Phylogenetic tree for the Triportheus genus (based on the data generated by [63]) highlighting the presence of a ZZ/ZW sex chromosome system in all analyzed species. Note that Lignobrycon myersi corresponds to the sister group of all other Triportheidae species and also possesses a similar ZW sex chromosome system, suggesting an early origin of this system in the family.
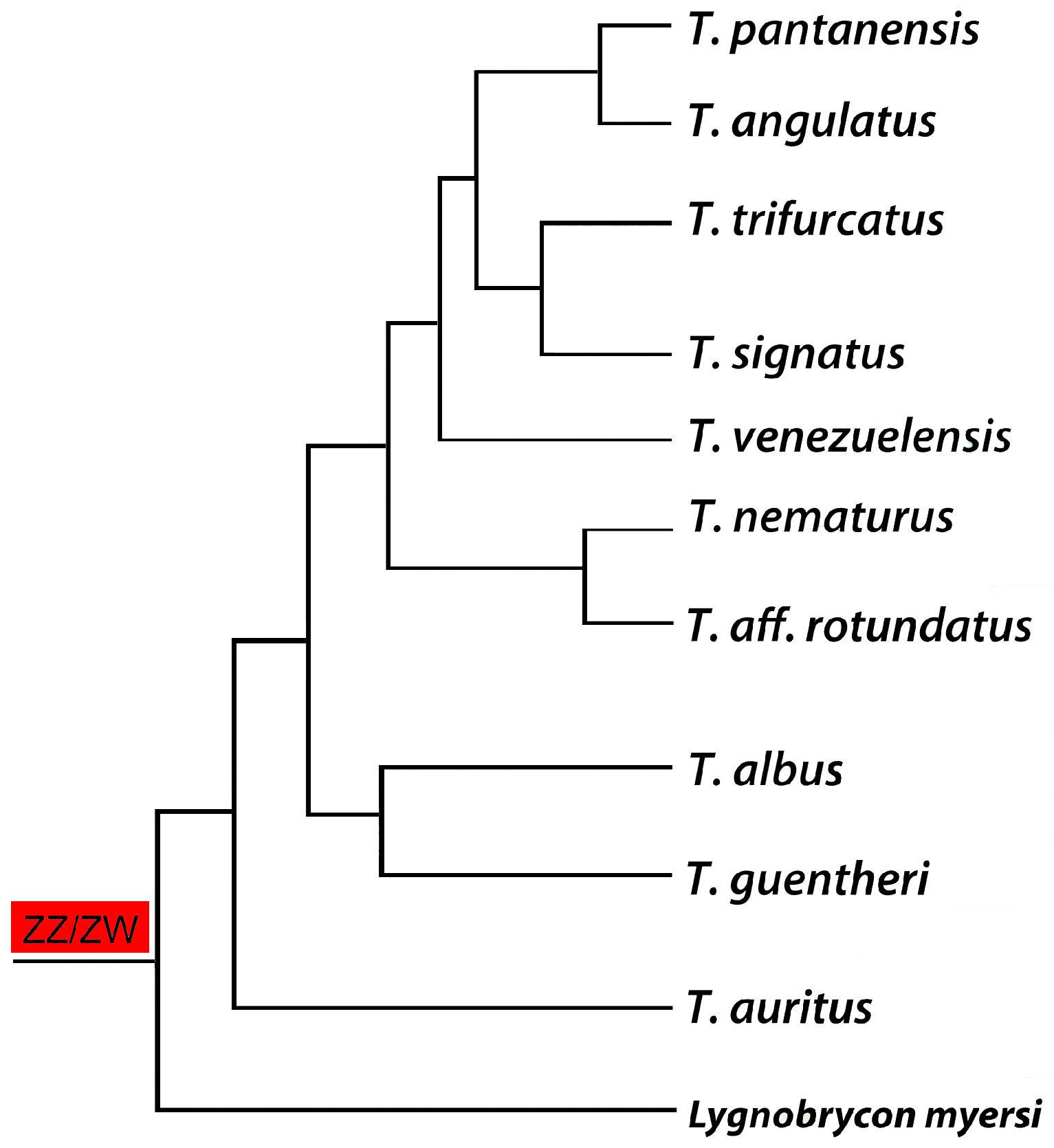
Figure 5.
