Amphibian and Avian Karyotype Evolution: Insights from Lampbrush Chromosome Studies

Saint-Petersburg State University, Saint-Petersburg 199034, Russia
*
Author to whom correspondence should be addressed.
†
These authors contributed equally to this work.
Genes 2017, 8(11), 311; https://doi.org/10.3390/genes8110311
Submission received: 15 September 2017
/
Revised: 29 October 2017
/
Accepted: 31 October 2017
/
Published: 8 November 2017
(This article belongs to the Special Issue Chromosomal Evolution)
{kind=link}
{kind=link}
{kind=link}
Abstract
:Amphibian and bird karyotypes typically have a complex organization, which makes them difficult for standard cytogenetic analysis. That is, amphibian chromosomes are generally large, enriched with repetitive elements, and characterized by the absence of informative banding patterns. The majority of avian karyotypes comprise a small number of relatively large macrochromosomes and numerous tiny morphologically undistinguishable microchromosomes. A good progress in investigation of amphibian and avian chromosome evolution became possible with the usage of giant lampbrush chromosomes typical for growing oocytes. Due to the giant size, peculiarities of organization and enrichment with cytological markers, lampbrush chromosomes can serve as an opportune model for comprehensive high-resolution cytogenetic and cytological investigations. Here, we review the main findings on chromosome evolution in amphibians and birds that were obtained using lampbrush chromosomes. In particular, we discuss the data on evolutionary chromosomal rearrangements, accumulation of polymorphisms, evolution of sex chromosomes as well as chromosomal changes during clonal reproduction of interspecies hybrids.
1. Peculiarities of Amphibian and Avian Genomes and Karyotypes
In general, amphibian and avian species are characterized by specific and complexly organized genomes. In particular, the size of amphibian genomes exhibits the greatest variety among vertebrates: in two orders, Anurans and Apoda, genome size ranges from 0.95 to 16 pg/N, while in Urodeles DNA value is extremely high and varies from 13.5 to 150 pg/N [1]. Such large genomes are enriched with repetitive sequences [2,3]. Moreover, certain Urodeles exhibit the longest intron length in comparison to other vertebrates [4]. Currently, only three amphibian genomes have been sequenced, Xenopus tropicalis, Xenopus laevis and Nanorana parkeri genomes, and all of them reveal quite small or average size [5,6,7].
Amphibian metaphase chromosomes are frequently characterized by the absence of informative banding patterns [8,9]. The number of chromosomes in diploid sets is generally low and usually does not exceed 26. Several primitive species of Urodels, Anurans and Apoda possess higher number of chromosomes sometimes including microchromosomes [10,11]. Similarities in sex chromosomes in the majority of amphibian species also complicate the karyotype analysis [12,13,14]. Furthermore, amphibians tend to hybridize and form viable and fertile interspecies hybrids that often become polyploid [15,16]. It makes investigation of amphibian genomes even more complex.
Birds have relatively small genomes with a diploid chromosome number being about 80 [17,18,19,20,21,22,23]. Typical avian karyotype comprises several pairs of relatively large macrochromosomes and numerous tiny morphologically undistinguishable microchromosomes. Notably, the microchromosomes represent a gene-dense part of the karyotype and possess about 50% of the genes [20,21,24,25]. At the same time, due to their size and DNA composition the cytogenetic and genomic data on microchromosomes remain quite limited. Currently, the most investigated avian karyotypes belong to representatives of the order Galliformes (including chicken, quails, turkey, paves, partridges, and pheasants), mainly due to their agricultural significance and importance as animal models in biological and biomedical research. Chicken (Gallus gallus domesticus) genome is the most comprehensively investigated avian genome. In particular, several improved drafts of chicken genome assembly have been released [21,26], the detailed description of chicken karyotype has been provided [22] and molecular fluorescent in situ hybridization (FISH) probes to individual chicken chromosomes and their particular regions have been generated [22,27,28,29,30]. It serves a reliable basis for comparative investigations of genomes and karyotypes in various bird species and representatives of other animal taxonomic groups [23,31,32,33,34,35,36,37]. At the same time, even in case of chicken, cytogenetic and genomic data on at least the microchromosomal part of the karyotype are still limited.
Taking into account the complexity of amphibian and avian karyotypes, standard cytogenetic and cytological approaches to their investigation using compact metaphase chromosomes often prove to be inappropriate. In avian and amphibian growing diplotene oocytes, chromosomes take the so-called lampbrush form. As compared with mitotic metaphase chromosomes, lampbrush chromosomes are highly decondensed, transcriptionally active and characterized by specific chromomere-loop organization [38,39,40,41]. Lampbrush chromosomes are considerably larger than their metaphase counterparts. As an example, the size of meiotic bivalents in urodeles can reach up to 700 µm as in case of a salamander Salamandra salamandra and a newt Lissotriton vulgaris (previously Triturus vulgaris) [38]. Lampbrush chromosomes may form loci-specific prominent loops and structures with complex morphology [38,39,40]. The nature and function of such entities mainly remain to be discovered. Nevertheless, the specific structures can serve as reliable landmarks for identification of lampbrush chromosomes and their particular regions. Based on chromomere-loop pattern, unique for each individual chromosome, as well as the distribution of marker structures, detailed cytological maps of lampbrush chromosomes can be constructed [38,42,43,44,45,46,47,48,49,50,51].
Thus, due to the enormous size, peculiarities of the organization and enrichment with cytological markers, lampbrush chromosomes can serve as an opportune model for comprehensive cytogenetic and cytological investigations. Currently, the detailed protocols for preparation of lampbrush chromosomes were developed [52]. In particular, both chromosomal spreads and intact growing oocyte nuclei (germinal vesicles) can be subjected to immunofluorescent staining or different procedures of FISH (Figure 1 and Figure 2) [52,53].
Lampbrush chromosomes are also known in other vertebrate species including fishes and reptilians [38,55,56]. However, there is a lack of data on lampbrush chromosomes in these classes of vertebrates. In fact, studies involving reptilian lampbrush chromosomes are restricted to histological observations of ovary development [56,57] or analysis of lampbrush chromosomes in intact oocyte nuclei [58]. However, Lukina and Kupriyanova demonstrated the possibility of lampbrush chromosomes isolation for several lizard species [59,60]. Nevertheless, an adapted protocol for preparation of lampbrush chromosomes spreads, the description of chromosomal morphology and construction of detailed cytological maps are still required. Rare usage of reptilian lampbrush chromosomes is hardly explained by technical difficulties in chromosome preparation but is rather due to low interest from the cytogenetic point of view, absence of model organisms and until recently lack of reliable molecular markers (e.g., bacterial artificial chromosome (BAC)-clones and paints) [61,62,63,64]. Thus, here we focus on the main findings on chromosomal evolution that were obtained using avian and amphibian lampbrush chromosomes.
2. Lampbrush Chromosomes as a Tool to Study Amphibian Chromosomal Evolution
2.1. Interspecies Differences
From pioneer works, amphibian lampbrush chromosomes represent an appropriate and convenient tool for analysis of amphibian karyotypes due to the abundance and variety of marker structures [38,43,51,65,66,67]. To facilitate lampbrush chromosomes identification authors constructed cytological maps with relative position of landmarks for variety of Urodeles and Anuran species [38,43,47,50,51,65,68]. Here, we focus on some of the most fascinating cases concerning karyotype evolution, accumulation of polymorphisms and sex chromosome origin provided by lampbrush chromosomes studies.
Chromosomal rearrangements acquired through amphibian karyotypes evolution are less frequent compared to mammalian karyotypes [69]. Nevertheless, the level of chromosomal rearrangements in amphibian karyotypes had been underestimated for the long time and proved to be comparable with birds, reptilians and fishes [69]. Advanced salamanders and frogs species underwent quite massive karyotype rearrangements, including fusion of the ancestral chromosomes [7,69,70]. Intra- and interchromosomal rearrangements such as inversions, fusions and translocations can be detected by the analysis of orthologous lampbrush chromosomes (Figure 3) [38,51,71,72]. Based on analysis of marker structures distribution on lampbrush chromosomes, Callan [38] observed no translocations in interspecific hybrids of European newt species (genus Triturus). He concluded that reciprocal translocations had not occurred in chromosomes of European newts, which is in agreement with generally conservative chromosome evolution described in this genus [51,73].
Positions and distribution of various landmark loops on lampbrush chromosomes correlate with phylogenetic relationships between different species [38,51,74]. Thus, closely related species usually have more similar landmark patterns [51,74]. Comparison of landmarks on lampbrush chromosomes allowed to find phylogenetic relationships between different species from Urodeles and Anura, even before reliable phylogenetic markers were obtained [51,66,74]. Such comparison for three closely related Triturus species Triturus cristatus, Triturus carnifer and Triturus karelinii revealed similar distribution patterns of landmarks [51,75]. Lampbrush chromosomes of a sister species Triturus marmoratus strongly differ from lampbrush chromosomes of the species within Triturus christatus groups [51,75]. Nevertheless, phylogeny of more distant species is difficult to unravel via lampbrush chromosome analysis [51,66,67].
2.2. Interpopulation Differences
In addition to interspecies divergence, intraspecies genetic polymorphisms between populations can also be investigated using lampbrush chromosomes as an instrument [50,66,76,77]. Variability of lampbrush chromosome landmarks in animals from isolated populations indicates accumulation of polymorphisms (Figure 3). Such polymorphisms inevitably appear during reduced genetic flow between separate populations [78]. Thus, lampbrush chromosomes can be used as a tool for studying polymorphisms accumulation that is considered as an initial stage of allopatric speciation [78].
Moreover, analysis of landmarks polymorphisms allows to track distribution of species in new habitants [76]. Thorough analysis of landmarks on lampbrush chromosomes from continental and Japan populations of Pelophylax nigromaculata allowed to conclude that the species spread after invasion [76]. Japanese P. nigromaculata genetically differentiated into four groups that appeared after the termination of migration caused by geographic obstacles. Ohtani detected similarities of lampbrush marker structures between frogs from continental and certain Japanese populations [76]. This fact was considered as an evidence of introgression of genetic material in the Japanese populations after a secondary contact with the continental population of P. nigromaculata [76].
Another example of interpopulational polymorphism together with the detection of chromosomal rearrangements came from the analysis of Rana rugosa sex chromosomes in the lampbrush form. R. rugosa represents a system where sex determination type differs in distinct populations: in some populations, males are heterogametic (XX/XY) while in the other females are heterogametic (ZZ/ZW) [79]. However, in two additional populations, these frogs exhibit homomorphic sex chromosomes [79]. Analysis of marker structures on sex chromosomes in the lampbrush form revealed similarities between all different types of R. rugosa sex chromosomes indicating their common origin from the ancestral chromosome similar to the homomorphic one [71,72,79]. Data on distribution of chiasmata and patterns of landmarks on lampbrush chromosomes demonstrated two independent inversions resulted in the emergence of several types of sex chromosomes in this species: W/X sex chromosomes appeared after a terminal inversion, while Z/Y chromosomes appeared after a pericentric inversion of the ancestral chromosomes and subsequent deletion of approximately 10% of the chromosome [71,72,79].
