
Control of Genome Integrity by RFC Complexes; Conductors of PCNA Loading onto and Unloading from Chromatin during DNA Replication


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. PCNA, the DNA Sliding Clamp in Eukaryotic Cells
2.1. Structure and Primary Function of PCNA
2.2. Post-Translational Modifications of PCNA
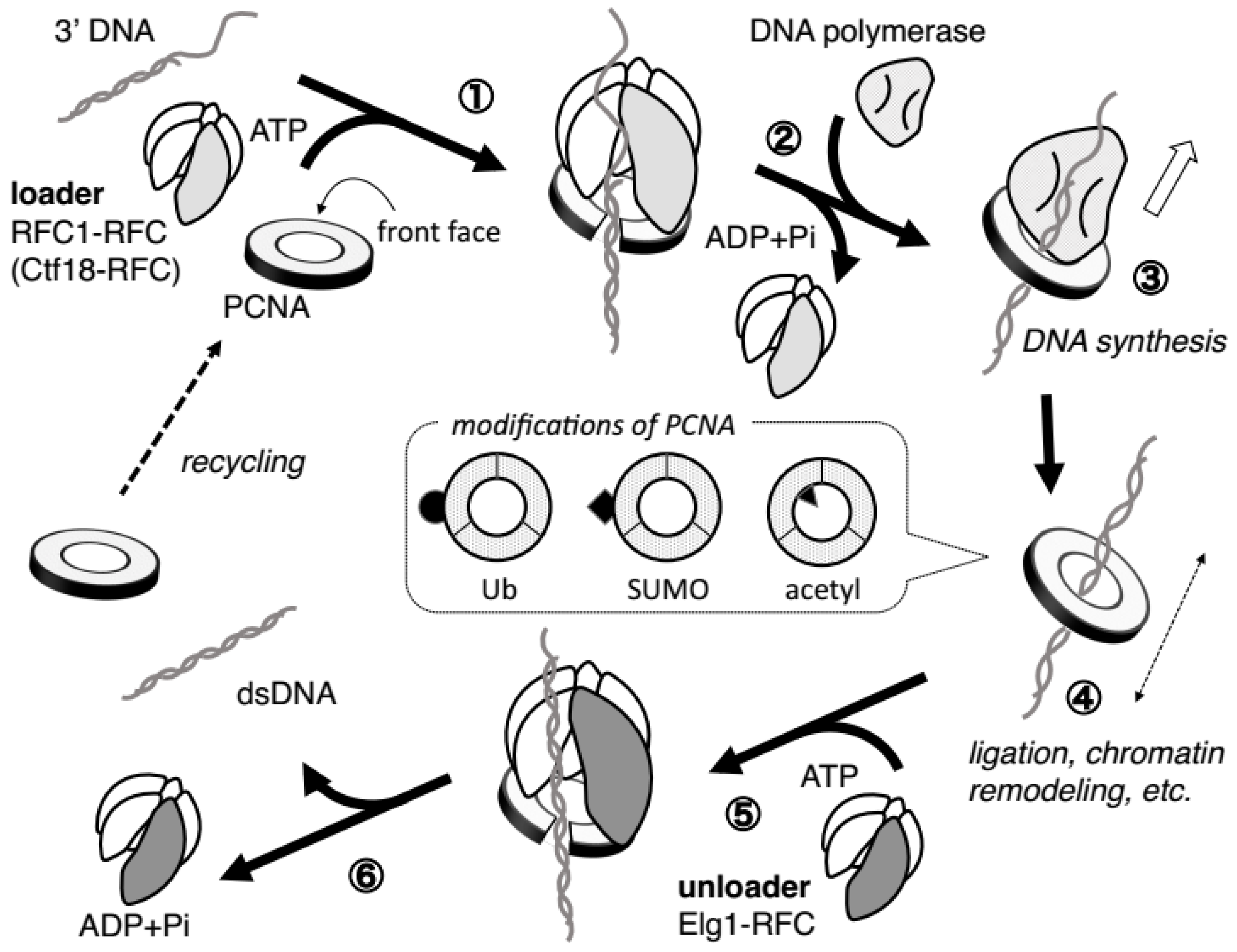
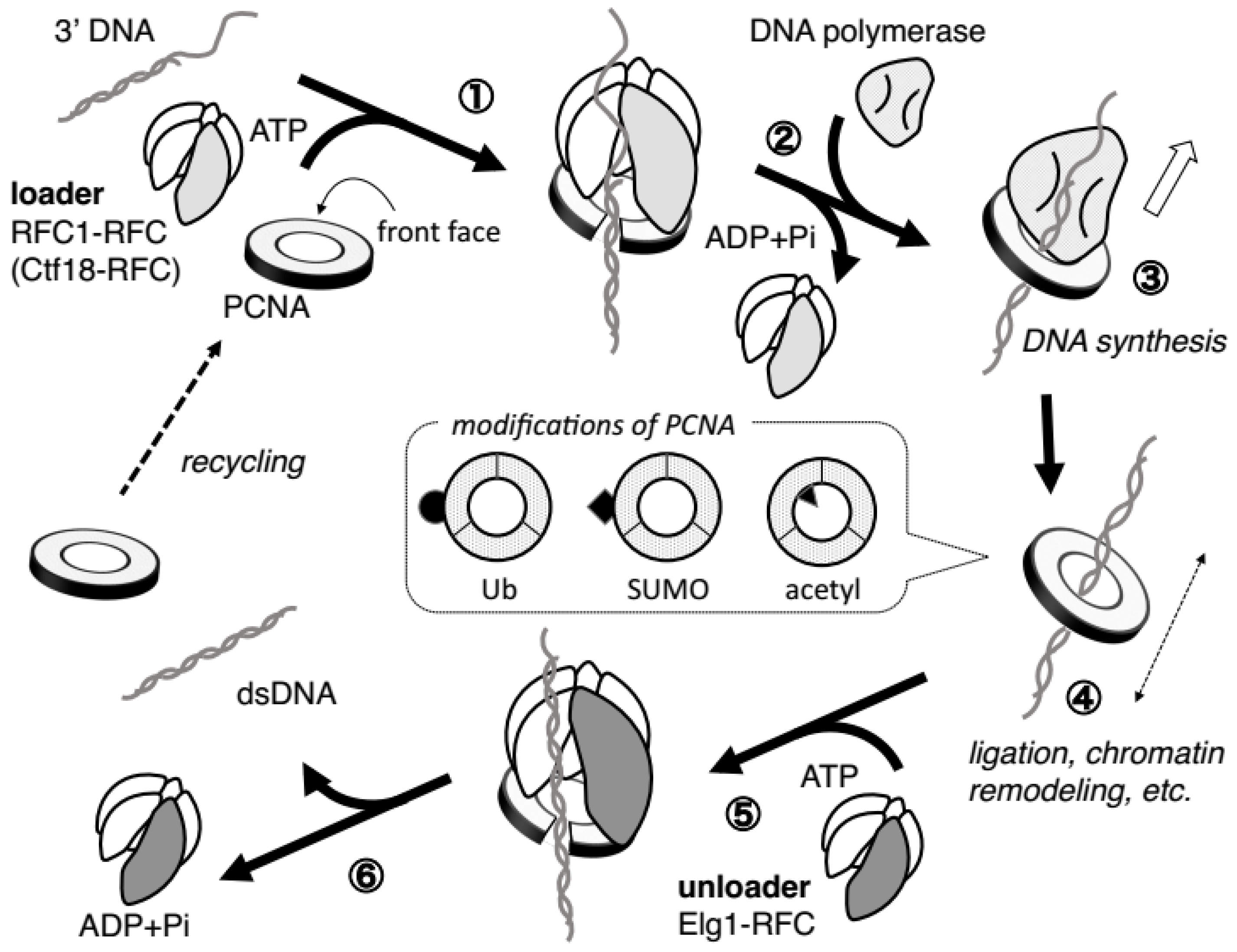
2.3. PCNA Requires Ring-Opening Factors to Regulate Its ON–OFF DNA Binding
3. Fundamental Features of RFC Complexes as PCNA Loaders/Unloaders
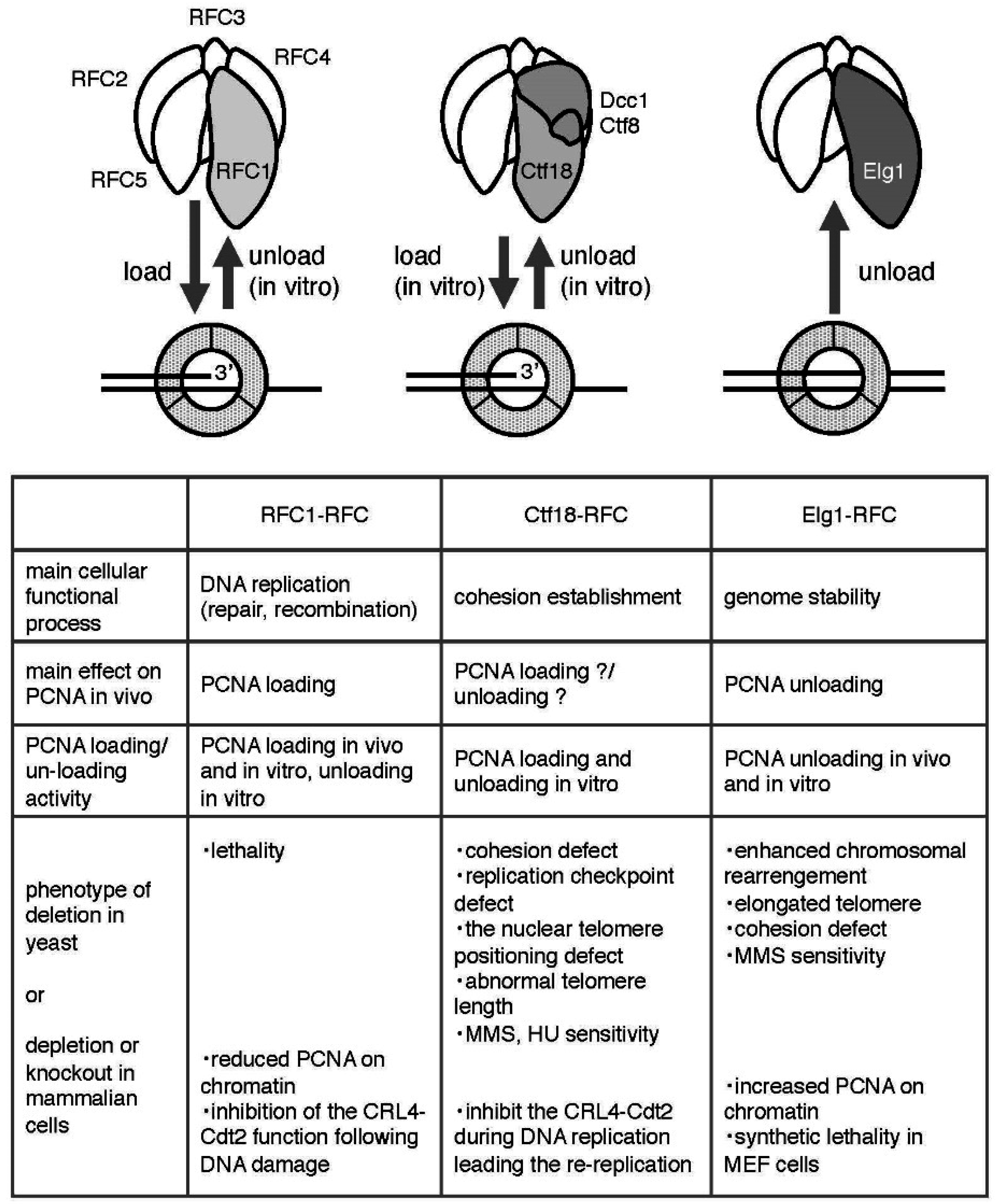
3.1. RFC1-RFC
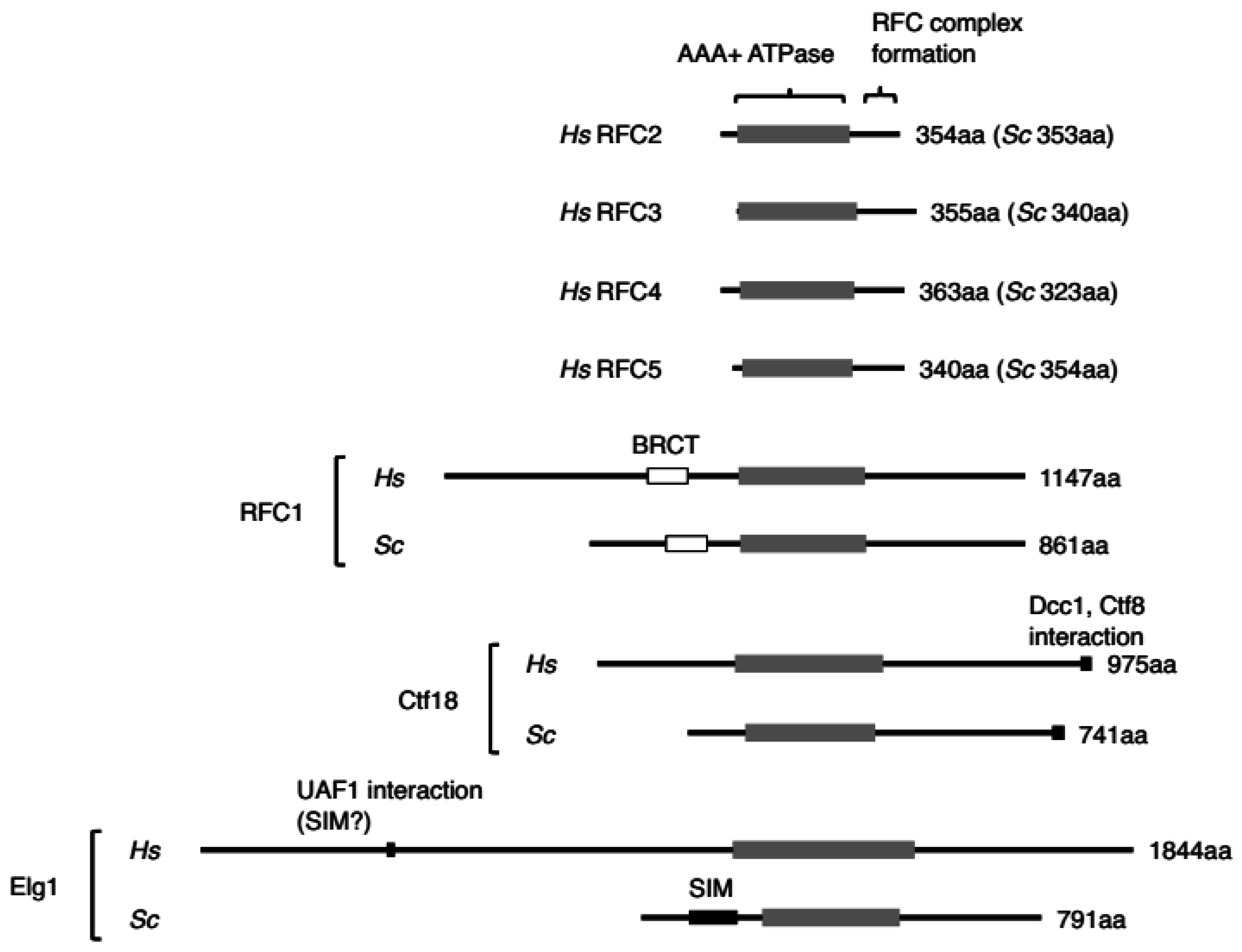
3.1.1. Fundamental Features and Structure
3.1.2. PCNA Loading/Unloading Activity of RFC1-RFC
3.2. Ctf18-RFC
3.2.1. Fundamental Features and Structure
3.2.2. PCNA Loading/Unloading Activity of Ctf18-RFC
3.3. Elg1-RFC
3.3.1. Fundamental Features and Structure
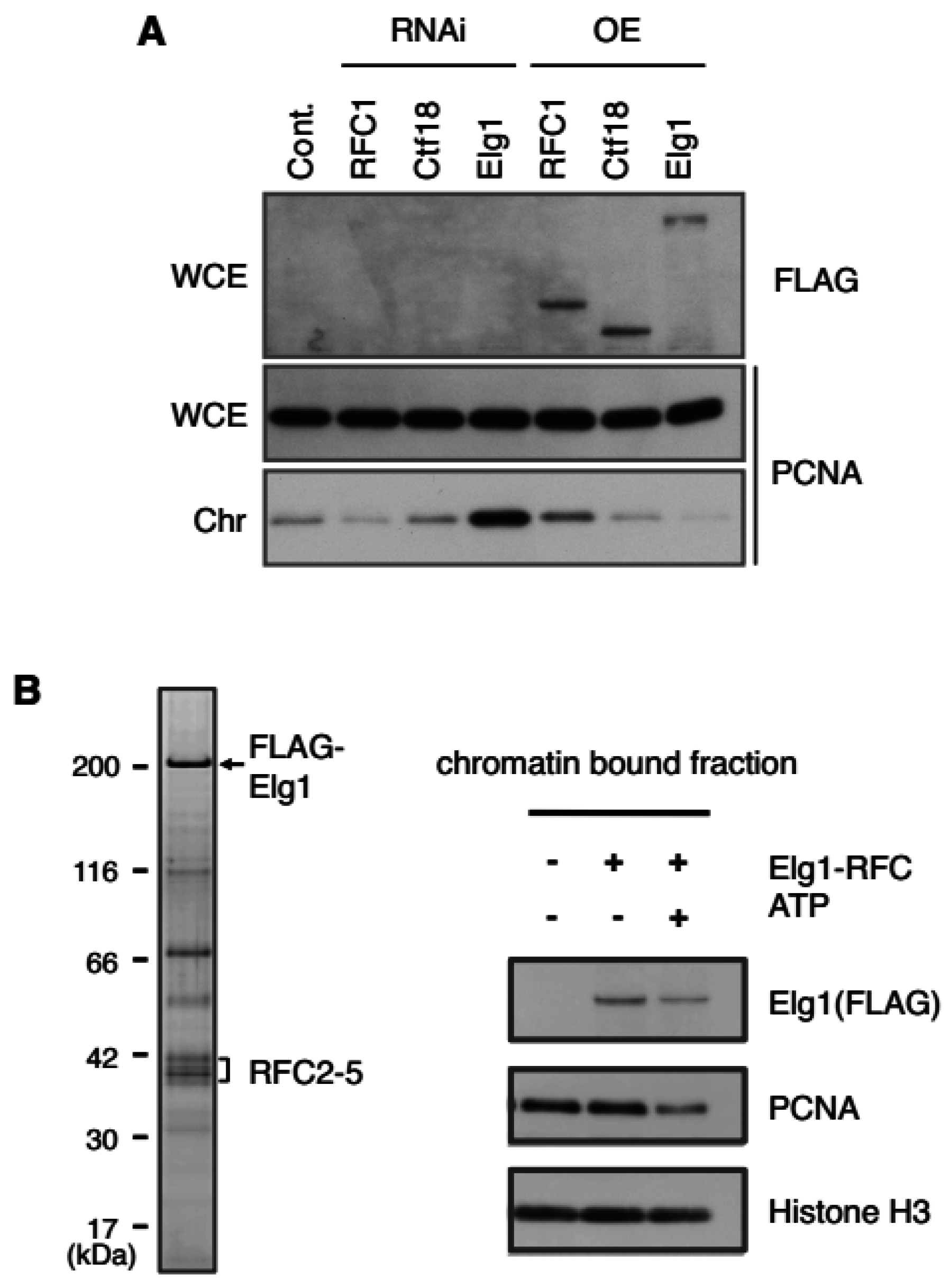
3.3.2. PCNA Unloading Activity of Elg1-RFC
4. The Three RFC Complexes Contribute to Genomic Integrity by Controlling PCNA Loading/Unloading
4.1. Roles during DNA Replication Progression
4.2. Roles in Sister Chromatid Cohesion
4.3. Roles for Proteolysis to Prevent DNA Re-Replication
4.4. Roles of RFCs in Other Events
5. RFC Complexes May Play Roles beyond PCNA Loading/Unloading
5.1. Extended Region of the Large Subunits of RFCs
5.2. RFC Complexes Interact with and Regulate Proteins Other than PCNA
6. Conclusions and Perspective
Acknowledgments
Conflicts of Interest
References
- Nurse, P. Ordering S phase and M phase in the cell cycle. Cell 1994, 79, 547–550. [Google Scholar] [CrossRef]
- Nishitani, H.; Taraviras, S.; Lygerou, Z.; Nishimoto, T. The human licensing factor for DNA replication Cdt1 accumulates in G1 and is destabilized after initiation of S-phase. J. Biol. Chem. 2001, 276, 44905–44911. [Google Scholar] [CrossRef] [PubMed]
- Masai, H.; Matsumoto, S.; You, Z.; Yoshizawa-Sugata, N.; Oda, M. Eukaryotic chromosome DNA replication: Where, when, and how? Annu. Rev. Biochem. 2010, 79, 89–130. [Google Scholar] [CrossRef] [PubMed]
- Nasmyth, K.; Haering, C.H. The structure and function of SMC and kleisin complexes. Annu. Rev. Biochem. 2005, 74, 595–648. [Google Scholar] [CrossRef] [PubMed]
- Nasmyth, K.; Haering, C.H. Cohesin: Its roles and mechanisms. Annu. Rev. Genet. 2009, 43, 525–558. [Google Scholar] [CrossRef] [PubMed]
- Johnson, A.; O’Donnell, M. Cellular DNA replicases: Components and dynamics at the replication fork. Annu. Rev. Biochem. 2005, 74, 283–315. [Google Scholar] [CrossRef] [PubMed]
- Moldovan, G.-L.; Pfander, B.; Jentsch, S. PCNA, the maestro of the replication fork. Cell 2007, 129, 665–679. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.; MacNeill, S.A. Genome stability: A new member of the RFC family. Curr. Biol. CB 2003, 13, R873–R875. [Google Scholar] [CrossRef] [PubMed]
- Waga, S.; Stillman, B. The DNA replication fork in eukaryotic cells. Annu. Rev. Biochem. 1998, 67, 721–751. [Google Scholar] [CrossRef] [PubMed]
- Bruck, I.; O’Donnell, M. The ring-type polymerase sliding clamp family. Genome Biol. 2001, 2. reviews3001.1–3001.3. [Google Scholar] [CrossRef] [PubMed]
- Bravo, R.; Frank, R.; Blundell, P.A.; Macdonald-Bravo, H. Cyclin/PCNA is the auxiliary protein of DNA polymerase-delta. Nature 1987, 326, 515–517. [Google Scholar] [CrossRef] [PubMed]
- Prelich, G.; Tan, C.K.; Kostura, M.; Mathews, M.; So, A.G.; Downey, K.M.; Stillman, B. Functional identity of proliferating cell nuclear antigen and a DNA polymerase-delta auxiliary protein. Nature 1987, 326, 517–520. [Google Scholar] [CrossRef] [PubMed]
- Krishna, T.S.; Kong, X.P.; Gary, S.; Burgers, P.M.; Kuriyan, J. Crystal structure of the eukaryotic DNA polymerase processivity factor PCNA. Cell 1994, 79, 1233–1243. [Google Scholar] [CrossRef]
- Gulbis, J.M.; Kelman, Z.; Hurwitz, J.; O’Donnell, M.; Kuriyan, J. Structure of the C-terminal region of p21(WAF1/CIP1) complexed with human PCNA. Cell 1996, 87, 297–306. [Google Scholar] [CrossRef]
- Tsurimoto, T. PCNA, a multifunctional ring on DNA. Biochim. Biophys. Acta 1998, 1443, 23–39. [Google Scholar] [CrossRef]
