Characterization of TTN Novex Splicing Variants across Species and the Role of RBM20 in Novex-Specific Exon Splicing

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Animals and Tissue Samples
2.2. Genomic Sequences in Multiple Species and Computational Analysis
2.3. RT-PCR and DNA Gel Electrophoresis
2.4. DNA Sequencing
2.5. Statistics
3. Results
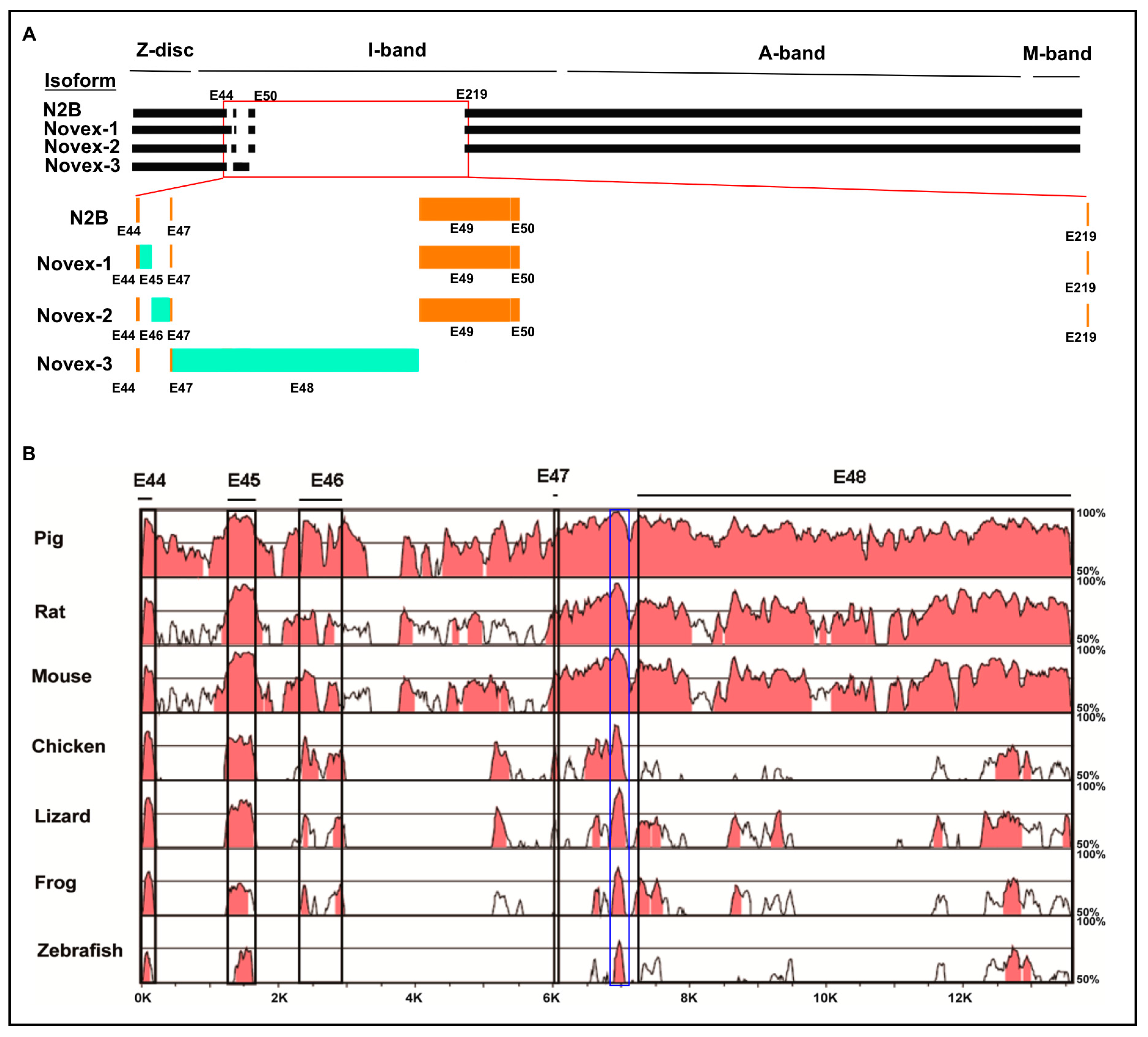
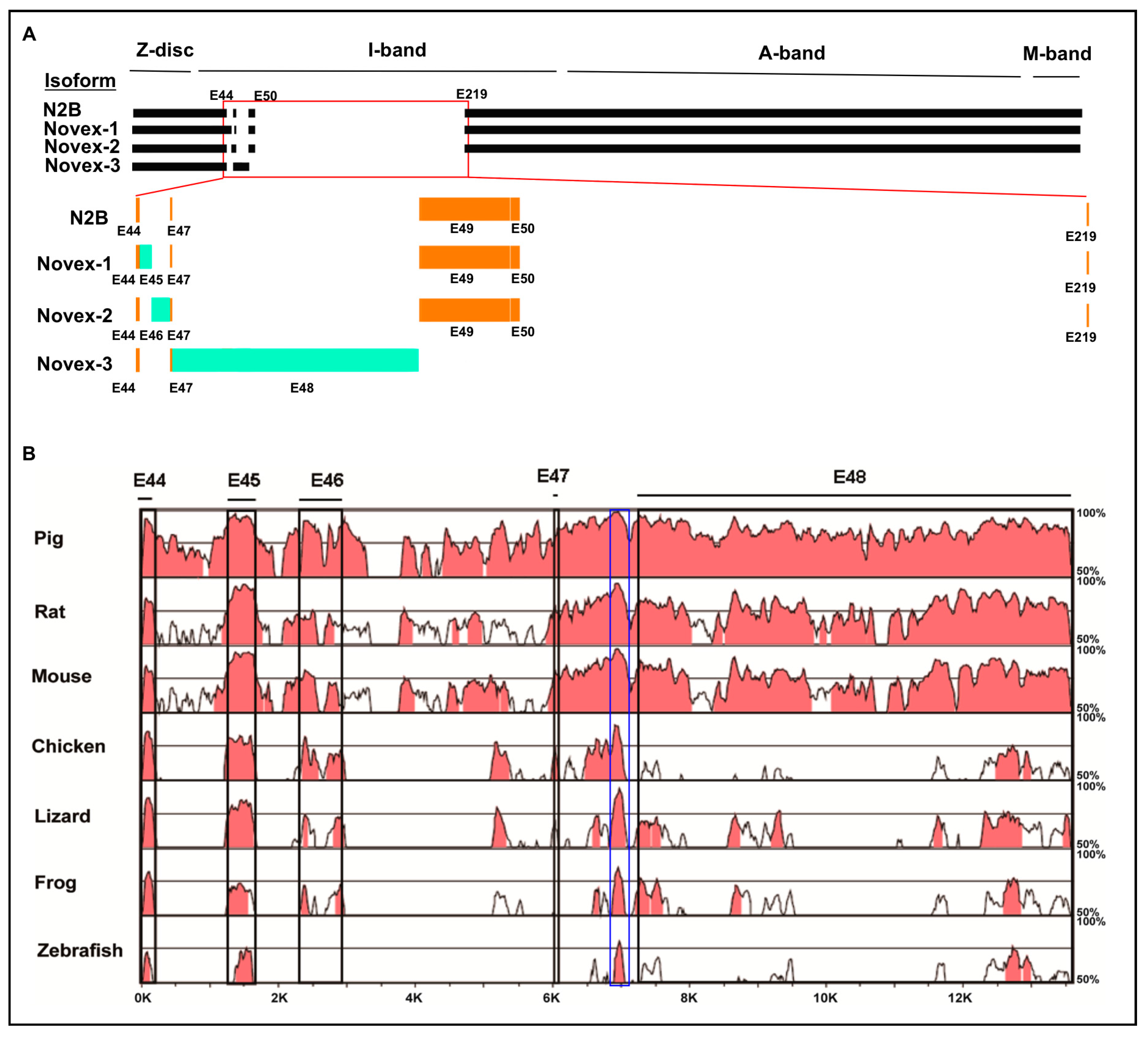
3.1. Cross-Species Comparison of TTN Isoform Novex 1, 2, and 3 Splicing Variants
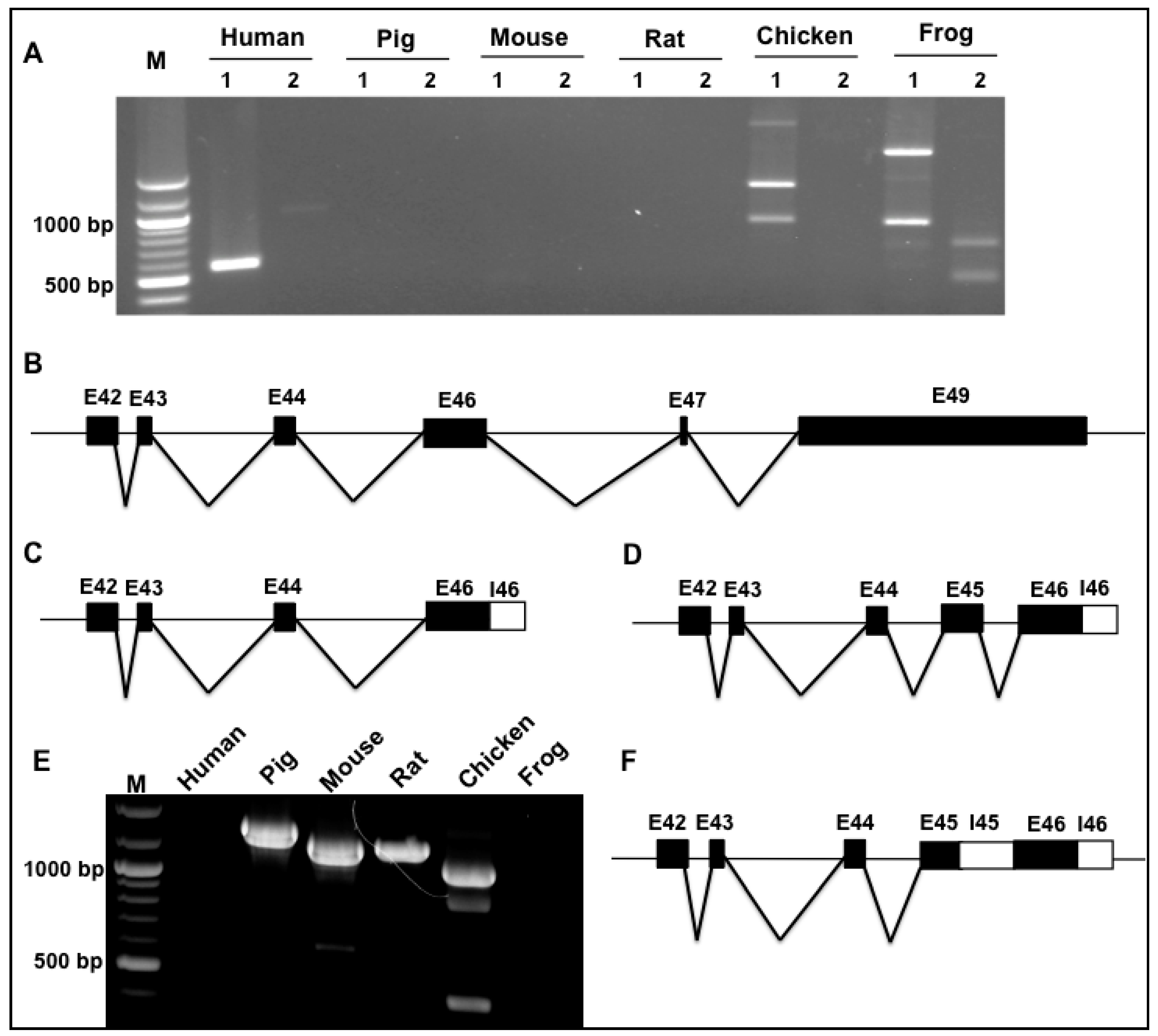
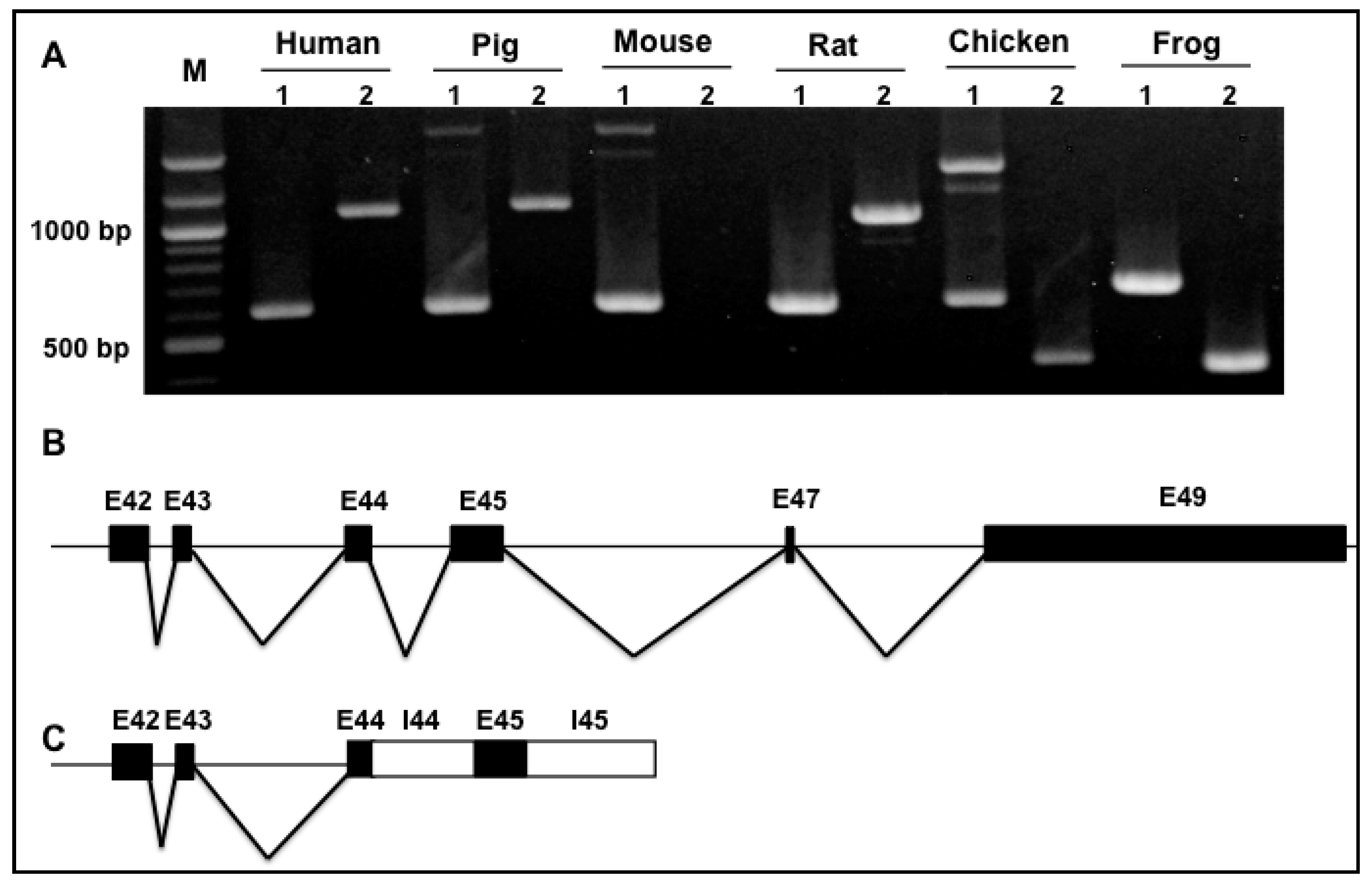
3.2. Splicing Variants of the Novex-1-Specific Exon 45 across Species
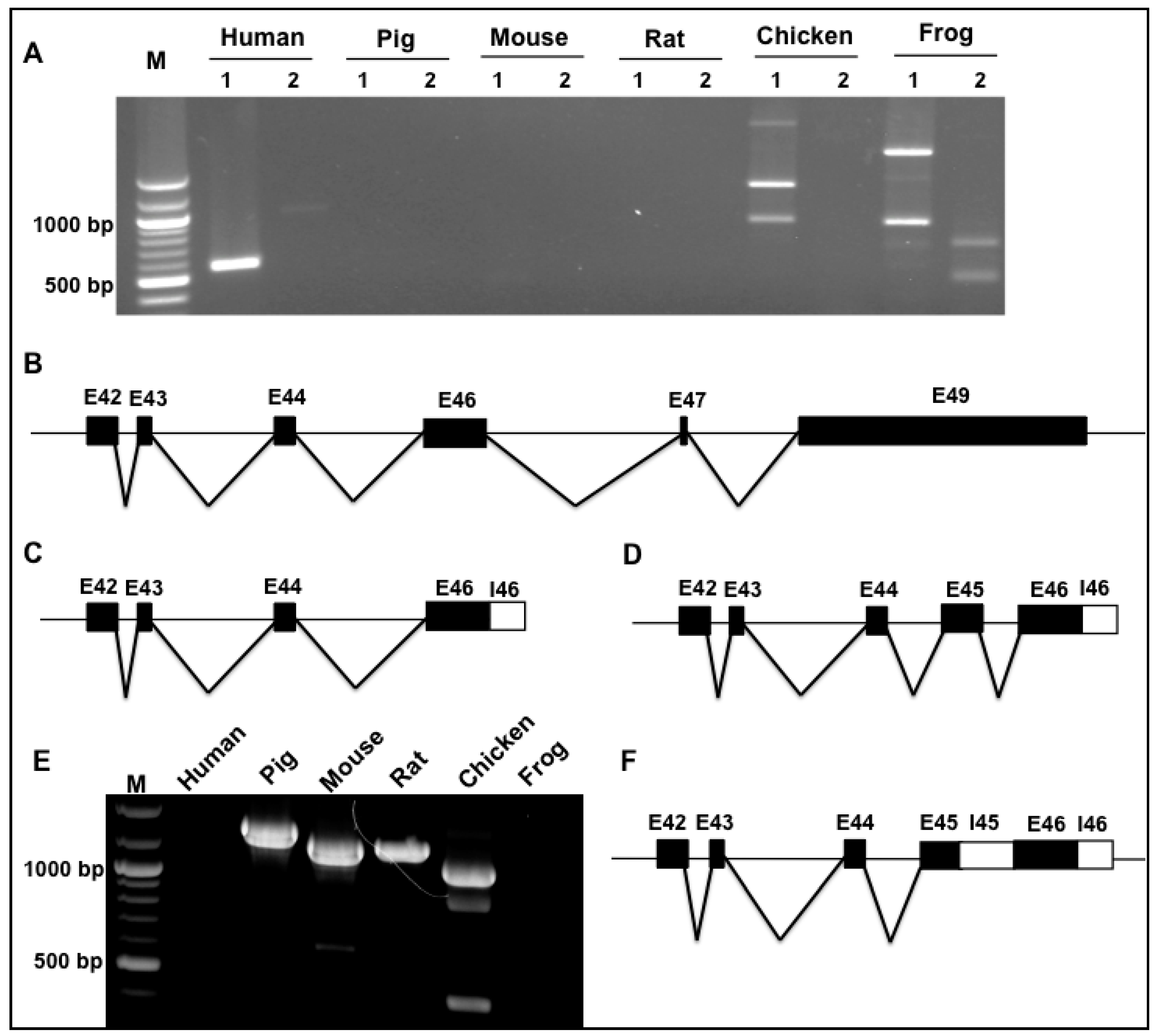
3.3. Splicing Variants of the Novex-2-Specific Exon 46 across Species
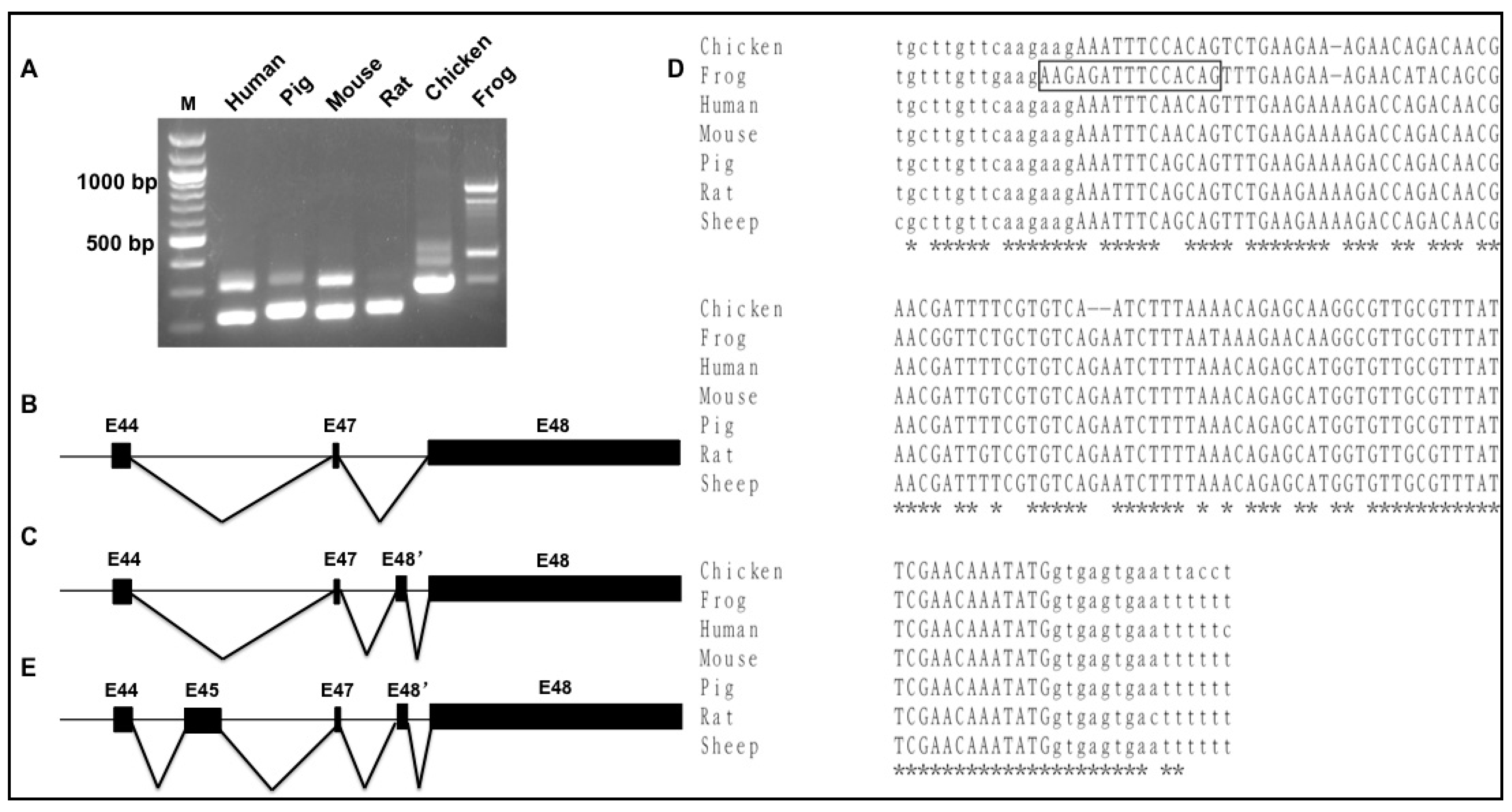
3.4. Splicing Variants of the Novex-3-Specific Exon 48 across Species
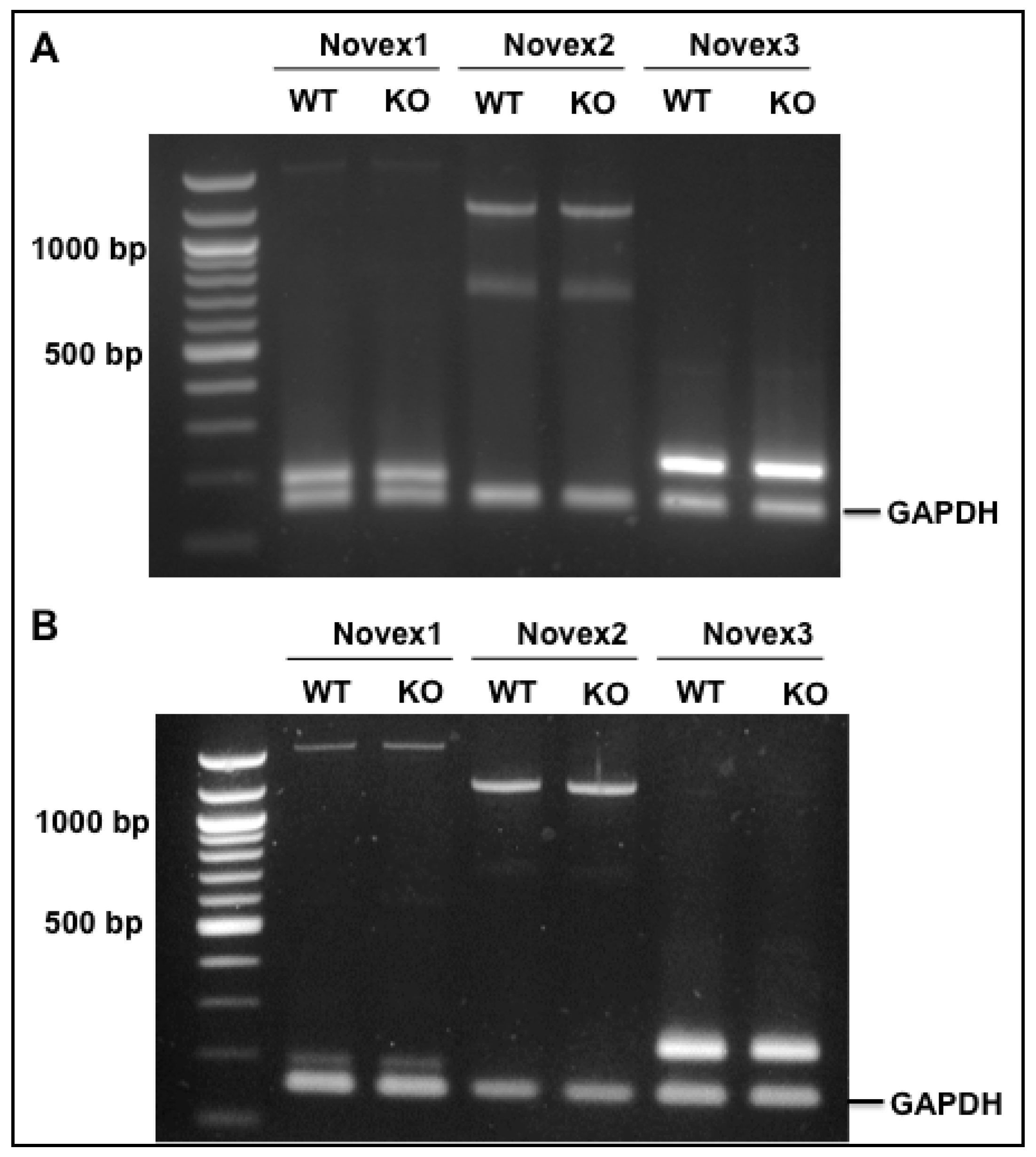
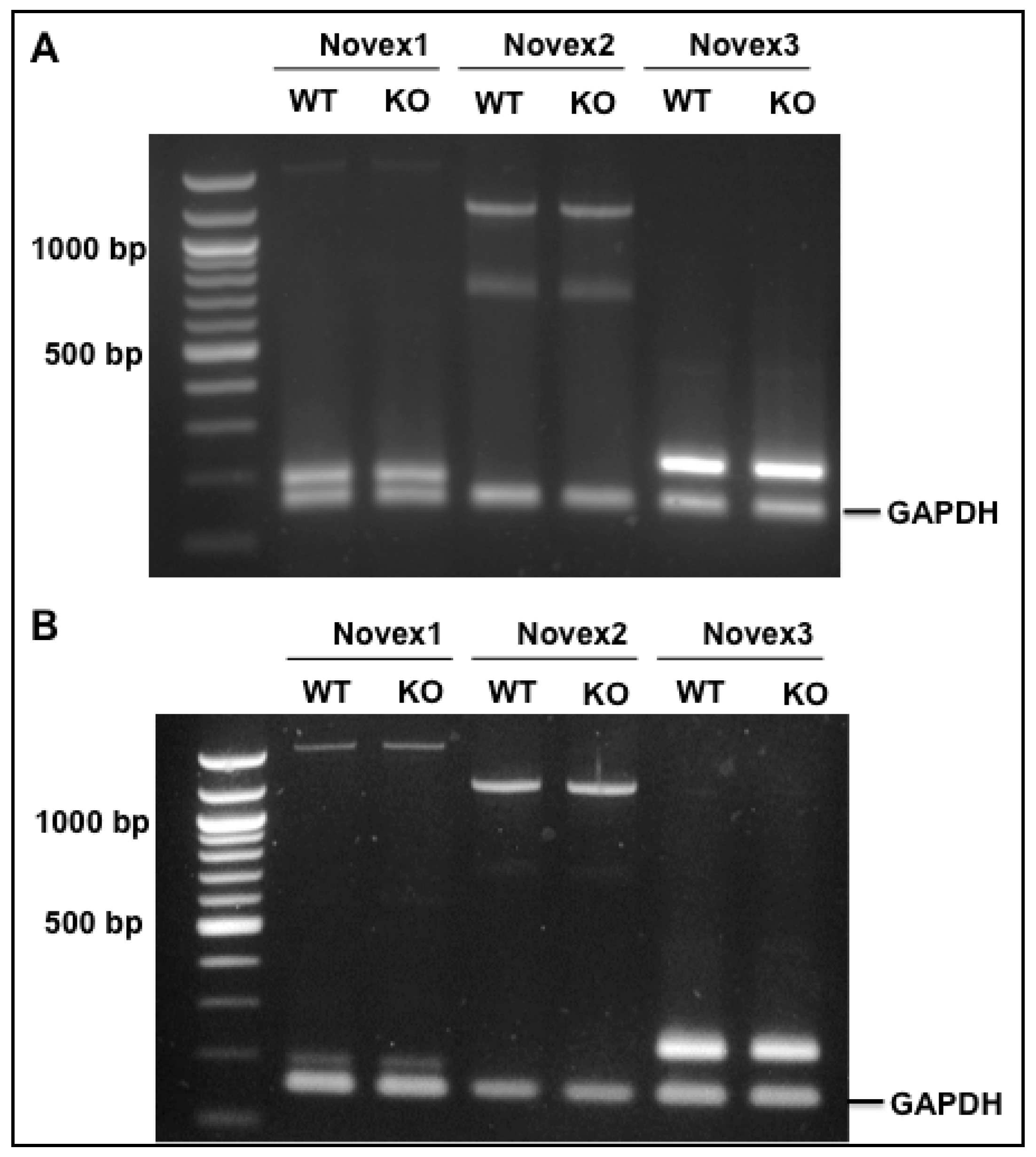
3.5. Splicing Regulation of RBM20 in the Novex 1, 2, and 3 Isoforms
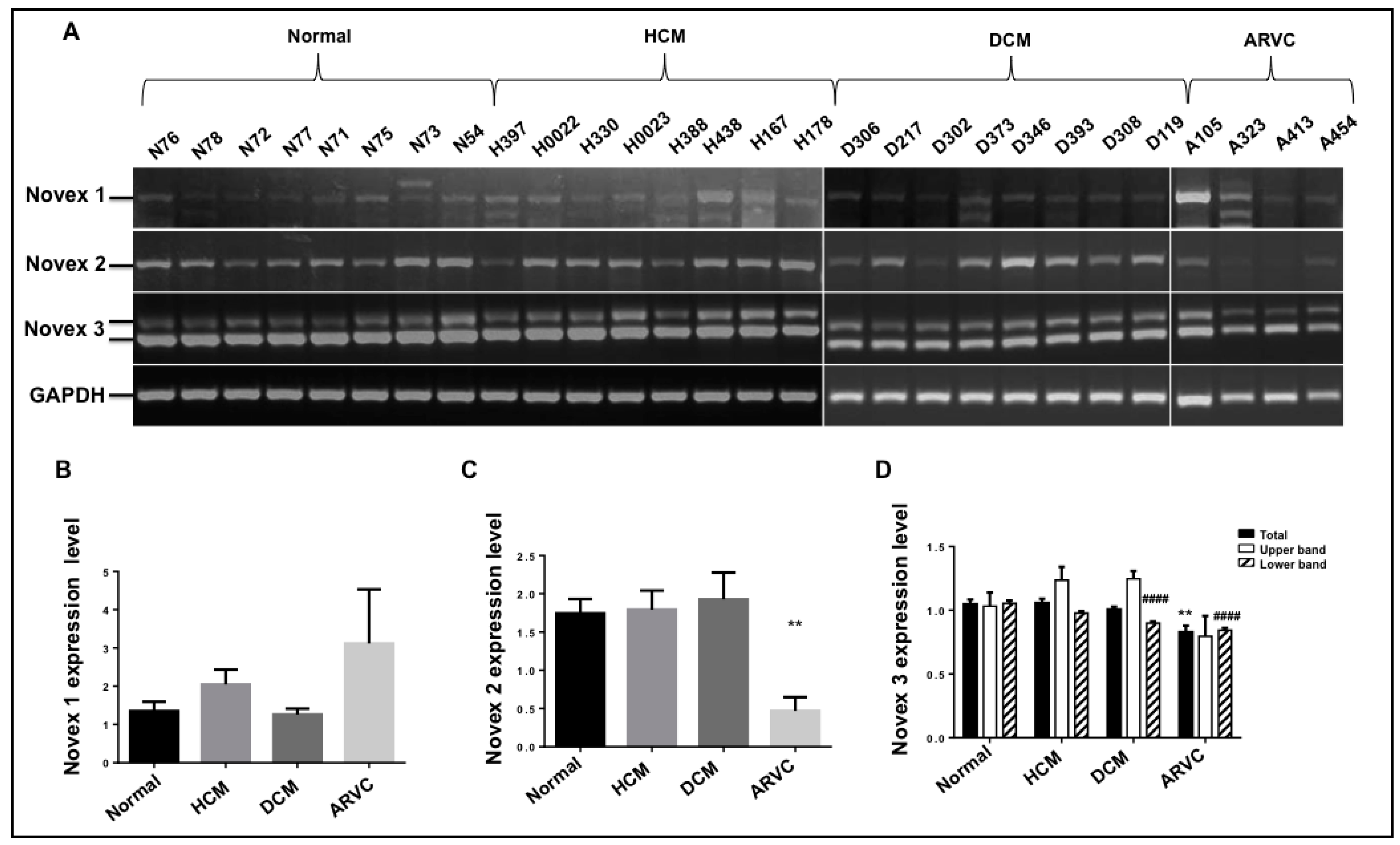
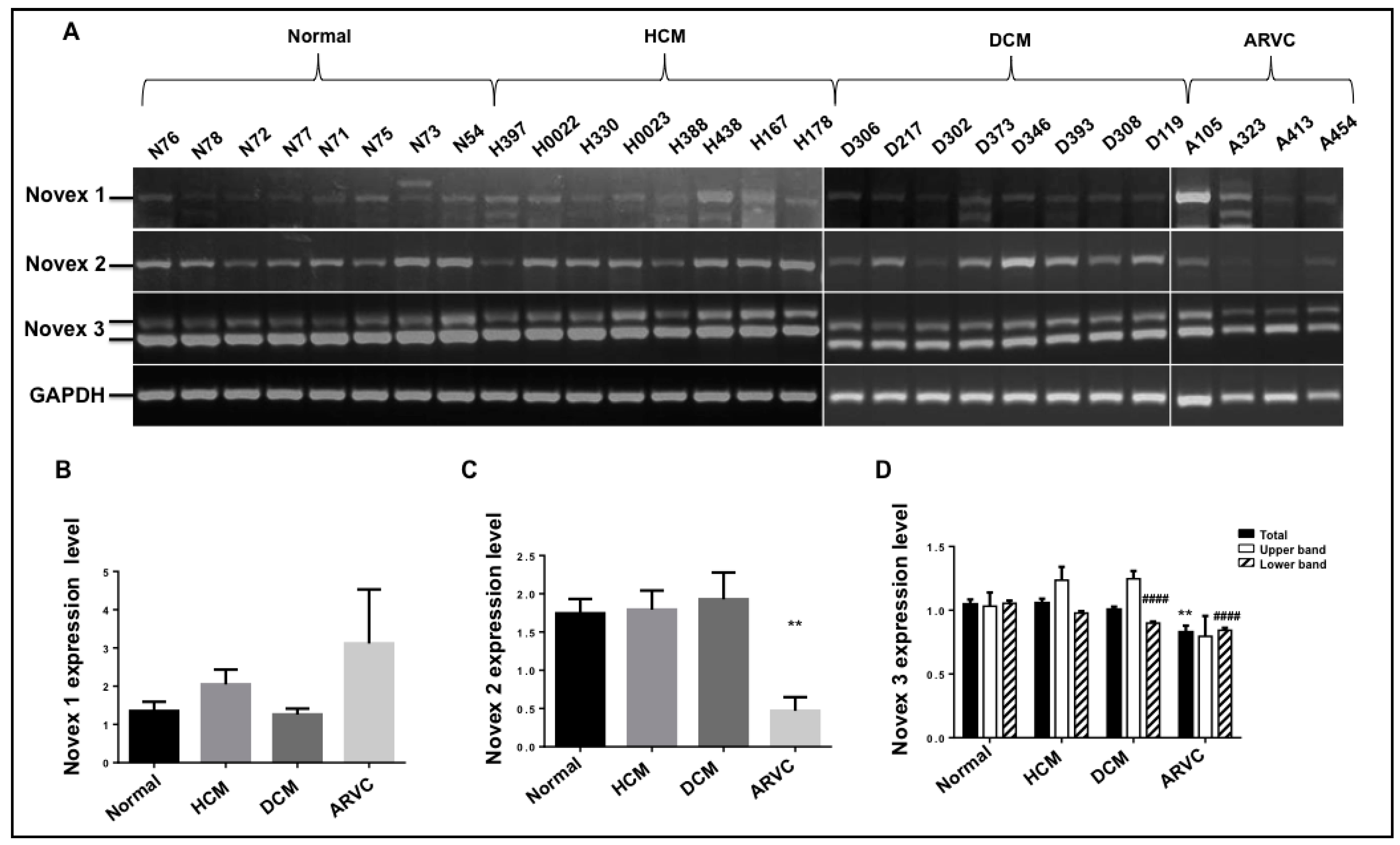
3.6. Splicing Pattern of the Novexes 1, 2, and 3 in Human Cardiomyopathies
3.7. In Silico Analysis of Titin Novex-1, -2, and -3 Exons 42–50
4. Discussion
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Wang, E.T.; Sandberg, R.; Luo, S.; Khrebtukova, I.; Zhang, L.; Mayr, C.; Kingsmore, S.F.; Schroth, G.P.; Burge, C.B. Alternative isoform regulation in human tissue transcriptomes. Nature 2008, 456, 470–476. [Google Scholar] [CrossRef] [PubMed]
- Johnson, J.M.; Castle, J.; Garrett-Engele, P.; Kan, Z.; Loerch, P.M.; Armour, C.D.; Santos, R.; Schadt, E.E.; Stoughton, R.; Shoemaker, D.D. Genome-wide survey of human alternative pre-mRNA splicing with exon junction microarrays. Science 2003, 302, 2141–2144. [Google Scholar] [CrossRef] [PubMed]
