Differential Expression of Six Rnase2 and Three Rnase3 Paralogs Identified in Blunt Snout Bream in Response to Aeromonas hydrophila Infection

1
Key Lab of Agricultural Animal Genetics, Breeding and Reproduction of Ministry of Education/Key Lab of Freshwater Animal Breeding, College of Fisheries, Huazhong Agricultural University, Wuhan 430070, China
2
Key Laboratory of Freshwater Fisheries and Germplasm Resources Utilization, Chinese Academy of Fishery Sciences, Wuxi 214081, China
3
Collaborative Innovation Center for Efficient and Health Production of Fisheries in Hunan Province, Changde 415000, China
*
Authors to whom correspondence should be addressed.
Genes 2018, 9(2), 95; https://doi.org/10.3390/genes9020095
Submission received: 22 December 2017
/
Revised: 5 February 2018
/
Accepted: 6 February 2018
/
Published: 14 February 2018
(This article belongs to the Section Animal Genetics and Genomics)
Abstract
:Ribonucleases (Rnases)2 and Rnase3 belong to the ribonuclease A (RnaseA) superfamily. Apart from their role in molecular evolutionary and functional biological studies, these genes have also been studied in the context of defense against pathogen infection in mammals. However, expression patterns, structures and response to bacterial infection of the two genes in blunt snout bream (Megalobrama amblycephala) remain unknown. In this study, we identified multiple copies of Rnase2 (six) and Rnase3 (three) in the M. amblycephala genome. The nine genes all possess characteristics typical of the RnaseA superfamily. No expression was detected in the early developmental stages, while a weak expression was observed at 120 and 140 h post-fertilization (hpf) for Rnase2b, Rnase2c, Rnase2e and Rnase3a, suggesting that only three copies of Rnase2 and one of Rnase3 are expressed. Interestingly, only Rnase2e was up-regulated in the kidney of M. amblycephala after Aeromonas hydrophila infection, while Rnase3a was significantly up-regulated in liver, gut and blood after the infection. We conclude that the paralogs of Rnase3 are more susceptible to A. hydrophila infection than Rnase2. These results indicate that different Rnase2 and Rnase3 paralogs suggest a role in the innate immune response of M. amblycephala to bacterial infection.
1. Introduction
The ribonuclease A (RnaseA) superfamily, also known as the pancreatic ribonuclease (Rnase) family, is the sole vertebrate-specific enzyme family [1,2]. The superfamily has been studied rather extensively in mammals, including humans, cattle, monkeys and mice, but studies in fishes remain comparatively scarce. In fishes, Rnase1 to 5 from Danio rerio [3,4,5,6], Rnase1 to 3 from Oryzias latipes and Rnase1-2 from Salmo salar [7] have been reported. All members of this superfamily have a few common characteristics: the entire coding sequence is contained within a single exon, a peptide signal at the N-terminal that directs protein biosynthesis within the endoplasmic reticulum, 6–8 conserved cysteines which form disulfide bonds, a catalytic triad composed of the conserved “CKXXNTF” functional motif, and finally two histidine residues and one lysine residue with catalytic activity [8]. However, physiological functions of the superfamily members, as well as their expression patterns in tissues, are distinct. Rnase1 is expressed in the pancreas, brain and kidney; Rnase4 mainly in liver and gut; Rnase5 in liver and testis; and Rnase6 in liver and brain [9]. RnaseA superfamily members have a wide variety of different functions, including the degrading of dietary RNAs [1], antibacterial activities [10], stimulation of blood vessel formation [11], host defense [12], and reproductive functions [13,14].
Rnase2 is a cytotoxic protein secreted together with the Rnase3 by cytoplasmic granules of eosinophils in mammals [15]. It is known to be expressed in the liver, spleen, placenta in humans and ruminants [1,16]. In mammals, Rnase2 shows the specific antiviral activity in vitro experiments [17]. Rnase2 was able to reduce the infection rate of the respiratory syncytial virus (RSV) and human immunodeficiency virus (HIV) in cell culture experiments [18,19]. Rnase2 exhibits chemotactic properties and plays an important role in the regulation of autoimmunity [20]. However, the expression patterns and biological functions of Rnase2 in mammals are different from the fish species. In fishes, gene expression analysis showed that D. rerio Rnase1 and Rnase2 are mainly in the adult liver and gut, and weakly expressed in the heart. Functional analysis of recombinant proteins demonstrated that D. rerio Rnases have strong antibacterial activity in vitro [3]. Rnase1 and Rnase2 in adult S. salar, mainly expressed in the thymus, have both angiogenic and bactericidal activities [7]. Rnase2 has 100-fold higher ribonuclease activity than Rnase3 [21].
Rnase3 is a host defense ribonuclease involved in inflammatory responses in humans. Molecular cloning experiments confirmed that both Rnase2 and Rnase3 (67% amino acid sequence similarity) are members of the RnaseA superfamily [22], and both have since been shown to be active ribonucleases [23]. Rnase3 exhibits high antibacterial activity against both Gram-positive and Gram-negative bacteria, as well as antiviral activity and antiparasitic functions against helminths and protozoa [24,25,26]. In fishes, Rnase3 of D. rerio is highly expressed in heart and gut, and has bactericidal activity [3]. Rnase1 gene, a member of RnaseA superfamily, has multiple copies. Duplications of Rnase1 have been observed in Myotis altarium (Rnase1α, Rnase1β and Rnase1γ), Martes flavigula (Rnase1-1, Rnase1-2 and Rnase1-3), Callosciurus prevostii (Rnase1α and Rnase1β), and Colobus guereza (Rnase1α, Rnase1β and Rnase1γ), etc. [27,28,29]. However, there is no genetic duplication of Rnase1, Rnase2 and Rnase3 in teleost. It has been suggested that these varied functions emerged after gene duplication under a positive Darwinian selection [27].
