
Impact of Repetitive Elements on the Y Chromosome Formation in Plants
by
Roman Hobza
1,2,*, 
Radim Cegan
1,
Wojciech Jesionek
1,
Eduard Kejnovsky
1,
Boris Vyskot
1 and
Zdenek Kubat
1 1
Department of Plant Developmental Genetics, Institute of Biophysics, Academy of Sciences of the Czech Republic, Kralovopolska 135, 61200 Brno, Czech Republic
2
Centre of the Region Hana for Biotechnological and Agricultural Research, Institute of Experimental Botany, 78371 Olomouc, Czech Republic
*
Author to whom correspondence should be addressed.
Genes 2017, 8(11), 302; https://doi.org/10.3390/genes8110302
Submission received: 21 August 2017
/
Revised: 19 October 2017
/
Accepted: 26 October 2017
/
Published: 1 November 2017
(This article belongs to the Special Issue Chromosomal Evolution)
Abstract
:In contrast to animals, separate sexes and sex chromosomes in plants are very rare. Although the evolution of sex chromosomes has been the subject of numerous studies, the impact of repetitive sequences on sex chromosome architecture is not fully understood. New genomic approaches shed light on the role of satellites and transposable elements in the process of Y chromosome evolution. We discuss the impact of repetitive sequences on the structure and dynamics of sex chromosomes with specific focus on Rumex acetosa and Silene latifolia. Recent papers showed that both the expansion and shrinkage of the Y chromosome is influenced by sex-specific regulation of repetitive DNA spread. We present a view that the dynamics of Y chromosome formation is an interplay of genetic and epigenetic processes.
1. Introduction
The evolution of the Y chromosome architecture has historically been of interest mainly at the cytogenetic level, with a recent switch to genomics and epigenomics. The Y chromosome is a unique part of the genome since it does not recombine over some or most of its length and represents male-limited transmission [1]. The main stages of the Y chromosome evolution are manifested by an establishment of the sex-determining region, local suppression of recombination, accumulation of repeats, degeneration of genes, and shrinkage by deletions. Expansion and shrinkage are parallel processes in shaping the Y chromosome structure, varying in the impact on Y chromosome dynamics in individual stages of sex chromosome evolution [2] (Table 1). Although a number of important studies on sex determination and sex chromosome evolution have emerged recently [3,4,5], we focus mainly on the impact of repetitive DNA on Y chromosome dynamics in advanced stages of sex chromosome divergence. Since there is a lack of information in plant models possessing ZZ/ZW [6,7], we review mainly XX/XY chromosomal systems with heteromorphic and/or heterochromatin-containing sex chromosomes.
In plants, sex chromosomes have been identified in 40 species and heteromorphic sex chromosomes have been revealed in just 19 species [8,9]. Even plants with homomorphic sex chromosomes reveal structural changes in non-recombining region(s). In the case of papaya homomorphic sex chromosomes, the accumulation of repeats accounts for much of Y chromosome specific region expansion. Surprisingly, in young papaya Y chromosome (7 million years ago (mya)) at least 20 specific repeats have been identified which are absent from the autosomes and have no homology in other plant species [10]. Dynamic changes of repetitive DNA content can also influence X chromosome evolution, as shown in the case of transposable elements accumulation on the X chromosome in papaya [11]. It is not clear whether the accumulation of retrotransposing sequences on the X chromosome is a result of more efficient selection against this process in autosomes or preferential targeting to the X chromosome [12]. Interestingly, young papaya Y chromosome already reveals signatures of heterochromatization [13].
A global genomic impact of sex chromosome evolution was most studied in species with heteromorphic sex chromosomes, namely in Cannabis sativa (hemp) [14], Hippophae rhamnoides (sea buckthorn) [15], Coccinia grandis [16], and classical models Rumex acetosa (sorrel) and Silene latifolia (white campion) [17]. Surprisingly, even in these plants sex chromosomes are relatively evolutionarily young in comparison with many animal species—e.g., Silene latifolia 6 mya [18], Rumex acetosa 12–13 mya [19], or Coccinia grandis 3 mya [20].
Although none of the plant species possessing heteromorphic sex chromosomes have been sequenced yet, large-scale genome analyses have been accelerated using a combination of next generation sequencing with comparative graph-based clustering analysis of sequenced reads by RepeatExplorer pipeline [21,22]. This approach enables us to characterize the structure of the repetitive fraction of the genome without deep sequencing as well as to determine an accurate assembly of the genome (Figure 1).
In this review, we focus on the main processes that influence the heteromorphic character of sex chromosomes in plants while paying an attention to sex chromosome systems in S. latifolia and R. acetosa that represent the most studied species in this aspect.
2. Promiscuous DNA—Frequent Passengers Colonizing Non-Recombining Regions
Chloroplast and mitochondrial DNA sequences are frequently transferred into the nuclear genome in plants. The endosymbiotic transfer is usually dependent on recombination-based insertions of large fragments of organellar DNA into the nucleus, resulting in either nuclear insertions of plastid DNA (NUPTs) or nuclear insertions of mitochondrial DNA (NUMTs).
In plants with sex chromosomes, the chloroplast DNA has frequently accumulated on the Y chromosomes. This accumulation has been observed on the Y chromosomes of sorrel (XY1Y2 system) and white campion (XY system), being more marked in older and more degenerated Y1 and Y2 chromosomes of sorrel [24,25]. A several-fold enrichment of both chloroplast and mitochondrial sequences was also observed in the male-specific regions of the Y chromosome (MSY) and the hermaphrodite-specific region of the Yh chromosome (HSY) of papaya, a plant with incipient sex chromosomes [26]. NUPTs are more frequent than NUMTs on the Y chromosome in papaya. Similar to other species possessing large Y chromosomes, organellar DNA has also accumulated in C. grandis [16].
3. Satellites and Chromatin Changes
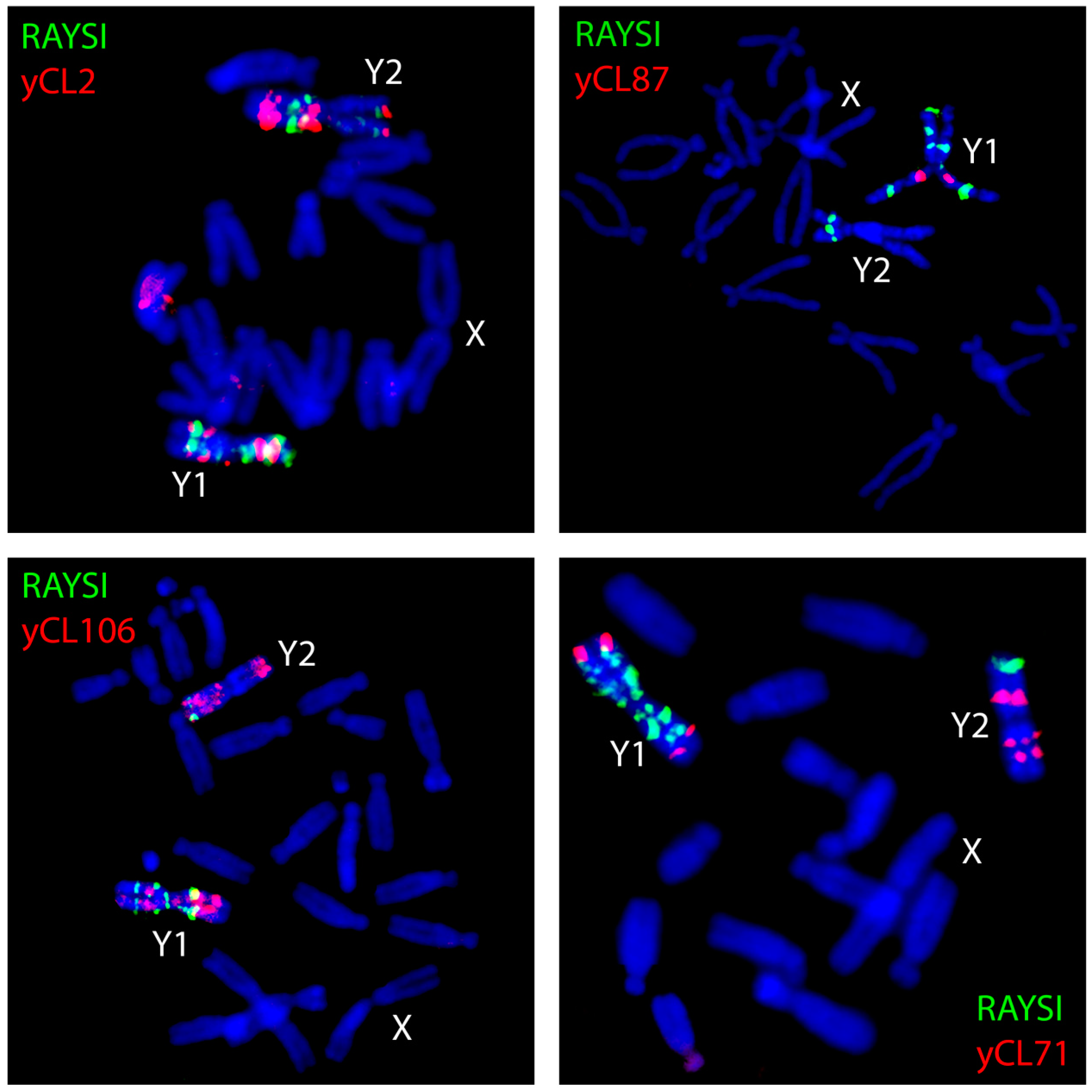
The accumulation of satellites accompanies the evolution of sex chromosomes in some plant species with heteromorphic sex chromosomes. Satellites mostly exhibit discrete chromosomal localization. The most prominent are the heterochromatic regions in R. acetosa (Figure 2) that are formed by various types of tandem repeats [19,27,28,29]. In sorrel, most of the satellites show the Y-specific pattern of distribution, either exhibiting an accumulating on both Y chromosomes like RAYSI and RAYSIII [27,29], or showing the Y chromosome-specific localization such as RAYSII [27,30], indicating that the Y chromosome probably represents a suitable chromosomal context for satellite expansion. It was thought that the heterochromatic nature of Y chromosomes in R. acetosa reflects an advanced degeneration process, but recent results showed that a number of Y-linked genes are transcribed in this species [31]. In white campion, TRAYC and STAR-Y satellites have accumulated on the Y chromosome [32,33]. Surprisingly, these satellite regions do not correlate with heterochromatic regions in this species. This observation suggests that the accumulation of satellites predates chromatin changes in the non-recombining region of the Y chromosome.