- Havens, C.G.; Walter, J.C. Mechanism of CRL4(Cdt2), a PCNA-dependent E3 ubiquitin ligase. Genes Dev. 2011, 25, 1568–1582. [Google Scholar] [CrossRef] [PubMed]
- Kubota, T.; Myung, K.; Donaldson, A.D. Is PCNA unloading the central function of the Elg1/ATAD5 replication factor C-like complex? Cell Cycle Georget. Tex 2013, 12, 2570–2579. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stelter, P.; Ulrich, H.D. Control of spontaneous and damage-induced mutagenesis by SUMO and ubiquitin conjugation. Nature 2003, 425, 188–191. [Google Scholar] [CrossRef] [PubMed]
- Hoege, C.; Pfander, B.; Moldovan, G.-L.; Pyrowolakis, G.; Jentsch, S. RAD6-dependent DNA repair is linked to modification of PCNA by ubiquitin and SUMO. Nature 2002, 419, 135–141. [Google Scholar] [CrossRef] [PubMed]
- Prakash, S.; Johnson, R.E.; Prakash, L. Eukaryotic translesion synthesis DNA polymerases: Specificity of structure and function. Annu. Rev. Biochem. 2005, 74, 317–353. [Google Scholar] [CrossRef] [PubMed]
- Matsuda, T.; Bebenek, K.; Masutani, C.; Hanaoka, F.; Kunkel, T. A. Low fidelity DNA synthesis by human DNA polymerase-eta. Nature 2000, 404, 1011–1013. [Google Scholar] [PubMed]
- Masutani, C.; Kusumoto, R.; Iwai, S.; Hanaoka, F. Mechanisms of accurate translesion synthesis by human DNA polymerase eta. EMBO J. 2000, 19, 3100–3109. [Google Scholar] [CrossRef] [PubMed]
- Papouli, E.; Chen, S.; Davies, A.A.; Huttner, D.; Krejci, L.; Sung, P.; Ulrich, H.D. Crosstalk between SUMO and ubiquitin on PCNA is mediated by recruitment of the helicase Srs2p. Mol. Cell 2005, 19, 123–133. [Google Scholar] [CrossRef] [PubMed]
- Pfander, B.; Moldovan, G.-L.; Sacher, M.; Hoege, C.; Jentsch, S. SUMO-modified PCNA recruits Srs2 to prevent recombination during S phase. Nature 2005, 436, 428–433. [Google Scholar] [CrossRef] [PubMed]
- Naryzhny, S.N.; Lee, H. The post-translational modifications of proliferating cell nuclear antigen: Acetylation, not phosphorylation, plays an important role in the regulation of its function. J. Biol. Chem. 2004, 279, 20194–20199. [Google Scholar] [CrossRef] [PubMed]
- Cazzalini, O.; Sommatis, S.; Tillhon, M.; Dutto, I.; Bachi, A.; Rapp, A.; Nardo, T.; Scovassi, A.I.; Necchi, D.; Cardoso, M.C.; Stivala, L.A.; Prosperi, E. CBP and p300 acetylate PCNA to link its degradation with nucleotide excision repair synthesis. Nucleic Acids Res. 2014, 42, 8433–8448. [Google Scholar] [CrossRef] [PubMed]
- Billon, P.; Li, J.; Lambert, J.-P.; Chen, Y.; Tremblay, V.; Brunzelle, J.S.; Gingras, A.-C.; Verreault, A.; Sugiyama, T.; Couture, J.-F.; et al. Acetylation of PCNA Sliding Surface by Eco1 Promotes Genome Stability through Homologous Recombination. Mol. Cell 2017, 65, 78–90. [Google Scholar] [CrossRef] [PubMed]
- Ogura, T.; Wilkinson, A.J. AAA+ superfamily ATPases: Common structure--diverse function. Genes Cells Devoted Mol. Cell. Mech. 2001, 6, 575–597. [Google Scholar] [CrossRef]
- Davey, M.J.; Jeruzalmi, D.; Kuriyan, J.; O’Donnell, M. Motors and switches: AAA+ machines within the replisome. Nat. Rev. Mol. Cell Biol. 2002, 3, 826–835. [Google Scholar] [CrossRef] [PubMed]
- Sancar, A.; Lindsey-Boltz, L.A.; Unsal-Kaçmaz, K.; Linn, S. Molecular mechanisms of mammalian DNA repair and the DNA damage checkpoints. Annu. Rev. Biochem. 2004, 73, 39–85. [Google Scholar] [CrossRef] [PubMed]
- Parrilla-Castellar, E.R.; Arlander, S.J.H.; Karnitz, L. Dial 9-1-1 for DNA damage: The Rad9-Hus1-Rad1 (9-1-1) clamp complex. DNA Repair 2004, 3, 1009–1014. [Google Scholar] [CrossRef] [PubMed]
- Mossi, R.; Hübscher, U. Clamping down on clamps and clamp loaders—The eukaryotic replication factor C. Eur. J. Biochem. 1998, 254, 209–216. [Google Scholar] [PubMed]
- Cullmann, G.; Fien, K.; Kobayashi, R.; Stillman, B. Characterization of the five replication factor C genes of Saccharomyces cerevisiae. Mol. Cell. Biol. 1995, 15, 4661–4671. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.; Robertson, K.; Mylonas, K.J.L.; Gray, F.C.; Charapitsa, I.; MacNeill, S.A. Contrasting effects of Elg1-RFC and Ctf18-RFC inactivation in the absence of fully functional RFC in fission yeast. Nucleic Acids Res. 2005, 33, 4078–4089. [Google Scholar] [CrossRef] [PubMed]