The blunt snout bream (Megalobrama amblycephala) has recently become one of the economically most important freshwater cultured fishes in China. It is recognized as an ecofriendly and resource-conserving fish because of its fast growth, herbivorous diet, ease and low costs of breeding and high nutritional value. With rapid development of the M. amblycephala aquaculture industry, infectious diseases are becoming increasingly prevalent, resulting in substantial economic losses [30,31]. Ma-Rnase1 has been cloned and characterized, and its recombinant protein was found to have digestive activity and antibacterial function [29]. Recently, our laboratory has published the entire genome of this species [32], so now we can identify all members of the RnaseA superfamily present in its genome. Here, we identified six Rnase2 paralogs (Rnase2a, Rnase2b, Rnase2c, Rnase2d, Rnase2e and Rnase2f) and three Rnase3 paralogs (Rnase3a, Rnase3b and Rnase3c) in the genome of M. amblycephala. Three-dimensional structures of all proteins were modeled and compared. The messenger RNA (mRNA) expression levels of all nine genes (6+3) were determined by quantitative polymerase chain reaction (qPCR) in the early developmental stages, as well as in different tissues of one-year-old and two-year-old M. amblycephala. In order to infer their functional roles, we also studied the expression patterns of these genes in five different tissues at 6 and 12 h after an immune-challenge with Aeromonas hydrophila. Our results are not only useful for the research of Rnase2 and Rnase3 evolution, but also help further elucidation of the regulation mechanisms in the innate immune response of M. amblycephala to bacterial infection.
2. Materials and Methods
2.1. Collection of Samples
Sample collection and experiments were conducted in accordance with the national legislation of China and approved by the ethics committee of Huazhong Agricultural University (No. HZAUFI-2017-006).
Healthy one-year-old (n = 10) and two-year-old (n = 70) M. amblycephala specimens, as well as three female and three male adult specimens for obtaining embryos via artificial breeding, were obtained from the Tuanfeng fish farm (Huanggang city, Hubei province, China), and kept in tanks at the Freshwater Fish Genetics Breeding Center of Huazhong Agricultural University (Wuhan, China). The fish (one- and two-year-old) were acclimated for two weeks, while maintaining water temperature at 26 ± 2 °C and an abundant oxygen level. They were fed commercial pellet feed twice daily for seven days before the onset of the study. Healthy two-year-old fish were divided into three groups: 20 “blank” (non-injected), 30 “challenge” (A. hydrophila injected) and 30 “control” (phosphate-buffered saline (PBS) injected). After the specimens from the blank group were anesthetized with 100 mg/L tricaine methane sulfonate (MS-222) and sterilized with 75% alcohol, eight type-tissue samples were immediately collected under sterile conditions from one- and two-year-old (adult) M. amblycephala: brain, heart, liver, spleen, kidney, muscle, gut, and testis. Seven different early developmental embryonic stages were also sampled: fertilized egg (0 h post-fertilization (hpf)), late gastrula stage (16 hpf), heart appearance (38 hpf), hatching (48 hpf), eye appearance (88 hpf), air bladder formation (120 hpf) and intestine appearance stage (140 hpf). All samples collected were immediately frozen in liquid nitrogen and stored at −80 °C.
2.2. Identification of Rnase2 and Rnase3 Genes
The coding sequences of Ma-Rnase1, Ma-Rnase2 and Ma-Rnase3 were obtained from our previous transcriptomic data for M. amblycephala (GenBank accession numbers: SRX731259 and SRA045792). The full-length sequences of Ma-Rnase1, Ma-Rnase2 and Ma-Rnase3 were obtained from the genome data of M. amblycephala (Figure S1).
2.3. Protein Alignment and Physicochemical Properties
The protein sequences of Ma-Rnase1, Ma-Rnase2 and Ma-Rnase3 were inferred from the genome data of M. amblycephala. The protein sequences of Danio rerio used for the analysis were obtained from the GenBank: Rnase1 (ABQ23783.1), Rnase2 (ABQ23784.1) and Rnase3 (ABQ2385.1). Orthologs were then aligned by CLC Sequence Viewer 6.8 (Qiagen, Hilden, Germany). Physicochemical properties, as well as functional sites and domains, of all nine proteins (Rnase2a, Rnase2b, Rnase2c, Rnase2d, Rnase2e, Rnase2f, Rnase3a, Rnase3b and Rnase3c) were predicted using different tools available from the ExPASy web server [33]. SignalP 4.1 server [34] was used to predict the signal peptide cleavage sites of the N-terminals of proteins. The isoelectric point (pI) and molecular weight (Mw) of the mature proteins were calculated with the ExPASy online tool Compute pI/Mw.
2.4. Structural Predictions
Three-dimensional structures of proteins were modeled with the SWISS-MODEL server, selecting nine gene models in the protein structure model database to build the models [35]. The sequence identity values between the template (Rnase1) and the nine studied genes (Rnase2a-f and Rnase3a-c) were 60%, 62%, 58%, 50%, 57%, 50%, 33%, 33% and 25%, respectively. Next, the secondary structures of Ma-Rnase1-3 protein sequences were created using ESPript software [36]. Superimposed three-dimensional structures and functional sites of the nine studied genes (with template) were modeled by PYMOL (DeLano Scientific) [37].
2.5. Phylogenetic Analysis
MEGA 7.0.21 [38] was used to conduct phylogenetic analysis aimed to better understand the evolutionary relationships of Rnase1-3 among the teleosts (M. amblycephala, D. rerio, O. latipes, S. salar, Oncorhynchus mykiss) and mammals (Bos taurus, Mus musculus, Homo sapiens, Rattus norvegicus, Macaca mulatta and Colobus angolensis). All species sequences were downloaded from the GenBank [39]. Amino acid sequences were aligned with the Muscle program. The phylogenetic tree was reconstructed using the Neighbor-Joining (NJ) method with the protein Jones-Taylor-Thornton (JTT) matrix model and 1000 bootstrap replications [40].