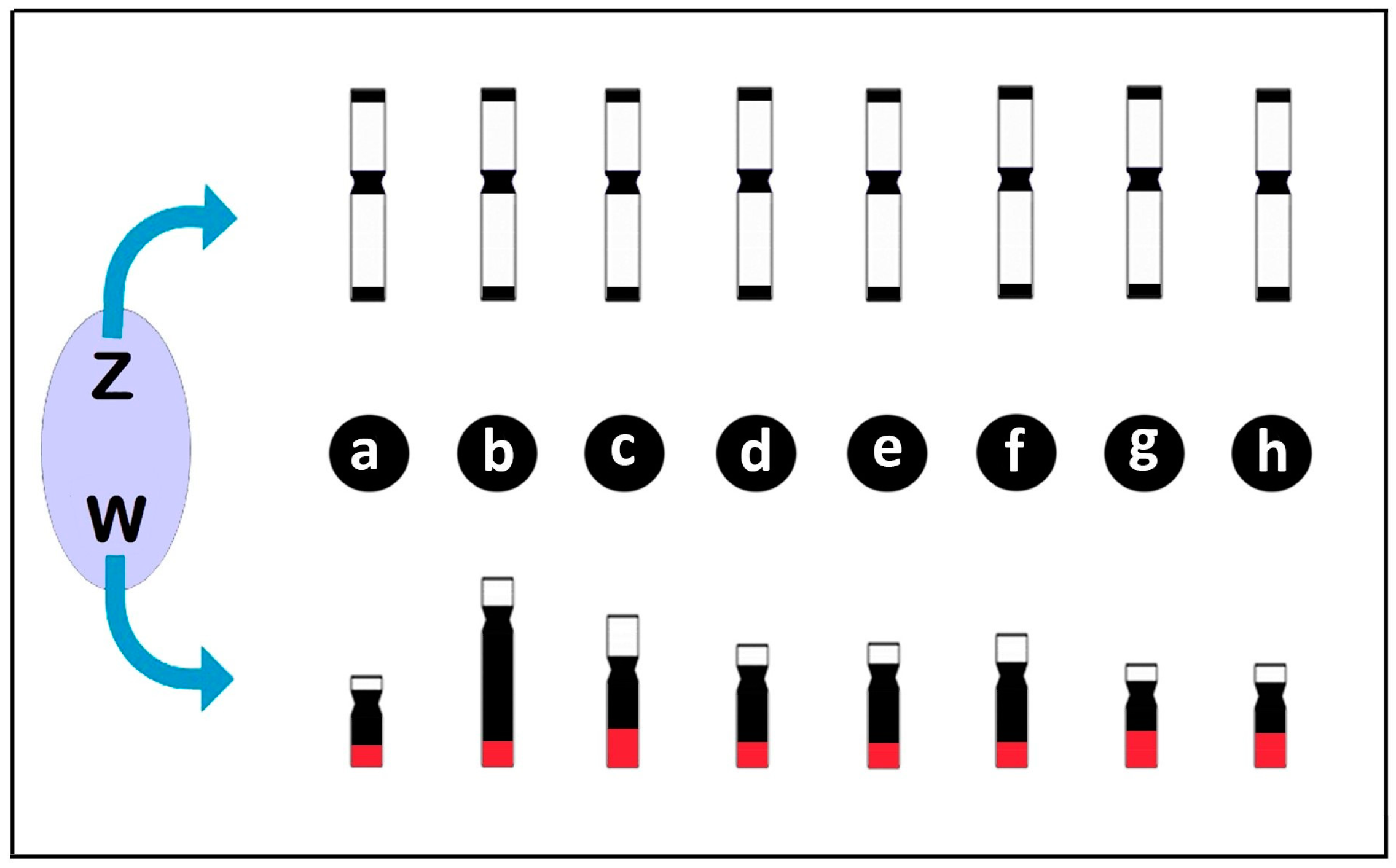
Diagrams illustrating the differentiation of the Z and W chromosomes in Triportheus species highlighting the conserved pattern of the Z chromosome in relation to morphology, size and heterochromatin content (in black), in opposite to the divergent patterns of the W chromosome among species. Note that the W chromosome varies in size, morphology, and heterochromatin content, but carries a 18S rDNA cluster on the long (q) arms (in red) in all species. (a) T. albus; (b) T. auritus; (c) T. guentheri; (d) T. nematurus; (e) T. pantanensis; (f) Triportheus aff. rotundatus; (g) T. signatus; and (h) T. trifurcatus. Data from [42,69,71,76].
