Role of Non-Coding RNAs in the Etiology of Bladder Cancer


Abstract
:1. Introduction
2. Classification and Etiology of BC
3. Epigenetics and BC
4. Non-Coding RNAs: An Overview
5. Long Non-Coding RNA
5.1. UCA1
5.2. MALAT1
5.3. H19
5.4. TUG1
5.5. MEG3
5.6. MIR31HG
5.7. Linc-UBC1
5.8. LOC572558
5.9. PANDAR
5.10. GHET1
5.11. ncRAN
5.12. GAS5
5.13. ANRIL
5.14. HIF1A-AS2
5.15. HOTAIR and HOXD-AS1
5.16. MDC1-AS1
5.17. PCAT-1
5.18. PVT1
5.19. SChLAP1
5.20. SPRY4-IT1
5.21. ZEB2-AS1
5.22. T-UCR 8+
5.23. NEAT1
5.24. Other lncRNA Deregulated in Bladder Cancer
6. Small Non-Coding RNA
6.1. Micro-RNA and Bladder Cancer
6.2. The FGFR3 Pathway
6.3. The TP53 Pathway
6.4. Other Molecular Pathways Causing miR-Mediated BC Formation
6.5. New Approaches for Assessing the miR-Mediated BC Formation
7. Other ncRNA Involved in BC Etiology
7.1. Y RNA
7.2. circRNA
8. Conclusions
Author Contributions
Conflicts of Interest
References
- US National Cancer Institute (NCI). Available online: http://www.cancer.gov (accessed on 19 May 2017).
- Erlich, A.; Zlotta, A.R. Treatment of bladder cancer in the elderly. Investig. Clin. Urol. 2016, 57, S26–S35. [Google Scholar] [CrossRef] [PubMed]
- Pasin, E.; Josephson, D.Y.; Mitra, A.P.; Cote, R.J.; Stein, J.P. Superficial bladder cancer: An update on etiology, molecular development, classification, and natural history. Rev. Urol. 2008, 10, 31–43. [Google Scholar] [PubMed]
- Brooks, N.A.; O’Donnell, M.A. Treatment options in non-muscle-invasive bladder cancer after BCG failure. Indian J. Urol. 2015, 31, 312–319. [Google Scholar] [CrossRef] [PubMed]
- Scarpato, K.R.; Morgans, A.K.; Moses, K.A. Optimal management of muscle-invasive bladder cancer—A review. Res. Rep. Urol. 2015, 7, 143–151. [Google Scholar] [CrossRef] [PubMed]
- Li, H.-T.; Duymich, C.E.; Weisenberger, D.J.; Liang, G. Genetic and epigenetic alterations in bladder cancer. Int. Neurourol. J. 2016, 20, S84–S94. [Google Scholar] [CrossRef] [PubMed]
- Hernández, S.; López-Knowles, E.; Lloreta, J.; Kogevinas, M.; Jaramillo, R.; Amorós, A.; Tardón, A.; García-Closas, R.; Serra, C.; Carrato, A.; et al. FGFR3 and Tp53 mutations in T1G3 transitional bladder carcinomas: Independent distribution and lack of association with prognosis. Clin. Cancer Res. 2005, 11, 5444–5450. [Google Scholar] [CrossRef] [PubMed]
- McKnight, J.J.; Gray, S.B.; O’Kane, H.F.; Johnston, S.R.; Williamson, K.E. Apoptosis and chemotherapy for bladder cancer. J. Urol. 2005, 173, 683–690. [Google Scholar] [CrossRef] [PubMed]
- Margulis, V.; Lotan, Y.; Montorsi, F.; Shariat, S.F. Predicting survival after radical cystectomy for bladder cancer. BJU Int. 2008, 102, 15–22. [Google Scholar] [CrossRef] [PubMed]
- Yamana, K.; Bilim, V.; Hara, N.; Kasahara, T.; Itoi, T.; Maruyama, R.; Nishiyama, T.; Takahashi, K.; Tomita, Y. Prognostic impact of FAS/CD95/APO-1 in urothelial cancers: Decreased expression of Fas is associated with disease progression. Br. J. Cancer 2005, 93, 544–551. [Google Scholar] [CrossRef] [PubMed]
- Karam, J.A.; Lotan, Y.; Karakiewicz, P.I.; Ashfaq, R.; Sagalowsky, A.I.; Roehrborn, C.G.; Shariat, S.F. Use of combined apoptosis biomarkers for prediction of bladder cancer recurrence and mortality after radical cystectomy. Lancet Oncol. 2007, 8, 128–136. [Google Scholar] [CrossRef]
- Shariat, S.F.; Ashfaq, R.; Karakiewicz, P.I.; Saeedi, O.; Sagalowsky, A.I.; Lotan, Y. Survivin expression is associated with bladder cancer presence, stage, progression, and mortality. Cancer 2007, 109, 1106–1113. [Google Scholar] [CrossRef] [PubMed]
- Shariat, S.F.; Karam, J.A.; Lerner, S.P. Molecular markers in bladder cancer. Curr. Opin. Urol. 2008, 18, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Shariat, S.F.; Youssef, R.F.; Gupta, A.; Chade, D.C.; Karakiewicz, P.I.; Isbarn, H.; Jeldres, C.; Sagalowsky, A.I.; Ashfaq, R.; Lotan, Y. Association of angiogenesis related markers with bladder cancer outcomes and other molecular markers. J. Urol. 2010, 183, 1744–1750. [Google Scholar] [CrossRef] [PubMed]
- Huang, H.-Y.; Shariat, S.F.; Sun, T.-T.; Lepor, H.; Shapiro, E.; Hsieh, J.-T.; Ashfaq, R.; Lotan, Y.; Wu, X.-R. Persistent uroplakin expression in advanced urothelial carcinomas: Implications in urothelial tumor progression and clinical outcome. Hum. Pathol. 2007, 38, 1703–1713. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.T.; Pak, J.; Huang, H.Y.; Shapiro, E.; Sun, T.T.; Pellicer, A.; Wu, X.R. Role of Ha-ras activation in superficial papillary pathway of urothelial tumor formation. Oncogene 2001, 20, 1973–1980. [Google Scholar] [CrossRef] [PubMed]
- Di Pierro, G.B.; Gulia, C.; Cristini, C.; Fraietta, G.; Marini, L.; Grande, P.; Gentile, V.; Piergentili, R. Bladder cancer: A simple model becomes complex. Curr. Genom. 2012, 13, 395–415. [Google Scholar] [CrossRef] [PubMed]
- Malats, N.; Real, F.X. Epidemiology of bladder cancer. Hematol. Oncol. Clin. N. Am. 2015, 29, 177–189. [Google Scholar] [CrossRef] [PubMed]
- Zeegers, M.P.; Tan, F.E.; Dorant, E.; van den Brandt, P.A. The impact of characteristics of cigarette smoking on urinary tract cancer risk: A meta-analysis of epidemiologic studies. Cancer 2000, 89, 630–639. [Google Scholar] [CrossRef]
- Sun, X.; Deng, Q.; Liang, Z.; Liu, Z.; Geng, H.; Zhao, L.; Zhou, Q.; Liu, J.; Ma, J.; Wang, D.; et al. Cigarette smoke extract induces epithelial-mesenchymal transition of human bladder cancer T24 cells through activation of ERK1/2 pathway. Biomed. Pharmacother. 2017, 86, 457–465. [Google Scholar] [CrossRef] [PubMed]
- Shaham, J.; Melzer, A.; Kaufman, Z.; Ribak, J. Occupation and bladder cancer. Harefuah 1996, 131, 382–386. [Google Scholar] [PubMed]
- Marrett, L.D.; Hartge, P.; Meigs, J.W. Bladder cancer and occupational exposure to leather. Br. J. Ind. Med. 1986, 43, 96–100. [Google Scholar] [CrossRef] [PubMed]
- Silverman, D.T.; Hartge, P.; Morrison, A.S.; Devesa, S.S. Epidemiology of bladder cancer. Hematol. Oncol. Clin. N. Am. 1992, 6, 1–30. [Google Scholar]
- Zhang, Y.; Kim, C.; Zheng, T. Hair dye use and risk of human cancer. Front. Biosci. (Elite Ed) 2012, 4, 516–528. [Google Scholar] [CrossRef] [PubMed]
- Huncharek, M.; Kupelnick, B. Personal use of hair dyes and the risk of bladder cancer: Results of a meta-analysis. Public Health Rep. 2005, 120, 31–38. [Google Scholar] [CrossRef] [PubMed]
- Letašiová, S.; Medveďová, A.; Šovčíková, A.; Dušinská, M.; Volkovová, K.; Mosoiu, C.; Bartonová, A. Bladder cancer, a review of the environmental risk factors. Environ. Health 2012, 11, S11. [Google Scholar] [CrossRef] [PubMed]
- Saint-Jacques, N.; Parker, L.; Brown, P.; Dummer, T.J.B. Arsenic in drinking water and urinary tract cancers: A systematic review of 30 years of epidemiological evidence. Environ. Health 2014, 13, 44. [Google Scholar] [CrossRef] [PubMed]
- Mostafa, M.H.; Sheweita, S.A.; O’Connor, P.J. Relationship between schistosomiasis and bladder cancer. Clin. Microb. Rev. 1999, 12, 97–111. [Google Scholar]
- Handy, D.E.; Castro, R.; Loscalzo, J. Epigenetic modifications: Basic mechanisms and role in cardiovascular disease. Circulation 2011, 123, 2145–2156. [Google Scholar] [CrossRef] [PubMed]
- Williams, S.C.P. Epigenetics. Proc. Natl. Acad. Sci. USA 2013, 110, 3209. [Google Scholar] [CrossRef] [PubMed]
- Jaenisch, R.; Bird, A. Epigenetic regulation of gene expression: How the genome integrates intrinsic and environmental signals. Nat. Genet. 2003, 33, 245–254. [Google Scholar] [CrossRef] [PubMed]
- Murrell, A.; Hurd, P.J.; Wood, I.C. Epigenetic mechanisms in development and disease. Biochem. Soc. Trans. 2013, 41, 697–699. [Google Scholar] [CrossRef] [PubMed]
- Bannister, A.J.; Kouzarides, T. Regulation of chromatin by histone modifications. Cell Res. 2011, 21, 381–395. [Google Scholar] [CrossRef] [PubMed]
- Collins, L.J.; Schönfeld, B.; Che, X.S. The Epigenetics of Non-coding RNA. In Handbook of Epigenetics: The New Molecular and Medical Genetics, 1st ed.; Tollefsbol, T., Ed.; Academic Press: Cambridge, MA, USA, 2010; pp. 49–61. [Google Scholar]
- Wang, Y.H.; Hu, H.N.; Weng, H.; Chen, H.; Luo, C.L.; Ji, J.; Yin, C.Q.; Yuan, C.H.; Wang, F.B. Association between polymorphisms in microRNAs and risk of urological cancer: A meta-analysis based on 17,019 subjects. Front. Physiol. 2017, 8, 325. [Google Scholar] [CrossRef] [PubMed]
- Arrigo, P.; Pulliero, A. Effect of environmental chemical stress on nuclear noncoding RNA involved in epigenetic control. Biomed. Res. Int. 2015, 2015, 761703. [Google Scholar] [CrossRef] [PubMed]
- IARC Monographs on the Evaluation of Carcinogenic Risks to Humans. International Agency for Research on Cancer. Available online: https://monographs.iarc.fr (accessed on 2 August 2017).