2.6. Quantitative Analysis of Rnase2 and Rnase3 Messenger RNA Expression
Total RNA was isolated from each sample using TRIzol Reagent (TaKaRa Bio Inc., Dalian, China), following the manufacturer’s instructions. All RNA samples were treated with RNase-free DNase (TaKaRa Bio Inc.). The quality and quantity of RNA were evaluated with 1% agarose gel electrophoresis and the NanoDrop ND-2000 spectrophotometer (Thermo Fisher Scientific, Waltham, MA, USA), and then stored at −80 °C. We amplified the mRNA sequences of these nine genes with the primers presented in Table 1. The primers for the reference gene β-actin were 5’-ACCCACACCGTGCCCATCTA-3’ (forward) and 5’-CGGACAATTTCTCTTTCGGCTG-3’ (reverse) [41]. The first-strand cDNA was synthesized using the Prime Script™ RT reagent Kit with genomic DNA (gDNA) eraser (TaKaRa Bio Inc.), according to the manufacturer’s instructions. To analyze the expression patterns of Rnase2 and Rnase3 in healthy fish tissues and developing fish, all samples were analyzed using Light Cycler® 480 II qPCR detection system (Roche Diagnostics, Mannheim, Germany). qPCR analysis were performed in a 96-well plate, where each well contained 20 μL of reaction mixture consisting of: 10 μL SYBR® PreMix Ex Tapm II (TaKaRa Bio Inc., Dalian, China), 0.4 μL each primer (10 μM), 2 μL complementary DNA (cDNA) template and 7.2 μL sterilized double distilled water (ddH2O). The qPCR conditions were as follows: pre-denaturation at 95 °C for 5 min, followed by 40 cycles of amplification at 95 °C for 10 s, 60 °C for 10 s, and 72 °C for 15 s. Each sample was tested in triplicate. The average value per gene was calculated from three replicates. Finally, the expression of each gene was calculated using the 2−△△Ct method [42].
2.7. Bacterial Challenge Experiment
For the bacterial challenge experiment, A. hydrophila was obtained from the Laboratory of Aquatic Medicine of the College of Fisheries at Huazhong Agricultural University (Hubei, China). A. hydrophila was incubated at 28 °C for 24 h on a Luria-Bertani Medium (LB) plate. We intraperitoneally injected the experimental group (two-year-old healthy M. amblycephala) with 1.0 × 107 colony-forming units/mL of A. hydrophila (0.1 mL). The control group (two-year-old healthy M. amblycephala) was injected with 0.1 mL of sterilized PBS (pH 7.4). Six and twelve hours post-injection (hpi), each group was anesthetized with 100 mg/L MS-222 and dissected. Liver, spleen, kidney, gut and blood samples were then collected from each fish at 6 and 12 h after injection and control group. All samples collected were immediately frozen in liquid nitrogen and stored at −80 °C for RNA extraction and subsequent qPCR.
2.8. Statistical Analyses
All quantitative data are presented as the means of three individual experiments ± standard errors (SE). Statistical analyses were performed using SPSS 19.0 software (IBM Analytics, Richmond, VA, USA). Significant differences among samples were determined by the one-way analysis of variance (ANOVA) using least significant difference (LSD)'s multiple range test. p < 0.05 and p < 0.01 were considered as statistically significantly and highly significant, respectively. After analyzing the data, Origin9.0 mapping software (OriginLab, Northampton, MA, USA) was used to produce the expression map.
3. Results
3.1. Protein Alignment
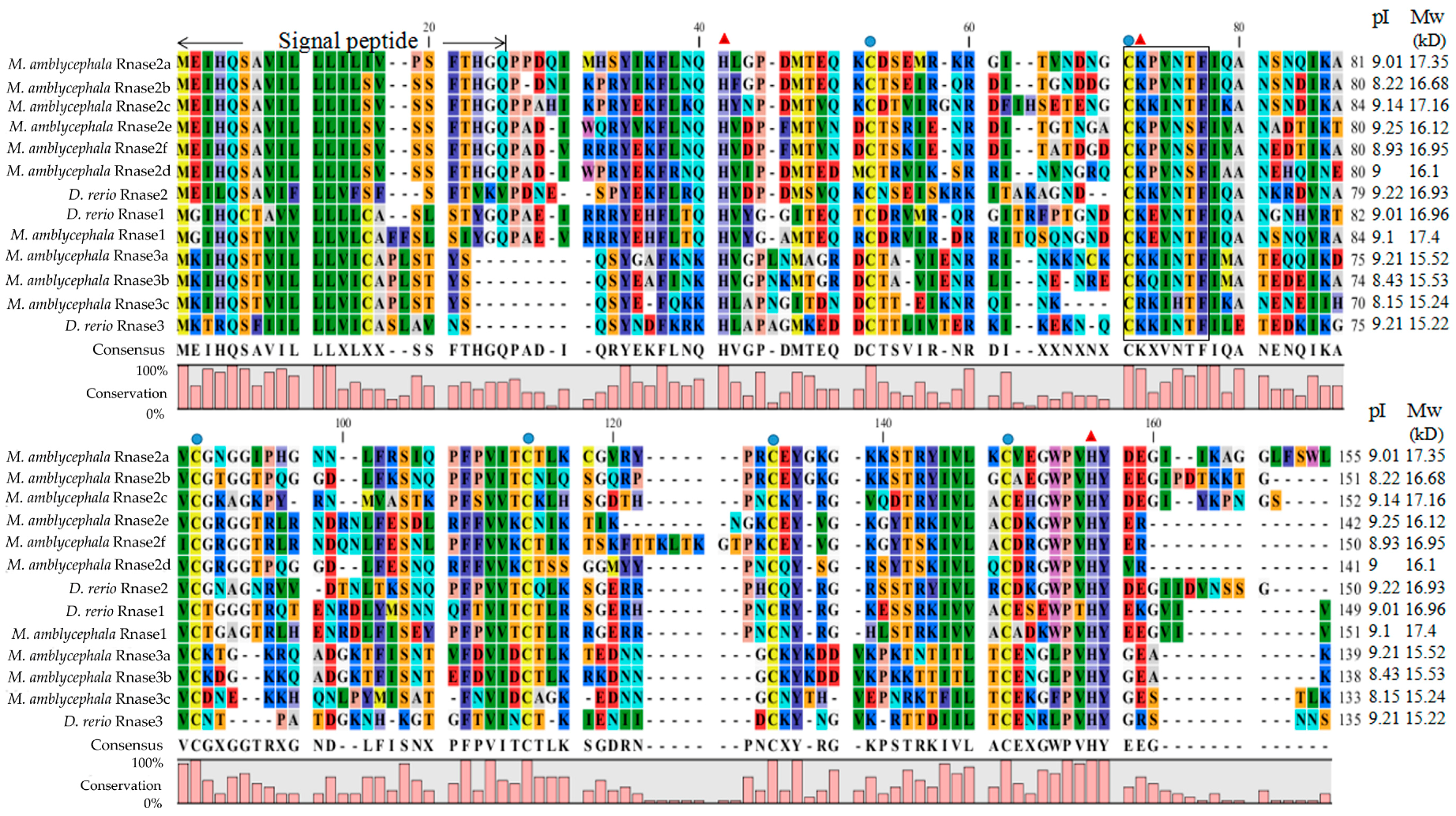
The alignment of M. amblycephala and D. rerio Rnase superfamily members revealed that all proteins possess a structure common to the RnaseA superfamily, including a specific signal peptide of more than 20 amino acids at the N-terminal, coding sequences encoded on a single exon, the conserved “CKXXNTF” signature motif, two histidine residues and a lysine residue constituting a conserved catalytic triad, and six cysteines forming disulfide bonds (Figure 1). The calculated putative pI and Mw of the mature peptides of all nine genes in M. amblycephala and D. rerio were ranging from 8.15 to 9.25 and 15.22 to 17.39 kD, respectively (Figure 1). Moreover, we calculated the number of lysines and arginines in Rnase1 to 3 of fishes, humans and cattle (Table S1).