- Bunz, F.; Kobayashi, R.; Stillman, B. cDNAs encoding the large subunit of human replication factor C. Proc. Natl. Acad. Sci. USA 1993, 90, 11014–11018. [Google Scholar] [CrossRef] [PubMed]
- Uhlmann, F.; Cai, J.; Flores-Rozas, H.; Dean, F.B.; Finkelstein, J.; O’Donnell, M.; Hurwitz, J. In vitro reconstitution of human replication factor C from its five subunits. Proc. Natl. Acad. Sci. USA 1996, 93, 6521–6526. [Google Scholar] [CrossRef] [PubMed]
- Uhlmann, F.; Cai, J.; Gibbs, E.; O’Donnell, M.; Hurwitz, J. Deletion analysis of the large subunit p140 in human replication factor C reveals regions required for complex formation and replication activities. J. Biol. Chem. 1997, 272, 10058–10064. [Google Scholar] [PubMed]
- Bowman, G.D.; O’Donnell, M.; Kuriyan, J. Structural analysis of a eukaryotic sliding DNA clamp-clamp loader complex. Nature 2004, 429, 724–730. [Google Scholar] [CrossRef] [PubMed]
- Yao, N.Y.; O’Donnell, M. The RFC clamp loader: Structure and function. Subcell. Biochem. 2012, 62, 259–279. [Google Scholar] [PubMed]
- Shiomi, Y.; Usukura, J.; Masamura, Y.; Takeyasu, K.; Nakayama, Y.; Obuse, C.; Yoshikawa, H.; Tsurimoto, T. ATP-dependent structural change of the eukaryotic clamp-loader protein, replication factor C. Proc. Natl. Acad. Sci. USA 2000, 97, 14127–14132. [Google Scholar] [CrossRef] [PubMed]
- O’Donnell, M.; Jeruzalmi, D.; Kuriyan, J. Clamp loader structure predicts the architecture of DNA polymerase III holoenzyme and RFC. Curr. Biol. 2001, 11, R935–R946. [Google Scholar] [CrossRef]
- Tsurimoto, T.; Stillman, B. Functions of replication factor C and proliferating-cell nuclear antigen: functional similarity of DNA polymerase accessory proteins from human cells and bacteriophage T4. Proc. Natl. Acad. Sci. USA 1990, 87, 1023–1027. [Google Scholar] [CrossRef] [PubMed]
- Tsurimoto, T.; Stillman, B. Purification of a cellular replication factor, RF-C, that is required for coordinated synthesis of leading and lagging strands during simian virus 40 DNA replication in vitro. Mol. Cell. Biol. 1989, 9, 609–619. [Google Scholar] [CrossRef] [PubMed]
- Kelch, B.A.; Makino, D.L.; O’Donnell, M.; Kuriyan, J. Clamp loader ATPases and the evolution of DNA replication machinery. BMC Biol. 2012. [Google Scholar] [CrossRef] [PubMed]
- Hedglin, M.; Kumar, R.; Benkovic, S.J. Replication clamps and clamp loaders. Cold Spring Harb. Perspect. Biol. 2013. [Google Scholar] [CrossRef] [PubMed]
- Kelch, B.A. Review: The lord of the rings: Structure and mechanism of the sliding clamp loader. Biopolymers 2016, 105, 532–546. [Google Scholar] [CrossRef] [PubMed]
- Yao, N.; Turner, J.; Kelman, Z.; Stukenberg, P.T.; Dean, F.; Shechter, D.; Pan, Z.Q.; Hurwitz, J.; O’Donnell, M. Clamp loading, unloading and intrinsic stability of the PCNA, beta and gp45 sliding clamps of human, E. coli and T4 replicases. Genes Cells Devoted Mol. Cell. Mech. 1996, 1, 101–113. [Google Scholar] [CrossRef]
- Shibahara, K.; Stillman, B. Replication-dependent marking of DNA by PCNA facilitates CAF-1-coupled inheritance of chromatin. Cell 1999, 96, 575–585. [Google Scholar] [CrossRef]
- Yao, N.Y.; Johnson, A.; Bowman, G.D.; Kuriyan, J.; O’Donnell, M. Mechanism of proliferating cell nuclear antigen clamp opening by replication factor C. J. Biol. Chem. 2006, 281, 17528–17539. [Google Scholar] [CrossRef] [PubMed]
- Mayer, M.L.; Gygi, S.P.; Aebersold, R.; Hieter, P. Identification of RFC (Ctf18p, Ctf8p, Dcc1p): An alternative RFC complex required for sister chromatid cohesion in S. cerevisiae. Mol. Cell 2001, 7, 959–970. [Google Scholar] [CrossRef]
- Naiki, T.; Kondo, T.; Nakada, D.; Matsumoto, K.; Sugimoto, K. Chl12 (Ctf18) forms a novel replication factor C-related complex and functions redundantly with Rad24 in the DNA replication checkpoint pathway. Mol. Cell. Biol. 2001, 21, 5838–5845. [Google Scholar] [CrossRef] [PubMed]
- Bermudez, V.P.; Maniwa, Y.; Tappin, I.; Ozato, K.; Yokomori, K.; Hurwitz, J. The alternative Ctf18-Dcc1-Ctf8-replication factor C complex required for sister chromatid cohesion loads proliferating cell nuclear antigen onto DNA. Proc. Natl. Acad. Sci. USA 2003, 100, 10237–10242. [Google Scholar] [CrossRef] [PubMed]