Recently, a small Y chromosome containing several satellite DNAs and a large X chromosome resembling a mammalian sex chromosome system (rare in plants) was revealed in sea buckthorn [15]. Some satellites accumulated specifically on individual sex chromosomes (HRTR8 on the X chromosome and HRTR12 on the Y chromosome) and some satellites are shared in both X and Y chromosomes (HRTR2). Generally, the sea buckthorn genome contains an unusually larger number of satellites than the majority of plant genomes [34]. Contrary to sea buckthorn, Coccinia grandis has only one accumulated satellite in its entire genome, without specific pattern on the Y chromosome [16]. Taken together, satellites gather both in heterochromatic and euchromatic parts of sex chromosomes, and it remains a question as to which role they play in the heterochromatinization process.
4. Transposable Elements—Major Force in Sex Chromosome Dynamics
The amplification of Transposable Elements (TEs) is the dominant force increasing the plant genome size. The accumulation of retrotransposons is often responsible for the enormous size of evolutionarily young plant Y chromosomes. Various families of retrotransposons contribute unequally to the size of the X and Y chromosomes in R. acetosa and S. latifolia [18,30,35,36,37], and the proliferation of individual retrotransposons may significantly vary between closely related species with and without sex chromosomes, as was shown in S. latifolia and S. vulgaris [38] as well as in Asparagus [39,40]. Transposable elements thus have a strong influence on the genome architecture of evolving sex chromosomes.
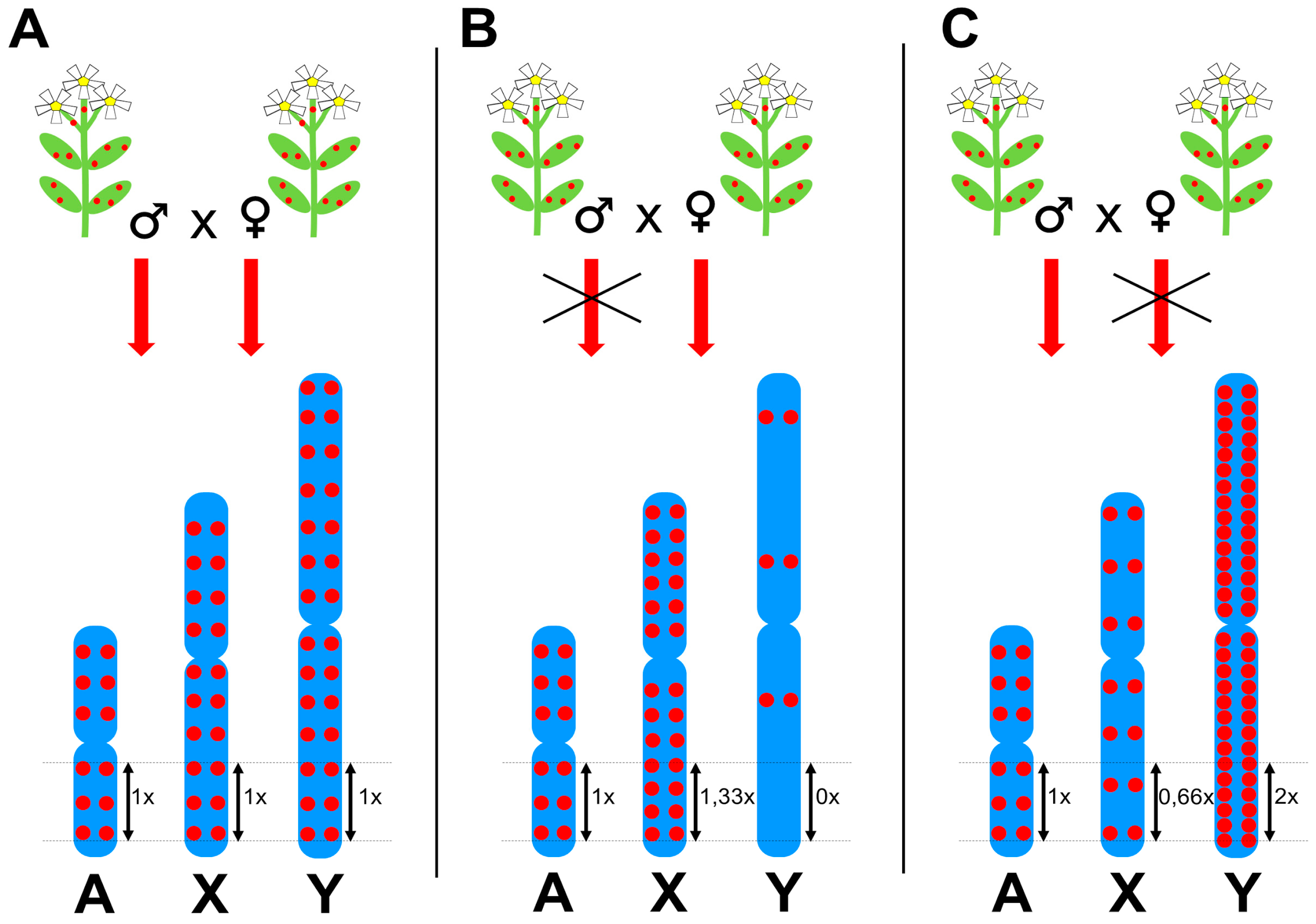
Although some TEs prefer insertion into specific chromosomal regions such as subtelomeres, microsatellite loci [41], other transposons [42], or gene promoters [43], which probably evolved to prevent insertion into protein coding genes, most plant TEs are randomly distributed along chromosomes with different density on sex chromosomes compared to autosomes. This phenomenon is widespread and applies to the majority of highly abundant TEs in S. latifolia and R. acetosa [18,30,35,36,37]. Figure 3 shows a schematic representation of idealized chromosomal distributions of Long Terminal Repeat (LTR) retrotransposons in dioecious plants and a hypothetical causal mechanism; i.e., restricted TE proliferation in either male or female reproductive organs. Three scenarios of TE behavior can be inferred from fluorescence in situ hybridization (FISH) experiments in dioecious plants. (i) The first group of TEs are homogeneously distributed along all chromosomes, thus the average insertion density (number of insertions per unit of length) is identical for all chromosomes (Figure 3A). These TEs proliferate equally in males and females and represent a minority of TEs. (ii) The TEs of the second group are underrepresented on the Y chromosome and simultaneously their insertion density is higher on X chromosomes compared to autosomes (Figure 3B). TEs of this group vary extensively in their effect strength; some TEs are almost completely absent on the Y and strongly accumulated on X chromosomes, while others show weak underrepresentation on the Y chromosome together with a slight accumulation on X chromosomes. Such chromosomal distribution is probably caused by the prevention of TE proliferation in male reproductive cells. Although not always clearly visible in FISH data, this might be the prevalent behavior among highly abundant and transcriptionally active plant TEs, primarily Ty3/gypsy LTR retrotransposons, indicating a strong selection to inhibit most active TEs preferentially in males. (iii) TEs of the third group are accumulated on the Y chromosome and underrepresented on X chromosomes compared to autosomes (Figure 3C). Similarly to the previous case, high variability of their effect strength is observed. This behavior might be caused by TE silencing in female reproductive cells and is frequent among Ty1/copia type LTR retrotransposons, long interspersed nuclear elements (LINEs), and DNA transposons; all TEs with comparatively lower insertional activity.