- Ponting, C.P.; Hardison, R.C. What fraction of the human genome is functional? Genome Res. 2011, 21, 1769–1776. [Google Scholar] [CrossRef] [PubMed]
- Ling, H.; Vincent, K.; Pichler, M.; Fodde, R.; Berindan-Neagoe, I.; Slack, F.J.; Calin, G.A. Junk DNA and the long non-coding RNA twist in cancer genetics. Oncogene 2015, 34, 5003–5011. [Google Scholar] [CrossRef] [PubMed]
- Cao, J. The functional role of long non-coding RNAs and epigenetics. Biol. Proced. Online 2014, 16, 11. [Google Scholar] [CrossRef] [PubMed]
- Bhan, A.; Soleimani, M.; Mandal, S.S. Long noncoding RNA and cancer: A new paradigm. Cancer Res. 2017, 77, 3965–3981. [Google Scholar] [CrossRef] [PubMed]
- Peng, J.C.; Shen, J.; Ran, Z.H. Transcribed ultraconserved region in human cancers. RNA Biol. 2013, 10, 1771–1777. [Google Scholar] [CrossRef] [PubMed]
- Jeck, W.R.; Sharpless, N.E. Detecting and characterizing circular RNAs. Nat. Biotechnol. 2014, 32, 453–461. [Google Scholar] [CrossRef] [PubMed]
- Carthew, R.W.; Sontheimer, E.J. Origins and mechanisms of miRNAs and siRNAs. Cell 2009, 136, 642–655. [Google Scholar] [CrossRef] [PubMed]
- Hall, A.E.; Turnbull, C.; Dalmay, T. Y RNAs: Recent developments. Biomol. Concepts 2013, 4, 103–110. [Google Scholar] [CrossRef] [PubMed]
- Ha, M.; Kim, V.N. Regulation of microRNA biogenesis. Nat. Rev. Mol. Cell Biol. 2014, 15, 509–524. [Google Scholar] [CrossRef] [PubMed]
- Sohel, M.H. Extracellular/circulating microRNAs: Release mechanisms, functions and challenges. Achiev. Life Sci. 2016, 10, 175–186. [Google Scholar] [CrossRef]
- Siomi, M.C.; Sato, K.; Pezic, D.; Aravin, A.A. PIWI-interacting small RNAs: The vanguard of genome defence. Nat. Rev. Mol. Cell. Biol. 2011, 12, 246–258. [Google Scholar] [CrossRef] [PubMed]
- Bratkovič, T.; Rogelj, B. The many faces of small nucleolar RNAs. Biochim. Biophys. Acta 2014, 1839, 438–443. [Google Scholar] [CrossRef] [PubMed]
- Matera, A.G.; Terns, R.M.; Terns, M.P. Non-coding RNAs: Lessons from the small nuclear and small nucleolar RNAs. Nat. Rev. Mol. Cell Biol. 2007, 8, 209–220. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Peng, G. Non-coding RNAs: An emerging player in DNA damage response. Mutat. Res. Rev. Mutat. Res. 2015, 763, 202–211. [Google Scholar] [CrossRef] [PubMed]
- D’Adda di Fagagna, F. A direct role for small non-coding RNAs in DNA damage response. Trends Cell Biol. 2014, 24, 171–178. [Google Scholar] [CrossRef] [PubMed]
- Okada, H.; Hayashizaki, Y. World of functional RNA. Rinsho Shinkeigaku 2013, 53, 957–961. [Google Scholar] [CrossRef] [PubMed]
- Martens-Uzunova, E.S.; Böttcher, R.; Croce, C.M.; Jenster, G.; Visakorpi, T.; Calin, G.A. Long noncoding RNA in prostate, bladder, and kidney cancer. Eur. Urol. 2014, 65, 1140–1151. [Google Scholar] [CrossRef] [PubMed]
- Sültmann, H.; Diederichs, S. Long noncoding RNA: “LNCs” to cancer. Eur. Urol. 2014, 65, 1152–1153. [Google Scholar] [CrossRef] [PubMed]
- Olivieri, M.; Ferro, M.; Terreri, S.; Durso, M.; Romanelli, A.; Avitabile, C.; De Cobelli, O.; Messere, A.; Bruzzese, D.; Vannini, I.; et al. Long non-coding RNA containing ultraconserved genomic region 8 promotes bladder cancer tumorigenesis. Oncotarget 2016, 7, 20636–20654. [Google Scholar] [CrossRef] [PubMed]
- Ning, S.; Zhang, J.; Wang, P.; Zhi, H.; Wang, J.; Liu, Y.; Gao, Y.; Guo, M.; Yue, M.; Wang, L.; et al. Lnc2Cancer: A manually curated database of experimentally supported lncRNAs associated with various human cancers. Nucleic Acids Res. 2016, 44, D980–D985. Available online: http://www.bio-bigdata.com/lnc2cancer/home.jsp (accessed on 17 October 2017). [CrossRef] [PubMed]
- GeneCards: The Human Gene Database. Available online: http://www.genecards.org (accessed on 27 July 2017).
- Online Mendelian Inheritance in Man (OMIM). Available online: https://www.omim.org/ (accessed on 26 July 2017).
- Yang, C.; Li, X.; Wang, Y.; Zhao, L.; Chen, W. Long non-coding RNA UCA1 regulated cell cycle distribution via CREB through PI3-K dependent pathway in bladder carcinoma cells. Gene 2012, 496, 8–16. [Google Scholar] [CrossRef] [PubMed]
- Xue, M.; Li, X.; Li, Z.; Chen, W. Urothelial carcinoma associated 1 is a hypoxia-inducible factor-1α-targeted long noncoding RNA that enhances hypoxic bladder cancer cell proliferation, migration, and invasion. Tumour Biol. 2014, 35, 6901–6912. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Jiang, Y.; Wan, Y.; Zhang, L.; Qiu, J.; Zhou, S.; Cheng, W. UCA1 functions as a competing endogenous RNA to suppress epithelial ovarian cancer metastasis. Tumour Biol. 2016, 37, 10633–10641. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.; Li, Q.; Wang, S.; Zhang, J. miR-485-5p inhibits bladder cancer metastasis by targeting HMGA2. Int. J. Mol. Med. 2015, 36, 1136–1142. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.-S. Rapid identification of UCA1 as a very sensitive and specific unique marker for human bladder carcinoma. Clin. Cancer Res. 2006, 12, 4851–4858. [Google Scholar] [CrossRef] [PubMed]
- Cui, X.; Jing, X.; Long, C.; Yi, Q.; Tian, J.; Zhu, J. Accuracy of the urine UCA1 for diagnosis of bladder cancer: A meta-analysis. Oncotarget 2017, 8, 35222–35233. [Google Scholar] [CrossRef] [PubMed]
- Milowich, D.; Le Mercier, M.; de Neve, N.; Sandras, F.; Roumeguere, T.; Decaestecker, C.; Salmon, I.; Rorive, S. Diagnostic value of the UCA1 test for bladder cancer detection: a clinical study. SpringerPlus 2015, 4, 349. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Peng, F.; Cheng, L.; Yang, G.; Zhang, D.; Liu, J.; Chen, X.; Zhao, S. Prognostic and clinicopathological role of long non-coding RNA UCA1 in various carcinomas. Oncotarget 2017, 8, 28373–28384. [Google Scholar] [CrossRef] [PubMed]
- Lamouille, S.; Xu, J.; Derynck, R. Molecular mechanisms of epithelial-mesenchymal transition. Nat. Rev. Mol. Cell Biol. 2014, 15, 178–196. [Google Scholar] [CrossRef] [PubMed]
- Chiyomaru, T.; Enokida, H.; Tatarano, S.; Kawahara, K.; Uchida, Y.; Nishiyama, K.; Fujimura, L.; Kikkawa, N.; Seki, N.; Nakagawa, M. miR-145 and miR-133a function as tumour suppressors and directly regulate FSCN1 expression in bladder cancer. Br. J. Cancer 2010, 102, 883–891. [Google Scholar] [CrossRef] [PubMed]
- Xue, M.; Pang, H.; Li, X.; Li, H.; Pan, J.; Chen, W. Long non-coding RNA urothelial cancer-associated 1 promotes bladder cancer cell migration and invasion by way of the hsa-miR-145-ZEB1/2-FSCN1 pathway. Cancer Sci. 2016, 107, 18–27. [Google Scholar] [CrossRef] [PubMed]
- Hutchinson, J.N.; Ensminger, A.W.; Clemson, C.M.; Lynch, C.R.; Lawrence, J.B.; Chess, A. A screen for nuclear transcripts identifies two linked noncoding RNAs associated with SC35 splicing domains. BMC Genomics 2007, 8, 39. [Google Scholar] [CrossRef] [PubMed]
- Wilusz, J.E.; Freier, S.M.; Spector, D.L. 3’ End processing of a long nuclear-retained noncoding RNA yields a tRNA-like cytoplasmic RNA. Cell 2008, 135, 919–932. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Huang, C.; Meng, X.; Li, J. Long noncoding RNA MALAT1: Insights into its biogenesis and implications in human disease. Curr. Pharm. Des. 2015, 21, 5017–5028. [Google Scholar] [CrossRef] [PubMed]
- Tian, X.; Xu, G. Clinical value of lncRNA MALAT1 as a prognostic marker in human cancer: Systematic review and meta-analysis. BMJ Open 2015, 5, e008653. [Google Scholar] [CrossRef] [PubMed]
- Wei, Y.; Niu, B. Role of MALAT1 as a prognostic factor for survival in various cancers: A systematic review of the literature with meta-analysis. Dis. Markers 2015, 2015, 164635. [Google Scholar] [CrossRef] [PubMed]
- Ying, L.; Chen, Q.; Wang, Y.; Zhou, Z.; Huang, Y.; Qiu, F. Upregulated MALAT-1 contributes to bladder cancer cell migration by inducing epithelial-to-mesenchymal transition. Mol. Biosyst. 2012, 8, 2289–2294. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Lu, W.; Xu, J.; Shi, Y.; Zhang, H.; Xia, D. Prognostic value of long non-coding RNA MALAT1 in cancer patients. Tumour Biol. 2016, 37, 897–903. [Google Scholar] [CrossRef] [PubMed]
- Smits, G.; Mungall, A.J.; Griffiths-Jones, S.; Smith, P.; Beury, D.; Matthews, L.; Rogers, J.; Pask, A.J.; Shaw, G.; VandeBerg, J.L.; et al. Conservation of the H19 noncoding RNA and H19-IGF2 imprinting mechanism in therians. Nat. Genet. 2008, 40, 971–976. [Google Scholar] [CrossRef] [PubMed]
- Gabory, A.; Jammes, H.; Dandolo, L. The H19 locus: Role of an imprinted non-coding RNA in growth and development. Bioessays 2010, 32, 473–480. [Google Scholar] [CrossRef] [PubMed]
- Ariel, I.; Sughayer, M.; Fellig, Y.; Pizov, G.; Ayesh, S.; Podeh, D.; Libdeh, B.A.; Levy, C.; Birman, T.; Tykocinski, M.L.; et al. The imprinted H19 gene is a marker of early recurrence in human bladder carcinoma. Mol. Pathol. 2000, 53, 320–323. [Google Scholar] [CrossRef] [PubMed]
- Byun, H.-M.; Wong, H.-L.; Aaron, E.A.; Wolff, E.M.; Ling, G.; Yang, A.S. Examination of IGF2 and H19 loss of imprinting in bladder cancer. Cancer Res 2007, 67, 10753–10758. [Google Scholar] [CrossRef] [PubMed]