- Merkle, C.J.; Karnitz, L.M.; Henry-Sánchez, J.T.; Chen, J. Cloning and characterization of hCTF18, hCTF8, and hDCC1. Human homologs of a Saccharomyces cerevisiae complex involved in sister chromatid cohesion establishment. J. Biol. Chem. 2003, 278, 30051–30056. [Google Scholar] [CrossRef] [PubMed]
- Shiomi, Y.; Shinozaki, A.; Sugimoto, K.; Usukura, J.; Obuse, C.; Tsurimoto, T. The reconstituted human Chl12-RFC complex functions as a second PCNA loader. Genes Cells Devoted Mol. Cell. Mech. 2004, 9, 279–290. [Google Scholar] [CrossRef] [PubMed]
- Murakami, T.; Takano, R.; Takeo, S.; Taniguchi, R.; Ogawa, K.; Ohashi, E.; Tsurimoto, T. Stable interaction between the human proliferating cell nuclear antigen loader complex Ctf18-replication factor C (RFC) and DNA polymerase {epsilon} is mediated by the cohesion-specific subunits, Ctf18, Dcc1, and Ctf8. J. Biol. Chem. 2010, 285, 34608–34615. [Google Scholar] [CrossRef] [PubMed]
- Bylund, G.O.; Burgers, P.M.J. Replication protein A-directed unloading of PCNA by the Ctf18 cohesion establishment complex. Mol. Cell. Biol. 2005, 25, 5445–5455. [Google Scholar] [CrossRef] [PubMed]
- McLellan, J.; O′Neil, N.; Tarailo, S.; Stoepel, J.; Bryan, J.; Rose, A.; Hieter, P. Synthetic lethal genetic interactions that decrease somatic cell proliferation in Caenorhabditis elegans identify the alternative RFC CTF18 as a candidate cancer drug target. Mol. Biol. Cell 2009, 20, 5306–5313. [Google Scholar] [CrossRef] [PubMed]
- Spencer, F.; Gerring, S.L.; Connelly, C.; Hieter, P. Mitotic chromosome transmission fidelity mutants in Saccharomyces cerevisiae. Genetics 1990, 124, 237–249. [Google Scholar] [PubMed]
- Kouprina, N.; Tsouladze, A.; Koryabin, M.; Hieter, P.; Spencer, F.; Larionov, V. Identification and genetic mapping of CHL genes controlling mitotic chromosome transmission in yeast. Yeast Chichester Engl. 1993, 9, 11–19. [Google Scholar] [CrossRef] [PubMed]
- Kouprina, N.; Kroll, E.; Kirillov, A.; Bannikov, V.; Zakharyev, V.; Larionov, V. CHL12, a gene essential for the fidelity of chromosome transmission in the yeast Saccharomyces cerevisiae. Genetics 1994, 138, 1067–1079. [Google Scholar] [PubMed]
- Hanna, J.S.; Kroll, E.S.; Lundblad, V.; Spencer, F.A. Saccharomyces cerevisiae CTF18 and CTF4 are required for sister chromatid cohesion. Mol. Cell. Biol. 2001, 21, 3144–3158. [Google Scholar] [CrossRef] [PubMed]
- Lengronne, A.; McIntyre, J.; Katou, Y.; Kanoh, Y.; Hopfner, K.-P.; Shirahige, K.; Uhlmann, F. Establishment of sister chromatid cohesion at the S. cerevisiae replication fork. Mol. Cell 2006, 23, 787–799. [Google Scholar] [CrossRef] [PubMed]
- Ohta, S.; Shiomi, Y.; Sugimoto, K.; Obuse, C.; Tsurimoto, T. A proteomics approach to identify proliferating cell nuclear antigen (PCNA)-binding proteins in human cell lysates. Identification of the human CHL12/RFCs2-5 complex as a novel PCNA-binding protein. J. Biol. Chem. 2002, 277, 40362–40367. [Google Scholar] [CrossRef] [PubMed]
- Ben-Aroya, S.; Koren, A.; Liefshitz, B.; Steinlauf, R.; Kupiec, M. ELG1, a yeast gene required for genome stability, forms a complex related to replication factor C. Proc. Natl. Acad. Sci. USA 2003, 100, 9906–9911. [Google Scholar] [CrossRef] [PubMed]
- Bellaoui, M.; Chang, M.; Ou, J.; Xu, H.; Boone, C.; Brown, G.W. Elg1 forms an alternative RFC complex important for DNA replication and genome integrity. EMBO J. 2003, 22, 4304–4313. [Google Scholar] [CrossRef] [PubMed]
- Kanellis, P.; Agyei, R.; Durocher, D. Elg1 forms an alternative PCNA-interacting RFC complex required to maintain genome stability. Curr. Biol. 2003, 13, 1583–1595. [Google Scholar] [CrossRef]
- Huang, M.-E.; Rio, A.-G.; Nicolas, A.; Kolodner, R.D. A genomewide screen in Saccharomyces cerevisiae for genes that suppress the accumulation of mutations. Proc. Natl. Acad. Sci. USA 2003, 100, 11529–11534. [Google Scholar] [CrossRef] [PubMed]
- Smolikov, S.; Mazor, Y.; Krauskopf, A. ELG1, a regulator of genome stability, has a role in telomere length regulation and in silencing. Proc. Natl. Acad. Sci. USA 2004, 101, 1656–1661. [Google Scholar] [CrossRef] [PubMed]