Figure 5.
Diagrams illustrating the differentiation of the Z and W chromosomes in Triportheus species highlighting the conserved pattern of the Z chromosome in relation to morphology, size and heterochromatin content (in black), in opposite to the divergent patterns of the W chromosome among species. Note that the W chromosome varies in size, morphology, and heterochromatin content, but carries a 18S rDNA cluster on the long (q) arms (in red) in all species. (a) T. albus; (b) T. auritus; (c) T. guentheri; (d) T. nematurus; (e) T. pantanensis; (f) Triportheus aff. rotundatus; (g) T. signatus; and (h) T. trifurcatus. Data from [42,69,71,76].
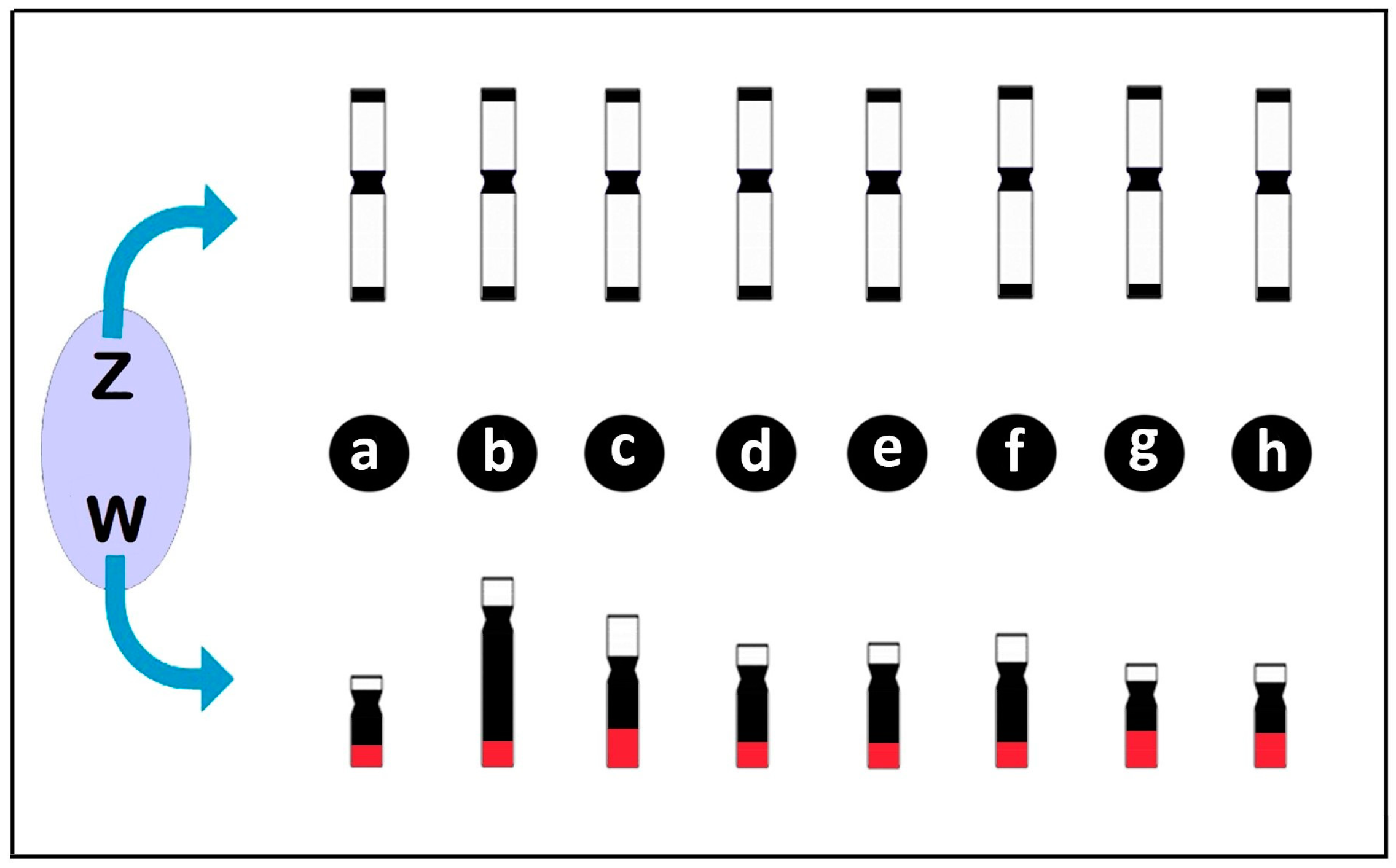
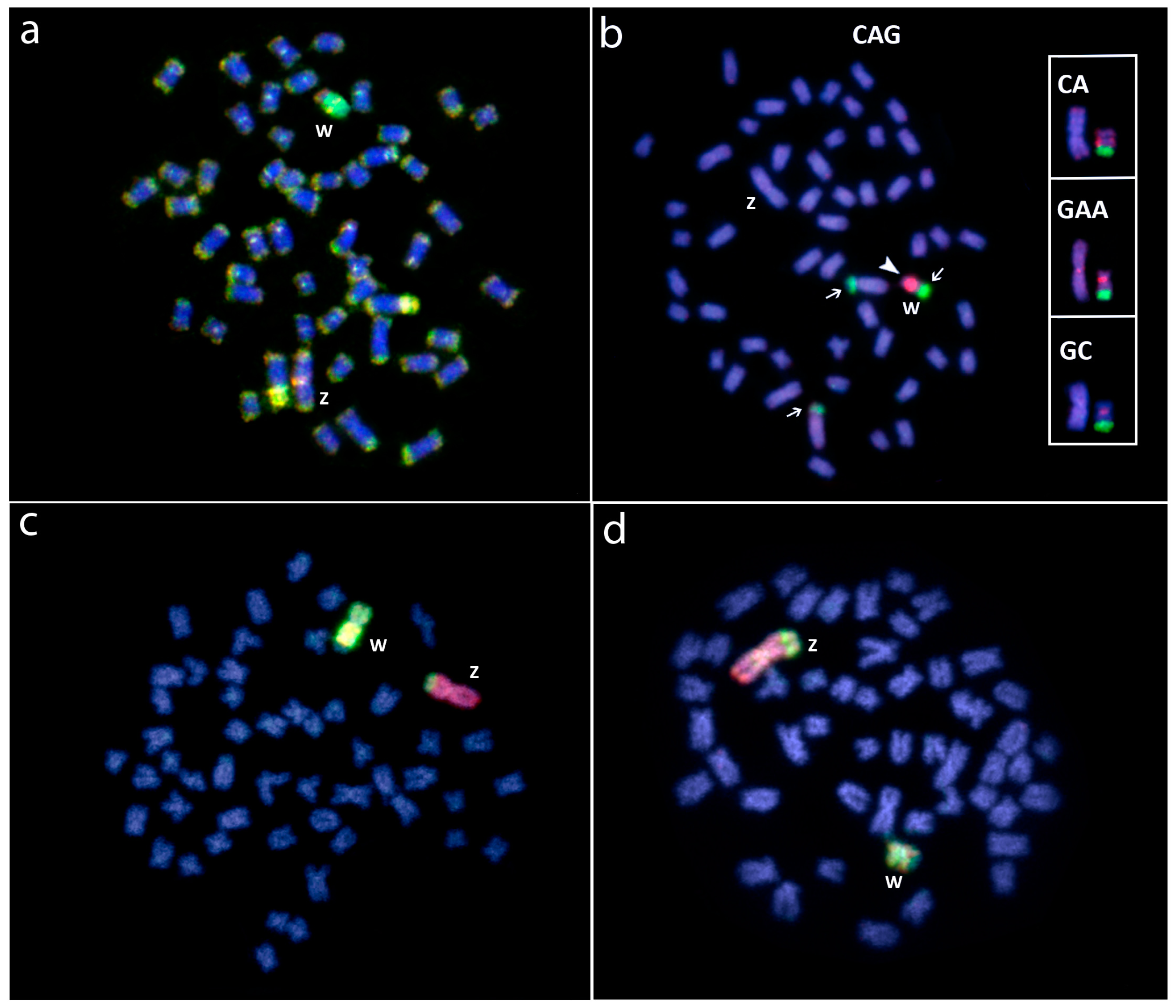
Figure 6.
Molecular cytogenetic data in Triportheus species. (a) Female T. signatus showing the chromosome superposition of female (green) and male (red) gDNA probes after CGH experiments. Note the large accumulation of female-specific sequences in the W chromosome; (b) Female T. signatus chromosomes