- Ariel, I.; de Groot, N.; Hochberg, A. Imprinted H19 gene expression in embryogenesis and human cancer: The oncofetal connection. Am. J. Med. Genet. 2000, 91, 46–50. [Google Scholar] [CrossRef]
- Huang, M.; Zhong, Z.; Lv, M.; Shu, J.; Tian, Q.; Chen, J. Comprehensive analysis of differentially expressed profiles of lncRNAs and circRNAs with associated co-expression and ceRNA networks in bladder carcinoma. Oncotarget 2016, 7, 47186–47200. [Google Scholar] [CrossRef] [PubMed]
- Gielchinsky, I.; Gilon, M.; Abu-Lail, R.; Matouk, I.; Hochberg, A.; Gofrit, O.N.; Ben-Dov, I.Z. H19 non-coding RNA in urine cells detects urothelial carcinoma: A pilot study. Biomarkers 2017, 22, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Cai, X.; Cullen, B.R. The imprinted H19 noncoding RNA is a primary microRNA precursor. RNA 2007, 13, 313–316. [Google Scholar] [CrossRef] [PubMed]
- Bartholdi, D.; Krajewska-Walasek, M.; Ounap, K.; Gaspar, H.; Chrzanowska, K.H.; Ilyana, H.; Kayserili, H.; Lurie, I.W.; Schinzel, A.; Baumer, A. Epigenetic mutations of the imprinted IGF2-H19 domain in Silver-Russell syndrome (SRS): Results from a large cohort of patients with SRS and SRS-like phenotypes. J. Med. Genet. 2009, 46, 192–197. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Forné, T.; Oswald, J.; Dean, W.; Saam, J.R.; Bailleul, B.; Dandolo, L.; Tilghman, S.M.; Walter, J.; Reik, W. Loss of the maternal H19 gene induces changes in Igf2 methylation in both cis and trans. Proc. Natl. Acad. Sci. USA 1997, 94, 10243–10248. [Google Scholar] [CrossRef] [PubMed]
- Ripoche, M.A.; Kress, C.; Poirier, F.; Dandolo, L. Deletion of the H19 transcription unit reveals the existence of a putative imprinting control element. Genes Dev. 1997, 11, 1596–1604. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.; Ohsumi, T.K.; Kung, J.T.; Ogawa, Y.; Grau, D.J.; Sarma, K.; Song, J.J.; Kingston, R.E.; Borowsky, M.; Lee, J.T. Genome-wide identification of polycomb-associated RNAs by RIP-seq. Mol. Cell 2010, 40, 939–953. [Google Scholar] [CrossRef] [PubMed]
- Liao, B.; Hu, Y.; Herrick, D.J.; Brewer, G. The RNA-binding protein IMP-3 is a translational activator of insulin-like growth factor II leader-3 mRNA during proliferation of human K562 leukemia cells. J. Biol. Chem. 2005, 280, 18517–18524. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Chen, Z.; Fang, J.; Xu, A.; Zhang, W.; Wang, Z. H19-derived miR-675 contributes to bladder cancer cell proliferation by regulating p53 activation. Tumour Biol. 2016, 37, 263–270. [Google Scholar] [CrossRef] [PubMed]
- Tsang, W.P.; Ng, E.K.; Ng, S.S.; Jin, H.; Yu, J.; Sung, J.J.; Kwok, T.T. Oncofetal H19-derived miR-675 regulates tumor suppressor RB in human colorectal cancer. Carcinogenesis 2010, 31, 350–358. [Google Scholar] [CrossRef] [PubMed]
- Devor, E.J.; De Mik, J.N.; Schickling, B.M. Conserved sequences and secondary RNA structures confirm that H19 functions as a pri-miRNA and as an RNA-binding protein platform. Open Genom. J. 2013, 6, 1–7. [Google Scholar] [CrossRef]
- Young, T.L.; Matsuda, T.; Cepko, C.L. The noncoding RNA taurine upregulated gene 1 is required for differentiation of the murine retina. Curr. Biol. 2005, 15, 501–512. [Google Scholar] [CrossRef] [PubMed]
- Khalil, A.M.; Guttman, M.; Huarte, M.; Garber, M.; Raj, A.; Morales, D.R.; Thomas, K.; Presser, A.; Bernstein, B.E.; van Oudenaarden, A.; et al. Many human large intergenic noncoding RNAs associate with chromatin-modifying complexes and affect gene expression. Proc. Nat. Acad. Sci. USA 2009, 106, 11667–11672. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Shen, J.; Chan, M.T.V.; Wu, W.K.K. TUG1: A pivotal oncogenic long non-coding RNA of human cancers. Cell Prolif. 2016, 49, 471–475. [Google Scholar] [CrossRef] [PubMed]
- Liu, Q.; Liu, H.; Cheng, H.; Li, Y.; Li, X.; Zhu, C. Downregulation of long noncoding RNA TUG1 inhibits proliferation and induces apoptosis through the TUG1/miR-142/ZEB2 axis in bladder cancer cells. Onco. Targets Ther. 2017, 10, 2461–2471. [Google Scholar] [CrossRef] [PubMed]
- Iliev, R.; Kleinova, R.; Juracek, J.; Dolezel, J.; Ozanova, Z.; Fedorko, M.; Pacik, D.; Svoboda, M.; Stanik, M.; Slaby, O. Overexpression of long non-coding RNA TUG1 predicts poor prognosis and promotes cancer cell proliferation and migration in high-grade muscle-invasive bladder cancer. Tumor Biol. 2016, 37, 13385–13390. [Google Scholar] [CrossRef] [PubMed]
- Guttman, M.; Amit, I.; Garber, M.; French, C.; Lin, M.F.; Feldser, D.; Huarte, M.; Zuk, O.; Carey, B.W.; Cassady, J.P.; et al. Chromatin signature reveals over a thousand highly conserved large non-coding RNAs in mammals. Nature 2009, 458, 223–227. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Rice, K.; Wang, Y.; Chen, W.; Zhong, Y.; Nakayama, Y.; Zhou, Y.; Klibanski, A. Maternally expressed gene 3 (MEG3) noncoding ribonucleic acid: Isoform structure, expression, and functions. Endocrinology 2010, 151, 939–947. [Google Scholar] [CrossRef] [PubMed]
- Miyoshi, N.; Wagatsuma, H.; Wakana, S.; Shiroishi, T.; Nomura, M.; Aisaka, K.; Kohda, T.; Surani, M.A.; Kaneko-Ishino, T.; Ishino, F. Identification of an imprinted gene, MEG3/GTL2 and its human homologue MEG3, first mapped on mouse distal chromosome 12 and human chromosome 14q. Genes Cells 2000, 5, 211–220. [Google Scholar] [CrossRef] [PubMed]
- Takada, S.; Paulsen, M.; Tevendale, M.; Tsai, C.-E.; Kelsey, G.; Cattanach, B.M.; Ferguson-Smith, A.C. Epigenetic analysis of the DLK1-GTL2 imprinted domain on mouse chromosome 12: Implications for imprinting control from comparison with IGF2-H19. Hum. Mol. Genet. 2002, 11, 77–86. [Google Scholar] [CrossRef] [PubMed]
- McLaughlin, D.; Vidaki, M.; Renieri, E.; Karagogeos, D. Expression pattern of the maternally imprinted gene GTL2 in the forebrain during embryonic development and adulthood. Gene Expr. Patterns 2006, 6, 394–399. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Zhou, Y.; Mehta, K.R.; Danila, D.C.; Scolavino, S.; Johnson, S.R.; Klibanski, A. A pituitary-derived MEG3 isoform functions as a growth suppressor in tumor cells. J. Clin. Endocrinol. Metab. 2003, 88, 5119–5126. [Google Scholar] [CrossRef] [PubMed]
- Michelhaugh, S.K.; Lipovich, L.; Blythe, J.; Jia, H.; Kapatos, G.; Bannon, M.J. Mining Affymetrix microarray data for long non-coding RNAs: Altered expression in the nucleus accumbens of heroin abusers. J. Neurochem. 2011, 116, 459–466. [Google Scholar] [CrossRef] [PubMed]
- Johnson, R. Long non-coding RNAs in Huntington’s disease neurodegeneration. Neurobiol. Dis. 2012, 46, 245–254. [Google Scholar] [CrossRef] [PubMed]
- Mondal, T.; Rasmussen, M.; Pandey, G.K.; Isaksson, A.; Kanduri, C. Characterization of the RNA content of chromatin. Genome Res. 2010, 20, 899–907. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Gejman, R.; Mahta, A.; Zhong, Y.; Rice, K.A.; Zhou, Y.; Cheunsuchon, P.; Louis, D.N.; Klibanski, A. Maternally expressed gene 3, an imprinted noncoding RNA gene, is associated with meningioma pathogenesis and progression. Cancer Res. 2010, 70, 2350–2358. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Zhong, Y.; Wang, Y.; Zhang, X.; Batista, D.L.; Gejman, R.; Ansell, P.J.; Zhao, J.; Weng, C.; Klibanski, A. Activation of p53 by MEG3 non-coding RNA. J. Biol. Chem. 2007, 282, 24731–24742. [Google Scholar] [CrossRef] [PubMed]
- Gordon, F.E.; Nutt, C.L.; Cheunsuchon, P.; Nakayama, Y.; Provencher, K.A.; Rice, K.A.; Zhou, Y.; Zhang, X.; Klibanski, A. Increased expression of angiogenic genes in the brains of mouse MEG3-null embryos. Endocrinology 2010, 151, 2443–2452. [Google Scholar] [CrossRef] [PubMed]
- Ying, L.; Huang, Y.; Chen, H.; Wang, Y.; Xia, L.; Chen, Y.; Liu, Y.; Qiu, F. Downregulated MEG3 activates autophagy and increases cell proliferation in bladder cancer. Mol. Biosyst. 2013, 9, 407–411. [Google Scholar] [CrossRef] [PubMed]
- Greife, A.; Knievel, J.; Ribarska, T.; Niegisch, G.; Schulz, W.A. Concomitant downregulation of the imprinted genes DLK1 and MEG3 at 14q32.2 by epigenetic mechanisms in urothelial carcinoma. Clin. Epigenetics 2014, 6, 29. [Google Scholar] [CrossRef] [PubMed]
- Duan, W.; Du, L.; Jiang, X.; Wang, R.; Yan, S.; Xie, Y.; Yan, K.; Wang, Q.; Wang, L.; Zhang, X.; et al. Identification of a serum circulating lncRNA panel for the diagnosis and recurrence prediction of bladder cancer. Oncotarget 2016, 7, 78850–78858. [Google Scholar] [CrossRef] [PubMed]
- Shi, Y.; Lu, J.; Zhou, J.; Tan, X.; He, Y.; Ding, J.; Tian, Y.; Wang, L.; Wang, K. Long non-coding RNA Loc554202 regulates proliferation and migration in breast cancer cells. Biochem. Biophys. Res. Commun. 2014, 446, 448–453. [Google Scholar] [CrossRef] [PubMed]