- Smith, S.; Hwang, J.-Y.; Banerjee, S.; Majeed, A.; Gupta, A.; Myung, K. Mutator genes for suppression of gross chromosomal rearrangements identified by a genome-wide screening in Saccharomyces cerevisiae. Proc. Natl. Acad. Sci. USA 2004, 101, 9039–9044. [Google Scholar] [CrossRef] [PubMed]
- Maradeo, M.E.; Skibbens, R.V. The Elg1-RFC clamp-loading complex performs a role in sister chromatid cohesion. PLoS ONE 2009, 4, e4707. [Google Scholar] [CrossRef] [PubMed]
- Parnas, O.; Zipin-Roitman, A.; Mazor, Y.; Liefshitz, B.; Ben-Aroya, S.; Kupiec, M. The ELG1 clamp loader plays a role in sister chromatid cohesion. PLoS ONE 2009, 4, e5497. [Google Scholar] [CrossRef] [PubMed]
- Shkedy, D.; Singh, N.; Shemesh, K.; Amir, A.; Geiger, T.; Liefshitz, B.; Harari, Y.; Kupiec, M. Regulation of Elg1 activity by phosphorylation. Cell Cycle Georget. Tex 2015, 14, 3689–3697. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.; Fu, H.; Aladjem, M.I.; Myung, K. ATAD5 regulates the lifespan of DNA replication factories by modulating PCNA level on the chromatin. J. Cell Biol. 2013, 200, 31–44. [Google Scholar] [CrossRef] [PubMed]
- Kubota, T.; Nishimura, K.; Kanemaki, M.T.; Donaldson, A. D. The Elg1 replication factor C-like complex functions in PCNA unloading during DNA replication. Mol. Cell 2013, 50, 273–280. [Google Scholar] [CrossRef] [PubMed]
- Shiomi, Y.; Nishitani, H. Alternative replication factor C protein, Elg1, maintains chromosome stability by regulating PCNA levels on chromatin. Genes Cells 2013, 18, 946–959. [Google Scholar] [CrossRef] [PubMed]
- Kubota, T.; Hiraga, S.; Yamada, K.; Lamond, A.I.; Donaldson, A.D. Quantitative proteomic analysis of chromatin reveals that Ctf18 acts in the DNA replication checkpoint. Mol. Cell. Proteomics 2011. [Google Scholar] [CrossRef] [PubMed]
- Nishimura, K.; Fukagawa, T.; Takisawa, H.; Kakimoto, T.; Kanemaki, M. An auxin-based degron system for the rapid depletion of proteins in nonplant cells. Nat. Methods 2009, 6, 917–922. [Google Scholar] [CrossRef] [PubMed]
- Kubota, T.; Katou, Y.; Nakato, R.; Shirahige, K.; Donaldson, A.D. Replication-Coupled PCNA Unloading by the Elg1 Complex Occurs Genome-wide and Requires Okazaki Fragment Ligation. Cell Rep. 2015, 12, 774–787. [Google Scholar] [CrossRef] [PubMed]
- Yu, C.; Gan, H.; Han, J.; Zhou, Z.-X.; Jia, S.; Chabes, A.; Farrugia, G.; Ordog, T.; Zhang, Z. Strand-specific analysis shows protein binding at replication forks and PCNA unloading from lagging strands when forks stall. Mol. Cell 2014, 56, 551–563. [Google Scholar] [CrossRef] [PubMed]
- Sikdar, N.; Banerjee, S.; Lee, K.; Wincovitch, S.; Pak, E.; Nakanishi, K.; Jasin, M.; Dutra, A.; Myung, K. DNA damage responses by human ELG1 in S phase are important to maintain genomic integrity. Cell Cycle Georget. Tex 2009, 8, 3199–3207. [Google Scholar] [CrossRef] [PubMed]
- Bell, D.W.; Sikdar, N.; Lee, K.-Y.; Price, J.C.; Chatterjee, R.; Park, H.-D.; Fox, J.; Ishiai, M.; Rudd, M.L.; Pollock, L.M.; et al. Predisposition to cancer caused by genetic and functional defects of mammalian Atad5. PLoS Genet. 2011, 7, e1002245. [Google Scholar] [CrossRef] [PubMed]
- Johnson, C.; Gali, V.K.; Takahashi, T.S.; Kubota, T. PCNA Retention on DNA into G2/M Phase Causes Genome Instability in Cells Lacking Elg1. Cell Rep. 2016, 16, 684–695. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mejlvang, J.; Feng, Y.; Alabert, C.; Neelsen, K.J.; Jasencakova, Z.; Zhao, X.; Lees, M.; Sandelin, A.; Pasero, P.; Lopes, M.; et al. New histone supply regulates replication fork speed and PCNA unloading. J. Cell Biol. 2014, 204, 29–43. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rowland, B.D.; Roig, M.B.; Nishino, T.; Kurze, A.; Uluocak, P.; Mishra, A.; Beckouët, F.; Underwood, P.; Metson, J.; Imre, R.; et al. Building sister chromatid cohesion: Smc3 acetylation counteracts an antiestablishment activity. Mol. Cell 2009, 33, 763–774. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Shi, X.; Li, Y.; Kim, B.-J.; Jia, J.; Huang, Z.; Yang, T.; Fu, X.; Jung, S.Y.; Wang, Y.; et al. Acetylation of Smc3 by Eco1 is required for S phase sister chromatid cohesion in both human and yeast. Mol. Cell 2008, 31, 143–151. [Google Scholar] [CrossRef] [PubMed]