Diverse TE behavior is apparently a consequence of epigenetic regulation during the development of reproductive cells and embryogenesis [18,44]. Since plants do not set aside germlines early in embryogenesis, reproductive cells differentiate from the meristematic tissues of the flower. Epigenetic marks that contributed to plant development and TE silencing have to be removed to restore totipotent state in the zygote. It is assumed that TEs make use of the opportunity of temporary deficiency in epigenetic control for transposition. However, plants have defensive mechanisms utilizing companion cells of plant gametes. While epigenetic information is gradually lost during gamete formation [45,46,47] and re-established again in the sperm and embryo, companion cells, vegetative cells in pollen and central cells in ovuli, retain low levels of heterochromatic marks, resulting in the active transcription of TEs. TE transcripts are thought to be processed to small RNA molecules that migrate to the sperm and embryo, where they reinforce both transcriptional and post-transcriptional TE silencing [48,49,50]. Epigenetic regulation is in current view a complex array of mutually interconnected pathways, sharing signal molecules such as small RNAs (sRNAs) and long non-coding (lncRNAs) as well as proteins and enzymes [51,52]. Thus, the way of particular TE silencing might be strongly individualized, which results in diverse chromosomal distribution of TEs in dioecious plants. Nevertheless, certain rules can be inferred from previous FISH experiments. The most transpositionally active TEs tend to be more intensively regulated in males, whereas less active TEs are more regulated in females. Although the clear evidence of sex-specific epigenetic regulation of TEs is still unavailable in dioecious plants, in Arabidopsis thaliana, a hermaphroditic model plant species, LTR retrotransposon Evade (EVD) expands if transmitted through the paternal germline but is suppressed when the active EVD element is maternally inherited [44]. It was suggested that EVD is silenced in the maternal sporophytic tissues of the flower by an epigenetic mechanism, most probably sRNA mediated post-transcriptional mRNA degradation. Such regulation of retrotransposon proliferation would result in Y chromosome expansion in the dioecious systems with XX/XY sex determination. The opposite TE behavior, female-specific transposition, has been examined in S. latifolia. LTR retrotransposon Ogre is the main genome size driver in S. latifolia [38] and is virtually absent on the Y chromosome [35,36], which is probably caused by epigenetic silencing in the sperm and embryo [18].
A crucial question resonates here: what is the reason for this apparent sex-specific epigenetic regulation of TEs? (i) Is the diverse epigenetic regulation of TEs a hit-and-miss affair? (ii) Is it an unintentional consequence of different modes of function of epigenetic mechanisms in male and female reproductive organs? (iii) Or a third explanation; is it a result of natural selection that prefers male gametes with the lowest possible number of new TE insertions, i.e. pollen grains that eliminate the most active TEs in order to decrease the risk of deleterious changes in haploid genetic information indispensable for pollen tube growths [53]? Is this a common feature of angiosperms or is it specific to evolutionarily young dioecious species possessing non-recombining Y chromosomes that still carry genes participating in pollen tube growths?
Several recent publications suggest that male reproductive organs adopted unique epigenetic pathways in A. thaliana. These pathways utilize micro RNAs and transfer RNA (tRNAs) for the efficient silencing of TEs in pollen grains [50,54]. Namely, tRNA-derived sRNAs were proved to target mainly Ty3/gypsy LTR retrotransposons, dominant TEs in dioecious plants. Thus, the male germline might possess a reinforced epigenetic barrier against TEs, a barrier that might point to the most active TEs often underrepresented on Y chromosomes in dioecious species. It is difficult to monitor long-term effects in hermaphroditic species without sex-specific DNA regions (sex chromosomes), but dioecious plants provide a plethora of TEs showing signs of sex-specific regulation. Nevertheless, without deep studies of long-term TE behavior in hermaphrodites, we cannot conclude whether those enigmatic distributions of TEs on sex chromosomes are a consequence of common mechanisms in reproductive organs or a consequence of the independence of individual sexes at the genome level that lead to the evolution of specific regulatory mechanisms shaping the architecture of male and female genomes.
5. Unusual DNA Structures on the Y Chromosome and Their Possible Function
Human and chimp Y chromosomes contain large palindromes in their non-recombining region [57,58] containing genes important for sex determination. Palindromes enable intrachromosomal gene conversion and thus help to eliminate deleterious mutations and protect the Y chromosome against degeneration [57,59,60]. Although the large palindromes present on the Y chromosome have different behavior compared to their shorter counterparts, these motifs generally readily form DNA hairpins that belong to non-B DNA conformations together with e.g., triplex and quadruplex DNA. An important role of these structures in the function and evolution of genomes is now only being recognized. For example, it has been suggested that palindromes on the Y chromosome (and on the X chromosome) favor the formation of unusual chromatin structures such as cruciforms [59], and that this conformation regulates the expression of palindrome genes by avoiding transcriptional inactivation [61,62]. Unfortunately, the complete sequence of a plant Y chromosome is not available, yet and the information of whether the palindromes are present also in plant Y chromosomes is still missing.
Recent studies showed that unusual DNA conformations are present not only in genes and their regulatory regions, but also within transposable elements [63,64]. The sequence motifs forming quadruplexes are often located within LTRs of plant LTR retrotransposons, and the higher abundance of quadruplexes within evolutionarily younger retrotransposon suggests their functional role [63,64]. Since these quadruplexes can represent the hot-spots of recombination, it would be interesting to know whether their abundance on the non-recombining Y chromosome is different compared to the rest of the genome.
The activity of LTR retrotransposons on the Y chromosome is regulated by a number of factors—not only by regulatory motifs within the elements (e.g., PBS – primer binding site, PPT – polypurine tract, psi element), but also by epigenetic mechanisms [56,65]—e.g., the Y chromosome of R. acetosa is formed by heterochromatin [66]. Moreover, DNA conformation can also affect TE activity [67], and the secondary DNA structure thus represents another factor playing a role in the life cycle of transposable elements and involves not only transcription and translation but also the reverse transcription and integration phases.
The Y chromosomes of several plant species (S. latifolia, R. acetosa) accumulate microsatellites that are known to readily adopt unusual DNA conformation like e.g., A-DNA or Z-DNA [68,69]. Such DNA with changed conformation is often nucleosome-free [70] and represents a preferred target for transposable elements insertions. Therefore, microsatellites are often more common in the neighborhood of transposable elements [69,71]. Moreover, DNA conformation plays a role also in microsatellite expansion when hairpins are formed during replication slippage, resulting in an increase in the number of monomer units [72].
The unique feature of the Y chromosomes is the reduced rate of recombination, but some recombination processes are still in action there. In this low- or no-recombination context, the strong recombination activity evolved in the form of gene conversion [58], homogenizing the large palindromes on the human and chimp Y chromosomes, and a similar process is expected to be present in the evolutionarily younger plant Y chromosomes. Then, the Y chromosome represents a mosaic of regions shaped by two opposing evolutionary dynamics: (i) the decay of genes and suppressed recombination in X-degenerate regions and (ii) gene acquisitions and the activation of intra-chromosomal recombination in palindromic (ampliconic) regions [59]. Transposable elements and satellites that form a majority of the Y chromosomes are also suitable targets for gene conversion [73,74].
Moreover, TEs and satellites are, thanks to their repetitive nature, good subjects for another recombination process—ectopic recombination. The turnover of transposable elements in plant genomes is much higher than in animals [55]. Ectopic recombination between long terminal repeats of the same retrotransposon results in the deletion of an internal part of LTR retrotransposons, leaving behind “solo LTRs”. Similarly, a region located between two elements can be deleted by ectopic recombination, leading to genome size reduction [75]. Since the recombination is lower on the Y chromosome than on other chromosomes, and the downsizing is less efficient, one can expect a lower proportion of solo LTRs compared to full-length elements on the Y chromosome.
6. Conclusions
Although the advent of genomic methods has shed light on many aspects of heteromorphic sex chromosome formation in dioecious plants, there is still limited information about the impact of structural changes on the function of sex-linked genes. Transposable elements can affect sex chromosome evolution directly via insertion into a specific site, and indirectly by affecting the expression of closely linked genes by epigenetic mechanisms. Large-scale genomic response to repetitive DNA accumulation results in changes in chromatin status, which can in some species lead to heterochromatinization. Is an elevated rate of transposon accumulation the cause or consequence of sex chromosome degeneration? How much does cross-talk of transposable elements with genic regions affect dosage compensation evolution? How much are epigenetic processes involved in the degeneration of sex chromosomes? Surprisingly, structurally divergent sex chromosomes in S. latifolia are euchromatic while papaya homomorphic sex chromosomes reveal clear signs of heterochromatization. It is likely that sex chromosome evolution is affected by a number of mechanisms that vary in individual dioecious species such as population size, genome dynamics, regulation of TEs, etc. It remains to be answered which processes are shared among the species and which mechanisms are unique in individual species. Recent studies clearly show that plants possessing sex chromosomes can regulate the activity of TEs and subsequently regulate their spread in non-recombining regions. Whether this phenomenon is specific for dioecious plants or it is a common attribute of angiosperms remains to be elucidated. It is tempting to speculate that not only RNAi (RNA interference) machinery but also specific DNA conformation such as quadruplexes may play a role in the dynamics of the spread of repetitive elements within sex chromosomes.