- Augoff, K.; McCue, B.; Plow, E.F.; Sossey-Alaoui, K. miR-31 and its host gene lncRNA LOC554202 are regulated by promoter hypermethylation in triple-negative breast cancer. Mol. Cancer 2012, 11, 5. [Google Scholar] [CrossRef] [PubMed]
- He, A.; Chen, Z.; Mei, H.; Liu, Y. Decreased expression of LncRNA MIR31HG in human bladder cancer. Cancer Biomark. 2016, 17, 231–236. [Google Scholar] [CrossRef] [PubMed]
- He, W.; Cai, Q.; Sun, F.; Zhong, G.; Wang, P.; Liu, H.; Luo, J.; Yu, H.; Huang, J.; Lin, T. linc-UBC1 physically associates with polycomb repressive complex 2 (PRC2) and acts as a negative prognostic factor for lymph node metastasis and survival in bladder cancer. Biochim. Biophys. Acta 2013, 1832, 1528–1537. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.P.; Bian, X.J.; Ye, D.W.; Yao, X.D.; Zhang, S.L.; Dai, B.; Zhang, H.L.; Shen, Y.J. Long noncoding RNA expression signatures of bladder cancer revealed by microarray. Oncol. Lett. 2014, 7, 1197–1202. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.; Dai, B.; Zhang, H.; Shi, G.; Shen, Y.; Ye, D. Long non-coding RNA LOC572558 inhibits bladder cancer cell proliferation and tumor growth by regulating the AKT-MDM2-p53 signaling axis. Cancer Lett. 2016, 380, 369–374. [Google Scholar] [CrossRef] [PubMed]
- Hung, T.; Wang, Y.; Lin, M.F.; Koegel, A.K.; Kotake, Y.; Grant, G.D.; Horlings, H.M.; Shah, N.; Umbricht, C.; Wang, P.; et al. Extensive and coordinated transcription of noncoding RNAs within cell-cycle promoters. Nature Genet. 2011, 43, 621–629. [Google Scholar] [CrossRef] [PubMed]
- Zhan, Y.; Lin, J.; Liu, Y.; Chen, M.; Chen, X.; Zhuang, C.; Liu, L.; Xu, W.; Chen, Z.; He, A.; et al. Up-regulation of long non-coding RNA PANDAR is associated with poor prognosis and promotes tumorigenesis in bladder cancer. J. Exp. Clin. Cancer Res. 2016, 35, 83. [Google Scholar] [CrossRef] [PubMed]
- Kotake, Y.; Kitagawa, K.; Ohhata, T.; Sakai, S.; Uchida, C.; Niida, H.; Naemura, M.; Kitagawa, M. Long non-coding RNA, PANDA, contributes to the stabilization of p53 tumor suppressor protein. Anticancer Res. 2016, 36, 1605–1612. [Google Scholar] [PubMed]
- Yang, F.; Xue, X.; Zheng, L.; Bi, J.; Zhou, Y.; Zhi, K.; Gu, Y.; Fang, G. Long non-coding RNA GHET1 promotes gastric carcinoma cell proliferation by increasing c-Myc mRNA stability. FEBS J. 2014, 281, 802–813. [Google Scholar] [CrossRef] [PubMed]
- Li, L.J.; Zhu, J.L.; Bao, W.S.; Chen, D.K.; Huang, W.W.; Weng, Z.L. Long noncoding RNA GHET1 promotes the development of bladder cancer. Int. J. Clin. Exp. Pathol. 2014, 7, 7196–7205. [Google Scholar] [PubMed]
- Zhu, Y.; Yu, M.; Li, Z.; Kong, C.; Bi, J.; Li, J.; Gao, Z.; Li, Z. ncRAN, a newly identified long noncoding RNA, enhances human bladder tumor growth, invasion, and survival. Urology 2011, 77, 510.e1–510.e5. [Google Scholar] [CrossRef] [PubMed]
- Yu, M.; Ohira, M.; Li, Y.; Niizuma, H.; Oo, M.L.; Zhu, Y.; Ozaki, T.; Isogai, E.; Nakamura, Y.; Koda, T.; et al. A high expression of ncRAN, a novel non-coding RNA mapped to chromosome 17q25.1, is associated with poor prognosis in neuroblastoma. Int. J. Oncol. 2009, 34, 931–938. [Google Scholar] [PubMed]
- Smith, C.M.; Steitz, J.A. Classification of gas5 as a multi-small-nucleolar-RNA (snoRNA) host gene and a member of the 5-prime terminal oligopyrimidine gene family reveals common features of snoRNA host genes. Mol. Cell. Biol. 1998, 18, 6897–6909. [Google Scholar] [CrossRef] [PubMed]
- Raho, G.; Barone, V.; Rossi, D.; Philipson, L.; Sorrentino, V. The gas 5 gene shows four alternative splicing patterns without coding for a protein. Gene 2000, 256, 13–17. [Google Scholar] [CrossRef]
- Liu, Z.; Wang, W.; Jiang, J.; Bao, E.; Xu, D.; Zeng, Y.; Tao, L.; Qiu, J. Downregulation of GAS5 promotes bladder cancer cell proliferation, partly by regulating CDK6. PLoS ONE 2013, 8, e73991. [Google Scholar] [CrossRef] [PubMed]
- Mourtada-Maarabouni, M.; Pickard, M.R.; Hedge, V.L.; Farzaneh, F.; Williams, G.T. GAS5, a non-protein-coding RNA, controls apoptosis and is downregulated in breast cancer. Oncogene 2009, 28, 195–208. [Google Scholar] [CrossRef] [PubMed]
- Pasmant, E.; Laurendeau, I.; Heron, D.; Vidaud, M.; Vidaud, D.; Bieche, I. Characterization of a germ-line deletion, including the entire INK4/ARF locus, in a melanoma-neural system tumor family: Identification of ANRIL, an antisense noncoding RNA whose expression coclusters with ARF. Cancer Res. 2007, 67, 3963–3969. [Google Scholar] [CrossRef] [PubMed]
- Zhu, H.; Li, X.; Song, Y.; Zhang, P.; Xiao, Y.; Xing, Y. Long non-coding RNA ANRIL is up-regulated in bladder cancer and regulates bladder cancer cell proliferation and apoptosis through the intrinsic pathway. Biochem. Biophys. Res. Commun. 2015, 467, 223–228. [Google Scholar] [CrossRef] [PubMed]
- Thrash-Bingham, C.A.; Tartof, K.D. aHIF: A natural antisense transcript overexpressed in human renal cancer and during hypoxia. J. Nat. Cancer Inst. 1999, 91, 143–151. [Google Scholar] [CrossRef] [PubMed]
- Baranello, L.; Bertozzi, D.; Fogli, M.V.; Pommier, Y.; Capranico, G. DNA topoisomerase I inhibition by camptothecin induces escape of RNA polymerases II from promoter-proximal pause site, antisense transcription and histone acetylation at the human HIF-1-alpha gene locus. Nucleic Acids Res. 2010, 38, 159–171. [Google Scholar] [CrossRef] [PubMed]
- Bertozzi, D.; Iurlaro, R.; Sordet, O.; Marinello, J.; Zaffaroni, N.; Capranico, G. Characterization of novel antisense HIF-1-alpha transcripts in human cancers. Cell Cycle 2011, 10, 3189–3197. [Google Scholar] [CrossRef] [PubMed]
- Chen, M.; Zhuang, C.; Liu, Y.; Li, J.; Dai, F.; Xia, M.; Zhan, Y.; Lin, J.; Chen, Z.; He, A.; et al. Tetracycline-inducible shRNA targeting antisense long non-coding RNA HIF1A-AS2 represses the malignant phenotypes of bladder cancer. Cancer Lett. 2016, 376, 155–164. [Google Scholar] [CrossRef] [PubMed]
- Rinn, J.L.; Kertesz, M.; Wang, J.K.; Squazzo, S.L.; Xu, X.; Brugmann, S.A.; Goodnough, L.H.; Helms, J.A.; Farnham, P.J.; Segal, E.; et al. Functional demarcation of active and silent chromatin domains in human HOX loci by noncoding RNAs. Cell 2007, 129, 1311–1323. [Google Scholar] [CrossRef] [PubMed]
- Tsai, M.-C.; Manor, O.; Wan, Y.; Mosammaparast, N.; Wang, J.K.; Lan, F.; Shi, Y.; Segal, E.; Chang, H.Y. Long noncoding RNA as modular scaffold of histone modification complexes. Science 2010, 329, 689–693. [Google Scholar] [CrossRef] [PubMed]
- Gupta, R.A.; Shah, N.; Wang, K.C.; Kim, J.; Horlings, H.M.; Wong, D.J.; Tsai, M.-C.; Hung, T.; Argani, P.; Rinn, J.L.; et al. Long non-coding RNA HOTAIR reprograms chromatin state to promote cancer metastasis. Nature 2010, 464, 1071–1076. [Google Scholar] [CrossRef] [PubMed]
- Yan, T.H.; Lu, S.W.; Huang, Y.Q.; Que, G.B.; Chen, J.H.; Chen, Y.P.; Zhang, H.B.; Liang, X.L.; Jiang, J.H. Upregulation of the long noncoding RNA HOTAIR predicts recurrence in stage Ta/T1 bladder cancer. Tumour Biol. 2014, 35, 10249–10257. [Google Scholar] [CrossRef] [PubMed]
- Martínez-Fernández, M.; Feber, A.; Dueñas, M.; Segovia, C.; Rubio, C.; Fernandez, M.; Villacampa, F.; Duarte, J.; López-Calderón, F.F.; Gómez-Rodríguez, M.J.; et al. Analysis of the Polycomb-related lncRNAs HOTAIR and ANRIL in bladder cancer. Clin. Epigenetics 2015, 7, 109. [Google Scholar] [CrossRef] [PubMed]
- Sun, X.; Du, P.; Yuan, W.; Du, Z.; Yu, M.; Yu, X.; Hu, T. Long non-coding RNA HOTAIR regulates cyclin J via inhibition of microRNA-205 expression in bladder cancer. Cell Death Dis. 2015, 6, e1907. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Zhuang, C.; Liu, Y.; Chen, M.; Chen, Y.; Chen, Z.; He, A.; Lin, J.; Zhan, Y.; Liu, L.; et al. Synthetic tetracycline-controllable shRNA targeting long non-coding RNA HOXD-AS1 inhibits the progression of bladder cancer. J. Exp. Clin. Cancer Res. 2016, 35, 99. [Google Scholar] [CrossRef] [PubMed]
- Xue, Y.; Ma, G.; Zhang, Z.; Hua, Q.; Chu, H.; Tong, N.; Yuan, L.; Qin, C.; Yin, C.; Zhang, Z.; et al. A novel antisense long noncoding RNA regulates the expression of MDC1 in bladder cancer. Oncotarget 2015, 6, 484–493. [Google Scholar] [CrossRef] [PubMed]
- Prensner, J.R.; Iyer, M.K.; Balbin, O.A.; Dhanasekaran, S.M.; Cao, Q.; Brenner, J.C.; Laxman, B.; Asangani, I.A.; Grasso, C.S.; Kominsky, H.D.; et al. Transcriptome sequencing across a prostate cancer cohort identifies PCAT-1, an unannotated lincRNA implicated in disease progression. Nat. Biotechnol. 2011, 29, 742–749. [Google Scholar] [CrossRef] [PubMed]
- Ge, X.; Chen, Y.; Liao, X.; Liu, D.; Li, F.; Ruan, H.; Jia, W. Overexpression of long noncoding RNA PCAT-1 is a novel biomarker of poor prognosis in patients with colorectal cancer. Med. Oncol. 2013, 30, 588. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Liu, Y.; Zhuang, C.; Xu, W.; Fu, X.; Lv, Z.; Wu, H.; Mou, L.; Zhao, G.; Cai, Z.; et al. Inducing cell growth arrest and apoptosis by silencing long non-coding RNA PCAT-1 in human bladder cancer. Tumour Biol. 2015, 36, 7685–7689. [Google Scholar] [CrossRef] [PubMed]