- Zhu, C.; Chen, Z.; Guo, W. Pre-mRNA mis-splicing of sarcomeric genes in heart failure. Biochim. Biophys. Acta (BBA) Mol. Basis Dis. 2016, 8, 2056–2063. [Google Scholar] [CrossRef] [PubMed]
- Davis, F.J.; Gupta, M.; Pogwizd, S.M.; Bacha, E.; Jeevanandam, V.; Gupta, M.P. Increased expression of alternatively spliced dominant-negative isoform of SRF in human failing hearts. Am. J. Physiol. Heart Circ. Physiol. 2002, 282, H1521–H1533. [Google Scholar] [CrossRef] [PubMed]
- Kong, S.W.; Hu, Y.W.; Ho, J.W.; Ikeda, S.; Polster, S.; John, R.; Hall, J.L.; Bisping, E.; Pieske, B.; Dos, R.C.; et al. Heart failure-associated changes in RNA splicing of sarcomere genes. Circ. Cardiovasc. Genet. 2010, 3, 138–146. [Google Scholar] [CrossRef] [PubMed]
- Bang, M.L.; Centner, T.; Fornoff, F.; Geach, A.J.; Gotthardt, M.; McNabb, M.; Witt, C.C.; Labeit, D.; Gregorio, C.C.; Granzier, H.; et al. The complete gene sequence of titin, expression of an unusual approximately 700-kDa titin isoform, and its interaction with obscuring identify a novel Z-line to I-band linking system. Circ. Res. 2001, 89, 1065–1072. [Google Scholar] [CrossRef] [PubMed]
- Granzier, H.L.; Irving, T.C. Passive tension in cardiac muscle: Contribution of collagen, titin, microtubules, and intermediate filaments. Biophys. J. 1995, 68, 1027–1044. [Google Scholar] [CrossRef]
- Herman, D.S.; Lam, L.; Taylor, M.R.; Wang, L.; Teekakirikul, P.; Christodoulou, D.; Conner, L.; DePalma, S.R.; McDonough, B.; Sparks, E.; et al. Truncations of titin causing dilated cardiomyopathy. N. Engl. J. Med. 2012, 366, 619–628. [Google Scholar] [CrossRef] [PubMed]
- LeWinter, M.M.; Granzier, H.L. Cardiac titin and heart disease. J. Cardiovasc. Pharmacol. 2014, 63, 207–212. [Google Scholar] [CrossRef] [PubMed]
- Taylor, M.; Graw, S.; Sinagra, G.; Barnes, C.; Slavov, D.; Brun, F.; Pinamonti, B.; Salcedo, E.E.; Sauer, W.; Pyxaras, S. Genetic variation in titin in arrhythmogenic right ventricular cardiomyopathy–overlap Syndromes Clinical perspective. Circulation 2011, 124, 876–885. [Google Scholar] [CrossRef] [PubMed]