2.3. Sex Chromosomes
Analysis of landmarks and chiasmata distribution in lampbrush chromosome allows to solve a problem of sex chromosomes identification if they are undistinguishable at metaphase. In the case of female heterogametic sex (ZW), one can detect Z and W sex chromosomes as a bivalent between two homologs with various patterns of marker loops and sometimes with a decreased level of chiasmata [12,14]. Using this approach, sex chromosomes in Pleurodeles potreti were identified due to a short heteromorphic region near the middle of the lampbrush bivalent 4 [14]. At the same time, in frog Buergeria buergeri, sex chromosomes were identified as a lampbrush bivalent with the only terminal chiasmata [80]. In another fascinating study, lampbrush chromosomes were obtained from males reversed into females after hormonal treatment [13]. Even when males’ sex is heterogametic, it is possible to apply lampbrush chromosomes to reveal differences in sex chromosomes [13]. Sex reversal experiments confirmed female heterogamety in P. potreti: neofemales (WW) exhibited similarity in the same region on the bivalent 4 [14]. Nevertheless, it is not always possible to observe differences between sex chromosomes during the lampbrush chromosome stage. Both females and reversed males of Triturus species that have XX/XY sex determination type did not exhibit any significant differences in sex chromosomes morphology [12].
Interesting example of sex chromosome evolution was observed after lampbrush chromosome analysis of seven species from Triturus genus [51,75]. In all of these species, bivalent 1 consists of a longer homolog with an extended achiasmatic region of compact loops and a shorter one with a more regular loop pattern [12,38,51,81]. Such unusual morphology is based on crucial differences in heterochromatic component of long arms between two homologous chromosomes 1: 1A and 1B [82]. Moreover, in homomorphic state (A1A1 or A2A2), all embryos usually die or exhibit severe development abnormalities [13,82]. One of the hypothesis aimed to explain the origin of this locus suggested a reciprocal translocation between two homologous autosomes in the common ancestor of species from T. cristatus subgenus [82,83]. Other more plausible explanation is based on the assumption that chromosome 1 is a relict sex chromosome [12]. After switching from WZ/WW to XY/XX sex determination system the majority of modern Triturus species are thought to eliminate Z chromosome and retain a WW chromosome pair while ancestors of species from a T. cristatus group probably retained ZW chromosomes which are currently known as A1 and A2 chromosomes [12,84].
An additional example of the application of lampbrush chromosomes in the field of karyotype evolution and sex chromosome emergence comes from investigations of frog Leopelma hochstetteri. Sex chromosome (W) of this species is represented by a supernumerary chromosome that highly varies in size, centromeric index and heterochromatin amount [85,86,87]. As Z chromosome was not found, authors suggested a W0/00 sex determination system for the species [85,86,88]. However, after lampbrush chromosomes analysis of frogs from one population researches did not find any supernumerary chromosomes but detected a heteromorphic lampbrush bivalent [89]. This bivalent was considered to represent a sex WZ bivalent where one homolog shared some kind of similarity in a marker loops pattern with supernumerary sex chromosomes in frogs from other populations [89]. Authors suggested that either loss of Z chromosome or its homogenization in occasional ZZW trisomy cases resulted in such a peculiar W0/00 or WZZ/ZZ sex determination type spread in other populations [86,89]. Subsequently, either translocation from W chromosome or degradation of W chromosome occurred independently in each population under Muller’s ratchet mechanism producing a variety of supernumerary W chromosomes [86,89]. In addition to supernumerary W chromosome, extra supernumerary B chromosomes varying in number, morphology and heterochromatin amount were revealed in the majority of populations [85,86]. These results indicate a higher chromosome evolution rate in L. hochstetteri than in other amphibians [88,90]. To explain this phenomenon, Bogart [90] proposed that rates of chromosomal evolution in amphibians might depend on the population size and animals reproductive behavior. For instance, in amphibians that form large breeding groups (Ranidae or Bufonidae) chromosomal evolution will be slower as compared to a small inbred population of neotropical frogs (such as Leopelma) with the unique breeding behavior [88,89,90].
2.4. Interspecies Hybrids
Amphibian species reveal a specific way of obtaining chromosomal novelties via interspecies hybridization [15,16,91]. Usually interspecies hybrids die during early development and even in case of survival cannot produce gametes [78]. After analysis of lampbrush chromosomes from oocytes of such interspecies hybrids, researchers found occasional chiasmata between orthologous chromosomes, which probably can provide chromosome separation and partial fertility [38,92]. Such hybrids represent a unique model for identification of homology regions between orthologous chromosomes after chiasmata analysis.
Some interspecies hybrids can produce progeny via modifications of their gametogenesis including selective elimination of chromosomes originating from one parental species and/or genome duplication [91,93,94]. Such alterations of gametogenesis frequently lead to the emergence of polyploid hybrids [15,16,93,94]. Owing to the presence of species-specific landmarks, lampbrush chromosomes analysis is a useful and reliable approach to identify genomes transmitted in oocytes of di- and polyploid hybrids and to reveal genome elimination and/or duplication during hybrid gametogenesis [50,66,67,95,96,97]. Based on analysis of lampbrush chromosomes from interspecies hybrid salamanders (genus Ambystoma), Macgregor and Uzzell [95] inferred that genome endoreplication occurred premeiotically in germline cells allowing gynogenetic reproduction. Moreover, both elimination of one parental genome and endoreplication of another one were revealed using lampbrush chromosomes analysis in diploid and triploid hybrids from Pelophylax esculentus complex that reproduces via hybridogenesis [66,96].
Genome elimination and duplication prevent recombination events between genomes of separate species [91,93,94]. Nevertheless, occasional introgression of genetic material was observed between their genomes [98,99]. For example, having applied genomic in situ hybridization (GISH) on lampbrush chromosomes of hybrid salamanders, Bogart and coauthors [99] demonstrated extensive chromosomal exchange between genomes of parental species. According to the hypothesis of Ohno [100], such occasional recombination events result in homogenization and diploidization of parental genomes within a hybrid individual. Otherwise, the absence of recombination in the allopolyploid hybrids can subsequently lead to the independent evolution of each parental chromosomal set within the allopolyploid hybrid genome. It was clearly demonstrated for an allopolyploid frog X. laevis [7]. Dissimilarity in centromeric repeats between two different chromosomal sets of X. laevis was proven by FISH on lampbrush chromosomes. This approach allowed to identify chromosomes bearing centromeric repeat whereas in mitosis these chromosomes are of an equal size [101]. Moreover, in X. laevis, two genomes are characterized by different chromosomal length, chromosomal rearrangements and transposon families spread after hybridization [7].
3. Lampbrush Chromosomes as a Tool to Study Avian Chromosomal Evolution
Comparative molecular–cytogenetic studies involving lampbrush chromosomes proved to be helpful to reveal new evolutional changes, both inter- and intrachromosomal rearrangements, and to specify the breakpoints with high-resolution in Galliform species. In particular, it is known that some variation in chromosome number among Galliform species is mainly caused by the interchromosomal rearrangements involving ancestral chromosomes 2 and 4 [23,102,103,104,105]. As an example, using standard cytogenetic and molecular–cytogenetic approaches, it had been earlier suggested that chicken (Gallus g. domesticus, GGA) and turkey (Meleagris gallopavo, MGA) karyotypes are discriminated by two interchromosomal rearrangements with the orthologs of chicken chromosomes 2 (GGA2) and 4 (GGA4) being composed of turkey chromosomes 3 (MGA3) and 6 (MGA6), and 4 (MGA4) and 9 (MGA9), correspondingly [23]. The application of turkey chromosome painting probes for MGA3 and MGA6, as well as for MGA4 to chicken lampbrush chromosomes clearly demonstrated that the breakpoint of the interchromosomal rearrangements corresponds to the centromere of chicken chromosome 2 (GGA2) and 4 (GGA4) [105].
The karyotypes of chicken and Japanese quail (Coturnix coturnix japonica, CCO) are very similar, with the same diploid number (2n = 78) and high synteny conservation demonstrated repeatedly by comparative physical mapping [32,44,49,106,107], chromosome painting [31,102] and genetic linkage analysis [107,108]. At the same time, centromere position on the majority of orthologous chromosomes differs between these two species. In particular, based on the mismatch of some blocks of G-banded chromosomes [109,110,111] and the pattern of comparative FISH with cloned chicken genome sequences [32,106,107] it was suggested that pericentric inversions are responsible for the discrepancy in centromere position between chicken and quail macrochromosomes 1, 2 and 4. Dense comparative FISH-mapping of chicken BACs to chicken and quail lampbrush chromosomes confirmed the presence of the pericentric inversion between GGA2 and CCO2, revealed the inversion between GGA11 and CCO11, and allowed to narrow down their breakpoint positions [49]. At the same time, FISH on lampbrush chromosomes demonstrated the same order of molecular markers along GGA1 and CCO1 [49] as well as GGA4 and CCO4 [44] with centromeres being flanked by different genomic material in the two species. Additionally, FISH on lampbrush chromosomes revealed the inversion on chromosome 3 but, again, the difference in centromere positions between GGA3 and CCO3 could not be explained by the inversion event only [49]. The phenomenon of “centromere repositioning” or “evolutionary new centromere” (ENC) formation, where a centromere could arise during the evolution in a new chromosomal locus without any changes in the gene order, has been described repeatedly in different taxonomic groups [112,113,114,115,116,117,118]. Among birds, single cases of ENCs events have been suggested in Galliformes by comparison of orthologous chromosomes 4 of chicken and red-legged partridge [119] as well as chromosomes Z of chicken and Pekin duck [120]. The usage of elongated lampbrush chromosomes for high-resolution comparative cytogenetic analysis clearly demonstrated that “centromere repositioning” events could also take place during the divergence of chicken and Japanese quail karyotypes.
Regarding the microchromosomal part of chicken and quail karyotypes, the usage of lampbrush chromosomes [121] and synaptonemal complex spreads from pachytene nuclei [122] allowed demonstrating that the majority of Japanese quail microchromosomes are submetacentric which differs them from the chicken orthologs known to be acrocentric [121,123,124]. In particular, the dissimilarity in centromere localization was unambiguously shown by immunofluorescent detection of cohesin-enriched protein granules that serve as a reliable marker of centromeres on Galliform lampbrush chromosomes as well as by FISH-mapping of pericentromeric chicken nuclear-membrane-associated repeat (CNM) and BglII- repeat [121]. Comprehensive investigation of epigenetic chromatin state of Japanese quail microbivalents demonstrated that short arms of submetacentric microchromosomes are not completely condensed but enriched with heterochromatin protein 1 (HP1β) and repressive histone modifications, including H3K27me3 [125]. Together with the data on high-resolution comparative BAC-clone mapping on the five largest chicken microchromosomes and their quail orthologs [49], these results suggest that the discrepancy in centromere position on microchromosomes between the two species might be due to the accumulation of species-specific distinct type of heterochromatin constituting the polymorphic short arms of quail microchromosomes [125].