- Tseng, Y.-Y.; Moriarity, B.S.; Gong, W.; Akiyama, R.; Tiwari, A.; Kawakami, H.; Ronning, P.; Reuland, B.; Guenther, K.; Beadnell, T.C.; et al. PVT1 dependence in cancer with MYC copy-number increase. Nature 2014, 512, 82–86. [Google Scholar] [CrossRef] [PubMed]
- Guan, Y.; Kuo, W.L.; Stilwell, J.L.; Takano, H.; Lapuk, A.V.; Fridlyand, J.; Mao, J.H.; Yu, M.; Miller, M.A.; Santos, J.L.; et al. Amplification of PVT1 contributes to the pathophysiology of ovarian and breast cancer. Clin. Cancer Res. 2007, 13, 5745–5755. [Google Scholar] [CrossRef] [PubMed]
- Zhuang, C.; Li, J.; Liu, Y.; Chen, M.; Yuan, J.; Fu, X.; Zhan, Y.; Liu, L.; Lin, J.; Zhou, Q.; et al. Tetracycline-inducible shRNA targeting long non-coding RNA PVT1 inhibits cell growth and induces apoptosis in bladder cancer cells. Oncotarget 2015, 6, 41194–41203. [Google Scholar] [CrossRef] [PubMed]
- Prensner, J.R.; Iyer, M.K.; Sahu, A.; Asangani, I.A.; Cao, Q.; Patel, L.; Vergara, I.A.; Davicioni, E.; Erho, N.; Ghadessi, M.; et al. The long noncoding RNA SChLAP1 promotes aggressive prostate cancer and antagonizes the SWI/SNF complex. Nat. Genet. 2013, 45, 1392–1398. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Shi, Z.; Nan, Y.; Li, M. Inhibiting malignant phenotypes of the bladder cancer cells by silencing long noncoding RNA SChLAP1. Int. Urol. Nephrol. 2016, 48, 711–716. [Google Scholar] [CrossRef] [PubMed]
- Khaitan, D.; Dinger, M.E.; Mazar, J.; Crawford, J.; Smith, M.A.; Mattick, J.S.; Perera, R.J. The melanoma-upregulated long noncoding RNA SPRY4-IT1 modulates apoptosis and invasion. Cancer Res. 2011, 71, 3852–3862. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.L.; Zhao, Z.H.; Xu, W.C.; Hou, J.Q.; Du, X.Y. Increased expression of SPRY4-IT1 predicts poor prognosis and promotes tumor growth and metastasis in bladder cancer. Int. J. Clin. Exp. Pathol. 2015, 8, 1954–1960. [Google Scholar] [PubMed]
- Beltran, M.; Puig, I.; Peña, C.; García, J.M.; Alvarez, A.B.; Peña, R.; Bonilla, F.; de Herreros, A.G. A natural antisense transcript regulates Zeb2/Sip1 gene expression during Snail1-induced epithelial-mesenchymal transition. Genes Dev. 2008, 22, 756–769. [Google Scholar] [CrossRef] [PubMed]
- Zhuang, J.; Lu, Q.; Shen, B.; Huang, X.; Shen, L.; Zheng, X.; Huang, R.; Yan, J.; Guo, H. TGFβ1 secreted by cancer-associated fibroblasts induces epithelial-mesenchymal transition of bladder cancer cells through lncRNA-ZEB2NAT. Sci. Rep. 2015, 5, 11924. [Google Scholar] [CrossRef] [PubMed]
- Katzman, S.; Kern, A.D.; Bejerano, G.; Fewell, G.; Fulton, L.; Wilson, R.K.; Salama, S.R.; Haussler, D. Human genome ultraconserved elements are ultraselected. Science 2007, 317, 915. [Google Scholar] [CrossRef] [PubMed]
- Reneker, J.; Lyons, E.; Conant, G.C.; Pires, J.C.; Freeling, M.; Shyu, C.R.; Korkin, D. Long identical multispecies elements in plant and animal genomes. Proc. Natl. Acad. Sci. USA 2012, 109, E1183–E1191. [Google Scholar] [CrossRef] [PubMed]
- Bejerano, G.; Pheasant, M.; Makunin, I.; Stephen, S.; Kent, W.J.; Mattick, J.S.; Haussler, D. Ultraconserved elements in the human genome. Science 2004, 304, 1321–1325. [Google Scholar] [CrossRef] [PubMed]
- McCole, R.B.; Fonseka, C.Y.; Koren, A.; Wu, C. Abnormal dosage of ultraconserved elements is highly disfavored in healthy cells but not cancer cells. PLoS Genet. 2014, 10, e1004646. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qian, K.; Liu, G.; Tang, Z.; Hu, Y.; Fang, Y.; Chen, Z.; Xu, X. The long non-coding RNA NEAT1 interacted with miR-101 modulates breast cancer growth by targeting EZH2. Arch. Biochem. Biophys. 2017, 615, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Ke, H.; Zhao, L.; Feng, X.; Xu, H.; Zou, L.; Yang, Q.; Su, X.; Peng, L.; Jiao, B. NEAT1 is required for survival of breast cancer cells through FUS and miR-548. Gene Regul. Syst. Biol. 2016, 10, 11–17. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Hao, Z.; Zhou, J.; Fan, S.; Luo, G.; Zhang, L.; Zhang, K.; Zhang, Y.; Liang, C. Promoting progression and clinicopathological significance of NEAT1 over-expression in bladder cancer. Oncotarget. Advance Publications 2016. [Google Scholar] [CrossRef]
- Zhao, F.; Lin, T.; He, W.; Han, J.; Zhu, D.; Hu, K.; Li, W.; Zheng, Z.; Huang, J.; Xie, W. Knockdown of a novel lincRNA AATBC suppresses proliferation and induces apoptosis in bladder cancer. Oncotarget 2015, 6, 1064–1078. [Google Scholar] [CrossRef] [PubMed]
- Chen, T.; Xie, W.; Xie, L.; Sun, Y.; Zhang, Y.; Shen, Z.; Sha, N.; Xu, H.; Wu, Z.; Hu, H.; et al. Expression of long noncoding RNA lncRNA-n336928 is correlated with tumor stage and grade and overall survival in bladder cancer. Biochem. Biophys. Res. Commun. 2015, 468, 666–670. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Zhong, G.; He, W.; Yu, H.; Huang, J.; Lin, T. lncRNA up-regulated in nonmuscle invasive bladder cancer facilitates tumor growth and acts as a negative prognostic factor of recurrence. J. Urol. 2016, 196, 1270–1278. [Google Scholar] [CrossRef] [PubMed]
- Peter, S.; Borkowska, E.; Drayton, R.M.; Rakhit, C.P.; Noon, A.; Chen, W.; Catto, J.W. Identification of differentially expressed long noncoding RNAs in bladder cancer. Clin. Cancer. Res. 2014, 20, 5311–5321. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Fu, D.; Qiu, Y.; Xing, X.; Xu, F.; Han, C.; Xu, X.; Wei, Z.; Zhang, Z.; Ge, J.; et al. Genome-wide screening and identification of long noncoding RNAs and their interaction with protein coding RNAs in bladder urothelial cell carcinoma. Cancer Lett. 2014, 349, 77–86. [Google Scholar] [CrossRef] [PubMed]
- Bartel, D.P. MicroRNAs: Genomics, biogenesis, mechanism, and function. Cell 2004, 116, 281–297. [Google Scholar] [CrossRef]
- International Human Genome Sequencing Consortium. Finishing the euchromatic sequence of the human genome. Nature 2004, 431, 931–945. [Google Scholar]
- Huttenhofer, A.; Schattner, P.; Polacek, N. Non-coding RNAs: Hope or hype? Trends Genet. 2005, 21, 289–297. [Google Scholar] [CrossRef] [PubMed]
- Iio, A.; Nakagawa, Y.; Hirata, I.; Naoe, T.; Akao, Y. Identification of non-coding RNAs embracing microRNA-143/145 cluster. Mol. Cancer 2010, 9, 136. [Google Scholar] [CrossRef] [PubMed]
- Enokida, H.; Yoshino, H.; Matsushita, R.; Nakagawa, M. The role of microRNAs in bladder cancer. Investig. Clin. Urol. 2016, 57, S60–S76. [Google Scholar] [CrossRef] [PubMed]
- Lai, E.C. Complementary miRNA pairs suggest a regulatory role for miRNA:miRNA duplexes. RNA 2004, 10, 171–175. [Google Scholar] [CrossRef] [PubMed]
- Bartel, D.P. MicroRNAs: Target recognition and regulatory functions. Cell 2009, 136, 215–233. [Google Scholar] [CrossRef] [PubMed]
- Moldovan, L.; Batte, K.E.; Trgovcich, J.; Wisler, J.; Marsh, C.B.; Piper, M. Methodological challenges in utilizing miRNA as circulating biomarkers. J. Cell. Mol. Med. 2014, 18, 371–390. [Google Scholar] [CrossRef] [PubMed]
- Christopher, A.F.; Kaur, R.P.; Kaur, G.; Kaur, A.; Gupta, V.; Bansal, P. MicroRNA therapeutics: Discovering novel targets and developing specific therapy. Perspect. Clin. Res. 2016, 7, 68–74. [Google Scholar] [CrossRef] [PubMed]
- Matullo, G.; Naccarati, A.; Pardini, B. MicroRNA expression profiling in bladder cancer: The challenge of next-generation sequencing in tissues and biofluids. Int. J. Cancer 2016, 138, 2334–2345. [Google Scholar] [CrossRef] [PubMed]
- Zhu, J.; Jiang, Z.; Gao, F.; Hu, X.; Zhou, L.; Chen, J.; Luo, H.; Sun, J.; Wu, S.; Han, Y.; et al. A systematic analysis on DNA methylation and the expression of both mRNA and microRNA in bladder cancer. PLoS ONE 2011, 6, e28223. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.H.; Wang, S.Q.; Wu, X.L.; Shen, M.; Chen, Z.G.; Chen, X.G.; Liu, Y.X.; Zhu, X.L.; Guo, F.; Duan, X.Z.; et al. Characterization of microRNAs expression profiling in one group of Chinese urothelial cell carcinoma identified by Solexa sequencing. Urol. Oncol. 2013, 31, 219–227. [Google Scholar] [CrossRef] [PubMed]
- Zabolotneva, A.A.; Zhavoronkov, A.; Garazha, A.V.; Roumiantsev, S.A.; Buzdin, A.A. Characteristic patterns of microRNA expression in human bladder cancer. Front. Genet. 2013, 3, 310. [Google Scholar] [CrossRef] [PubMed]
- Itesako, T.; Seki, N.; Yoshino, H.; Chiyomaru, T.; Yamasaki, T.; Hidaka, H.; Yonezawa, T.; Nohata, N.; Kinoshita, T.; Nakagawa, M.; et al. The microRNA expression signature of bladder cancer by deep sequencing: The functional significance of the miR-195/497 cluster. PLoS ONE 2014, 9, e84311. [Google Scholar] [CrossRef] [PubMed]
- Dong, F.; Xu, T.; Shen, Y.; Zhong, S.; Chen, S.; Ding, Q.; Shen, Z. Dysregulation of miRNA in bladder cancer: Altered expression with aberrant biogenesis procedure. Oncotarget 2017, 8, 27547–27568. [Google Scholar] [CrossRef] [PubMed]
- Mitash, N.; Tiwari, S.; Agnihotri, S.; Mandhani, A. Bladder cancer: Micro RNAs as biomolecules for prognostication and surveillance. Indian J. Urol. 2017, 33, 127–133. [Google Scholar] [CrossRef] [PubMed]