- Linke, W.A.; Hamdani, N. Gigantic business: Titin properties and function through thick and thin. Circ. Res. 2014, 114, 1052–1068. [Google Scholar] [CrossRef] [PubMed]
- Yin, Z.; Ren, J.; Guo, W. Sarcomeric protein isoform transitions in cardiac muscle: A journey to heart failure. Biochim. Biophys. Acta (BBA) Mol. Basis Dis. 2015, 1852, 47–52. [Google Scholar] [CrossRef] [PubMed]
- Warren, C.M.; Krzesinski, P.R.; Campbell, K.S.; Moss, R.L.; Greaser, M.L. Titin isoform changes in rat myocardium during development. Mech. Dev. 2004, 121, 1301–1312. [Google Scholar] [CrossRef] [PubMed]
- Guo, W.; Bharmal, S.J.; Esbona, K.; Greaser, M.L. Titin diversity—Alternative splicing gone wild. J. Biomed. Biotechnol. 2010, 2010, 675–753. [Google Scholar] [CrossRef] [PubMed]
- Linke, W.A. Sense and stretchability: The role of titin and titin-associated proteins in myocardial stress-sensing and mechanical dysfunction. Cardiovasc. Res. 2008, 77, 637–648. [Google Scholar] [CrossRef] [PubMed]
- Guo, W.; Sun, M. RBM20, a potential target for treatment of cardiomyopathy via titin isoform switching. Biophys. Rev. 2017. [Google Scholar] [CrossRef] [PubMed]
- Tskhovrebova, L.; Trinick, J. Properties of titin immunoglobulin and fibronectin-3 domains. J. Biol. Chem. 2004, 279, 46351–46354. [Google Scholar] [CrossRef] [PubMed]
- Puchner, E.M.; Alexandrovich, A.; Kho, A.L.; Hensen, U.; Schafer, L.V.; Brandmeier, B.; Grater, F.; Grubmuller, H.; Gaub, H.E.; Gautel, M. Mechanoenzymatics of titin kinase. Proc. Natl. Acad. Sci. USA 2008, 105, 13385–13390. [Google Scholar] [CrossRef] [PubMed]
- Guo, W.; Schafer, S.; Greaser, M.L.; Radke, M.H.; Liss, M.; Govindarajan, T.; Maatz, H.; Schulz, H.; Li, S.; Parrish, A.M.; et al. RBM20, a gene for hereditary cardiomyopathy, regulates titin splicing. Nat. Med. 2012, 18, 766–773. [Google Scholar] [CrossRef] [PubMed]