Thus, the usage of lampbrush chromosomes as a powerful tool for high-resolution physical mapping allows extending our knowledge of chromosomal rearrangements accompanied Galliform karyotype evolution. That is, the data obtained support the idea that the number of intrachromosomal imbalances affected highly conserved avian karyotypes seems to be higher than it could be assumed based on results of standard cytogenetic and molecular–cytogenetic analysis, which is consistent with the comprehensive molecular and in silico data on both macro- and microchromosomes rearrangements in avian genome evolution [126,127,128].
4. Conclusions
The application of classical cytological approaches, mainly based on the analysis of cytological maps and distribution of marker structures, to lampbrush chromosomes allowed to shed light on various aspects of chromosome evolution in diverse amphibian species. In comparison, avian lampbrush chromosomes possess a much lower number of cytological landmarks. In this regard, the detailed studies on avian chromosome evolution using lampbrush chromosomes became possible since modern molecular–cytogenetic techniques (FISH-mapping, chromosomal painting, etc.) became widespread. The application of lampbrush chromosomes as a tool for high-resolution cytological and cytogenetic analysis allows to obtain unique data on chromosome evolution and gives prospects for exploration of complex karyotypes, as in the case of amphibians and birds. Moreover, lampbrush chromosome analysis seems to be promising in investigation the details of chromosomal evolution in other animals including reptiles and fishes.
Acknowledgments
The research was supported by a grant of the President of the Russian Federation (MK-1630.2017.4). The work was partially performed using experimental equipment of the Environmental Safety Observatory and the Research Resource Centers “Chromas” and “Molecular and Cell Technologies” of St. Petersburg State University.
Author Contributions
A.Z., D.D., and A.K. wrote the first draft of the manuscript and prepared the figures. All authors were involved in the revision of the manuscript and have agreed to the final content.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Gregory, T.R. Genome size and evolution in animals. In The Evolution of the Genome; Gregory, T.R., Ed.; Elsevier: Burlington, NJ, USA, 2005; pp. 3–87. [Google Scholar]
- Morescalchi, A.; Olmo, E. Single-copy DNA and vertebrate phylogeny. Cytogenet. Cell Genet. 1982, 34, 93–101. [Google Scholar] [CrossRef] [PubMed]
- Sun, C.; Shepard, D.B.; Chong, R.A.; Lopez Arriaza, J.; Hall, K.; Castoe, T.A.; Feschotte, C.; Pollock, D.D.; Mueller, R.L. LTR transposons contribute to genomic gigantism in plethodontid salamanders. Genome Biol. Evol. 2012, 4, 168–183. [Google Scholar] [CrossRef] [PubMed]
- Smith, J.J.; Putta, S.; Zhu, W.; Pao, G.M.; Verma, I.M.; Hunter, T.; Bryant, S.V.; Gardiner, D.M.; Harkins, T.T.; Voss, S.R. Genic regions of a large salamander genome contain long introns and novel genes. BMC Genom. 2009, 10, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Hellsten, U.; Harland, R.M.; Gilchrist, M.J.; Hendrix, D.; Jurka, J.; Kapitonov, V.; Ovcharenko, I.; Putnam, N.H.; Shu, S.; Taher, L.; et al. The genome of the Western clawed frog Xenopus tropicalis. Science 2010, 328, 633–636. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.B.; Xiong, Z.J.; Xiang, X.Y.; Liu, S.P.; Zhou, W.W.; Tu, X.L.; Zhong, L.; Wang, L.; Wu, D.D.; Zhang, B.L.; et al. Whole-genome sequence of the Tibetan frog Nanorana parkeri and the comparative evolution of tetrapod genome. Proc. Natl. Acad. Sci. USA 2015, 112, 1257–1262. [Google Scholar] [CrossRef] [PubMed]
- Session, A.M.; Uno, Y.; Kwon, T.; Chapman, J.A.; Toyoda, A.; Takahashi, S.; Fukui, A.; Hikosaka, A.; Suzuki, A.; Kondo, M.; et al. Genome evolution in the allotetraploid frog Xenopus laevis. Nature 2016, 538, 336–343. [Google Scholar] [CrossRef] [PubMed]
- Schmid, M. Chromosome banding in Amphibia. I. Constitutive heterochromatin and nucleolus organizer regions in Bufo and Hyla. Chromosoma 1978, 66, 361–388. [Google Scholar] [CrossRef]
- Schempp, W.; Schmid, M. Chromosome banding in amphibia. VI. BrdU-replication patterns in anura and demonstration of XX/XY sex chromosomes in Rana esculenta. Chromosoma 1981, 83, 697–710. [Google Scholar] [CrossRef] [PubMed]
- Morescalchi, A. Evolution and karyology of the amphibians. Boll. Zool. 1980, 47, 113–126. [Google Scholar] [CrossRef]
- Schmid, M.; Steinlein, C.; Bogart, J.P.; Feichtinger, W.; León, P.; La Marca, E.; Díaz, L.M.; Sanz, A.; Chen, S.H.; Hedges, S.B. The chromosomes of terraranan frogs. Insights into vertebrate cytogenetics. Cytogenet. Genome Res. 2010, 130–131, 1–568. [Google Scholar] [CrossRef] [PubMed]
- Wallace, H.; Wallace, B.M.; Badawy, G.M. Lampbrush chromosomes and chiasmata of sex-reversed crested newts. Chromosoma 1997, 106, 526–533. [Google Scholar] [CrossRef] [PubMed]
- Wallace, H.; Badawy, G.M.; Wallace, B.M. Amphibian sex determination and sex reversal. Cell. Mol. Life Sci. 1999, 55, 901–909. [Google Scholar] [CrossRef] [PubMed]
- Lacroix, J.C.; Azzouz, R.; Simon, F.; Bellini, M.; Charlemagne, J.; Dournon, C. Lampbrush, W and Z heterochromosome characterization with a monoclonal antibody and heat-induced chromosomal markers in the newt Pleurodeles waltl: W chromosome plays a role in female sex determination. Chromosoma 1990, 99, 307–314. [Google Scholar] [CrossRef] [PubMed]
- Mable, B.K.; Alexandrou, M.A.; Taylor, M.I. Genome duplication in amphibians and fish: An extended synthesis. J. Zool. 2011, 284, 151–182. [Google Scholar] [CrossRef]
- Schmid, M.; Evans, B.J.; Bogart, J.P. Polyploidy in amphibia. Cytogenet. Genome Res. 2015, 145, 315–330. [Google Scholar] [CrossRef] [PubMed]
- Tegelström, H.; Ryttman, H. Chromosomes in birds (Aves): Evolutionary implications of macro-and microchromosome numbers and lengths. Hereditas 1981, 94, 225–233. [Google Scholar] [CrossRef]
- Christidis, L. Aves. In Animal Cytogenetics, Chordata 3 B; John, B., Kayano, H., Levan, A., Eds.; Gebrüder Borntraeger: Berlin, Germany, 1990; Volume 4. [Google Scholar]
- Rodionov, A.V. Evolution of avian chromosomes and linkage groups. Russ. J. Genet. 1997, 33, 605–617. [Google Scholar]
- Burt, D.W. Origin and evolution of avian microchromosomes. Cytogenet. Genome Res. 2002, 96, 97–112. [Google Scholar] [CrossRef] [PubMed]
- International Chicken Genome Sequencing Consortium (ICGSC). Sequence and comparative analysis of the chicken genome provide unique perspectives on vertebrate evolution. Nature 2004, 432, 695–716. [Google Scholar]
- Masabanda, J.S.; Burt, D.W.; O’Brien, P.C.M. Molecular cytogenetic definition of the chicken genome: The first complete avian karyotype. Genetics 2004, 166, 1367–1373. [Google Scholar] [CrossRef] [PubMed]
- Griffin, D.K.; Robertson, L.B.W.; Tempest, H.G.; Skinner, B.M. The evolution of the avian genome as revealed by comparative molecular cytogenetics. Cytogenet. Genome Res. 2007, 117, 64–77. [Google Scholar] [CrossRef] [PubMed]
- McQueen, H.A.; Siriaco, G.; Bird, A.P. Chicken microchromosomes are hyperacetylated, early replicating, and gene rich. Genome Res. 1998, 8, 621–630. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smith, J.; Bruley, C.K.; Paton, I.R.; Dunn, I.; Jones, C.T.; Windsor, D.; Morrice, D.R.; Law, A.S.; Masabanda, J.; Sazanov, A.; et al. Differences in gene density on chicken macrochromosomes and microchromosomes. Anim. Genet. 2000, 31, 96–103. [Google Scholar] [CrossRef] [PubMed]
- The National Center for Biotechnology Information (NCBI). Genomes. Available online: http://www.ncbi.nlm.nih.gov/genome/111 (accessed on 27 August 2017).
- Zoorob, R.; Billault, A.; Severac, V.; Fillon, V.; Vignal, A.; Auffray, C. Two chicken genomic libraries in the PAC and BAC cloning systems: Organization and characterization. Anim. Genet. 1996, 27, 2–69. [Google Scholar]
- Griffin, D.K.; Haberman, F.; Masabanda, J. Micro- and macrochromosome paints generated by flow cytometry and microdissection: Tools for mapping the chicken genome. Cytogenet. Cell Genet. 1999, 87, 278–281. [Google Scholar] [CrossRef] [PubMed]
- Crooijmans, R.P.M.A.; Vrebalov, J.; Dijkhof, R.J.; van der Poel, J.J.; Nicholls, R.D.; Bovenhuis, H.; Groenen, M.A. Two-dimensional screening of the Wageningen chicken BAC library. Mamm. Genome 2000, 11, 360–363. [Google Scholar] [CrossRef] [PubMed]
- BACPAC Resources Center (BPRC). Available online: https://bacpacresources.org (accessed on 20 August 2017).