Acknowledgments
This work was supported by the Czech Science Foundation (grants 16-08698S to RH and 15-21523Y to ZK).
Conflicts of Interest
The authors declare no conflict of interest.
References
- Bachtrog, D. Y-chromosome evolution: Emerging insights into processes of Y-chromosome degeneration. Nat. Rev. Genet. 2013, 14, 113–124. [Google Scholar] [CrossRef] [PubMed]
- Vyskot, B.; Hobza, R. The genomics of plant sex chromosomes. Plant Sci. 2015, 236, 126–315. [Google Scholar] [CrossRef] [PubMed]
- Akagi, T.; Henry, I.M.; Tao, R.; Comai, L. Plant genetics. A Y-chromosome-encoded small RNA acts as a sex determinant in persimmons. Science 2014, 346, 646–650. [Google Scholar] [CrossRef] [PubMed]
- Mohanty, J.N.; Nayak, S.; Jha, S.; Joshi, R.K. Transcriptome profiling of the floral buds and discovery of genes related to sex-differentiation in the dioecious cucurbit Coccinia grandis (L.) Voigt. Gene 2017, 626, 395–406. [Google Scholar] [CrossRef] [PubMed]
- Murase, K.; Shigenobu, S.; Fujii, S.; Ueda, K.; Murata, T.; Sakamoto, A.; Wada, Y.; Yamaguchi, K.; Osakabe, Y.; Osakabe, K.; et al. MYB transcription factor gene involved in sex determination in Asparagus officinalis. Genes Cells 2017, 22, 115–123. [Google Scholar] [CrossRef] [PubMed]
- Tennessen, J.A.; Govindarajulu, R.; Liston, A.; Ashman, T.-L. Homomorphic ZW chromosomes in a wild strawberry show distinctive recombination heterogeneity but a small sex-determining region. New Phytol. 2016, 211, 1412–1423. [Google Scholar] [CrossRef] [PubMed]
- Slancarova, V.; Zdanska, J.; Janousek, B.; Talianova, M.; Zschach, C.; Zluvova, J.; Siroky, J.; Kovacova, V.; Blavet, H.; Danihelka, J.; et al. Evolution of sex determination systems with heterogametic males and females in silene. Evolution 2013, 67, 3669–3677. [Google Scholar] [CrossRef] [PubMed]
- Ming, R.; Bendahmane, A.; Renner, S.S. Sex chromosomes in land plants. Annu. Rev. Plant Biol. 2011, 62, 485–514. [Google Scholar] [CrossRef] [PubMed]
- Renner, S.S. The relative and absolute frequencies of angiosperm sexual systems: Dioecy, monoecy, gynodioecy, and an updated online database. Am. J. Bot. 2014, 101, 1588–1596. [Google Scholar] [CrossRef] [PubMed]
- Na, J.-K.; Wang, J.; Ming, R. Accumulation of interspersed and sex-specific repeats in the non-recombining region of papaya sex chromosomes. BMC Genomics 2014, 15, 335. [Google Scholar] [CrossRef] [PubMed]
- Gschwend, A.R.; Yu, Q.; Tong, E.J.; Zeng, F.; Han, J.; VanBuren, R.; Aryal, R.; Charlesworth, D.; Moore, P.H.; Paterson, A.H.; et al. Rapid divergence and expansion of the X chromosome in papaya. Proc. Natl. Acad. Sci. USA 2012, 109, 13716–13721. [Google Scholar] [CrossRef] [PubMed]
- Kejnovsky, E.; Hobza, R.; Cermak, T.; Kubat, Z.; Vyskot, B. The role of repetitive DNA in structure and evolution of sex chromosomes in plants. Heredity 2009, 102, 533–541. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Wang, X.; Yu, Q.; Ming, R.; Jiang, J. DNA methylation and heterochromatinization in the male-specific region of the primitive Y chromosome of papaya. Genome Res. 2008, 18, 1938–1943. [Google Scholar] [CrossRef] [PubMed]
- Sakamoto, K.; Ohmido, N.; Fukui, K.; Kamada, H.; Satoh, S. Site-specific accumulation of a line-like retrotransposon in a sex chromosome of the dioecious plant Cannabis sativa. Plant Mol. Biol. 2000, 44, 723–732. [Google Scholar] [CrossRef] [PubMed]
- Puterova, J.; Razumova, O.; Martinek, T.; Alexandrov, O.; Divashuk, M.; Kubat, Z.; Hobza, R.; Karlov, G.; Kejnovsky, E. Satellite DNA and Transposable Elements in Seabuckthorn (Hippophae rhamnoides), a Dioecious Plant with Small Y and Large X Chromosomes. Genome Biol. Evol. 2017, 9, 197–212. [Google Scholar] [CrossRef] [PubMed]
- Sousa, A.; Bellot, S.; Fuchs, J.; Houben, A.; Renner, S.S. Analysis of transposable elements and organellar DNA in male and female genomes of a species with a huge Y chromosome reveals distinct Y centromeres. Plant J. 2016, 88, 387–396. [Google Scholar] [CrossRef] [PubMed]
- Vyskot, B.; Hobza, R. Gender in plants: Sex chromosomes are emerging from the fog. Trends Genet. 2004, 20, 432–438. [Google Scholar] [CrossRef] [PubMed]
- Kubat, Z.; Zluvova, J.; Vogel, I.; Kovacova, V.; Cermak, T.; Cegan, R.; Hobza, R.; Vyskot, B.; Kejnovsky, E. Possible mechanisms responsible for absence of a retrotransposon family on a plant Y chromosome. New Phytol. 2014, 202, 662–678. [Google Scholar] [CrossRef] [PubMed]
- Navajas-Pérez, R.; de la Herrán, R.; López González, G.; Jamilena, M.; Lozano, R.; Ruiz Rejón, C.; Ruiz Rejón, M.; Garrido-Ramos, M.A. The evolution of reproductive systems and sex-determining mechanisms within rumex (polygonaceae) inferred from nuclear and chloroplastidial sequence data. Mol. Biol. Evol. 2005, 22, 1929–1939. [Google Scholar] [CrossRef] [PubMed]
- Sousa, A.; Fuchs, J.; Renner, S.S. Molecular cytogenetics (FISH, GISH) of Coccinia grandis: A ca. 3 myr-old species of cucurbitaceae with the largest Y/autosome divergence in flowering plants. Cytogenet. Genome Res. 2013, 139, 107–118. [Google Scholar] [CrossRef] [PubMed]
- Novák, P.; Neumann, P.; Macas, J. Graph-based clustering and characterization of repetitive sequences in next-generation sequencing data. BMC Bioinformatics 2010, 11, 378. [Google Scholar] [CrossRef] [PubMed]
- Novak, P.; Neumann, P.; Pech, J.; Steinhaisl, J.; Macas, J. RepeatExplorer: A Galaxy-based web server for genome-wide characterization of eukaryotic repetitive elements from next-generation sequence reads. Bioinformatics 2013, 29, 792–793. [Google Scholar] [CrossRef] [PubMed]
- Novák, P.; Ávila Robledillo, L.; Koblížková, A.; Vrbová, I.; Neumann, P.; Macas, J. TAREAN: A computational tool for identification and characterization of satellite DNA from unassembled short reads. Nucleic Acids Res. 2017, 45, e111. [Google Scholar] [CrossRef] [PubMed]