3.2. Structural Predictions
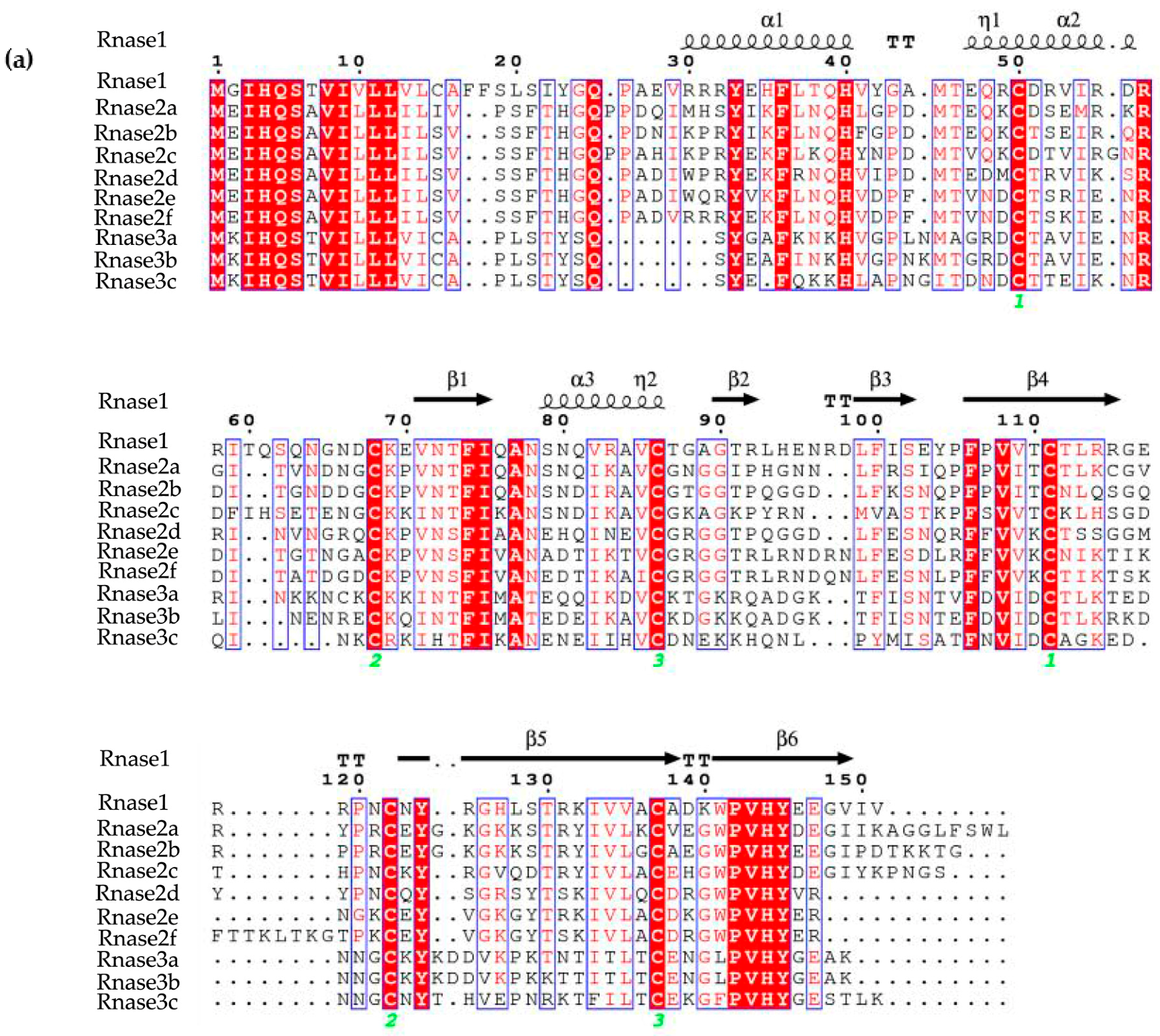
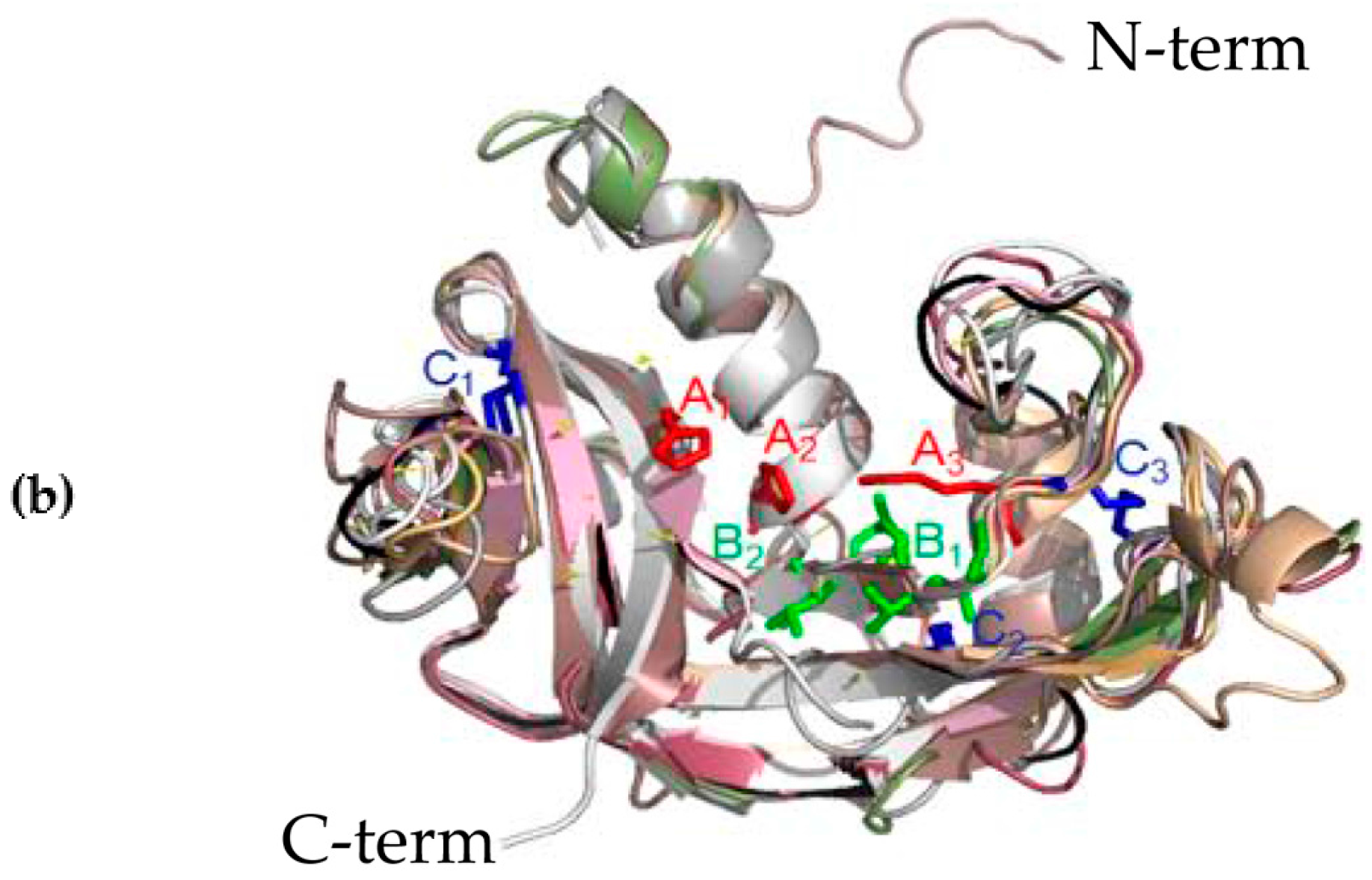
The three-dimensional (3D) structures of M. amblycephala Rnase1-3 proteins were predicted by homology modeling. Their alignment shows a typical fold with three α-helices and six β-strands for members of the RnaseA superfamily (Figure 2A). Moreover, all studied proteins show a high level of conservation and high homology. The high conservation is observed in the conserved positions of three catalytic residues (A1, A2 and A3 in Figure 2B), and pyrimidine (B1) and purine (B2) binding sites. More variability was seen in disulfide bond binding sites (C1, C2, C3). Three-dimensional structures of individual proteins are available from the Figure S2.
3.3. Phylogenetic Analysis
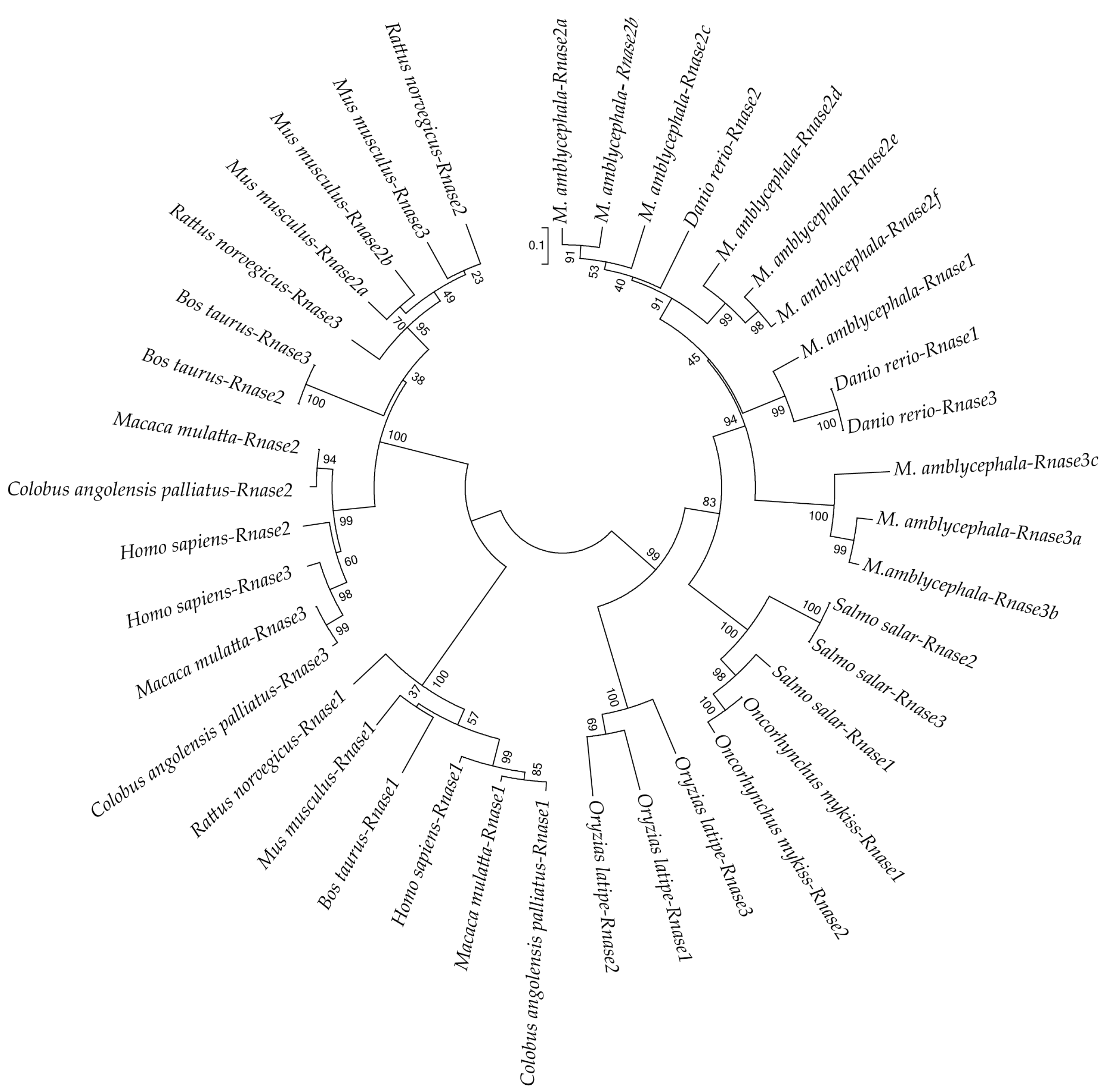
To better understand the evolutionary relationships of Rnase1 to 3 in vertebrates, we reconstructed a protein sequence-based NJ phylogenetic tree of homologs in Teleosts and mammals. The two groups (fish and mammals) are divided into two distinct clades (Figure 3). The mammalian clade is further clearly divided into Rnase1 and Rnase2+Rnase3 branches. In the fish clade, Ma-Rnase1 and Ma-Rnase2 protein sequences are more similar to the corresponding orthologs belonging to D. rerio