- Hidalgo, C.; Granzier, H. Tuning the molecular giant titin through phosphorylation: Role in health and disease. Trends Cardiovasc. Med. 2013, 23, 165–171. [Google Scholar] [CrossRef] [PubMed]
- Methawasin, M.; Hutchinson, K.R.; Lee, E.J.; Smith, J.R.; Saripalli, C.; Hidalgo, C.G.; Ottenheijm, C.A.; Granzier, H. Experimentally increasing titin compliance in a novel mouse model attenuates the Frank-Starling mechanism but has a beneficial effect on diastole. Circulation 2014, 129, 1924–1936. [Google Scholar] [CrossRef] [PubMed]
- Trombitas, K.; Redkar, A.; Centner, T.; Wu, Y.; Labeit, S.; Granzier, H. Extensibility of isoforms of cardiac titin: Variation in contour length of molecular subsegments provides a basis for cellular passive stiffness diversity. Biophys. J. 2000, 79, 3226–3234. [Google Scholar] [CrossRef]
- Bell, S.P.; Nyland, L.; Tischler, M.D.; McNabb, M.; Granzier, H.; LeWinter, M.M. Alterations in the determinants of diastolic suction during pacing tachycardia. Circ. Res. 2000, 87, 235–240. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Bell, S.P.; Trombitas, K.; Witt, C.C.; Labeit, S.; LeWinter, M.M.; Granzier, H. Changes in titin isoform expression in pacing-induced cardiac failure give rise to increased passive muscle stiffness. Circulation 2002, 106, 1384–1389. [Google Scholar] [CrossRef] [PubMed]
- Shapiro, B.P.; Lam, C.S.; Patel, J.B.; Mohammed, S.F.; Kruger, M.; Meyer, D.M.; Linke, W.A.; Redfield, M.M. Acute and chronic ventricular-arterial coupling in systole and diastole: Insights from an elderly hypertensive model. Hypertension 2007, 50, 503–511. [Google Scholar] [CrossRef] [PubMed]
- Hamdani, N.; Bishu, K.G.; von Frieling-Salewsky, M.; Redfield, M.M.; Linke, W.A. Deranged myofilament phosphorylation and function in experimental heart failure with preserved ejection fraction. Cardiovasc. Res. 2013, 97, 464–471. [Google Scholar] [CrossRef] [PubMed]