- Kejnovsky, E.; Kubat, Z.; Hobza, R.; Lengerova, M.; Sato, S.; Tabata, S.; Fukui, K.; Matsunaga, S.; Vyskot, B. Accumulation of chloroplast DNA sequences on the Y chromosome of Silene latifolia. Genetica 2006, 128, 167–175. [Google Scholar] [CrossRef] [PubMed]
- Steflova, P.; Hobza, R.; Vyskot, B.; Kejnovsky, E. Strong accumulation of chloroplast DNA in the Y chromosomes of Rumex acetosa and Silene latifolia. Cytogenet. Genome Res. 2014, 142, 59–65. [Google Scholar] [CrossRef] [PubMed]
- VanBuren, R.; Ming, R. Organelle DNA accumulation in the recently evolved papaya sex chromosomes. Mol. Genet. Genomics 2013, 288, 277–284. [Google Scholar] [CrossRef] [PubMed]
- Shibata, F.; Hizume, M.; Kuroki, Y. Chromosome painting of Y chromosomes and isolation of a Y chromosome-specific repetitive sequence in the dioecious plant Rumex acetosa. Chromosoma 1999, 108, 266–270. [Google Scholar] [CrossRef] [PubMed]
- Shibata, F.; Hizume, M.; Kuroki, Y. Differentiation and the polymorphic nature of the Y chromosomes revealed by repetitive sequences in the dioecious plant, Rumex acetosa. Chromosome Res. 2000, 8, 229–236. [Google Scholar] [CrossRef] [PubMed]
- Mariotti, B.; Manzano, S.; Kejnovský, E.; Vyskot, B.; Jamilena, M. Accumulation of Y-specific satellite DNAs during the evolution of Rumex acetosa sex chromosomes. Mol. Genet. Genomics 2009, 281, 249–259. [Google Scholar] [CrossRef] [PubMed]
- Steflova, P.; Tokan, V.; Vogel, I.; Lexa, M.; Macas, J.; Novak, P.; Hobza, R.; Vyskot, B.; Kejnovsky, E. Contrasting patterns of transposable element and satellite distribution on sex chromosomes (XY1Y2) in the dioecious plant Rumex acetosa. Genome Biol. Evol. 2013, 5, 769–782. [Google Scholar] [CrossRef] [PubMed]
- Michalovova, M.; Kubat, Z.; Hobza, R.; Vyskot, B.; Kejnovsky, E. Fully automated pipeline for detection of sex linked genes using RNA-Seq data. BMC Bioinformatics 2015, 16, 78. [Google Scholar] [CrossRef] [PubMed]
- Hobza, R.; Lengerova, M.; Svoboda, J.; Kubekova, H.; Kejnovsky, E.; Vyskot, B. An accumulation of tandem DNA repeats on the Y chromosome in Silene latifolia during early stages of sex chromosome evolution. Chromosoma 2006, 115, 376–382. [Google Scholar] [CrossRef] [PubMed]
- Hobza, R.; Kejnovsky, E.; Vyskot, B.; Widmer, A. The role of chromosomal rearrangements in the evolution of Silene latifolia sex chromosomes. Mol. Genet. Genomics 2007, 278, 633–638. [Google Scholar] [CrossRef] [PubMed]
- Mehrotra, S.; Goyal, V. Repetitive Sequences in Plant Nuclear DNA: Types, Distribution, Evolution and Function. Genom. Proteom. Bioinform. 2014, 12, 164–171. [Google Scholar] [CrossRef] [PubMed]
- Cermak, T.; Kubat, Z.; Hobza, R.; Koblizkova, A.; Widmer, A.; Macas, J.; Vyskot, B.; Kejnovsky, E. Survey of repetitive sequences in Silene latifolia with respect to their distribution on sex chromosomes. Chromosom. Res. 2008, 16, 961–976. [Google Scholar] [CrossRef] [PubMed]
- Filatov, D.A.; Howell, E.C.; Groutides, C.; Armstrong, S.J. Recent spread of a retrotransposon in the Silene latifolia genome, apart from the Y chromosome. Genetics 2009, 181, 811–817. [Google Scholar] [CrossRef] [PubMed]
- Kralova, T.; Cegan, R.; Kubat, Z.; Vrana, J.; Vyskot, B.; Vogel, I.; Kejnovsky, E.; Hobza, R. Identification of a novel retrotransposon with sex chromosome-specific distribution in Silene latifolia. Cytogenet. Genome Res. 2014, 143, 87–95. [Google Scholar] [CrossRef] [PubMed]
- Cegan, R.; Vyskot, B.; Kejnovsky, E.; Kubat, Z.; Blavet, H.; Šafář, J.; Doležel, J.; Blavet, N.; Hobza, R. Genomic diversity in two related plant species with and without sex chromosomes—Silene latifolia and S. vulgaris. PLoS ONE 2012, 7, e31898. [Google Scholar] [CrossRef] [PubMed]
- Kuhl, J.C.; Havey, M.J.; Martin, W.J.; Cheung, F.; Yuan, Q.; Landherr, L.; Hu, Y.; Leebens-Mack, J.; Town, C.D.; Sink, K.C. Comparative genomic analyses in Asparagus. Genome 2005, 48, 1052–1060. [Google Scholar] [CrossRef] [PubMed]
- Harkess, A.; Mercati, F.; Abbate, L.; McKain, M.; Pires, J.C.; Sala, T.; Sunseri, F.; Falavigna, A.; Leebens-Mack, J. Retrotransposon proliferation coincident with the evolution of dioecy in Asparagus. G3 Genes, Genomes, Genet. 2016, 6, 2679–2685. [Google Scholar] [CrossRef] [PubMed]
- Akagi, H.; Yokozeki, Y.; Inagaki, A.; Mori, K.; Fujimura, T. Micron, a microsatellite-targeting transposable element in the rice genome. Mol. Genet. Genomics 2001, 266, 471–480. [Google Scholar] [CrossRef] [PubMed]
- Jiang, N.; Wessler, S.R. Insertion preference of maize and rice miniature inverted repeat transposable elements as revealed by the analysis of nested elements. Plant Cell 2001, 13, 2553–2564. [Google Scholar] [CrossRef] [PubMed]
- Naito, K.; Monden, Y.; Yasuda, K.; Saito, H.; Okumoto, Y. mPing: The bursting transposon. Breed. Sci. 2014, 64, 109–114. [Google Scholar] [CrossRef] [PubMed]
- Reinders, J.; Mirouze, M.; Nicolet, J.; Paszkowski, J. Parent-of-origin control of transgenerational retrotransposon proliferation in Arabidopsis. EMBO Rep 2013, 14, 823–828. [Google Scholar] [CrossRef] [PubMed]
- Gehring, M.; Henikoff, S. DNA methylation dynamics in plant genomes. Biochim. Biophys. Acta BBA Gene Struct. Expr. 2002, 1769, 276–286. [Google Scholar] [CrossRef] [PubMed]
- Hsieh, T.-F.; Ibarra, C.A.; Silva, P.; Zemach, A.; Eshed-Williams, L.; Fischer, R.L.; Zilberman, D. Genome-wide demethylation of Arabidopsis endosperm. Science 2009, 324, 1451–1454. [Google Scholar] [CrossRef] [PubMed]
- Calarco, J.P.; Borges, F.; Donoghue, M.T.A.; Van Ex, F.; Jullien, P.E.; Lopes, T.; Gardner, R.; Berger, F.; Feijó, J.A.; Becker, J.D.; et al. Reprogramming of DNA methylation in pollen guides epigenetic inheritance via small RNA. Cell 2012, 151, 194–205. [Google Scholar] [CrossRef] [PubMed]