than to those of O. mykiss, O. latipes or S. salar. Rnase2 and Rnase3 of many species clustered together, including M. amblycephala, S. salar, B. taurus and M. musculus, which is a consequence of gene duplication. M. amblycephala has multiple copies of Rnase2 and Rnase3, both of which formed their respective clusters. Because the sequences of Rnase1 to 3 are short and quite divergent, the bootstrap values on the phylogenetic tree are not high at some nodes.
3.4. Rnase2 and Rnase3 Messenger RNA Expression Patterns
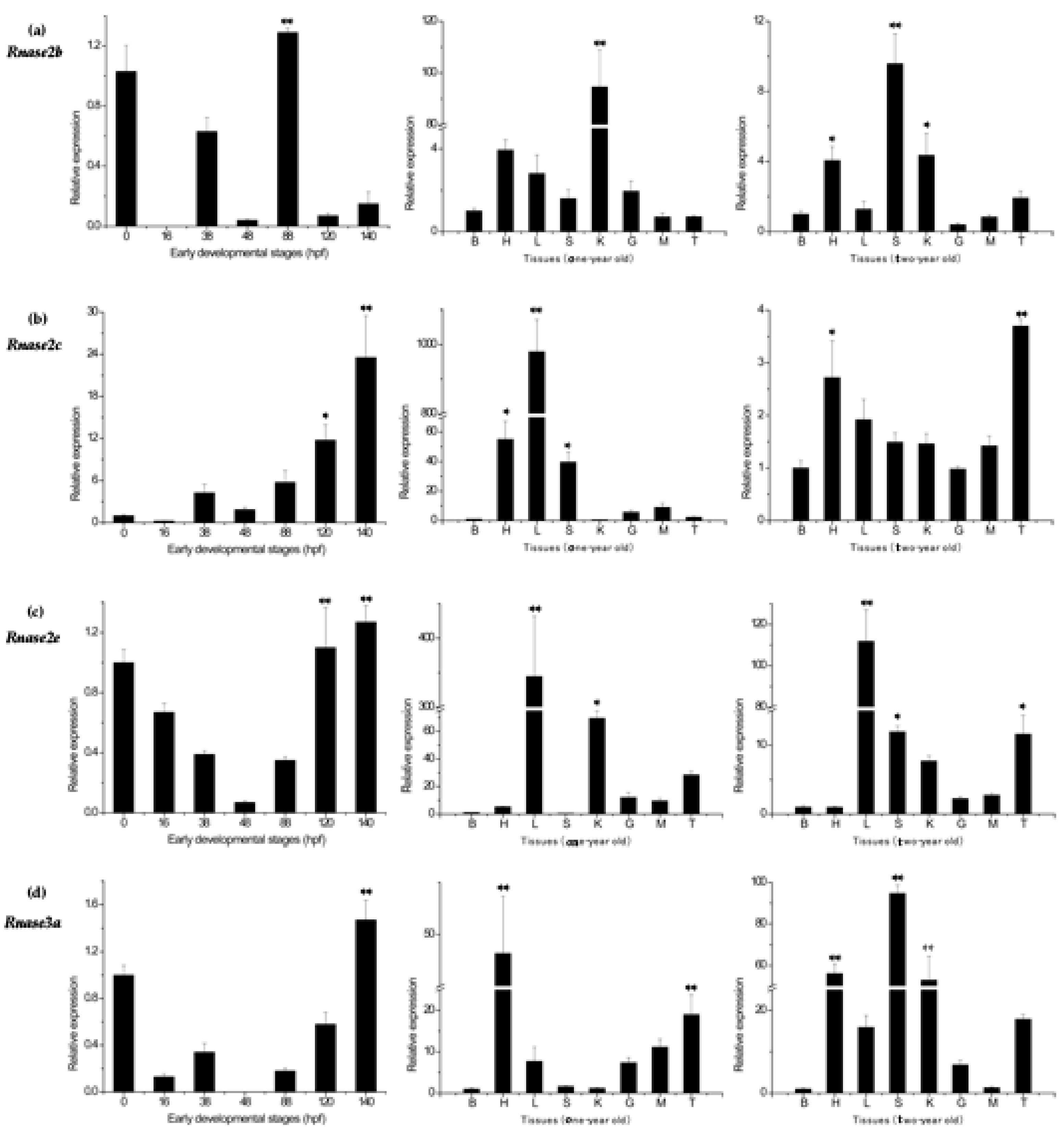
Expression of only four genes (Rnase2b, Rnase2c, Rnase2e and Rnase3a) could be detected in the eight different tissues of healthy M. amblycephala and during the early developmental stages (0, 16, 38, 48, 88, 120 and 140 hpf). No expression was detected for Rnase2a, Rnase2d, Rnase2f, Rnase3b or Rnase3c. The expression levels in the fertilized egg stage (0 h) were used as the standard against which the relative expression at other stages of development was calculated. Ma-Rnase2b was strongly expressed at 88 hpf, but weakly expressed at 120 and 140 hpf, with no expression at all at 16 hpf. However, Ma-Rnase2c and Ma-Rnase2e were strongly expressed at 120 and 140 hpf. During the remaining stages, expression levels were lower than during the