- Unal, E.; Heidinger-Pauli, J.M.; Kim, W.; Guacci, V.; Onn, I.; Gygi, S.P.; Koshland, D.E. A molecular determinant for the establishment of sister chromatid cohesion. Science 2008, 321, 566–569. [Google Scholar] [CrossRef] [PubMed]
- Rolef Ben-Shahar, T.; Heeger, S.; Lehane, C.; East, P.; Flynn, H.; Skehel, M.; Uhlmann, F. Eco1-dependent cohesin acetylation during establishment of sister chromatid cohesion. Science 2008, 321, 563–566. [Google Scholar] [CrossRef] [PubMed]
- García-Rodríguez, L.J.; De Piccoli, G.; Marchesi, V.; Jones, R.C.; Edmondson, R.D.; Labib, K. A conserved Polϵ binding module in Ctf18-RFC is required for S-phase checkpoint activation downstream of Mec1. Nucleic Acids Res. 2015, 43, 8830–8838. [Google Scholar] [CrossRef] [PubMed]
- Okimoto, H.; Tanaka, S.; Araki, H.; Ohashi, E.; Tsurimoto, T. Conserved interaction of Ctf18-RFC with DNA polymerase ε is critical for maintenance of genome stability in Saccharomyces cerevisiae. Genes Cells Devoted Mol. Cell. Mech. 2016, 21, 482–491. [Google Scholar] [CrossRef] [PubMed]
- Moldovan, G.-L.; Pfander, B.; Jentsch, S. PCNA controls establishment of sister chromatid cohesion during S phase. Mol. Cell 2006, 23, 723–732. [Google Scholar] [CrossRef] [PubMed]
- Jin, J.; Arias, E.E.; Chen, J.; Harper, J.W.; Walter, J.C. A family of diverse Cul4-Ddb1-interacting proteins includes Cdt2, which is required for S phase destruction of the replication factor Cdt1. Mol. Cell 2006, 23, 709–721. [Google Scholar] [CrossRef] [PubMed]
- Nishitani, H.; Lygerou, Z.; Nishimoto, T.; Nurse, P. The Cdt1 protein is required to license DNA for replication in fission yeast. Nature 2000, 404, 625–628. [Google Scholar] [PubMed]
- Havens, C.G.; Walter, J.C. Docking of a specialized PIP Box onto chromatin-bound PCNA creates a degron for the ubiquitin ligase CRL4Cdt2. Mol. Cell 2009, 35, 93–104. [Google Scholar] [CrossRef] [PubMed]
- Michishita, M.; Morimoto, A.; Ishii, T.; Komori, H.; Shiomi, Y.; Higuchi, Y.; Nishitani, H. Positively charged residues located downstream of PIP box, together with TD amino acids within PIP box, are important for CRL4(Cdt2) -mediated proteolysis. Genes Cells Devoted Mol. Cell. Mech. 2011, 16, 12–22. [Google Scholar] [CrossRef] [PubMed]
- Abbas, T.; Sivaprasad, U.; Terai, K.; Amador, V.; Pagano, M.; Dutta, A. PCNA-dependent regulation of p21 ubiquitylation and degradation via the CRL4Cdt2 ubiquitin ligase complex. Genes Dev. 2008, 22, 2496–2506. [Google Scholar] [CrossRef] [PubMed]
- Abbas, T.; Shibata, E.; Park, J.; Jha, S.; Karnani, N.; Dutta, A. CRL4(Cdt2) regulates cell proliferation and histone gene expression by targeting PR-Set7/Set8 for degradation. Mol. Cell 2010, 40, 9–21. [Google Scholar] [CrossRef] [PubMed]
- Centore, R.C.; Havens, C.G.; Manning, A.L.; Li, J.-M.; Flynn, R.L.; Tse, A.; Jin, J.; Dyson, N.J.; Walter, J.C.; Zou, L. CRL4(Cdt2)-mediated destruction of the histone methyltransferase Set8 prevents premature chromatin compaction in S phase. Mol. Cell 2010, 40, 22–33. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.; Starostina, N.G.; Kipreos, E.T. The CRL4Cdt2 ubiquitin ligase targets the degradation of p21Cip1 to control replication licensing. Genes Dev. 2008, 22, 2507–2519. [Google Scholar] [CrossRef] [PubMed]
- Nishitani, H.; Shiomi, Y.; Iida, H.; Michishita, M.; Takami, T.; Tsurimoto, T. CDK inhibitor p21 is degraded by a proliferating cell nuclear antigen-coupled Cul4-DDB1Cdt2 pathway during S phase and after UV irradiation. J. Biol. Chem. 2008, 283, 29045–29052. [Google Scholar] [CrossRef] [PubMed]
- Oda, H.; Hübner, M.R.; Beck, D.B.; Vermeulen, M.; Hurwitz, J.; Spector, D.L.; Reinberg, D. Regulation of the histone H4 monomethylase PR-Set7 by CRL4(Cdt2)-mediated PCNA-dependent degradation during DNA damage. Mol. Cell 2010, 40, 364–376. [Google Scholar] [CrossRef] [PubMed]
- Tardat, M.; Brustel, J.; Kirsh, O.; Lefevbre, C.; Callanan, M.; Sardet, C.; Julien, E. The histone H4 Lys 20 methyltransferase PR-Set7 regulates replication origins in mammalian cells. Nat. Cell Biol. 2010, 12, 1086–1093. [Google Scholar] [CrossRef] [PubMed]