- Xie, Y.; Ma, X.; Chen, L.; Li, H.; Gu, L.; Gao, Y.; Zhang, Y.; Li, X.; Fan, Y.; Chen, J.; et al. MicroRNAs with prognostic significance in bladder cancer: A systematic review and meta-analysis. Sci. Rep. 2017, 7, 5619. [Google Scholar] [CrossRef] [PubMed]
- Ichimi, T.; Enokida, H.; Okuno, Y.; Kunimoto, R.; Chiyomaru, T.; Kawamoto, K.; Kawahara, K.; Toki, K.; Kawakami, K.; Nishiyama, K.; et al. Identification of novel microRNA targets based on microRNA signatures in bladder cancer. Int. J. Cancer 2009, 125, 345–352. [Google Scholar] [CrossRef] [PubMed]
- Miyamoto, K.; Seki, N.; Matsushita, R.; Yonemori, M.; Yoshino, H.; Nakagawa, M.; Enokida, H. Tumour-suppressive miRNA-26a-5p and miR-26b-5p inhibit cell aggressiveness by regulating PLOD2 in bladder cancer. Br. J. Cancer 2016, 115, 354–363. [Google Scholar] [CrossRef] [PubMed]
- Homami, A.; Ghazi, F. MicroRNAs as biomarkers associated with bladder cancer. Med. J. Islam. Repub. Iran 2016, 30, 475. [Google Scholar] [PubMed]
- Guancial, E.A.; Werner, L.; Bellmunt, J.; Bamias, A.; Choueiri, T.K.; Ross, R.; Schutz, F.A.; Park, R.S.; O’Brien, R.J.; Hirsch, M.S.; et al. FGFR3 expression in primary and metastatic urothelial carcinoma of the bladder. Cancer Med. 2014, 3, 835–844. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Junker, K.; van Oers, J.M.M.; Zwarthoff, E.C.; Kania, I.; Schubert, J.; Hartmann, A. Fibroblast growth factor receptor 3 mutations in bladder tumors correlate with low frequency of chromosome alterations. Neoplasia 2008, 10, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Pollard, C.; Smith, S.C.; Theodorescu, D. Molecular genesis of non-muscle-invasive urothelial carcinoma (NMIUC). Expert Rev. Mol. Med. 2010, 12, e10. [Google Scholar] [CrossRef] [PubMed]
- Cappellen, D.; de Oliveira, C.; Ricol, D.; de Medina, S.; Bourdin, J.; Sastre-Garau, X.; Chopin, D.; Thiery, J.P.; Radvanyi, F. Frequent activating mutations of FGFR3 in human bladder and cervix carcinomas. Nat. Genet. 1999, 23, 18–20. [Google Scholar] [CrossRef] [PubMed]
- Sibley, K.; Stern, P.; Knowles, M.A. Frequency of fibroblast growth factor receptor 3 mutations in sporadic tumours. Oncogene 2001, 20, 4416–4418. [Google Scholar] [CrossRef] [PubMed]
- Morais, D.R.; Reis, S.T.; Viana, N.; Piantino, C.B.; Massoco, C.; Moura, C.; Dip, N.; Silva, I.A.; Srougi, M.; Leite, K.R. The involvement of miR-100 in bladder urothelial carcinogenesis changing the expression levels of mRNA and proteins of genes related to cell proliferation, survival, apoptosis and chromosomal stability. Cancer Cell Int. 2014, 14, 119. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Li, Q.; Wang, K.; Dai, Y.; Yang, J.; Xue, S.; Han, F.; Zhang, Q.; Liu, J.; Wu, W. Decreased expression of microRNA-31 associates with aggressive tumor progression and poor prognosis in patients with bladder cancer. Clin. Transl. Oncol. 2013, 15, 849–854. [Google Scholar] [CrossRef] [PubMed]
- Catto, J.W.F.; Miah, S.; Owen, H.C.; Bryant, H.; Myers, K.; Dudziec, E.; Larré, S.; Milo, M.; Rehman, I.; Rosario, D.J.; et al. Distinct microRNA alterations characterize high- and low-grade bladder cancer. Cancer Res. 2009, 69, 8472–8481. [Google Scholar] [CrossRef] [PubMed]
- Veerla, S.; Lindgren, D.; Kvist, A.; Frigyesi, A.; Staaf, J.; Persson, H.; Liedberg, F.; Chebil, G.; Gudjonsson, S.; Borg, A.; et al. MiRNA expression in urothelial carcinomas: Important roles of miR-10a, miR-222, miR-125b, miR-7 and miR-452 for tumor stage and metastasis, and frequent homozygous losses of miR-31. Int. J. Cancer 2009, 124, 2236–2242. [Google Scholar] [CrossRef] [PubMed]
- Bourdon, J.C. p53 and its isoforms in cancer. Br. J. Cancer 2007, 97, 277–282. [Google Scholar] [CrossRef] [PubMed]
- Muller, P.A.J.; Vousden, K.H. Mutant p53 in cancer: New functions and therapeutic opportunities. Cancer Cell 2014, 25, 304–317. [Google Scholar] [CrossRef] [PubMed]
- Rivlin, N.; Brosh, R.; Oren, M.; Rotter, V. Mutations in the p53 tumor suppressor gene: Important milestones at the various steps of tumorigenesis. Genes Cancer 2011, 2, 466–474. [Google Scholar] [CrossRef] [PubMed]
- Smal, M.P.; Rolevich, A.I.; Polyakov, S.L.; Krasny, S.A.; Goncharova, R.I. FGFR3 and TP53 mutations in a prospective cohort of Belarusian bladder cancer patients. Exp. Oncol. 2014, 36, 246–251. [Google Scholar] [PubMed]
- Porrello, A.; Soddu, S.; Zbilut, J.P.; Crescenzi, M.; Giuliani, A. Discrimination of single amino acid mutations of the p53 protein by means of deterministic singularities of recurrence quantification analysis. Proteins 2004, 55, 743–755. [Google Scholar] [CrossRef] [PubMed]
- Ano Bom, A.P.; Rangel, L.P.; Costa, D.C.; de Oliveira, G.A.; Sanches, D.; Braga, C.A.; Gava, L.M.; Ramos, C.H.; Cepeda, A.O.; Stumbo, A.C.; et al. Mutant p53 aggregates into prion-like amyloid oligomers and fibrils: Implications for cancer. J. Biol. Chem. 2012, 287, 28152–28162. [Google Scholar] [CrossRef] [PubMed]
- Ghosh, S.; Salot, S.; Sengupta, S.; Navalkar, A.; Ghosh, D.; Jacob, R.; Das, S.; Kumar, R.; Jha, N.N.; Sahay, S.; et al. p53 amyloid formation leading to its loss of function: Implications in cancer pathogenesis. Cell Death Differ. 2017. [Google Scholar] [CrossRef] [PubMed]
- Vosgha, H.; Salajegheh, A.; Smith, R.A.; Lam, A.K. The important roles of miR-205 in normal physiology, cancers and as a potential therapeutic target. Curr. Cancer Drug Targets 2014, 14, 621–637. [Google Scholar] [CrossRef] [PubMed]
- Neely, L.A.; Rieger-Christ, K.M.; Neto, B.S.; Eroshkin, A.; Garver, J.; Patel, S.; Phung, N.A.; McLaughlin, S.; Libertino, J.A.; Whitney, D.; et al. A microRNA expression ratio defining the invasive phenotype in bladder tumors. Urol. Oncol. 2010, 28, 39–48. [Google Scholar] [CrossRef] [PubMed]
- Catto, J.W.F.; Alcaraz, A.; Bjartell, A.S.; de Vere White, R.; Evans, C.P.; Fussel, S.; Hamdy, F.C.; Kallioniemi, O.; Mengual, L.; Schlomm, T.; et al. MicroRNA in prostate, bladder, and kidney cancer: A systematic review. Eur. Urol. 2011, 59, 671–681. [Google Scholar] [CrossRef] [PubMed]
- Dalmay, T.; Edwards, D.R. MicroRNAs and the hallmarks of cancer. Oncogene 2006, 25, 6170–6175. [Google Scholar] [CrossRef] [PubMed]
- Guancial, E.A.; Bellmunt, J.; Yeh, S.; Rosenberg, J.E.; Berman, D.M. The evolving understanding of microRNA in bladder cancer. Urol. Oncol. 2014, 32, 41.e31–41.e40. [Google Scholar] [CrossRef] [PubMed]
- Wei, F.; Cao, C.; Xu, X.; Wang, J. Diverse functions of miR-373 in cancer. J. Transl. Med. 2015, 13, 162. [Google Scholar] [CrossRef] [PubMed]
- Boominathan, L. The tumor suppressors p53, p63, and p73 are regulators of microRNA processing complex. PLoS ONE 2010, 5, e10615. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.-J.; Park, S.-J.; Choi, E.Y.; Kim, S.; Kwak, H.J.; Yoo, B.C.; Yoo, H.; Lee, S.-H.; Kim, D.; Park, J.B.; et al. PTEN modulates miR-21 processing via RNA-regulatory protein RNH1. PLoS ONE 2011, 6, e28308. [Google Scholar] [CrossRef] [PubMed]
- Van Rhijn, B.W.; van der Kwast, T.H.; Vis, A.N.; Kirkels, W.J.; Boevé, E.R.; Jöbsis, A.C.; Zwarthoff, E.C. FGFR3 and P53 characterize alternative genetic pathways in the pathogenesis of urothelial cell carcinoma. Cancer Res. 2004, 64, 1911–1914. [Google Scholar] [CrossRef] [PubMed]
- Hanke, M.; Hoefig, K.; Merz, H.; Feller, A.C.; Kausch, I.; Jocham, D.; Warnecke, J.M.; Sczakiel, G. A robust methodology to study urine microRNA as tumor marker: MicroRNA-126 and microRNA-182 are related to urinary bladder cancer. Urol. Oncol. 2010, 28, 655–661. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.; Wu, L.; Lin, Q.; Shi, J.; Lin, X.; Shi, L. Evaluation of miR-182/miR-100 ratio for diagnosis and survival prediction in bladder cancer. Arch. Iran. Med. 2016, 19, 645–651. [Google Scholar] [PubMed]
- Christov, C.P.; Gardiner, T.J.; Szuts, D.; Krude, T. Functional requirement of noncoding Y RNAs for human chromosomal DNA replication. Mol. Cell. Biol. 2006, 26, 6993–7004. [Google Scholar] [CrossRef] [PubMed]
- Christov, C.P.; Trivier, E.; Krude, T. Noncoding human Y RNAs are overexpressed in tumours and required for cell proliferation. Br. J. Cancer 2008, 98, 981–988. [Google Scholar] [CrossRef] [PubMed]
- Kowalski, M.P.; Krude, T. Functional roles of non-coding Y RNAs. Int. J. Biochem. Cell Biol. 2015, 66, 20–29. [Google Scholar] [CrossRef] [PubMed]
- Han, Y.N.; Xia, S.Q.; Zhang, Y.Y.; Zheng, J.H.; Li, W. Circular RNAs: A novel type of biomarker and genetic tools in cancer. Oncotarget 2017, 8, 64551–64563. [Google Scholar] [CrossRef] [PubMed]
- Dong, Y.; He, D.; Peng, Z.; Peng, W.; Shi, W.; Wang, J.; Li, B.; Zhang, C.; Duan, C. Circular RNAs in cancer: An emerging key player. J. Hematol. Oncol. 2017, 10, 2. [Google Scholar] [CrossRef] [PubMed]