fertilized egg stage (Figure 4). As the lowest expression was detected in brain tissue, expression levels in other tissues were normalized to this tissue. In the studied tissues of one-year-old fish, the highest expression levels of Ma-Rnase2b were detected in kidney (>90×), significantly higher (p < 0.01) than in any other tissue. Ma-Rnase2c and Ma-Rnase2e were expressed most highly in liver (979×, 344×, respectively) and Ma-Rnase3a was strongly expressed in heart (48×) and testis (19×). However, the expression patterns of the four genes in the two-year-old fish were dramatically different from those in the tissues of one-year-old fish. In the studied tissues of two-year-old fish, significantly higher (p < 0.01) expression levels of Ma-Rnase2b were detected in spleen (>9×). Ma-Rnase2c was expressed most highly in testis (>3.5×), followed by heart (>2.5×) and liver (>2.5×). Ma-Rnase2e was expressed in liver (>110×), followed by spleen (>10×) and testis (10×). Ma-Rnase3a was significantly higher (p < 0.01) expressed in spleen (>90×), heart (>55×) and kidney (>50×). The Rnase3 showed an expression pattern different from Rnase2. The highest expression levels of four genes were detected in heart, spleen, testis, liver and kidney tissues. Detailed cycle threshold (Ct) values can be found in Table S2.