- Schmid, M.; Nanda, I.; Guttenbach, M.; Steinlein, C.; Hoehn, M.; Schartl, M.; Haaf, T.; Weigend, S.; Fries, R.; Buerstedde, J.M.; et al. First report on chicken genes and chromosomes. Cytogenet. Cell Genet. 2000, 90, 169–218. [Google Scholar] [CrossRef] [PubMed]
- Schmid, M.; Nanda, I.; Hoehn, H.; Schartl, M.; Haaf, T.; Buerstedde, J.M.; Arakawa, H.; Caldwell, R.B.; Weigend, S.; Burt, D.W.; et al. Second report on chicken genes and chromosomes. Cytogenet. Genome Res. 2005, 109, 415–479. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schmid, M.; Smith, J.; Burt, D.W.; Aken, B.L.; Antin, P.B.; Archibald, A.L.; Ashwell, C.; Blackshear, P.J.; Boschiero, C.; Brown, C.T.; et al. Third report on chicken genes and chromosomes 2015. Cytogenet. Genome Res. 2015, 145, 78–179. [Google Scholar] [CrossRef] [PubMed]
- Larkin, D.M.; Pape, G.; Donthu, R.; Auvil, L.; Welge, M.; Lewin, H.A. Breakpoint regions and homologous synteny blocks in chromosomes have different evolutionary histories. Genome Res. 2009, 19, 770–777. [Google Scholar] [CrossRef] [PubMed]
- Zhang, G.; Li, C.; Li, Q.; Li, B.; Larkin, D.M.; Lee, C.; Storz, J.F.; Antunes, A.; Greenwold, M.J.; Meredith, R.W.; et al. Comparative genomics reveals insights into avian genome evolution and adaptation. Science 2014, 346, 1311–1320. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Farré, M.; Narayan, J.; Slavov, G.T.; Damas, J.; Auvil, L.; Li, C.; Jarvis, E.D.; Burt, D.W.; Griffin, D.K.; Larkin, D.M. Novel insights into chromosome evolution in birds, archosaurs, and reptiles. Genome Biol. Evol. 2016, 8, 2442–2451. [Google Scholar] [CrossRef] [PubMed]
- Damas, J.; O’Connor, R.; Farré, M.; Lenis, V.P.E.; Martell, H.J.; Mandawala, A.; Fowler, K.; Joseph, S.; Swain, M.T.; Griffin, D.K.; et al. Upgrading short-read animal genome assemblies to chromosome level using comparative genomics and a universal probe set. Genome Res. 2017, 27, 875–884. [Google Scholar] [CrossRef] [PubMed]
- Callan, H.G. Lampbrush Chromosomes. Mol. Biol. Biochem. Biophys. 1986, 36, 1–254. [Google Scholar] [PubMed]
- Morgan, G.T. Lampbrush chromosomes and associated bodies: New insights into principles of nuclear structure and function. Chromosom. Res. 2002, 10, 177–200. [Google Scholar] [CrossRef]
- Gaginskaya, E.; Kulikova, T.; Krasikova, A. Avian lampbrush chromosomes: A powerful tool for exploration of genome expression. Cytogenet. Genome Res. 2009, 124, 251–267. [Google Scholar] [CrossRef] [PubMed]
- Macgregor, H.C. Chromomeres revisited. Chromosom. Res. 2012, 20, 911–924. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chelysheva, L.A.; Solovei, I.V.; Rodionov, A.V.; Yakovlev, A.F.; Gaginskaya, E.R. The lampbrush chromosomes of the chicken. The cytological map of macrobivalents. Tsitologiia 1990, 32, 303–316. [Google Scholar] [PubMed]
- Callan, H.G.; Gall, J.G.; Berg, A. The lampbrush chromosomes of Xenopus laevis: Preparation, identification, and distribution of 5s DNA sequences. Chromosoma 1987, 95, 236–250. [Google Scholar] [CrossRef] [PubMed]
- Galkina, S.; Deryusheva, S.; Fillon, V.; Vignal, A.; Crooijmans, R.; Groenen, M.; Rodionov, A.; Gaginskaya, E. FISH on avian lampbrush chromosomes produces higher resolution gene mapping. Genetica 2006, 128, 241–251. [Google Scholar] [CrossRef] [PubMed]
- Daks, A.; Deryusheva, S.; Krasikova, A.; Zlotina, A.; Gaginskaya, E.; Galkina, S. Lampbrush Chromosomes of the Japanese Quail (Coturnix coturnix japonica): A New Version of Cytogenetic Maps. Genetika 2010, 46, 1335–1338. [Google Scholar] [CrossRef] [PubMed]
- Rodionov, A.V.; Chechik, M.S. Cytogenetic map of the Japanese quail lampbrush chromosomes. Genetika 2002, 38, 1246–1251. [Google Scholar] [PubMed]
- Penrad-Mobayed, M.; El Jamil, A.; Kanhoush, R.; Perrin, C. Working map of the lampbrush chromosomes of Xenopus tropicalis: A new tool for cytogenetic analyses. Dev. Dyn. 2009, 238, 1492–1501. [Google Scholar] [CrossRef] [PubMed]
- Solinhac, R.; Leroux, S.; Galkina, S.; Chazara, O.; Feve, K.; Vignoles, F.; Morisson, M.; Derjusheva, S.; Bed’hom, B.; Vignal, A.; et al. Integrative mapping analysis of chicken microchromosome 16 organization. BMC Genom. 2010, 11, 616. [Google Scholar] [CrossRef] [PubMed]
- Zlotina, A.; Galkina, S.; Krasikova, A.; Crooijmans, R.P.; Groenen, M.A.; Gaginskaya, E.; Deryusheva, S. Centromere positions in chicken and Japanese quail chromosomes: De novo centromere formation versus pericentric inversions. Chromosom. Res. 2012, 20, 1017–1032. [Google Scholar] [CrossRef] [PubMed]
- Dedukh, D.; Mazepa, G.; Shabanov, D.; Rosanov, J.; Litvinchuk, S.; Borkin, L.; Saifitdinova, A.; Krasikova, A. Cytological maps of lampbrush chromosomes of European water frogs (Pelophylax esculentus complex) from the Eastern Ukraine. BMC Genet. 2013, 14, 1–26. [Google Scholar] [CrossRef] [PubMed]
- Macgregor, H.C.; Sessions, S.K.; Arntzen, J.W. An integrative analysis of phylogenetic relationships among newts of the genus Triturus (family Salamandridae), using comparative biochemistry, cytogenetics, and reproductive interactions. J. Evol. Biol. 1990, 3, 329–373. [Google Scholar] [CrossRef]
- Lampbrush chromosome protocols. Available online: http://projects.exeter.ac.uk/lampbrush/protocols.htm (accessed on 7 September 2017).
- Zlotina, A.; Krasikova, A. FISH in Lampbrush Chromosomes. In Fluorescence in situ Hybridization (FISH)—Application Guide, 2nd ed.; Liehr, T., Ed.; Springer: Berlin, Germany, 2017; pp. 445–457. ISBN 978-3662529577. [Google Scholar]
- Zlotina, A.; Galkina, S.; Krasikova, A.; Crooijmans, R.P.; Groenen, M.A.; Gaginskaya, E.; Deryusheva, S. Precise centromere positioning on chicken chromosome 3. Cytogenet. Genome Res. 2010, 129, 310–313. [Google Scholar] [CrossRef] [PubMed]
- Guraya, S.S. The cell and molecular biology of fish oogenesis. Monogr. Dev. Biol. 1986, 18, 1–223. [Google Scholar] [PubMed]
- Callebaut, M.; Van Nassauw, L.; Harrisson, F. Comparison between oogenesis and related ovarian structures in a reptile, Pseudemys scripta elegans (turtle) and in a bird Coturnix coturnix japonica (quail). Reprod. Nutr. Dev. EDP Sci. 1997, 37, 233–252. [Google Scholar] [CrossRef]
- Moore, B.C.; Uribe-Aranabal, M.C.; Boggs, A.S.P.; Guillette, L.J.J. Developmental morphology of the neonatal alligator (Alligator mississippiensis) ovary. J. Morphol. 2008, 269, 302–312. [Google Scholar] [CrossRef] [PubMed]
- Lutes, A.A.; Neaves, W.B.; Baumann, D.P.; Wiegraebe, W.; Baumann, P. Sister chromosome pairing maintains heterozygosity in parthenogenetic lizards. Nature 2010, 464, 283–286. [Google Scholar] [CrossRef] [PubMed]
- Lukina, N.A. Characterization of meiotic chromosomes in the oocytes of some Lacertidae (Reptilia). Tsitologiia 1994, 36, 323–329. [Google Scholar]
- Kupriyanova, L. Cytogenetic and genetic trends in the evolution of unisexual lizards. Cytogenet. Genome Res. 2009, 127, 273–279. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Miyake, T.; Edwards, S.V.; Amemiya, C.T. Tuatara (Sphenodon) genomics: BAC library construction, sequence survey, and application to the DMRT gene family. J. Hered. 2006, 97, 541–548. [Google Scholar] [CrossRef] [PubMed]
- Olmo, E. Trends in the evolution of reptilian chromosomes. Integr. Comp. Biol. 2008, 48, 486–493. [Google Scholar] [CrossRef] [PubMed]
- Chapus, C.; Edwards, S.V. Genome evolution in Reptilia: In silico chicken mapping of 12,000 BAC-end sequences from two reptiles and a basal bird. BMC Genom. 2009, 10, S8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pokorna, M.; Giovannotti, M.; Kratochvil, L.; Caputo, V.; Olmo, E.; Ferguson-Smith, M.A.; Rens, W. Conservation of chromosomes syntenic with avian autosomes in squamate reptiles revealed by comparative chromosome painting. Chromosoma 2012, 121, 409–418. [Google Scholar] [CrossRef] [PubMed]
- Muller, W.P. The lampbrush chromosomes of Xenopus laevis. Chromosoma 1974, 47, 283–296. [Google Scholar] [CrossRef] [PubMed]