- Shiomi, Y.; Hayashi, A.; Ishii, T.; Shinmyozu, K.; Nakayama, J.; Sugasawa, K.; Nishitani, H. Two different replication factor C proteins, Ctf18 and RFC1, separately control PCNA-CRL4Cdt2-mediated Cdt1 proteolysis during S phase and following UV irradiation. Mol. Cell. Biol. 2012, 32, 2279–2288. [Google Scholar] [CrossRef] [PubMed]
- Aboussekhra, A.; Biggerstaff, M.; Shivji, M.K.; Vilpo, J.A.; Moncollin, V.; Podust, V.N.; Protić, M.; Hübscher, U.; Egly, J.M.; Wood, R.D. Mammalian DNA nucleotide excision repair reconstituted with purified protein components. Cell 1995, 80, 859–868. [Google Scholar] [CrossRef]
- Corrette-Bennett, S.E.; Borgeson, C.; Sommer, D.; Burgers, P.M.J.; Lahue, R.S. DNA polymerase delta, RFC and PCNA are required for repair synthesis of large looped heteroduplexes in Saccharomyces cerevisiae. Nucleic Acids Res. 2004, 32, 6268–6275. [Google Scholar] [CrossRef] [PubMed]
- Hashiguchi, K.; Matsumoto, Y.; Yasui, A. Recruitment of DNA repair synthesis machinery to sites of DNA damage/repair in living human cells. Nucleic Acids Res. 2007, 35, 2913–2923. [Google Scholar] [CrossRef] [PubMed]
- Holmes, A.M.; Haber, J.E. Double-strand break repair in yeast requires both leading and lagging strand DNA polymerases. Cell 1999, 96, 415–424. [Google Scholar] [CrossRef]
- Kadyrov, F.A.; Dzantiev, L.; Constantin, N.; Modrich, P. Endonucleolytic function of MutLalpha in human mismatch repair. Cell 2006, 126, 297–308. [Google Scholar] [CrossRef] [PubMed]
- Kolodner, R.D.; Marsischky, G.T. Eukaryotic DNA mismatch repair. Curr. Opin. Genet. Dev. 1999, 9, 89–96. [Google Scholar] [CrossRef]
- Matsumoto, Y. Molecular mechanism of PCNA-dependent base excision repair. Prog. Nucleic Acid Res. Mol. Biol. 2001, 68, 129–138. [Google Scholar] [PubMed]
- Ogi, T.; Limsirichaikul, S.; Overmeer, R.M.; Volker, M.; Takenaka, K.; Cloney, R.; Nakazawa, Y.; Niimi, A.; Miki, Y.; Jaspers, N.G.; et al. Three DNA polymerases, recruited by different mechanisms, carry out NER repair synthesis in human cells. Mol. Cell 2010, 37, 714–727. [Google Scholar] [CrossRef] [PubMed]
- Overmeer, R.M.; Gourdin, A.M.; Giglia-Mari, A.; Kool, H.; Houtsmuller, A.B.; Siegal, G.; Fousteri, M.I.; Mullenders, L.H.F.; Vermeulen, W. Replication factor C recruits DNA polymerase delta to sites of nucleotide excision repair but is not required for PCNA recruitment. Mol. Cell. Biol. 2010, 30, 4828–4839. [Google Scholar] [CrossRef] [PubMed]
- Peng, Z.; Liao, Z.; Dziegielewska, B.; Matsumoto, Y.; Thomas, S.; Wan, Y.; Yang, A.; Tomkinson, A.E. Phosphorylation of serine 51 regulates the interaction of human DNA ligase I with replication factor C and its participation in DNA replication and repair. J. Biol. Chem. 2012, 287, 36711–36719. [Google Scholar] [CrossRef] [PubMed]
- Pluciennik, A.; Dzantiev, L.; Iyer, R.R.; Constantin, N.; Kadyrov, F.A.; Modrich, P. PCNA function in the activation and strand direction of MutLα endonuclease in mismatch repair. Proc. Natl. Acad. Sci. USA 2010, 107, 16066–16071. [Google Scholar] [CrossRef] [PubMed]
- Shivji, M.K.; Podust, V.N.; Hübscher, U.; Wood, R.D. Nucleotide excision repair DNA synthesis by DNA polymerase epsilon in the presence of PCNA, RFC, and RPA. Biochemistry (Mosc.) 1995, 34, 5011–5017. [Google Scholar] [CrossRef]
- Ogiwara, H.; Ui, A.; Enomoto, T.; Seki, M. Role of Elg1 protein in double strand break repair. Nucleic Acids Res. 2007, 35, 353–362. [Google Scholar] [CrossRef] [PubMed]
- Ma, L.; Zhai, Y.; Feng, D.; Chan, T.; Lu, Y.; Fu, X.; Wang, J.; Chen, Y.; Li, J.; Xu, K.; et al. Identification of novel factors involved in or regulating initiation of DNA replication by a genome-wide phenotypic screen in Saccharomyces cerevisiae. Cell Cycle Georget. Tex 2010, 9, 4399–4410. [Google Scholar] [CrossRef] [PubMed]
- Crabbé, L.; Thomas, A.; Pantesco, V.; De Vos, J.; Pasero, P.; Lengronne, A. Analysis of replication profiles reveals key role of RFC-Ctf18 in yeast replication stress response. Nat. Struct. Mol. Biol. 2010, 17, 1391–1397. [Google Scholar] [CrossRef] [PubMed]
- Gellon, L.; Razidlo, D.F.; Gleeson, O.; Verra, L.; Schulz, D.; Lahue, R.S.; Freudenreich, C.H. New functions of Ctf18-RFC in preserving genome stability outside its role in sister chromatid cohesion. PLoS Genet. 2011, 7, e1001298. [Google Scholar] [CrossRef] [PubMed]