- Warren, C.M.; Jordan, M.C.; Roos, K.P.; Krzesinski, P.R.; Greaser, M.L. Titin isoform expression in normal and hypertensive myocardium. Cardiovasc. Res. 2003, 59, 86–94. [Google Scholar] [CrossRef]
- Hudson, B.; Hidalgo, C.; Saripalli, C.; Granzier, H. Hyperphosphorylation of mouse cardiac titin contributes to transverse aortic constriction-induced diastolic dysfunction. Circ. Res. 2011, 109, 858–866. [Google Scholar] [CrossRef] [PubMed]
- Neagoe, C.; Kulke, M.; Del, M.F.; Gwathmey, J.K.; de Tombe, P.P.; Hajjar, R.J.; Linke, W.A. Titin isoform switch in ischemic human heart disease. Circulation 2002, 106, 1333–1341. [Google Scholar] [CrossRef] [PubMed]
- Makarenko, I.; Opitz, C.A.; Leake, M.C.; Neagoe, C.; Kulke, M.; Gwathmey, J.K.; Del, M.F.; Hajjar, R.J.; Linke, W.A. Passive stiffness changes caused by upregulation of compliant titin isoforms in human dilated cardiomyopathy hearts. Circ. Res. 2004, 95, 708–716. [Google Scholar] [CrossRef] [PubMed]
- Nagueh, S.F.; Shah, G.; Wu, Y.; Torre-Amione, G.; King, N.M.; Lahmers, S.; Witt, C.C.; Becker, K.; Labeit, S.; Granzier, H.L. Altered titin expression, myocardial stiffness, and left ventricular function in patients with dilated cardiomyopathy. Circulation 2004, 110, 155–162. [Google Scholar] [CrossRef] [PubMed]
- Borbély, A.; Falcao-Pires, I.; Van Heerebeek, L.; Hamdani, N.; Edes, I.; Gavina, C.; Leite-Moreira, A.F.; Bronzwaer, J.G.; Papp, Z.; Van Der Velden, J. Hypophosphorylation of the Stiff N2B titin isoform raises cardiomyocyte resting tension in failing human myocardium. Circ. Res. 2009, 104, 780–786. [Google Scholar] [CrossRef] [PubMed]
- Hinze, F.; Dieterich, C.; Radke, M.H.; Granzier, H.; Gotthardt, M. Reducing RBM20 activity improves diastolic dysfunction and cardiac atrophy. J. Mol. Med. (Berl.) 2016, 94, 1349–1358. [Google Scholar] [CrossRef] [PubMed]
- Kent, W.J.; Sugnet, C.W.; Furey, T.S.; Roskin, K.M.; Pringle, T.H.; Zahler, A.M.; Haussler, D. The human genome browser at UCSC. Genome Res. 2002, 12, 996–1006. [Google Scholar]