- Bucci, S.; Ragghianti, M.; Mancino, G.; Berger, L.; Hotz, H.; Uzzell, T. Lampbrush and mitotic chromosomes of the hemiclonally reproducing hybrid Rana esculenta and its parental species. J. Exp. Zool. Suppl. 1990, 255, 37–56. [Google Scholar] [CrossRef] [PubMed]
- Guerrini, F.; Bucci, S.; Ragghianti, M.; Mancino, G.; Hotz, H.; Uzzell, T.; Berger, L. Genomes of two water frog species resist germ line exclusion in interspecies hybrids. J. Exp. Zool. 1997, 279, 163–176. [Google Scholar] [CrossRef]
- Vishnyakova, N.M.; Lacroix, J.-C.; Rodionov, A.V. Cytogenetic maps of lampbrush chromosomes of newts of the genus Pleurodeles: An algorithm of lampbrush chromosome identification in Pleurodeles waltl by immunocytochemical staining of landmark loops with polyclonal anti-Ro52 antisera. Russ. J. Genet. 2004, 40, 491–499. [Google Scholar]
- Smith, J.J.; Voss, S.R. Gene order data from a model amphibian (Ambystoma): New perspectives on vertebrate genome structure and evolution. BMC Genom. 2006, 7, 219. [Google Scholar] [CrossRef]
- Voss, S.R.; Kump, D.K.; Putta, S.; Pauly, N.; Reynolds, A.; Henry, R.J.; Basa, S.; Walker, J.A.; Smith, J.J. Origin of amphibian and avian chromosomes by fission, fusion, and retention of ancestral chromosomes. Genome Res. 2011, 21, 1306–1312. [Google Scholar] [CrossRef] [PubMed]
- Miura, I.; Ohtani, H.; Kashiwagi, A.; Hanada, H.; Nakamura, M. Structural differences between XX and ZW sex lampbrush chromosomes in Rana rugosa females (Anura: Ranidae). Chromosoma 1996, 105, 237–241. [Google Scholar] [CrossRef] [PubMed]
- Miura, I.; Ohtani, H.; Hanada, H.; Ichikawa, Y.; Kashiwagi, A.; Nakamura, M. Evidence for two successive pericentric inversions in sex lampbrush chromosomes of Rana rugosa (Anura: Ranidae). Chromosoma 1997, 106, 178–182. [Google Scholar] [CrossRef] [PubMed]
- Mancino, G. Evolutionary cytogenetics of newts of the genus Triturus as revealed using monoclonal antibodies. In Amphibian Cytogenetics and Evolution; Green, D.M., Sessions, S.K., Eds.; Academic Press: New York, NY, USA, 1991; pp. 197–215. [Google Scholar]
- Ohtani, H. Lampbrush chromosomes of Rana nigromaculata, R. brevipoda, R. plancyi chosenica, R. p. fukienensis and their reciprocal hybrids. Sci. Rep. Lab. Amphib. Biol. Hiroshima Univ. 1990, 10, 165–221. [Google Scholar]
- Wielstra, B.; Baird, A.B.; Arntzen, J.W. A multimarker phylogeography of crested newts (Triturus cristatus superspecies) reveals cryptic species. Mol. Phylogenet. Evol. 2013, 67, 167–175. [Google Scholar] [CrossRef] [PubMed]
- Ohtani, H. Polymorphism of lampbrush chromosomes in Japanese populations of Rana nigromaculata. Zool. Sci. 1994, 11, 337–342. [Google Scholar]
- Ohtani, H. Local variations in the lampbrush chromosomes of the Japanese pond frog, Rana porosa. Caryologia 1995, 48, 189–199. [Google Scholar] [CrossRef]
- Coyne, J.A.; Orr, H.A. The evolutionary genetics of speciation. Philos. Trans. R. Soc. Lond. Ser. B 1998, 353, 287–305. [Google Scholar] [CrossRef] [PubMed]
- Miura, I. An evolutionary witness: The frog Rana rugosa underwent change of heterogametic sex from XY male to ZW female. Sex. Dev. 2007, 1, 323–331. [Google Scholar] [CrossRef] [PubMed]
- Ohta, S. Sex-determining mechanism in Buergeria buergeri (Schlegel): I. Heterozygosity of chromosome pair No. 7 in the female. Sci. Rep. Lab. Amphib. Biol. Hiroshima Univ. 1986, 8, 29–43. [Google Scholar]
- Callan, H.G.; Lloyd, L. Lampbrush chromosomes of crested newts Triturus cristatus (Laurenti). Philos. Trans. R. Soc. B 1960, 243, 135–219. [Google Scholar] [CrossRef]
- Sims, S.H.; Macgregor, H.C.; Pellatt, P.S.; Horner, H.A. Chromosome 1 in crested and marbled newts (Triturus). An extraordinary case of heteromorphism and independent chromosome evolution. Chromosoma 1984, 89, 169–185. [Google Scholar] [CrossRef]
- Sessions, S.K.; Macgregor, H.C.; Schmid, M.; Haaf, T. Cytology, embryology, and evolution of the development arrest syndrome in newts of the genus Triturus (Caudata: Salamandridae). J. Exp. Zool. 1988, 248, 321–334. [Google Scholar] [CrossRef] [PubMed]
- Hillis, D.M.; Green, D.M. Evolutionary changes of heterogametic sex in the phylogenetic history of amphibians. J. Evol. Biol. 1990, 3, 49–64. [Google Scholar] [CrossRef]
- Green, D.M.; Kezer, J.; Nussbaum, R.A. Supernumerary chromosome variation and heterochromatin distribution in the endemic New Zealand frog Leopelma hochstetteri. Chromosoma 1987, 95, 339–344. [Google Scholar] [CrossRef] [PubMed]
- Green, D.M. Cytogenetics of the endemic New Zealand frog, Leopelma hochstetteri: Extraordinary supernumerary chromosome variation and a unique sex chromosome system. Chromosoma 1988, 97, 55–70. [Google Scholar] [CrossRef]
- Green, D.M. Heteromorphic sex chromosomes in the rare and primitive frog Leopelma hamiltonii from New Zealand. J. Heredity 1988, 79, 165–169. [Google Scholar] [CrossRef]
- Zeul, C. Genome Evolution in the Primitive Frog Leopelma hochstetteri. Ph.D. Thesis, McGill University, Montreal, QC, Canada, 1991. [Google Scholar]
- Green, D.M.; Zeyl, C.W.; Sharbel, T.F. The evolution of hypervariable sex and supernumerary (B) chromosomes in the relict New Zealand frog, Leiopelma hochstetteri. J. Evol. Biol. 1993, 6, 417–441. [Google Scholar] [CrossRef]
- Bogart, J.P. The influence of life history on karyotypic evolution in frogs. In Amphibian Cytogencttcs and Evolution; Green, D.M., Sessions, S.K., Eds.; Academic Press: San Diego, CA, USA, 1991; pp. 233–258. [Google Scholar]
- Dawley, R.M.; Bogart, J.P. Evolution and Ecology of Unisexual Vertebrates; New York State Museum Bulletin 466; New York State Museum: Albany, NY, USA, 1989; p. 302.
- Muller, W.P. Diplotene chromosomes of Xenopus hybrid oocytes. Chromosoma 1977, 59, 273–282. [Google Scholar] [CrossRef] [PubMed]
- Neaves, W.B.; Baumann, P. Unisexual reproduction among vertebrates. Trends Genet. 2011, 27, 81–88. [Google Scholar] [CrossRef] [PubMed]
- Stenberg, P.; Saura, A. Meiosis and its deviations in polyploid animals. Cytogenet. Genome Res. 2013, 140, 185–203. [Google Scholar] [CrossRef] [PubMed]
- Macgregor, H.C.; Uzzell, T.M. Gynogenesis in salamanders related to Ambystoma jeffersonianum. Science 1964, 143, 1043–1045. [Google Scholar] [CrossRef] [PubMed]
- Dedukh, D.; Litvinchuk, S.; Rosanov, J.; Mazepa, G.; Saifitdinova, A.; Shabanov, D.; Krasikova, A. Optional endoreplication and selective elimination of parental genomes during oogenesis in diploid and triploid hybrid European water frogs. PLoS ONE 2015, 10, e0123304. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stöck, M.; Lamatsch, K.; Steinlein, C.; Epplen, J.T.; Grosse, W.-R.; Klapperstück, T.; Lampert, K.P.; Scheer, U.; Schmid, M.; Schartl, M. A bisexually reproducing all-triploid vertebrate. Nat. Genet. 2002, 30, 325–328. [Google Scholar] [CrossRef] [PubMed]
- Bi, K.; Bogart, J.P.; Fu, J. Intergenomic translocations in unisexual salamanders of the genus Ambystoma (Amphibia, Caudata). Cytogenet. Genome Res. 2007, 116, 289–297. [Google Scholar] [CrossRef] [PubMed]
- Bi, K.; Bogart, J.P. Probing the meiotic mechanism of intergenomic exchanges by genomic in situ hybridization on lampbrush chromosomes of unisexual Ambystoma (Amphibia: Caudata). Chromosom. Res. 2010, 18, 371–382. [Google Scholar] [CrossRef] [PubMed]
- Ohno, S. Evolution by Gene Duplication; Springer: New York, NY, USA, 1970; p. 160. [Google Scholar]
- Edwards, N.S.; Murray, A.W. Identification of Xenopus CENP-A and an associated centromeric DNA repeat. Mol. Biol. Cell 2005, 16, 1800–1810. [Google Scholar] [CrossRef] [PubMed]
- Guttenbach, M.; Nanda, I.; Feichtinger, W.; Masabanda, J.S.; Griffin, D.K.; Schmid, M. Comparative chromosome painting of chicken autosomal paints 1-9 in nine different bird Species. Cytogenet. Genome Res. 2003, 103, 173–184. [Google Scholar] [CrossRef] [PubMed]