hybridized with the (CAG)10 microsatellite. The hybridization pattern on the Z and W chromosomes using (CA)15, (CAG)10, (GC)15 microsatellite probes are boxed. Note the preferential accumulation of these repeats on the W chromosome; (c) Whole chromosome painting in female T. auritus and (d) T. albus (d), using W-chromosome (green) and Z-chromosome (red) probes from T. auritus, providing evidence for the common origin of the ZW sex chromosome system in Triportheus. Bar = 10 μm. Data from [27,42].
Figure 6.
Molecular cytogenetic data in Triportheus species. (a) Female T. signatus showing the chromosome superposition of female (green) and male (red) gDNA probes after CGH experiments. Note the large accumulation of female-specific sequences in the W chromosome; (b) Female T. signatus chromosomes hybridized with the (CAG)10 microsatellite. The hybridization pattern on the Z and W chromosomes using (CA)15, (CAG)10, (GC)15 microsatellite probes are boxed. Note the preferential accumulation of these repeats on the W chromosome; (c) Whole chromosome painting in female T. auritus and (d) T. albus (d), using W-chromosome (green) and Z-chromosome (red) probes from T. auritus, providing evidence for the common origin of the ZW sex chromosome system in Triportheus. Bar = 10 μm. Data from [27,42].
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Cioffi, M.d.B.; Yano, C.F.; Sember, A.; Bertollo, L.A.C. Chromosomal Evolution in Lower Vertebrates: Sex Chromosomes in Neotropical Fishes. Genes 2017, 8, 258. https://doi.org/10.3390/genes8100258
AMA Style
Cioffi MdB, Yano CF, Sember A, Bertollo LAC. Chromosomal Evolution in Lower Vertebrates: Sex Chromosomes in Neotropical Fishes. Genes. 2017; 8(10):258. https://doi.org/10.3390/genes8100258
Chicago/Turabian StyleCioffi, Marcelo de Bello, Cassia Fernanda Yano, Alexandr Sember, and Luiz Antônio Carlos Bertollo. 2017. "Chromosomal Evolution in Lower Vertebrates: Sex Chromosomes in Neotropical Fishes" Genes 8, no. 10: 258. https://doi.org/10.3390/genes8100258
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.