- Zhong, Z.; Lv, M.; Chen, J. Screening differential circular RNA expression profiles reveals the regulatory role of circTCF25-miR-103a-3p/miR-107-CDK6 pathway in bladder carcinoma. Sci. Rep. 2016, 6, 30919. [Google Scholar] [CrossRef] [PubMed]
- Zhong, Z.; Huang, M.; Lv, M.; He, Y.; Duan, C.; Zhang, L.; Chen, J. Circular RNA MYLK as a competing endogenous RNA promotes bladder cancer progression through modulating VEGFA/VEGFR2 signaling pathway. Cancer Lett. 2017, 403, 305–317. [Google Scholar] [CrossRef] [PubMed]


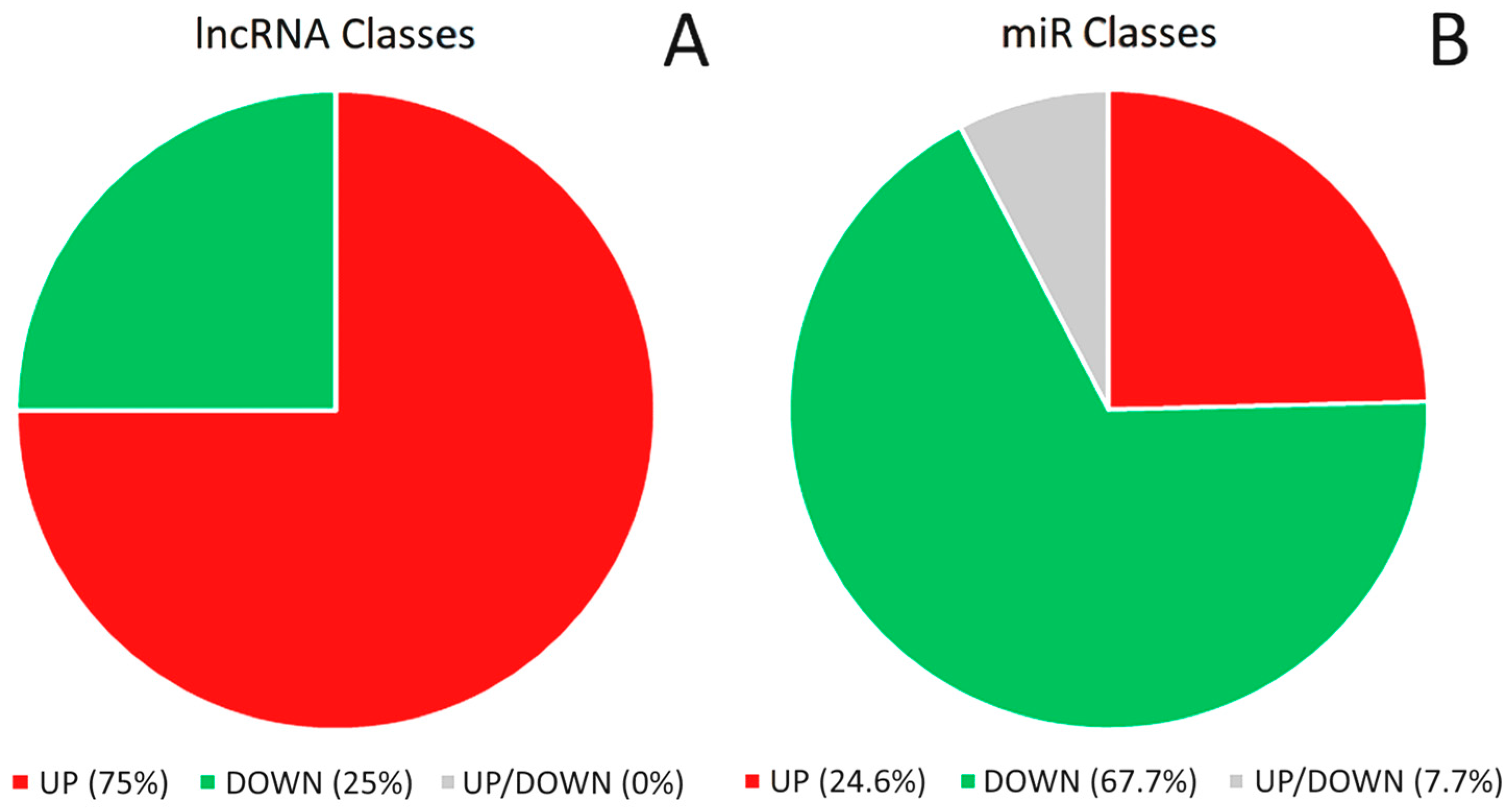{kind=link}
| Name | Acronym | Length (nt) | Notes | Refs |
|---|---|---|---|---|
| Long non-coding RNAs | lncRNA | >200 | Non-protein coding transcripts; heterogeneous class of RNAs | [41] |
| Transcribed ultraconserved region | T-UCR | ≈50–570 | Frequently located at fragile sites and cancer-associated genomic regions; possibly regulated by miR | [42] |
| Circular RNA | circRNA | ≈100–1600 | Covalently closed RNA rings; some have coding functions; potential gene regulators and miR “traps” | [43] |
| Small interfering RNA | siRNA | 20–25 | Double-stranded RNAs similar to miR, operating through RNA interference (RNAi) pathway; promote mRNA degradation | [44] |
| Y RNA | Y RNA | 21–24 | Necessary for DNA replication through interactions with chromatin and initiation proteins; target of autoimmune antibodies | [45] |
| Micro-RNA | miRNA; miR | 21–24 | Function in RNA silencing and post-transcriptional regulation of gene expression; may have an extracellular localization | [46,47] |
| Piwi-interacting RNA | piRNA | 26–31 | Epigenetic and post-transcriptional gene silencing of retrotransposons and other genetic elements in germ line cells | [48] |
| Small nucleolar RNAs | snoRNAs | 60–300 | Guide chemical modifications of other RNAs (rRNA, tRNA, snRNA) | [49] |
| Small nuclear ribonucleic acid | snRNA; U-RNA | ≈150 | Function in the processing of pre-messenger RNA (hnRNA) in the nucleus; aid in the regulation of transcription factors; telomere maintenance | [50] |
| Name | Acronym | Map | Approx. Length in Kb | Regulation in BC | Notes |
|---|---|---|---|---|---|
| Urothelial cancer-associated 1 | UCA1 | 19p13.12 | 1.4 | Up | Cell cycle regulation, cell proliferation, cell migration, cell invasion, apoptosis inhibition |
| Metastasis-associated lung adenocarcinoma transcript 1 | MALAT1 | 11q13.1 | 7 | Up | Alternative splicing, nuclear organization, modulation of gene expression |
| Imprinted maternally expressed noncoding transcript H19 | H19 | 11p15.5 | 6 | Up | Modulation of gene expression pre- and post-translation |
| Taurine Upregulated Gene 1 | TUG1 | 22q12.2 | 3.3, 5.9, 6.4, 9.7 | Up | Cell proliferation and migration |
| Maternally expressed gene 3 | MEG3 | 14q32.2 | 1.6 plus other putative sizes | Down | Maternally imprinted gene, chromatin function, angiogenesis, autophagy |
| MiR-31 host gene | MIR31HG | 9p21.3 | 2.2 | Down | Gene expression through DNA methylation, chromatin structure |
| Long intergenic noncoding RNA upregulated in bladder cancer 1; Bladder Cancer Associated Transcript 1 | Linc-UBC1, BLACAT1 | 1q32.1 | 3 | Up | Cell proliferation, cell motility, invasiveness, colony formation, chromatin structure |
| LOC572558 | LOC572558 | 9q13 | 2.6 | Down | Cell proliferation, cell motility |
| Promoter of CDKN1A antisense DNA damage-activated RNA | PANDAR | 6p21.2 | 1.5 | Up | Apoptotic response to DNA damage |
| Gastric carcinoma high expressed transcript 1 | GHET1 | 7q36.1 | 2.5 | Up | Cell proliferation, cell invasion, EMT (epithelial to mesenchymal transition) |
| Non-coding RNA expressed in aggressive neuroblastoma | ncRAN | 17q25.1 | 2.1, 2.2 | Up | Cell proliferation, cell migration, cell invasion, chemotherapy resistance |
| Growth-arrest-specific transcript 5 | GAS5 | 1q25.1 | 0.67 (exons only) | Down | Cell division through CDK6 (cyclin dependent kinase 6) control |
| Antisense noncoding RNA in the INK4 locus | ANRIL, CDKN2B-AS1 | 9p21.3 | 2.7, 3.8 | Up | Co-expressed with CDKN2A (cyclin dependent kinase inhibitor 2A); cell proliferation and apoptosis |
| Hypoxia inducible factor 1 alpha antisense RNA-2 | HIF1A-AS2, aHIF | 14q23.2 | 1.6 | Up | Cell proliferation, cell migration, apoptosis |
| HOX transcript antisense RNA | HOTAIR, HOXAS | 12q13.13 | 2.2 | Up | Regulation of cyclin J via inhibition of miR-205 |
| HOXD antisense growth-associated long non-coding RNA, HOXD cluster antisense RNA 1 | HAGLR, HOXD-AS1 | 2q31.1 | 15.8 | Up | Regulation of tumor size, histological grade and TNM (tumor-node-metastasis) stage |
| Mediator of DNA damage checkpoint protein 1 antisense RNA 1 | MDC1-AS1 | 6p21.33 | N/A | Down | DNA damage checkpoint, apoptosis |
| Prostate cancer associated transcript 1 | PCAT-1 | 8q24.21 | 2 | Up | Cell proliferation, apoptosis |
| Plasmacytoma variant translocation 1 (Pvt1) Oncogene | PVT1 | 8q24.21 | N/A | Up | Co-expression with MYC (myelocytomatosis viral oncogene homolog), cell proliferation, apoptosis |
| SWI/SNF complex antagonist associated with prostate cancer 1 | SChLAP1 | 2q31.3 | 1.7, 1.4, 1.3, 1.1 (major forms) | Up | Cell proliferation, apoptosis, cell migration |
| SPRY4 intronic transcript 1 | SPRY4-IT1 | 5q31.3 | 0.7 | Up | Cell proliferation, cell migration, cell invasion |
| ZEB2 Antisense RNA 1, Zeb2 natural antisense transcript | ZEB2-AS1, ZEB2NAT | 2q22.3 | 0.7 | Up | TGFβ1 (transforming growth factor beta 1) signaling and epithelial to mesenchymal transition (EMT) control |
| Ultraconserved RNA 8+, translated ultraconserved region 8+ | uc.8+, T-UCR 8+ | 1p36.22 | 0.2 | Up | Interaction with miR-596; cell invasion, migration, and proliferation |
| Nuclear enriched abundant transcript 1 | NEAT1 | 11q13.1 | 3.2 | Up | Cell proliferation, miR-101 interaction |
| Apoptosis associated transcript in bladder cancer | AATBC | 21q22.3 | 4.6 | Up | Cell proliferation, apoptosis |
| LncRNA-n336928 | lncRNA-n336928 | N/A | N/A | Up | N/A |
| Up-regulated in non-muscle invasive bladder cancer | lncRNA-UNMIBC | N/A | N/A | Up | Tumor relapse, chromatin structure |
| Tenascin XA pseudogene | TNXA | 6p21.33 | 4.6 | Down | N/A |
| CADM3 antisense RNA 1 | CADM3-AS1, CTA-134P22.2 | 1q23.2 | n/a | Down | N/A |
| C5orf66 antisense RNA 1 | C5orf66-AS1, CTC-276P9.1 | 5q31.1 | 1.2 | Down | N/A |
| Keratin 19 pseudogene 3 | KRT19P3 | 4q25 | 0.9 | Up | N/A |
| AB074278 | AB074278 | 17q23.2-23.3 | N/A | Up | Possible functional interactions with TANC2 (tetratricopeptide repeat, ankyrin repeat and coiled-coil containing 2) and EMP1 (epithelial membrane protein 1) |
| miR | Regulation in BC | Proposed Target Gene | Notes on Target Gene | Refs * |
|---|---|---|---|---|
| miR-1 | Down | SRSF9/SRp30c | Splicing, apoptosis | [17,173] |
| TAGLN2 | Neuronal protein | [173] | ||
| LASP1 | Oncogene | [173] | ||
| PNP | Purine nucleoside phosphorylase | [173] | ||
| PTMA | Hormone polypeptide precursor | [173] | ||
| miR-10 | Up | RASSF1 | Cell cycle inhibitor | [181] |
| MAPK1 | Cell growth, adhesion, survival, differentiation | [181] | ||
| PKC | Kinase | [181] | ||
| GRB2 | Growth factor-induced activation of RAS | [181] | ||
| FGFR3 | Cell cycle control; angiogenesis | [181] | ||
| ATM | DNA repair | [181] | ||
| MDM2/4 | Ubiquitin ligase targeting TP53 | [181] | ||
| miR-10b | Up | HOXD10 | Homeobox, transcription factor | [173] |
| KLF4 | Transcription factor | [173] | ||
| miR-100 | Down | FGFR3 | Cell cycle control; angiogenesis | [17] |
| MTOR | Protein kinase | [173] | ||
| miR-101 | Down | PLCG | Actin organization, cell migration | [181] |
| FGFR3 | Cell cycle control; angiogenesis | [181] | ||
| EZH2 | Histone methyltransferase | [185] | ||
| COX2 | Inflammation | [173] | ||
| MET | Oncogene, growth factor receptor | [173] | ||
| VEGFC | Growth factor | [173] | ||
| miR-103 | Up | MSK1 | Kinase | [181] |
| PKC | Kinase | [181] | ||
| FGFR3 | Cell cycle control; angiogenesis | [181] | ||
| miR-1182 | Down | TERT | Telomerase | [173] |
| miR-124-3p | Down | ROK1/DDX52 | Putative RNA helicase | [173] |
| CDK4 | Cyclin-dependent kinase | [173] | ||
| miR-125 | Down | RAF1 | Kinase, oncogene | [181] |
| KRAS | Activation of mitosis | [181] | ||
| FGFR3 | Cell cycle control; angiogenesis | [181] | ||
| CDKN2A | Proliferation | [181] | ||
| TP53 | Proliferation, apoptosis, angiogenesis | [181] | ||
| miR-125b | Down | E2F3 | Transcription factor | [17,173] |
| MMP13 | Matrix metalloproteinase | [173] | ||
| SPHK1 | Sphingosine kinase | [173] | ||
| miR-128 | Down | VEGFC | RAS regulator, growth factor | [173] |
| miR-129 | Down or up | GALNT1 | Post-transcriptional glycosylation | [17,173,185] |
| SOX4 | Transcription factor | [17,173,185] | ||
| SHC4 | Acetylcholine receptor clustering | [181] | ||
| PKC | Kinase | [181] | ||
| GRB2 | Growth factor-induced activation of RAS | [181] | ||
| MDM4 | Ubiquitin ligase targeting TP53 | [181] | ||
| ATM | DNA repair | [181] | ||