- Shibusawa, M.; Nishibori, M.; Nishida-Umehara, C.; Tsudzuki, M.; Masabanda, J.; Griffin, D.K.; Matsuda, Y. Karyotypic evolution in the Galliformes: An examination of the process of karyotypic evolution by comparison of the molecular cytogenetic findings with the molecular phylogeny. Cytogenet. Genome Res. 2004, 106, 111–119. [Google Scholar] [CrossRef] [PubMed]
- Shibusawa, M.; Nishida-Umehara, C.; Tsudzuki, M.; Masabanda, J.; Griffin, D.K.; Matsuda, Y. A comparative karyological study of the blue-breasted quail (Coturnix chinensis, Phasianidae) and California quail (Callipepla californica, Odontophoridae). Cytogenet. Genome Res. 2004, 106, 82–90. [Google Scholar] [CrossRef] [PubMed]
- Griffin, D.K.; Robertson, L.B.; Tempest, H.G.; Vignal, A.; Fillon, V.; Crooijmans, R.P.; Groenen, M.A.; Deryusheva, S.; Gaginskaya, E.; Carré, W.; et al. Whole genome comparative studies between chicken and turkey and their implications for avian genome evolution. BMC Genom. 2008, 9, 168. [Google Scholar] [CrossRef] [PubMed]
- Shibusawa, M.; Minai, S.; Nishida-Umehara, C.; Suzuki, T.; Mano, T.; Yamada, K.; Namikawa, T.; Matsuda, Y. A comparative cytogenetic study of chromosome homology between chicken and Japanese quail. Cytogenet. Cell Genet. 2001, 95, 103–109. [Google Scholar] [CrossRef] [PubMed]
- Kayang, B.B.; Fillon, V.; Inoue-Murayama, M.; Miwa, M.; Leroux, S.; Feve, K.; Monvoisin, J.; Pitel, F.; Vignoles, M.; Mouilhayrat, C.; et al. Integrated maps in quail (Coturnix japonica) confirm the high degree of synteny conservation with chicken (Gallus gallus) despite 35 million years of divergence. BMC Genom. 2006, 7, 101–111. [Google Scholar] [CrossRef] [PubMed]
- Sasazaki, S.; Hinenoya, T.; Lin, B.; Fujiwara, A.; Mannen, H. A comparative map of macrochromosomes between chicken and Japanese quail based on orthologous genes. Anim. Genet. 2006, 37, 316–320. [Google Scholar] [CrossRef] [PubMed]
- Ryttman, H.; Tegelström, H. G-banded karyotypes of three Galliformes species, domestic fowl (Gallus domesticus), Quail (Coturnix coturnix japonica), and turkey (Meleagris gallopavo). Hereditas 1981, 94, 165–170. [Google Scholar] [CrossRef]
- Sasaki, M. High resolution G-band karyotypes of the domestic fowl and the Japanese quail. Chromosom. Inf. Sevice 1981, 31, 26–28. [Google Scholar]
- Stock, A.D.; Bunch, T.D. The evolutionary implications of chromosome banding pattern homologies in the bird order Galliformes. Cytogenet. Cell Genet. 1982, 34, 136–148. [Google Scholar] [CrossRef] [PubMed]
- Montefalcone, G.; Tempesta, S.; Rocchi, M.; Archidiacono, N. Centromere repositioning. Genome Res. 1999, 9, 184–1188. [Google Scholar] [CrossRef]
- Nagaki, K.; Cheng, Z.; Ouyang, S.; Talbert, P.B.; Kim, M.; Jones, K.M.; Henikoff, S.; Buell, C.R.; Jiang, J. Sequencing of a rice centromere uncovers active genes. Nat. Genet. 2004, 36, 138–145. [Google Scholar] [CrossRef] [PubMed]
- O’Neill, R.J.; Eldridge, M.D.; Metcalfe, C.J. Centromere dynamics and chromosome evolution in marsupials. J. Hered. 2004, 95, 375–381. [Google Scholar] [CrossRef] [PubMed]
- Han, Y.; Zhang, Z.; Liu, C.; Liu, J.; Huang, S.; Jiang, J.; Jina, W. Centromere repositioning in cucurbit species: Implication of the genomic impact from centromere activation and inactivation. Proc. Natl. Acad. Sci. USA 2009, 106, 14937–14941. [Google Scholar] [CrossRef] [PubMed]
- Piras, F.M.; Nergadze, S.G.; Magnani, E.; Bertoni, L.; Attolini, C.; Khoriauli, L.; Raimondi, E.; Giulotto, E. Uncoupling of satellite DNA and centromeric function in the genus Equus. PLoS Genet. 2010, 6, e1000845. [Google Scholar] [CrossRef] [PubMed]
- Rocchi, M.; Archidiacono, N.; Schempp, W.; Capozzi, O.; Stanyon, R. Centromere repositioning in mammals. Heredity 2012, 108, 59–67. [Google Scholar] [CrossRef] [PubMed]
- Trifonov, V.A.; Musilova, P.; Kulemsina, A.I. Chromosome Evolution in Perissodactyla. Cytogenet. Genome Res. 2012, 137, 208–217. [Google Scholar] [CrossRef] [PubMed]
- Kasai, F.; Garcia, C.; Arruga, M.V.; Ferguson-Smith, M.A. Chromosome homology between chicken (Gallus gallus domesticus) and the red-legged partridge (Alectoris rufa); evidence of the occurrence of a neocentromere during evolution. Cytogenet. Genome Res. 2003, 102, 326–330. [Google Scholar] [CrossRef] [PubMed]
- Skinner, B.M.; Robertson, L.B.W.; Tempest, H.G.; Langley, E.J.; Ioannou, D.; Fowler, K.E.; Crooijmans, R.P.M.A.; Hall, A.D.; Griffin, D.K.; Völker, M. Comparative genomics in chicken and Pekin duck using FISH mapping and microarray analysis. BMC Genom. 2009, 10, 357. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krasikova, A.; Deryusheva, S.; Galkina, S.; Kurganova, A.; Evteev, A.; Gaginskaya, E. On the positions of centromeres in chicken lampbrush chromosomes. Chromosom. Res. 2006, 14, 777–789. [Google Scholar] [CrossRef] [PubMed]
- Calderon, P.L.; Pigozzi, M.I. MLH1-focus mapping in birds shows equal recombination between sexes and diversity of crossover patterns. Chromosom. Res. 2006, 14, 605–612. [Google Scholar] [CrossRef] [PubMed]
- Solari, A.J. Synaptosomal complexes and associated structures in microspread human spermatocytes. Chromosoma 1980, 81, 315–337. [Google Scholar] [CrossRef] [PubMed]
- Kaelbling, M.; Fechheimer, N.S. Synaptonemal complexes and the chromosomes of the domestic fowl, Gallus domesticus. Cytogenet. Cell Genet. 1983, 35, 87–92. [Google Scholar] [CrossRef] [PubMed]
- Krasikova, A.; Daks, A.; Zlotina, A.; Gaginskaya, E. Polymorphic heterochromatic segments in Japanese quail microchromosomes. Cytogenet. Genome Res. 2009, 126, 148–155. [Google Scholar] [CrossRef] [PubMed]
- Völker, M.; Backstrom, N.; Skinner, B.M.; Langley, E.J.; Bunzey, S.K.; Ellegren, H.; Griffin, D.K. Copy number variation, chromosome rearrangement, and their association with recombination during avian evolution. Genome Res. 2010, 20, 503–511. [Google Scholar] [CrossRef] [PubMed]
- Skinner, B.M.; Griffin, D.K. Intrachromosomal rearrangements in avian genome evolution: Evidence for regions prone to breakpoints. Heredity 2012, 108, 37–41. [Google Scholar] [CrossRef] [PubMed]
- Lithgow, P.E.; O’Connor, R.; Smith, D.; Fonseka, G.; Mutery, A.A.; Rathje, C.; Frodsham, R.; O’Brien, P.; Kasai, F.; Ferguson-Smith, M.A.; et al. Novel tools for characterising inter and intra chromosomal rearrangements in avian microchromosomes. Chromosom. Res. 2014, 22, 85–97. [Google Scholar] [CrossRef] [PubMed]
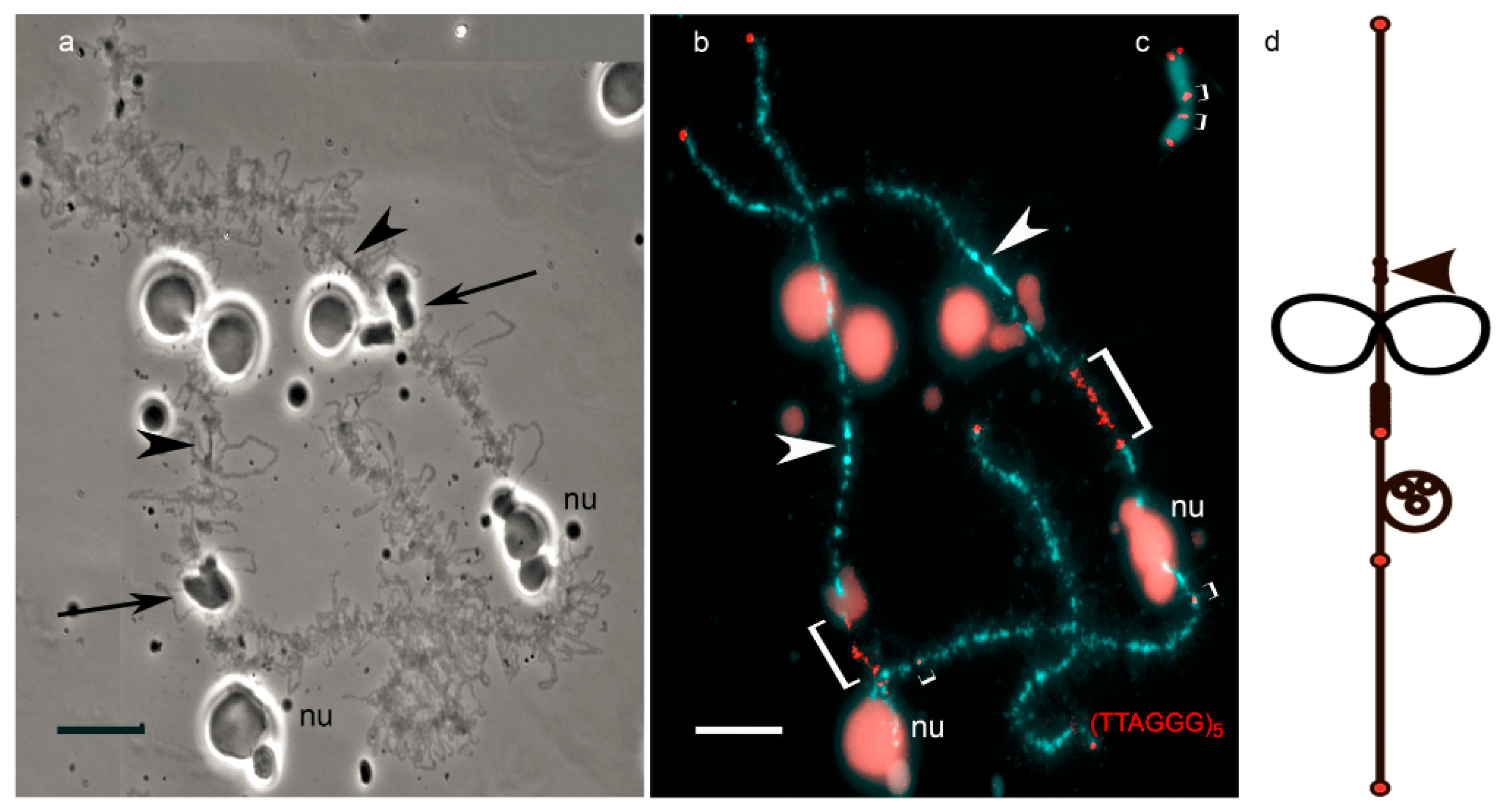
Figure 1.
Comparison of the usage of amphibian lampbrush chromosome (a,b) and metaphase chromosome (c), for high-resolution fluorescent in situ hybridization (FISH)-mapping. Lampbrush chromosomes exhibit marker loops (indicated by arrows in (a)) and other landmarks (including nucleolus, nu in (a,b)), which allows to construct cytological chromosomal maps (d). Such maps facilitate identification of individual chromosomes and their particular regions. Mapping of interstitial telomeric sites (shown by square brackets) in: lampbrush chromosome (b); and metaphase chromosome (c). Arrowheads indicate centromeres. Chromosomes are counterstained with DAPI (4′ 6-diamidino-2-phenylindole). Scale bar = 10 µm.
Figure 1.
Comparison of the usage of amphibian lampbrush chromosome (a,b) and metaphase chromosome (c), for high-resolution fluorescent in situ hybridization (FISH)-mapping. Lampbrush chromosomes exhibit marker loops (indicated by arrows in (a)) and other landmarks (including nucleolus, nu in (a,b)), which allows to construct cytological chromosomal maps (d). Such maps facilitate identification of individual chromosomes and their particular regions. Mapping of interstitial telomeric sites (shown by square brackets) in: lampbrush chromosome (b); and metaphase chromosome (c). Arrowheads indicate centromeres. Chromosomes are counterstained with DAPI (4′ 6-diamidino-2-phenylindole). Scale bar = 10 µm.