- Slotkin, R.K.; Vaughn, M.; Borges, F.; Tanurdžić, M.; Becker, J.D.; Feijó, J.A.; Martienssen, R.A. Epigenetic Reprogramming and Small RNA Silencing of Transposable Elements in Pollen. Cell 2009, 136, 461–472. [Google Scholar] [CrossRef] [PubMed]
- Ibarra, C.A.; Feng, X.; Schoft, V.K.; Hsieh, T.-F.; Uzawa, R.; Rodrigues, J.A.; Zemach, A.; Chumak, N.; Machlicova, A.; Nishimura, T.; et al. Active DNA Demethylation in Plant Companion Cells Reinforces Transposon Methylation in Gametes. Science 2012, 337, 1360–1364. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martínez, G.; Panda, K.; Köhler, C.; Slotkin, R.K. Silencing in sperm cells is directed by RNA movement from the surrounding nurse cell. Nat. Plants 2016, 2, 16030. [Google Scholar] [CrossRef] [PubMed]
- Fultz, D.; Choudury, S.G.; Slotkin, R.K. Silencing of active transposable elements in plants. Curr. Opin. Plant Biol. 2015, 27, 67–76. [Google Scholar] [CrossRef] [PubMed]
- Cuerda-Gil, D.; Slotkin, R.K. Non-canonical RNA-directed DNA methylation. Nat. Plants 2016, 2, 16163. [Google Scholar] [CrossRef] [PubMed]
- Chibalina, M.V.; Filatov, D.A. Plant Y chromosome degeneration is retarded by haploid purifying selection. Curr. Biol. 2011, 21, 1475–1479. [Google Scholar] [CrossRef] [PubMed]
- Creasey, K.M.; Zhai, J.; Borges, F.; Van Ex, F.; Regulski, M.; Meyers, B.C.; Martienssen, R.A. miRNAs trigger widespread epigenetically activated siRNAs from transposons in Arabidopsis. Nature 2014, 508, 411–415. [Google Scholar] [CrossRef] [PubMed]
- Kejnovsky, E.; Leitch, I.J.; Leitch, A.R. Contrasting evolutionary dynamics between angiosperm and mammalian genomes. Trends Ecol. Evol. 2009, 24, 572–582. [Google Scholar] [CrossRef] [PubMed]
- Ito, H. Small RNAs and transposon silencing in plants. Dev. Growth Differ. 2012, 54, 100–107. [Google Scholar] [CrossRef] [PubMed]
- Rozen, S.; Skaletsky, H.; Marszalek, J.D.; Minx, P.J.; Cordum, H.S.; Waterston, R.H.; Wilson, R.K.; Page, D.C. Abundant gene conversion between arms of palindromes in human and ape Y chromosomes. Nature 2003, 423, 873–876. [Google Scholar] [CrossRef] [PubMed]
- Trombetta, B.; Cruciani, F. Y chromosome palindromes and gene conversion. Hum. Genet. 2017, 136, 605–619. [Google Scholar] [CrossRef] [PubMed]
- Skaletsky, H.; Kuroda-Kawaguchi, T.; Minx, P.J.; Cordum, H.S.; Hillier, L.; Brown, L.G.; Repping, S.; Pyntikova, T.; Ali, J.; Bieri, T.; et al. The male-specific region of the human Y chromosome is a mosaic of discrete sequence classes. Nature 2003, 423, 825–837. [Google Scholar] [CrossRef] [PubMed]
- Betrán, E.; Demuth, J.P.; Williford, A. Why chromosome palindromes? Int. J. Evol. Biol. 2012, 2012, 207958. [Google Scholar] [CrossRef] [PubMed]
- Warburton, P.E.; Giordano, J.; Cheung, F.; Gelfand, Y.; Benson, G. Inverted repeat structure of the human genome: The X-chromosome contains a preponderance of large, highly homologous inverted repeats that contain testes genes. Genome Res. 2004, 14, 1861–1869. [Google Scholar] [CrossRef] [PubMed]
- De Vries, M.; Vosters, S.; Merkx, G.; D’Hauwers, K.; Wansink, D.G.; Ramos, L.; de Boer, P. Human male meiotic sex chromosome inactivation. PLoS ONE 2012, 7, e31485. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lexa, M.; Kejnovský, E.; Steflová, P.; Konvalinová, H.; Vorlícková, M.; Vyskot, B. Quadruplex-forming sequences occupy discrete regions inside plant LTR retrotransposons. Nucleic Acids Res. 2014, 42, 968–978. [Google Scholar] [CrossRef] [PubMed]
- Lexa, M.; Steflova, P.; Martinek, T.; Vorlickova, M.; Vyskot, B.; Kejnovsky, E. Guanine quadruplexes are formed by specific regions of human transposable elements. BMC Genomics 2014, 15, 1032. [Google Scholar] [CrossRef] [PubMed]
- Lisch, D. Epigenetic Regulation of Transposable Elements in Plants. Annu. Rev. Plant Biol. 2009, 60, 43–66. [Google Scholar] [CrossRef] [PubMed]
- Lengerova, M.; Vyskot, B. Sex chromatin and nucleolar analyses in Rumex acetosa L. Protoplasma 2001, 217, 147–153. [Google Scholar] [CrossRef] [PubMed]
- Sahakyan, A.B.; Murat, P.; Mayer, C.; Balasubramanian, S. G-quadruplex structures within the 3′ UTR of LINE-1 elements stimulate retrotransposition. Nat. Struct. Mol. Biol. 2017, 24, 243–247. [Google Scholar] [CrossRef] [PubMed]
- Kubat, Z.; Hobza, R.; Vyskot, B.; Kejnovsky, E. Microsatellite accumulation on the Y chromosome in Silene latifolia. Genome 2008, 51, 350–356. [Google Scholar] [CrossRef] [PubMed]
- Kejnovský, E.; Michalovova, M.; Steflova, P.; Kejnovska, I.; Manzano, S.; Hobza, R.; Kubat, Z.; Kovarik, J.; Jamilena, M.; Vyskot, B. Expansion of Microsatellites on Evolutionary Young Y Chromosome. PLoS ONE 2013, 8, e45519. [Google Scholar] [CrossRef] [PubMed]
- Huppert, J.L.; Balasubramanian, S. G-quadruplexes in promoters throughout the human genome. Nucleic Acids Res. 2007, 35, 406–413. [Google Scholar] [CrossRef] [PubMed]
- Nadir, E.; Margalit, H.; Gallily, T.; Ben-Sasson, S.A. Microsatellite spreading in the human genome: Evolutionary mechanisms and structural implications. Proc. Natl. Acad. Sci. USA 1996, 93, 6470–6475. [Google Scholar] [CrossRef] [PubMed]
- Ellegren, H. Microsatellites: Simple sequences with complex evolution. Nat. Rev. Genet. 2004, 5, 435–445. [Google Scholar] [CrossRef] [PubMed]
- Kass, D.H.; Batzer, M.A.; Deininger, P.L. Gene conversion as a secondary mechanism of short interspersed element (SINE) evolution. Mol. Cell. Biol. 1995, 15, 19–25. [Google Scholar] [CrossRef] [PubMed]
- Krzywinski, J.; Sangaré, D.; Besansky, N.J. Satellite DNA from the Y chromosome of the malaria vector Anopheles gambiae. Genetics 2005, 169, 185–196. [Google Scholar] [CrossRef] [PubMed]
- Hawkins, J.S.; Proulx, S.R.; Rapp, R.A.; Wendel, J.F. Rapid DNA loss as a counterbalance to genome expansion through retrotransposon proliferation in plants. Proc. Natl. Acad. Sci. USA 2009, 106, 17811–17816. [Google Scholar] [CrossRef] [PubMed]
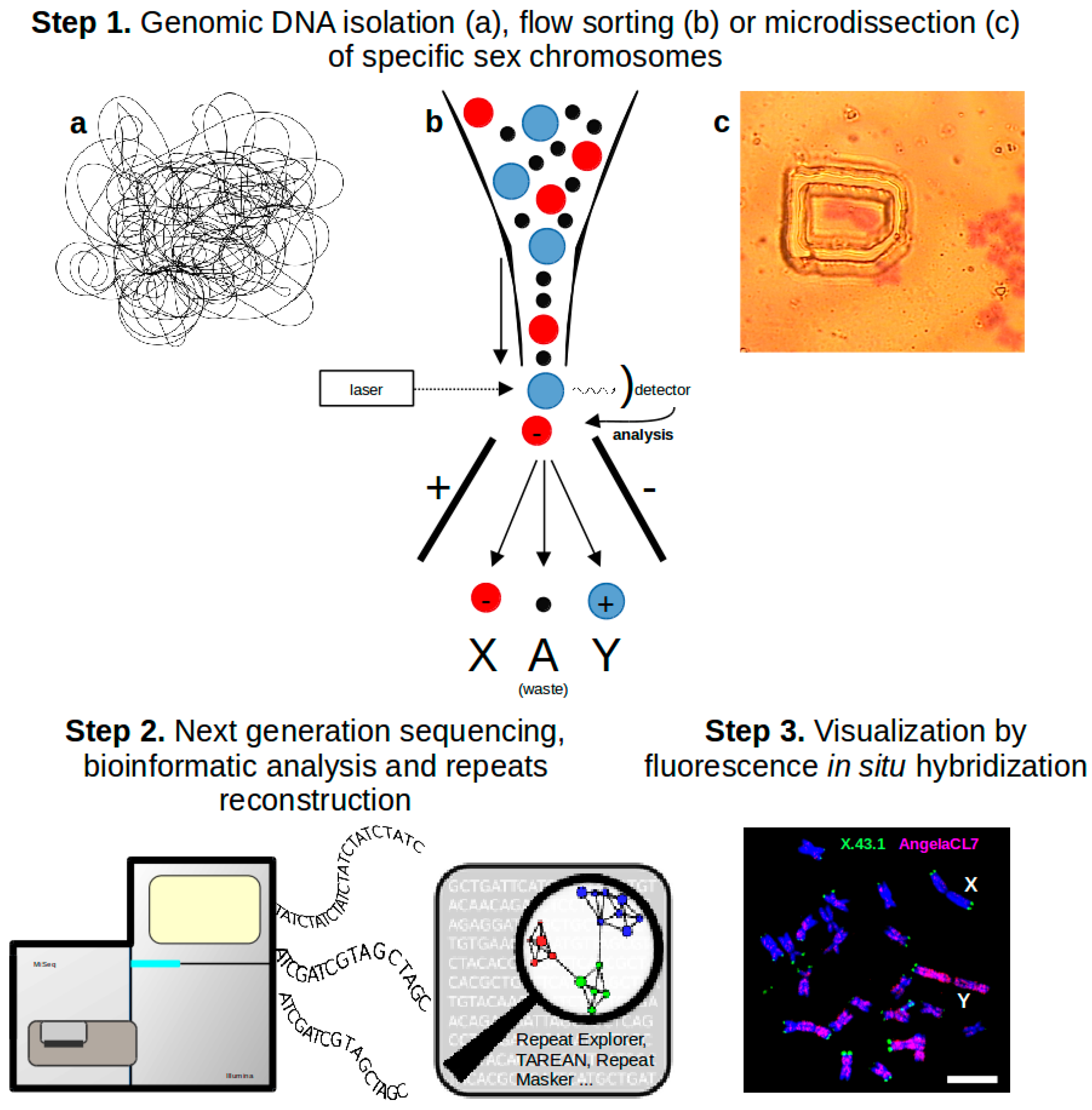
Figure 1.