3.5. Expression Profiles of Rnase2 and Rnase3 Messenger RNA after Aeromonas hydrophila Challenge
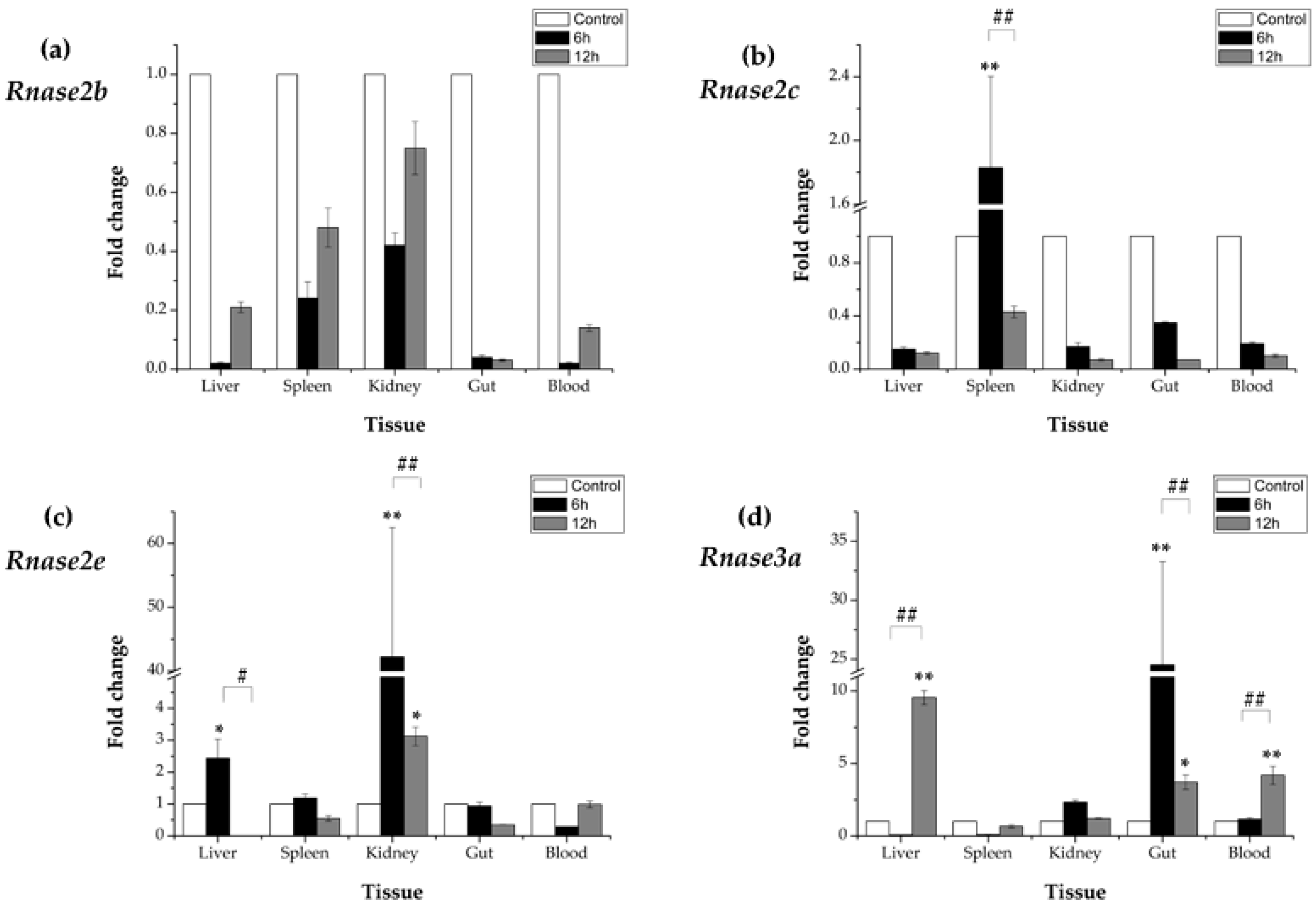
After an A. hydrophila immune-challenge, time-dependent relative (to the control group) expression patterns of Rnase2 and Rnase3 mRNA were observed in all five studied tissues (liver, spleen, kidney, gut and blood). The expression of Rnase2b showed down-regulated at 6 hpi (in comparison to the control) in liver (0.02×), spleen (0.24×), kidney (0.42×), gut (0.04×) and blood (0.02×) samples, and down-regulated at 12 hpi in liver (0.21×), spleen (0.48×), kidney (0.75×), gut (0.03×) and blood (0.14×) samples (Figure 5a), while the differences were not significant (p > 0.05). The expression of Rnase2c was only significantly (p < 0.01) up-regulated (1.83×) in spleen at 6 hpi compared to control, while down-regulated at 12hpi compared to 6hpi (p < 0.01). No significant differences (p > 0.05) were found in other tissues (Figure 5b). The expression of Rnase2e was significantly (p < 0.01) increased in kidney at 6hpi (42.27×) and 12 hpi (3.12×), followed by liver at 6hpi (2.44×) (p < 0.05), while no differences were found in spleen, gut and blood (p > 0.05) (Figure 5c). The expression of Rnase3a showed up-regulated at 6hpi in kidney (2.33×) and blood (1.16×), significantly up-regulated (p < 0.01) in gut (24.53×), but down-regulated in liver (0.07×) and spleen (0.09×) (p > 0.05) compared to control. However, at 12 hpi, the expression was significantly increased in liver (9.54×) (p < 0.01), blood (4.18×) (p < 0.01) and gut (3.71×) (p < 0.05) in comparison to control (Figure 5d). In addition, Rnase3a increased significantly in liver and blood, but decreased in gut at 12 hpi compared to 6 hpi (p < 0.01).