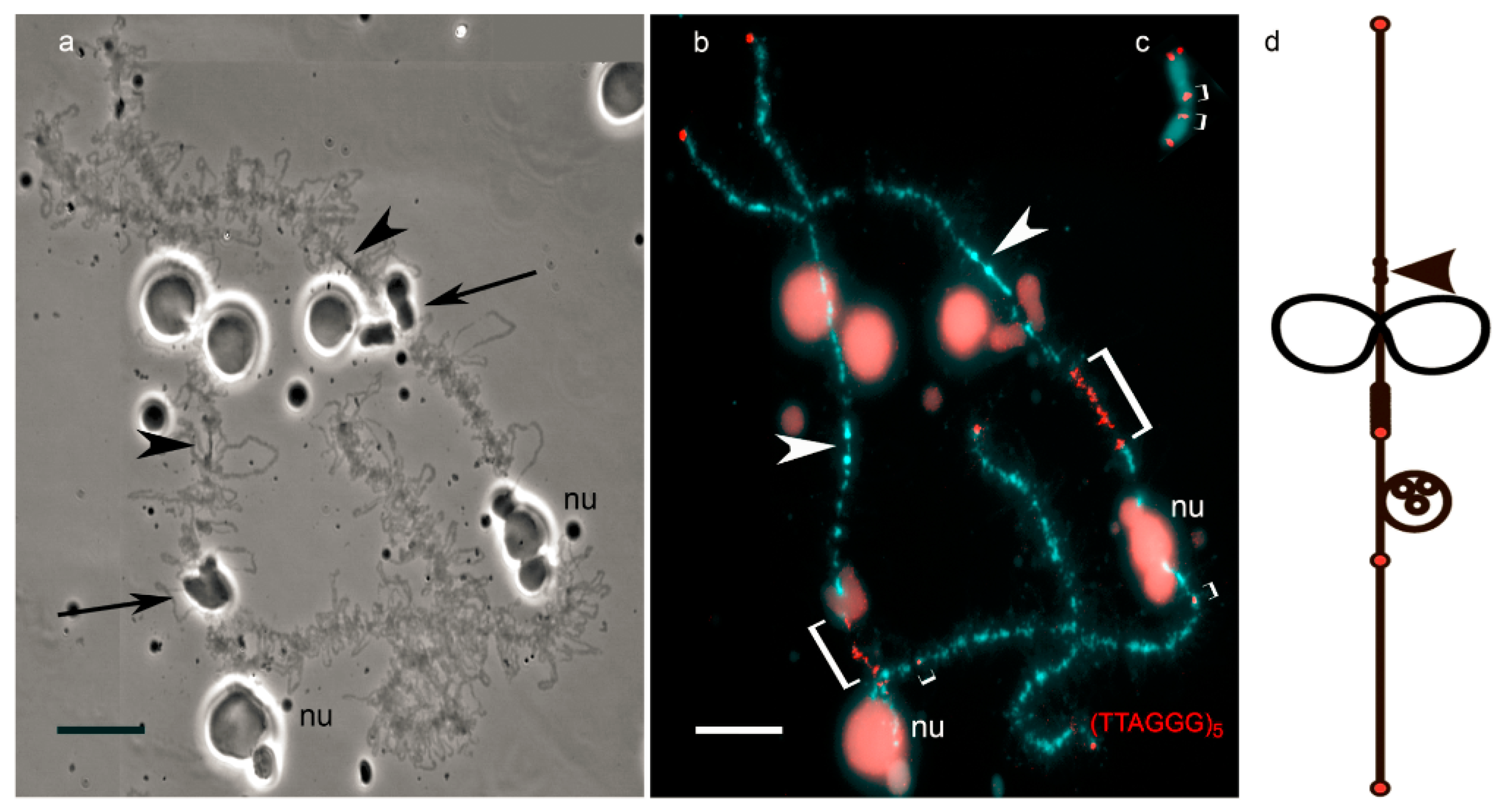
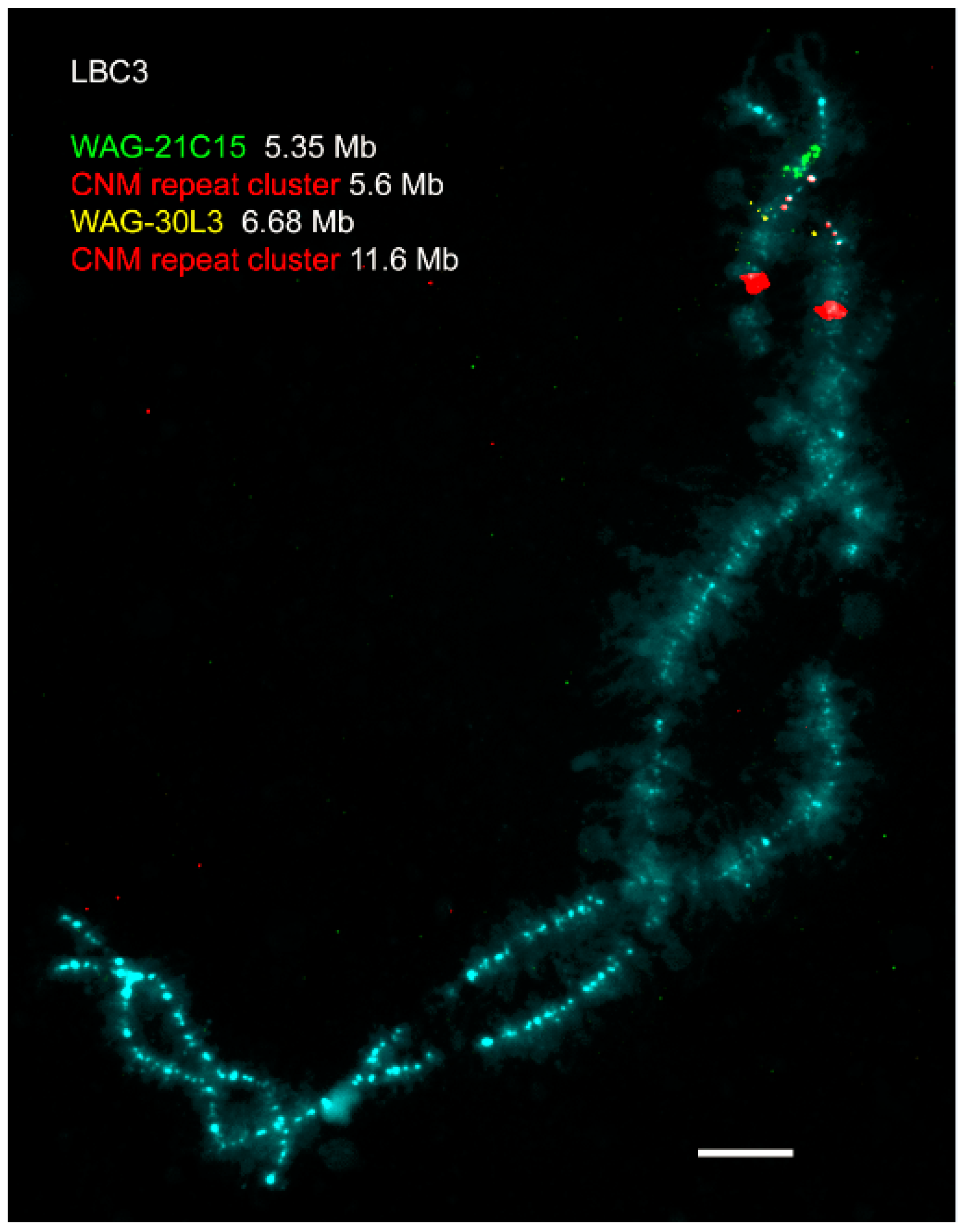
Figure 2.
High-resolution FISH-mapping on avian lampbrush chromosomes (an example). FISH with chicken nuclear-membrane-associated repeat (CNM repeat)-specific probe (red) and bacterial artificial chromosome (BAC) clones WAG12C15 (green) and WAG30L03 (yellow) on chicken lampbrush chromosome 3. Chromosome is counterstained with DAPI. Scale bar = 10 μm. Chromosome coordinates of BACs and CNM clusters are given in megabases (Mb) according to the chicken genome assembly Gallus_gallus-5.0 (https://www.ncbi.nlm.nih.gov/genome/111) [26]. BACs were kindly provided by Richard Crooijmans and Martin Groenen (Wageningen chicken BAC library, Crooijmans et al., 2000 [29]). The data on precise genome positioning of the centromere and two CNM-repeat clusters from chicken chromosome 3 are published in Zlotina et al., 2010 [54].
Figure 2.
High-resolution FISH-mapping on avian lampbrush chromosomes (an example). FISH with chicken nuclear-membrane-associated repeat (CNM repeat)-specific probe (red) and bacterial artificial chromosome (BAC) clones WAG12C15 (green) and WAG30L03 (yellow) on chicken lampbrush chromosome 3. Chromosome is counterstained with DAPI. Scale bar = 10 μm. Chromosome coordinates of BACs and CNM clusters are given in megabases (Mb) according to the chicken genome assembly Gallus_gallus-5.0 (https://www.ncbi.nlm.nih.gov/genome/111) [26]. BACs were kindly provided by Richard Crooijmans and Martin Groenen (Wageningen chicken BAC library, Crooijmans et al., 2000 [29]). The data on precise genome positioning of the centromere and two CNM-repeat clusters from chicken chromosome 3 are published in Zlotina et al., 2010 [54].