- Kersey, P.J.; Allen, J.E.; Allot, A.; Barba, M.; Boddu, S.; Bolt, B.J.; Carvalho-Silva, D.; Christensen, M.; Davis, P.; Grabmueller, C.; et al. Ensembl Genomes 2018: An integrated omics infrastructure for non-vertebrate species. Nucleic Acids Res. 2018, 46, D802–D808. [Google Scholar] [CrossRef] [PubMed]
- O’Leary, N.A.; Wright, M.W.; Brister, J.R.; Ciufo, S.; Haddad, D.; McVeigh, R.; Rajput, B.; Robbertse, B.; Smith-White, B.; Ako-Adjei, D.; et al. Reference sequence (RefSeq) database at NCBI: Current status, taxonomic expansion, and functional annotation. Nucleic Acids Res. 2016, 44, D733–D745. [Google Scholar] [CrossRef] [PubMed]
- McGinnis, S.; Madden, T.L. BLAST: At the core of a powerful and diverse set of sequence analysis tools. Nucleic Acids Res. 2004, 32, W20–W25. [Google Scholar] [CrossRef] [PubMed]
- Jeanmougin, F.; Thompson, J.D.; Gouy, M.; Higgins, D.G.; Gibson, T.J. Multiple sequence alignment with Clustal X. Trends Biochem. Sci. 1998, 23, 403–405. [Google Scholar] [CrossRef]
- Frazer, K.A.; Pachter, L.; Poliakov, A.; Rubin, E.M.; Dubchak, I. VISTA: Computational tools for comparative genomics. Nucleic Acids Res. 2004, 32, W273–W279. [Google Scholar] [CrossRef] [PubMed]
- Schneider, C.A.; Rasband, W.S.; Eliceiri, K.W. NIH Image to ImageJ: 25 years of image analysis. Nat. Methods 2012, 9, 671–675. [Google Scholar] [CrossRef] [PubMed]
- Kellermayer, D.; Smith, J.R.; Granzier, H. Novex-3, the tiny titin of muscle. Biophys. Rev. 2017, 9, 201–206. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Guo, W.; Dewey, C.N.; Greaser, M.L. Rbm20 regulates titin alternative splicing as a splicing repressor. Nucleic Acids Res. 2013, 41, 2659–2672. [Google Scholar] [CrossRef] [PubMed]
- Guo, W.; Pleitner, J.M.; Saupe, K.W.; Greaser, M.L. Pathophysiological Defects and Transcriptional Profiling in the RBM20-/- Rat Model. PLoS ONE 2013, 8, e84281. [Google Scholar] [CrossRef] [PubMed]