| miR-133a | Down | EGFR | Cell proliferation, differentiation, motility, survival | [173] |
| FSCN1 | Actin-bundling | [173] | ||
| GSTP1 | Detoxification | [173] | ||
| LASP1 | Oncogene | [173] | ||
| PNP | Purine nucleoside phosphorylase | [173] | ||
| PTMA | Hormone polypeptide precursor | [173] | ||
| TAGLN2 | Neuronal protein | [173] | ||
| KRT | Keratin | [186] | ||
| miR-133b | Down | AKT1 | Protein kinase | [173] |
| BCL2L2 | Apoptosis | [173] | ||
| EGFR | Cell proliferation, differentiation, motility, survival | [173] | ||
| KRT | Keratin | [186] | ||
| miR-135a | Down | FOXO1 | Cell cycle regulation, apoptosis | [173] |
| miR-138 | Down | ZEB2 | Transcription repression | [173] |
| miR-143 | Down or up | ERK5/MAPK7 | Kinase | [17,185] |
| AKT | Kinase | [17,173,185] | ||
| PDGFB | Growth factor | [181] | ||
| PDGRFB | Inorganic phosphate transporter | [181] | ||
| PKC | Kinase | [181] | ||
| SOS1/2 | Positive regulator of RAS | [181] | ||
| KRAS | Activation of mitosis | [181] | ||
| RAF1 | Kinase, oncogene | [181] | ||
| ATM | DNA repair | [181] | ||
| TP53 | Proliferation, apoptosis, angiogenesis | [181] | ||
| SERPIN | Serine proteinase inhibitors | [173] | ||
| miR-144-5p/3p | Down | CCNE1 | Cyclin | [173] |
| CCNE2 | Cyclin | [173] | ||
| CDC25A | Phosphatase, cell cycle | [173] | ||
| PKMYT1 | Membrane-associated cdc2-inhibitory kinase | [173] | ||
| miR-145 | Down | PKC | Kinase | [181] |
| FGFR3 | Cell cycle control; angiogenesis | [181] | ||
| CBFB | Transcription factor | [173] | ||
| CLINT1 | Early and recycling endosomes | [173] | ||
| FSCN1 | Actin-bundling | [173] | ||
| ILK | Protein kinase | [173] | ||
| PAK1 | Protein kinase | [173] | ||
| PPP3CA | Protein phosphatase | [173] | ||
| SERPIN1 | Serine proteinase inhibitors | [173] | ||
| SOCS7 | Cell signaling, cytoskeleton | [173] | ||
| IGF1R | Growth factor receptor | [173] | ||
| miR-150 | Up | PDCD4 | Tumor suppressor | [173] |
| miR-152 | Up | DNMT1 | DNA methylation | [185] |
| miR-155 | Up | CASP3 | Apoptosis | [185] |
| TP53BP1 | Apoptosis | [185] | ||
| SOCS1 | Cytokine response | [185] | ||
| PTEN | Tumor suppressor gene | [185] | ||
| PDCD4 | Tumor suppressor | [185] | ||
| SHIP1 | Cell differentiation | [185] | ||
| DMTF1 | Transcription factor | [173] | ||
| miR-16 | Down | CCND1 | Cyclin | [173] |
| miR-182-5p | Up | RECK | Tumor suppressor, cell shape | [173,184] |
| SMAD4 | Signal transduction of the transforming growth Factor-beta | [173,184] | ||
| miR-186 | Down | HMGN5 | Nucleosome, transcription activation | [173] |
| miR-1826 | Down | CTNNB1 | Wnt/beta-catenin regulator | [17] |
| MEK1 | RAS regulator | [17] | ||
| VECFG | RAS regulator, growth factor | [17] | ||
| miR-19a | Up | PTEN | Tumor suppressor gene | [173] |
| miR-193a-3p | Down | LOXL4 | Extracellular matrix formation | [173] |
| PSEN1 | NOTCH receptor cleavage | [173] | ||
| HOXC9 | Homeobox, transcription factor | [173] | ||
| miR-195 | Down | CDK-4 | Cyclin-dependent kinase | [17,173] |
| RAF1 | Kinase, oncogene | [181] | ||
| MAP2K1/2 | Kinase, cell growth | [181] | ||
| MAPK1 | Cell growth, adhesion, survival, differentiation | [181] | ||
| SOS1/2 | Positive regulator of RAS | [181] | ||
| GRB2 | Growth factor-induced activation of RAS | [181] | ||
| FGFR3 | Cell cycle control; angiogenesis | [181] | ||
| BIRC5 | Apoptosis | [173] | ||
| CDC42 | GTPase | [173] | ||
| GLUT3 | Glucose transporter | [173] | ||
| WNT7A | Cell signaling | [173] | ||
| miR-200b/c | Down or up | ERRFI-1 | Regulator of EGFR | [17,185] |
| ZEB1 | Transcriptional repressor | [17,184,185] | ||
| MMP16 | Matrix metalloproteinase | [173] | ||
| BMI1 | Oncogene | [173] | ||
| E2F3 | Transcription factor | [173] | ||
| miR-203 | Down | BCL2L2 | Apoptosis | [173,185] |
| BIRC5 | Apoptosis | [173,185] | ||
| miR-205 | Down | TP53 | Proliferation, apoptosis, angiogenesis | [17,185] |
| PTEN | Tumor suppressor gene | [17] | ||
| C-ERB-B-3 | Receptor tyrosine kinase | [17] | ||
| CDC42 | GTPase | [17] | ||
| YES | Tyrosine kinase | [17] | ||
| ZEB1/2 | Transcription repression | [184] | ||
| miR-21 | Up | TP53 | Proliferation, apoptosis, angiogenesis | [17] |
| TIMP3 | Degradation of extracellular matrix | [17] | ||
| BCL2 | Apoptosis | [17] | ||
| PTEN | Tumor suppressor gene | [17,185] | ||
| TPM1 | Tumor suppressor gene | [17] | ||
| MSH2 | DNA repair | [17] | ||
| E2F3 | Transcription factor | [17] | ||
| ATM | DNA repair | [181] | ||
| VEGFC | Growth factor | [185] | ||
| PDCD4 | Tumor suppressor | [185] | ||
| TPM1 | Tumor suppressor | [185] | ||
| miR-210 | Up | VEGF | Growth factor | [185] |
| miR-214 | Down | PDRG1 | Oncogene | [173,185] |
| miR-218 | Down | BMI1 | Oncogene | [173] |
| LASP1 | Oncogene | [173] | ||
| miR-221 | Down or up | TRAIL | Apoptosis | [17] |
| ATM | DNA repair | [181] | ||
| MDM2 | Ubiquitin ligase targeting TP53 | [181] | ||
| STMN1 | Microtubule dynamics | [173] | ||
| miR-222 | Up | PTEN | Tumor suppressor gene | [185] |
| miR-223 | Up | TP53 | Proliferation, apoptosis, angiogenesis | [181] |
| miR-224 | Up | SUFU | Tumor suppressor | [185] |
| miR-23a/b | Up | MAPK1 | Cell growth, adhesion, survival, differentiation | [181] |
| FGFR3 | Cell cycle control; angiogenesis | [181] | ||
| EGFR | Cell proliferation, differentiation, motility, survival | [173] | ||
| MET | Oncogene, growth factor receptor | [173] | ||
| ZEB1 | Transcription factor | [173,184] | ||
| miR-24 | Down | CARD10 | Apoptosis | [173] |
| FOXM1 | Transcription factor | [173] | ||
| miR-26a | Down | HMGA1 | Non-histone chromosomal protein | [185] |
| PLOD2 | Collagen crosslinking enzyme | [187] | ||
| miR-27a | Down | SLC7A11 | Cystine/glutamate exchanger | [173,185] |
| RUNX1 | Transcription factor | [173] | ||
| miR-27b | Down | DROSHA | miR biogenesis | [173] |
| EGFR | Cell proliferation, differentiation, motility, survival | [173] | ||
| MET | Oncogene, growth factor receptor | [173] | ||
| miR-27a-3p | Down | EGFR | Cell proliferation, differentiation, motility, survival | [185] |
| miR-29a/b/c | Down or up | PDGFRA | Growth factor | [181] |
| FGFR3 | Cell cycle control; angiogenesis | [181] | ||
| MCL1 | Apoptosis | [185] | ||
| DNMT3A/B | DNA methyltransferase | [185] | ||
| CDK6 | Cyclin-dependent protein kinase | [173] | ||
| miR-30a/c | Down | PLCG | Actin organization, cell migration | [181] |
| MAPK1 | Cell growth, adhesion, survival, differentiation | [181] | ||
| PDGFA | Growth factor | [181] | ||
| ATM | DNA repair | [181] | ||
| TP53 | Proliferation, apoptosis, angiogenesis | [181] | ||
| NOTCH1 | Cell signaling | [173] | ||
| miR-31 | Down | FGFR3 | Cell cycle control; angiogenesis | [17,185] |
| miR-320a/c | Down | ITGB3 | Cell adhesion | [173] |
| CDK6 | Cyclin-dependent protein kinase | [173] | ||
| miR-34a | Down | NOTCH1 | Cell signaling | [173,185] |
| CD44 | Cell adhesion | [173] | ||
| HNF4G | Nuclear receptor | [173] | ||
| miR-424 | Down | EGFR | Cell proliferation, differentiation, motility, survival | [185] |
| mir-449a | Down | CDK6 | RB control | [17] |
| CDC25a | RB control | [17,173] | ||
| TP130 | RB control | [17] | ||
| miR-485-5p | Down | HMGA2 | Non-histone chromosomal proteins | [173] |
| miR-490-5p | Down | FOS | Oncogene, transcription factor | [173] |
| miR-493 | Down | FZD4 | Transmembrane receptor | [17,173] |
| RhoC | G protein | [17,173] | ||
| miR-497 | Down | BIRC5 | Apoptosis | [173] |
| WNT7A | Cell signaling | [173] | ||
| miR-574-3p | Down | MESDC1 | Mesoderm development | [173] |
| miR-576-3p | Down | CCND1 | Cyclin | [173] |
| miR-590-3p | Down | TFAM | Mitochondrial transcription factor | [173] |
| miR-7 | Down | SHC1 | Cell signaling | [181] |
| PKC | Kinase | [181] | ||
| HOXB5 | Homeobox, transcription factor | [184] | ||
| miR-708 | Up | CASP2 | Apoptosis | [173] |
| miR-9 | Up | CDH-1 | Cell differentiation | [185] |
| CBX7 | Chromatin structure, transcription repression | [173] | ||
| CERS2 | Cell signaling | [173] | ||
| miR-96 | Down | CDKN1A | Proliferation | [173] |
| miR-99a | Down | FGFR3 | Cell cycle control; angiogenesis | [17,173] |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gulìa, C.; Baldassarra, S.; Signore, F.; Rigon, G.; Pizzuti, V.; Gaffi, M.; Briganti, V.; Porrello, A.; Piergentili, R. Role of Non-Coding RNAs in the Etiology of Bladder Cancer. Genes 2017, 8, 339. https://doi.org/10.3390/genes8110339
Gulìa C, Baldassarra S, Signore F, Rigon G, Pizzuti V, Gaffi M, Briganti V, Porrello A, Piergentili R. Role of Non-Coding RNAs in the Etiology of Bladder Cancer. Genes. 2017; 8(11):339. https://doi.org/10.3390/genes8110339
Chicago/Turabian StyleGulìa, Caterina, Stefano Baldassarra, Fabrizio Signore, Giuliano Rigon, Valerio Pizzuti, Marco Gaffi, Vito Briganti, Alessandro Porrello, and Roberto Piergentili. 2017. "Role of Non-Coding RNAs in the Etiology of Bladder Cancer" Genes 8, no. 11: 339. https://doi.org/10.3390/genes8110339