Typical workflow used for repeats reconstruction and characterization. Step 1—Genomic DNA for subsequent analysis can be isolated by three different approaches. Flow sorting and microdissection in our case are usually used for sex chromosomes separation. Step 2—Genomic DNA or separated sex chromosomes are sequenced with low coverage. Repeats are reconstructed and characterized by the clustering algorithm employed in the RepeatExplorer pipeline [22] or by the TAREAN tool [23]. Step 3—Reconstructed repeats are used as probes for Fluorescence In Situ Hybridization (FISH) localization on Silene latifolia metaphase chromosomes. The X and Y chromosomes are indicated, bar indicates 10 μm. Red probe illuminates the Y-biased repetitive element (Angela CL7), green probe represents the internal FISH control (subtelomeric tandem repeat X43.1).
Figure 1.
Typical workflow used for repeats reconstruction and characterization. Step 1—Genomic DNA for subsequent analysis can be isolated by three different approaches. Flow sorting and microdissection in our case are usually used for sex chromosomes separation. Step 2—Genomic DNA or separated sex chromosomes are sequenced with low coverage. Repeats are reconstructed and characterized by the clustering algorithm employed in the RepeatExplorer pipeline [22] or by the TAREAN tool [23]. Step 3—Reconstructed repeats are used as probes for Fluorescence In Situ Hybridization (FISH) localization on Silene latifolia metaphase chromosomes. The X and Y chromosomes are indicated, bar indicates 10 μm. Red probe illuminates the Y-biased repetitive element (Angela CL7), green probe represents the internal FISH control (subtelomeric tandem repeat X43.1).
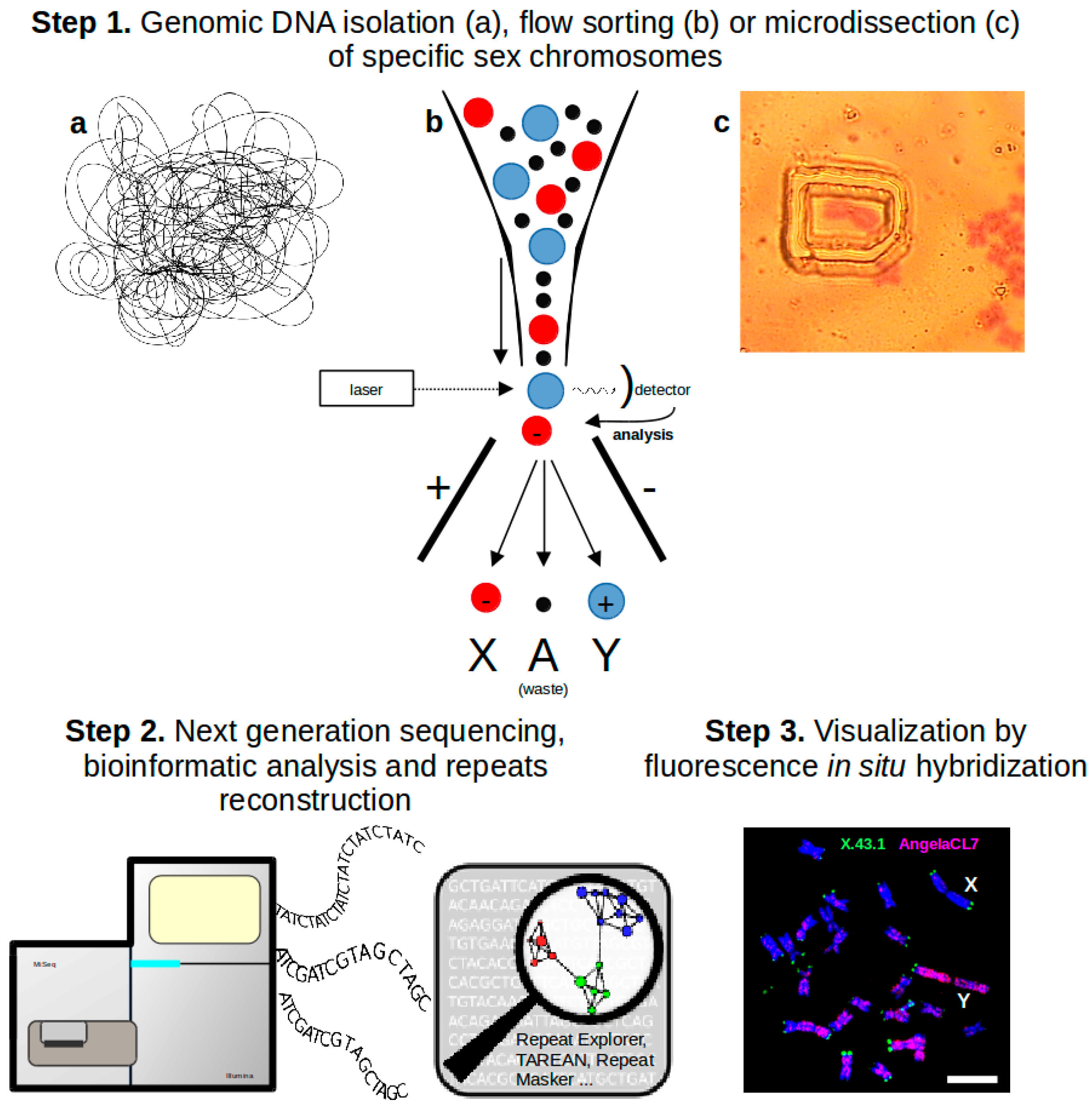
Figure 2.
Localization of satellites on metaphase chromosomes of Rumex acetosa using FISH. The name of the satellite or number of satellite cluster is inside each figure.
Figure 2.
Localization of satellites on metaphase chromosomes of Rumex acetosa using FISH. The name of the satellite or number of satellite cluster is inside each figure.