- Furst, D.O.; Osborn, M.; Nave, R.; Weber, K. The organization of titin filaments in the half-sarcomere revealed by monoclonal antibodies in immunoelectron microscopy: A map of ten nonrepetitive epitopes starting at the Z line extends close to the M line. J. Cell Biol. 1988, 106, 1563–1572. [Google Scholar] [CrossRef] [PubMed]
- Labeit, S.; Barlow, D.P.; Gautel, M.; Gibson, T.; Holt, J.; Hsieh, C.L.; Francke, U.; Leonard, K.; Wardale, J.; Whiting, A.; et al. A regular pattern of two types of 100-residue motif in the sequence of titin. Nature 1990, 345, 273–276. [Google Scholar] [CrossRef] [PubMed]
- Freiburg, A.; Gautel, M. A molecular map of the interactions between titin and myosin-binding protein C. Implications for sarcomeric assembly in familial hypertrophic cardiomyopathy. Eur. J. Biochem. 1996, 235, 317–323. [Google Scholar] [CrossRef] [PubMed]
- Granzier, H.L.; Labeit, S. The giant protein titin: A major player in myocardial mechanics, signaling, and disease. Circ. Res. 2004, 94, 284–295. [Google Scholar] [CrossRef] [PubMed]
- Tskhovrebova, L.; Trinick, J. Titin: Properties and family relationships. Nat. Rev. Mol. Cell Biol. 2003, 4, 679–689. [Google Scholar] [CrossRef] [PubMed]
- Granzier, H.; Labeit, S. Cardiac titin: An adjustable multi-functional spring. J. Physiol. 2002, 541, 335–342. [Google Scholar] [CrossRef] [PubMed]
- Linke, W.A.; Fernandez, J.M. Cardiac titin: Molecular basis of elasticity and cellular contribution to elastic and viscous stiffness components in myocardium. J. Muscle Res. Cell Motil. 2002, 23, 483–497. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Regions | Human | Pig | Rat | Mouse | Chicken | Lizard | Frog | Zebrafish | |
|---|---|---|---|---|---|---|---|---|---|
| Species | |||||||||
| Exon42 | 285 | 285 | 285 | 285 | 285 | 285 | 285 | 285 | |
| Intron42 | 224 | 222 | 177 | 188 | 100 | 124 | 108 | 83 | |