- Hiraga, S.; Robertson, E.D.; Donaldson, A.D. The Ctf18 RFC-like complex positions yeast telomeres but does not specify their replication time. EMBO J. 2006, 25, 1505–1514. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Terret, M.-E.; Sherwood, R.; Rahman, S.; Qin, J.; Jallepalli, P.V. Cohesin acetylation speeds the replication fork. Nature 2009, 462, 231–234. [Google Scholar] [CrossRef] [PubMed]
- Gomes, X.V.; Gary, S.L.; Burgers, P.M. Overproduction in Escherichia coli and characterization of yeast replication factor C lacking the ligase homology domain. J. Biol. Chem. 2000, 275, 14541–14549. [Google Scholar] [CrossRef] [PubMed]
- Kobayashi, M.; Ab, E.; Bonvin, A.M. J.J.; Siegal, G. Structure of the DNA-bound BRCA1 C-terminal region from human replication factor C p140 and model of the protein-DNA complex. J. Biol. Chem. 2010, 285, 10087–10097. [Google Scholar] [CrossRef] [PubMed]
- Huang, F.; Saraf, A.; Florens, L.; Kusch, T.; Swanson, S.K.; Szerszen, L.T.; Li, G.; Dutta, A.; Washburn, M.P.; Abmayr, S.M.; et al. The Enok acetyltransferase complex interacts with Elg1 and negatively regulates PCNA unloading to promote the G1/S transition. Genes Dev. 2016, 30, 1198–1210. [Google Scholar] [CrossRef] [PubMed]
- Davidson, M.B.; Brown, G.W. The N- and C-termini of Elg1 contribute to the maintenance of genome stability. DNA Repair 2008, 7, 1221–1232. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.-Y.; Yang, K.; Cohn, M.A.; Sikdar, N.; D′Andrea, A.D.; Myung, K. Human ELG1 regulates the level of ubiquitinated proliferating cell nuclear antigen (PCNA) through Its interactions with PCNA and USP1. J. Biol. Chem. 2010, 285, 10362–10369. [Google Scholar] [CrossRef] [PubMed]
- Levin, D.S.; Vijayakumar, S.; Liu, X.; Bermudez, V.P.; Hurwitz, J.; Tomkinson, A.E. A conserved interaction between the replicative clamp loader and DNA ligase in eukaryotes: Implications for Okazaki fragment joining. J. Biol. Chem. 2004, 279, 55196–55201. [Google Scholar] [CrossRef] [PubMed]
- Franco, A.A.; Lam, W.M.; Burgers, P.M.; Kaufman, P.D. Histone deposition protein Asf1 maintains DNA replisome integrity and interacts with replication factor C. Genes Dev. 2005, 19, 1365–1375. [Google Scholar] [CrossRef] [PubMed]
- Anderson, L.A.; Perkins, N.D. The large subunit of replication factor C interacts with the histone deacetylase, HDAC1. J. Biol. Chem. 2002, 277, 29550–29554. [Google Scholar] [CrossRef] [PubMed]
- Pennaneach, V.; Salles-Passador, I.; Munshi, A.; Brickner, H.; Regazzoni, K.; Dick, F.; Dyson, N.; Chen, T.T.; Wang, J.Y.; Fotedar, R.; et al. The large subunit of replication factor C promotes cell survival after DNA damage in an LxCxE motif- and Rb-dependent manner. Mol. Cell 2001, 7, 715–727. [Google Scholar] [CrossRef]
- Hong, S.; Park, S.J.; Kong, H.J.; Shuman, J.D.; Cheong, J. Functional interaction of bZIP proteins and the large subunit of replication factor C in liver and adipose cells. J. Biol. Chem. 2001, 276, 28098–28105. [Google Scholar] [CrossRef] [PubMed]
- Anderson, L.A.; Perkins, N.D. Regulation of RelA (p65) function by the large subunit of replication factor C. Mol. Cell. Biol. 2003, 23, 721–732. [Google Scholar] [CrossRef] [PubMed]
- Shiomi, Y.; Masutani, C.; Hanaoka, F.; Kimura, H.; Tsurimoto, T. A second proliferating cell nuclear antigen loader complex, Ctf18-replication factor C, stimulates DNA polymerase eta activity. J. Biol. Chem. 2007, 282, 20906–20914. [Google Scholar] [CrossRef] [PubMed]

© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shiomi, Y.; Nishitani, H. Control of Genome Integrity by RFC Complexes; Conductors of PCNA Loading onto and Unloading from Chromatin during DNA Replication. Genes 2017, 8, 52. https://doi.org/10.3390/genes8020052
Shiomi Y, Nishitani H. Control of Genome Integrity by RFC Complexes; Conductors of PCNA Loading onto and Unloading from Chromatin during DNA Replication. Genes. 2017; 8(2):52. https://doi.org/10.3390/genes8020052
Chicago/Turabian StyleShiomi, Yasushi, and Hideo Nishitani. 2017. "Control of Genome Integrity by RFC Complexes; Conductors of PCNA Loading onto and Unloading from Chromatin during DNA Replication" Genes 8, no. 2: 52. https://doi.org/10.3390/genes8020052