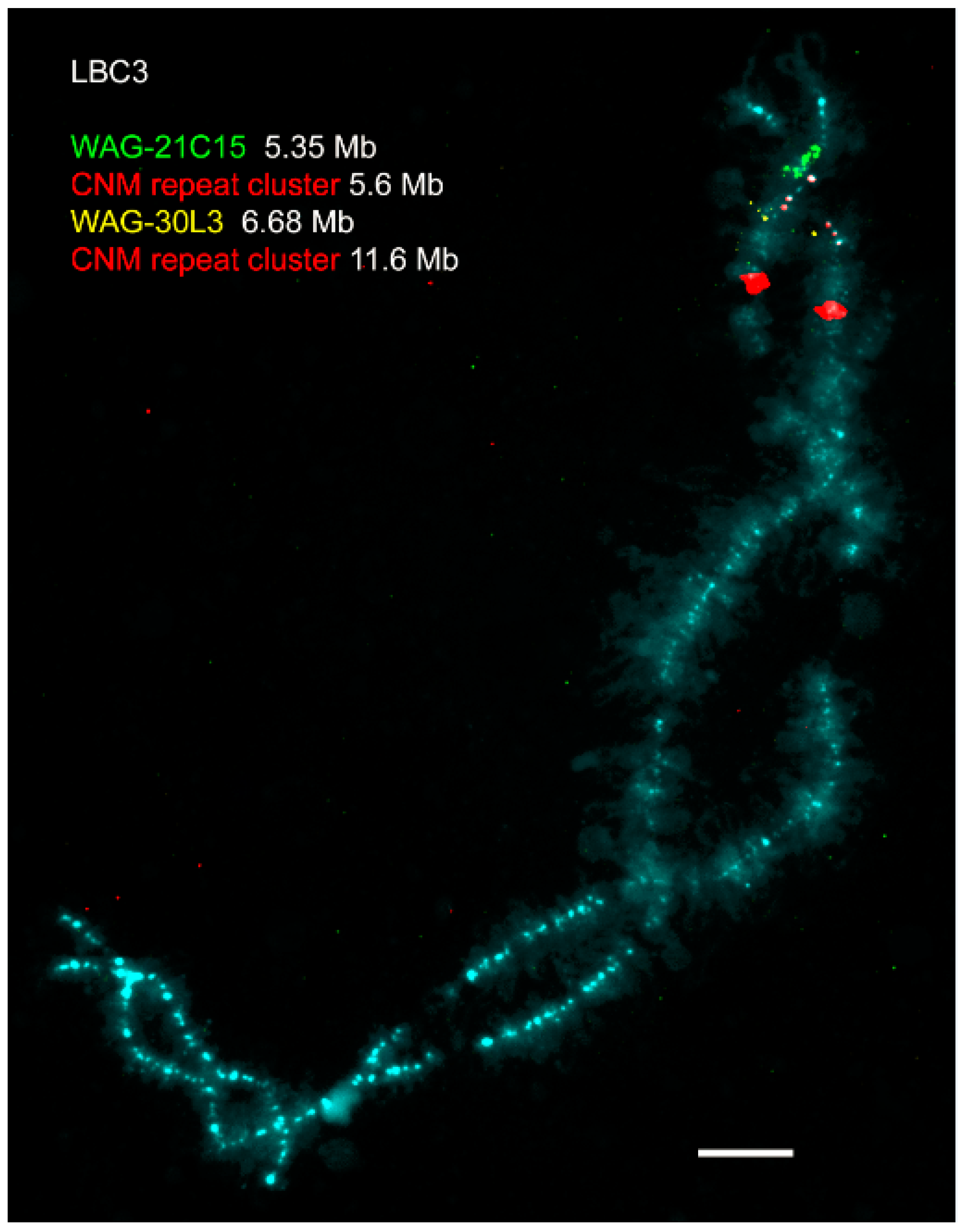
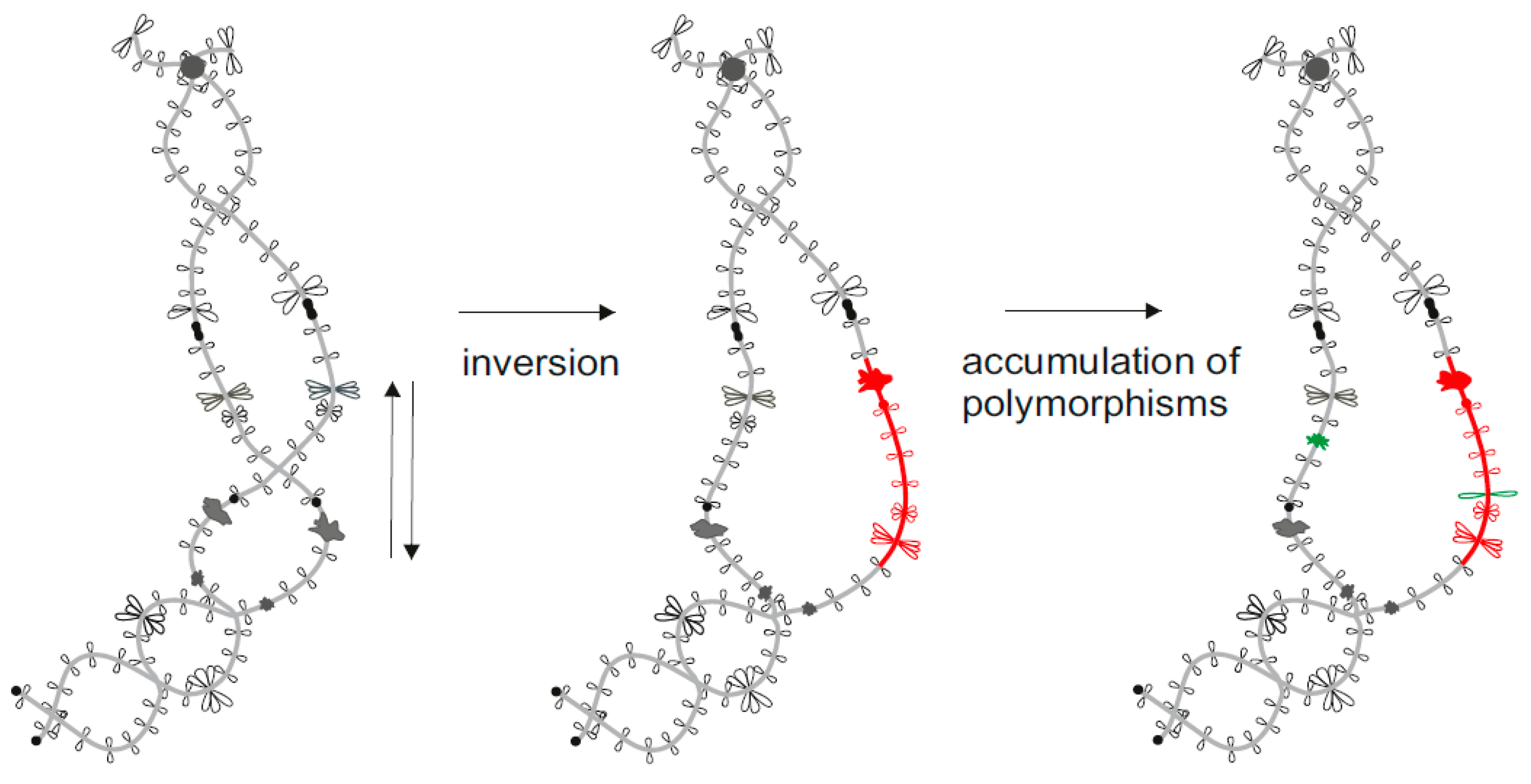
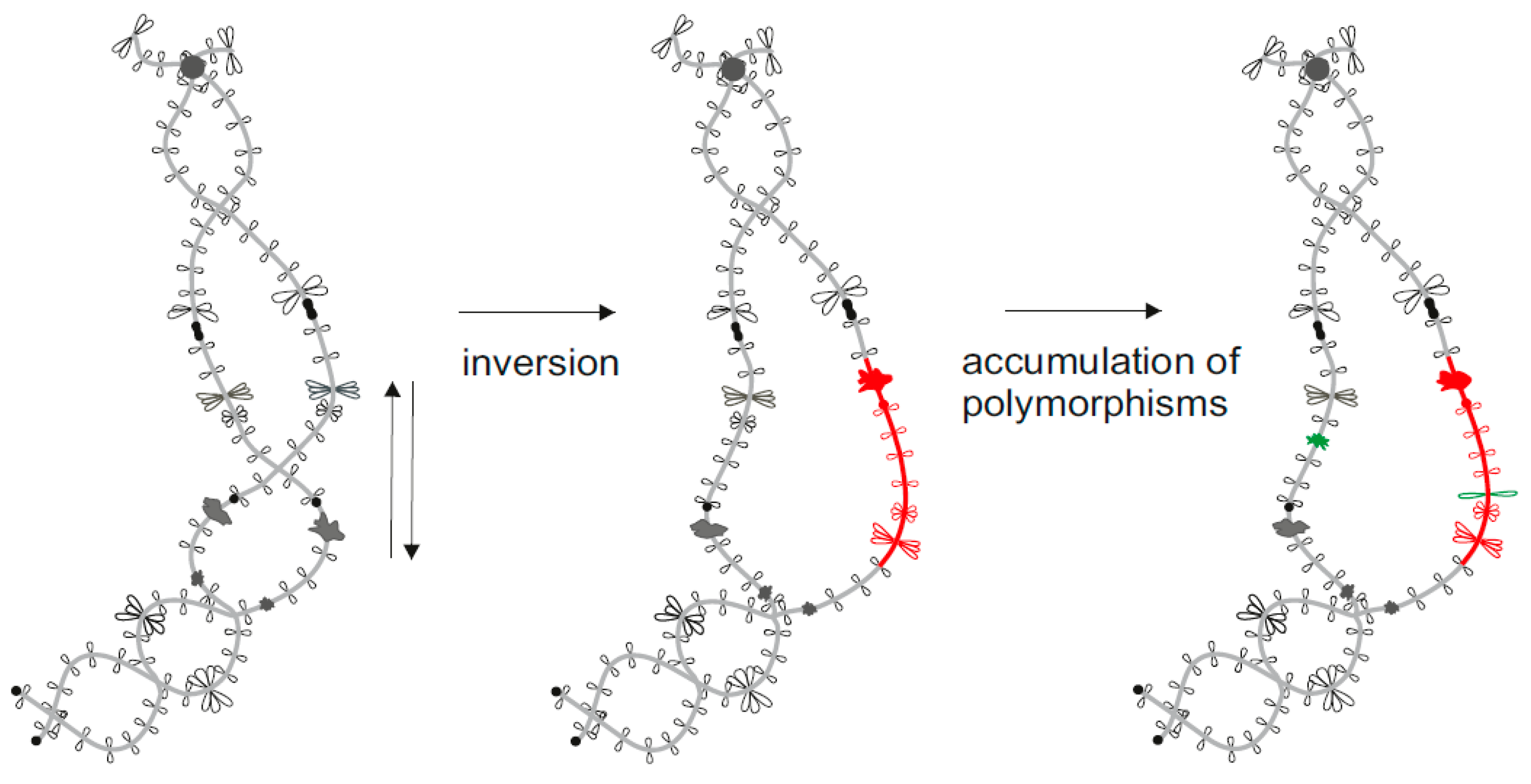
Figure 3.
Example of lampbrush chromosome application to study chromosomal rearrangements. Inverted order of marker structures (special loops and granules) indicates the inversion of a chromosomal segment. Additional marker structures appear after accumulation of genetic polymorphisms in the absence of recombination.
Figure 3.
Example of lampbrush chromosome application to study chromosomal rearrangements. Inverted order of marker structures (special loops and granules) indicates the inversion of a chromosomal segment. Additional marker structures appear after accumulation of genetic polymorphisms in the absence of recombination.
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Zlotina, A.; Dedukh, D.; Krasikova, A. Amphibian and Avian Karyotype Evolution: Insights from Lampbrush Chromosome Studies. Genes 2017, 8, 311. https://doi.org/10.3390/genes8110311
AMA Style
Zlotina A, Dedukh D, Krasikova A. Amphibian and Avian Karyotype Evolution: Insights from Lampbrush Chromosome Studies. Genes. 2017; 8(11):311. https://doi.org/10.3390/genes8110311
Chicago/Turabian StyleZlotina, Anna, Dmitry Dedukh, and Alla Krasikova. 2017. "Amphibian and Avian Karyotype Evolution: Insights from Lampbrush Chromosome Studies" Genes 8, no. 11: 311. https://doi.org/10.3390/genes8110311
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.