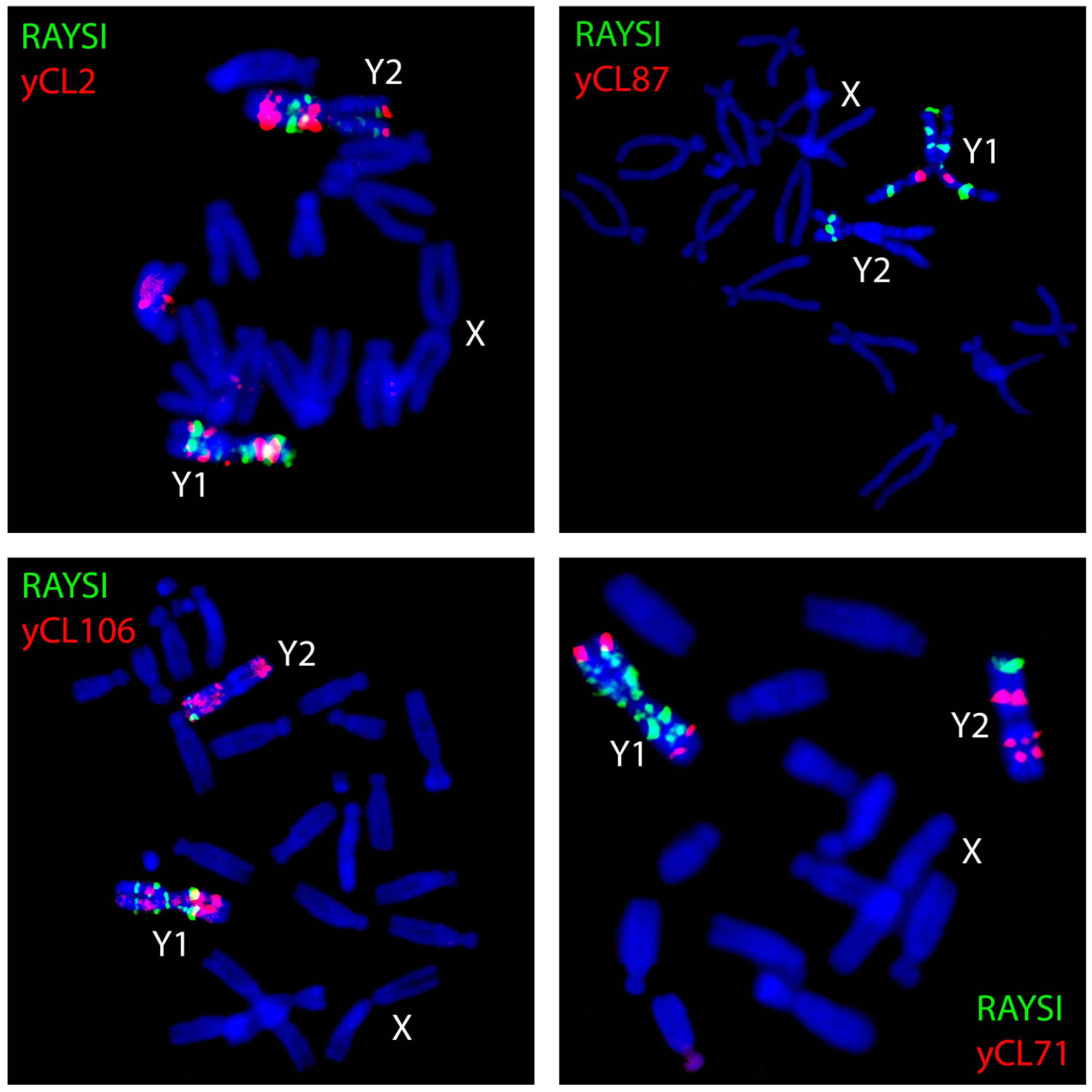
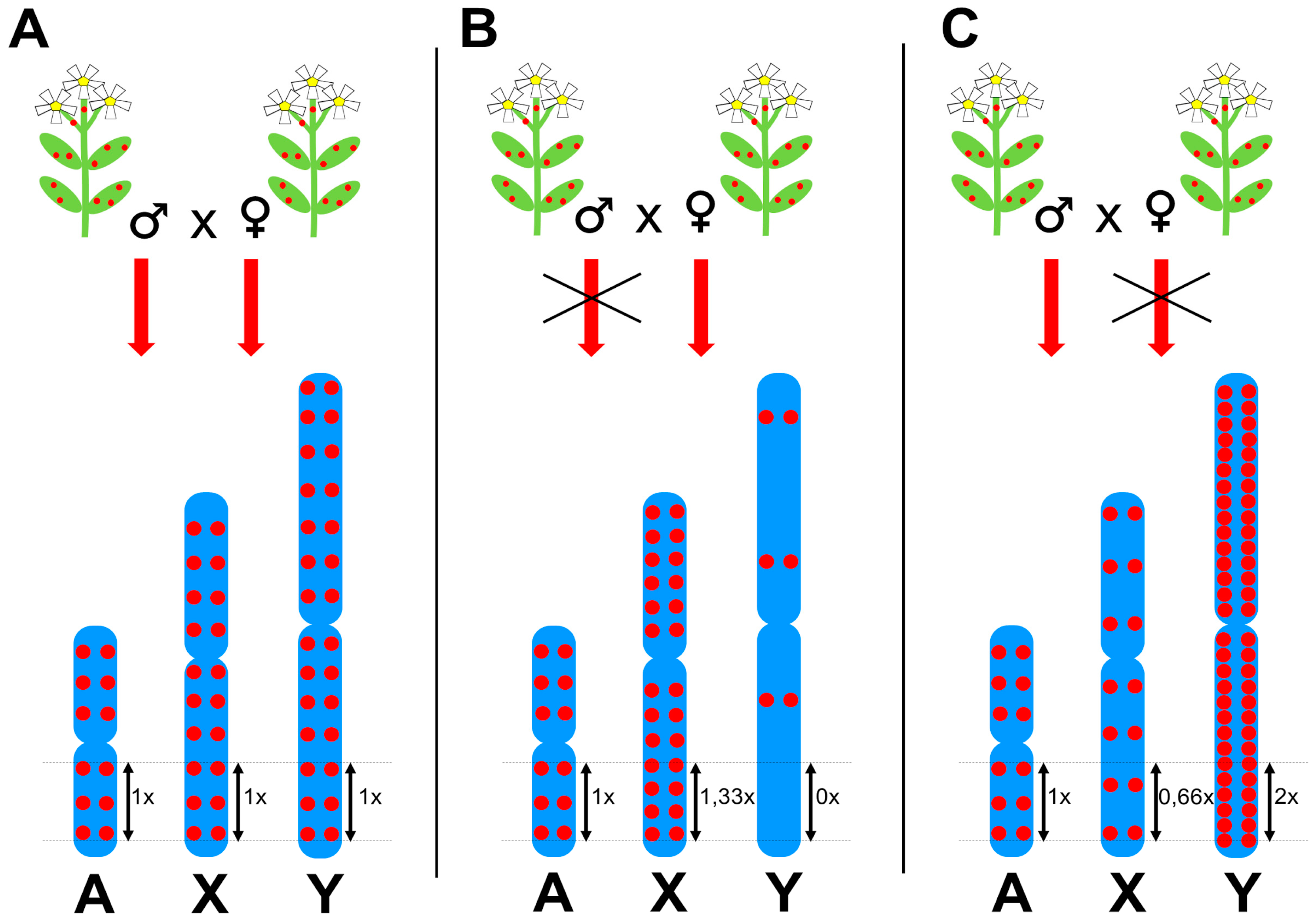
Figure 3.
Three scenarios of transgenerational proliferation of transposable elements (TEs) in plants and their impact on the chromosomal distribution of a TE. Red dots indicate TE insertions—in blue-colored meiotic chromosomes, A-autosomes, X-chromosomes, and Y-chromosomes. Numbers next to double arrows indicate the expected density of TE insertions per one unit of length of a respective chromosome. (A) If a TE is passed down to offspring equally in males and females, TE insertion density is identical on all chromosomes. (B) If TE proliferation is disrupted in males, TE density is 1,33 times higher on the X chromosome than on autosomes, but nearly zero on the Y chromosome. (C) If TE proliferation is disrupted in females, TE insertion density is lower on the X chromosome compared to autosomes and twice as high on the Y chromosome than on autosomes. These three scenarios represent extreme cases of sex-specific TE activity. Real world TEs range from TEs that are almost fully sex-specifically inheritable to TEs with only slight sex-dependent inheritance.
Figure 3.
Three scenarios of transgenerational proliferation of transposable elements (TEs) in plants and their impact on the chromosomal distribution of a TE. Red dots indicate TE insertions—in blue-colored meiotic chromosomes, A-autosomes, X-chromosomes, and Y-chromosomes. Numbers next to double arrows indicate the expected density of TE insertions per one unit of length of a respective chromosome. (A) If a TE is passed down to offspring equally in males and females, TE insertion density is identical on all chromosomes. (B) If TE proliferation is disrupted in males, TE density is 1,33 times higher on the X chromosome than on autosomes, but nearly zero on the Y chromosome. (C) If TE proliferation is disrupted in females, TE insertion density is lower on the X chromosome compared to autosomes and twice as high on the Y chromosome than on autosomes. These three scenarios represent extreme cases of sex-specific TE activity. Real world TEs range from TEs that are almost fully sex-specifically inheritable to TEs with only slight sex-dependent inheritance.

{kind=link}
{kind=link}
{kind=link}
Table 1.
Summary of processes forming sex chromosome structure in the expansion and shrinkage phases.
Table 1.
Summary of processes forming sex chromosome structure in the expansion and shrinkage phases.
| Mechanism | References |
|---|---|
| Expansion phase | |
| Satellites expansion | Sousa et al. (2016) [16], Shibata et al. (1999) [26], |
| Kubat et al. (2008) [18], Puterova et al. (2017) [15], | |
| Retrotranspositions | Na et al. (2014) [10], Cermak et al. (2008) [34], |
| Sousa et al. (2016) [16] | |
| Promiscuous DNA insertions | VanBuren et al. (2013) [25], Kejnovsky et al. (2006) [23] |
| Shrinkage phase | |
| Ectopic recombination | Kejnovsky et al. (2009) [12] |
| Deletions | Hawkins et al. (2009) [55] |
| Both expansion and shrinkage phase | |
| Epigenetic regulation of TEs | Kubat et al. (2014) [18] |
| Chromatin modification | Zhang et al. (2008) [13], Lengerova et al. (2001) [56] |
TE: Transposable element.
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Hobza, R.; Cegan, R.; Jesionek, W.; Kejnovsky, E.; Vyskot, B.; Kubat, Z. Impact of Repetitive Elements on the Y Chromosome Formation in Plants. Genes 2017, 8, 302. https://doi.org/10.3390/genes8110302
AMA Style
Hobza R, Cegan R, Jesionek W, Kejnovsky E, Vyskot B, Kubat Z. Impact of Repetitive Elements on the Y Chromosome Formation in Plants. Genes. 2017; 8(11):302. https://doi.org/10.3390/genes8110302
Chicago/Turabian StyleHobza, Roman, Radim Cegan, Wojciech Jesionek, Eduard Kejnovsky, Boris Vyskot, and Zdenek Kubat. 2017. "Impact of Repetitive Elements on the Y Chromosome Formation in Plants" Genes 8, no. 11: 302. https://doi.org/10.3390/genes8110302
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.