| Exon43 | 126 | 126 | 126 | 126 | 126 | 126 | 126 | 126 | |
| Intron43 | 5004 | 3844 | 4470 | 4856 | 1795 | 3190 | 584 | 634 | |
| Exon44 | 189 | 189 | 189 | 189 | 189 | 189 | 189 | 189 | |
| Intron44 | 1067 | 1095 | 1382 | 1107 | 723 | 1564 | 559 | 2350 | |
| Exon45 (Novex 1 exon) | 375 | 375 | 375 | 375 | 375 | 372 | 375 | 390 | |
| Intron45 | 744 | 3990 | 3779 | 3997 | 628 | 671 | 571 | 357 | |
| Exon46 (Novex 2 exon) | 573 a | 582 | 558 | 552 | |||||
| Intron46 | 3044 | 1898 | 3257 | 2337 | |||||
| Exon47 | 57 | 45 | 57 | 57 | 57 | 57 | 48 | 51 | |
| Intron47 | 1084 | 1091 | 1094 | 1087 | 1102 | 1105 | 1021 | 1022 | |
| Exon48 b (Novex 3 exon) | 6455 | 6443 | 6266 | 6287 | 6077 | 8039 | 7088 | 6866 | |
| Intron47 c | 11,202 | 12,096 | 10,890 | 11,597 | 11,301 | 14,386 | 12,571 | 8954 | |
| Exon49 (N2B exon) | 2781 | 2742 | 2628 | 2646 | 2544 | 3846 | 3630 | 3771 | |
| Intron49 | 780 | 773 | 792 | 775 | 562 | 876 | 1298 | 919 | |
| Exon50 | 279 | 279 | 279 | 279 | 279 | 279 | 276 | 279 | |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, Z.; Song, J.; Chen, L.; Zhu, C.; Cai, H.; Sun, M.; Stern, A.; Mozdziak, P.; Ge, Y.; Means, W.J.; et al. Characterization of TTN Novex Splicing Variants across Species and the Role of RBM20 in Novex-Specific Exon Splicing. Genes 2018, 9, 86. https://doi.org/10.3390/genes9020086
Chen Z, Song J, Chen L, Zhu C, Cai H, Sun M, Stern A, Mozdziak P, Ge Y, Means WJ, et al. Characterization of TTN Novex Splicing Variants across Species and the Role of RBM20 in Novex-Specific Exon Splicing. Genes. 2018; 9(2):86. https://doi.org/10.3390/genes9020086
Chicago/Turabian StyleChen, Zhilong, Jiangping Song, Liang Chen, Chaoqun Zhu, Hanfang Cai, Mingming Sun, Allysa Stern, Paul Mozdziak, Ying Ge, Warrie J. Means, and et al. 2018. "Characterization of TTN Novex Splicing Variants across Species and the Role of RBM20 in Novex-Specific Exon Splicing" Genes 9, no. 2: 86. https://doi.org/10.3390/genes9020086