4. Discussion
RnaseA has been widely studied in the context of biological activities, molecular evolution, structural function and physiological diseases in mammals [43], amphibians [44], primates [45], and teleosts [7]. Many members of the RnaseA superfamily have antibacterial and antiviral activities. It has been reported that human Rnase 2, 3, 5 and 7 [46], chicken Rnase A-2 [47] and mouse Rnase4 [48] possess bactericidal activity. Rnase1-3 and 5 from zebrafish (D. rerio) and Rnase1-2 from Atlantic salmon (S. salar) have angiogenic and bactericidal activities [3,7]. Our previous study found that Rnase1 of M. amblycephala has digestive and antibacterial activity [31]. The present study reports Rnase2 (six) and Rnase3 (three) genes in the blunt snout bream genome, and their expression in healthy and bacterially challenged M. amblycephala. Only three copies of Rnase2 and one Rnase3 are expressed, but they have different patterns of expression. Rnase3 are more susceptible to A. hydrophila infection than Rnase2.
In this research, multiple-protein-sequence alignment of M. amblycephala and D. rerio homologs showed that protein sequences are mostly similar to each other, consistent in size (133–150 amino acids) and possessing the characteristics of the RnaseA superfamily. As opposed to most mammalian Rnases, which have eight conserved cysteines, non-mammalian Rnases, including fishes, have only six [6]. Catalytic residues of Rnase2 and Rnase3 are located in the first and second α-helices and sixth β-strands. The N-terminal domain (1–45) of human Rnase3 includes the main determinants for the protein’s antimicrobial activity [49], so we speculate that M. amblycephala Rnase3 antimicrobial activity may be related to the N-terminus, and the helical extension of the N- and C-termini may provide a flexible structure. It has been suggested and experimentally verified in several antibacterial Rnases that positively charged amino acid residues are important for the disruption of negatively charged bacterial cell membranes, and thus are crucial for the bactericidal activity [27]. Indeed, all known bactericidal Rnases, including mammalian Rnase3, 5, 6 and 7, as well as chicken Rnases, have relatively high isoelectric points [28]. As the isoelectric points of M. amblycephala of Rnase2 and 3 are relatively high (Figure 1), this suggests that these Rnases may also be bactericidal but requires further assessment. The Rnase1 gene has multiple copies in mammals, especially in foregut-fermenting monkeys and bovines, suggesting that the duplication of the Rnase1 gene was an adaptation to their digestive physiology [45,50]. However, there is no genetic duplication of Rnase1 in teleosts. The two groups (fish and mammals) are divided into two distinct clades in the NJ tree of Rnase2 and Rnase3 which is similar to Rnase1 (Figure 3) [31], In addition, the two genes (Rnase2 and Rnase3) are divided into two distinct clades in the NJ tree of mammals and fishes in our studies (Figure 3), which is consistent with studies reported [2,16]. Rnase2 of M. musculus has two homologous genes, suggesting that the duplication of the Rnase2 gene was an adaptation to their host defense [51]. Rnase2 and 3 of other animals don’t have genetic homology. In this study, we found that M. amblycephala eosinophils comprise nine genes (Rnase2a, Rnase2b, Rnase2c, Rnase2d, Rnase2e, Rnase2f, Rnase3a, Rnase3b and Rnase3c), but D. rerio and S. salar possess only one copy of each. There are also many gene duplications of Rnase1, Rnase4, Rnase5 and Rnase6 in Chiroptera [52]. It shows that they have unique evolutionary patterns and functional differentiation. Expression profiles of genes can also be strongly influenced by gene duplications. qPCR was used to further explore the expression patterns of these nine genes.
Our qPCR analysis showed that Ma-Rnase2 and 3 genes have differential expression patterns. Rnases2b, 2c, 2e and 3a were not expressed in the early stages of development, but are weakly expressed at 120 and 140 hpf, which is similar to D. rerio [6]. Rnase2b, Rnase2c and Rnase2e were highly expressed in the endocrine tissues (kidney, liver and spleen) and heart of one-year-old fish, and highly expressed in liver, spleen, kidney of two-year-old fish. In addition, Rnase3a was most strongly expressed in the heart and testis of one-year-old fish, whereas in two-year-old fish it was highly expressed in all studied tissues, except for the brain and muscle. This expression pattern is different from the one reported for Rnase2 and Rnase3 orthologs in adult D. rerio: Rnase3 is highly expressed in heart and gut, but Rnase2 weakly expressed in heart, liver and gut. However, neither of the two orthologs was expressed in the brain tissue of either of the two species (D. rerio and M. amblycephala). Rnase2 of D. rerio has a weak digestive activity and antimicrobial activity, whereas Rnase3 has high antibacterial activity against both gram-positive and gram-negative bacteria. Compared with D. rerio, Rnase3 of M. amblycephala has higher expression and stronger antibacterial activity. Rnase2 in adult S. salar is mainly expressed in the thymus, but no expression was detected in other tissues; functionally, Ss-Rnase2 has strong antibacterial and angiogenic activity [7]. Rnase2 and Rnase3 expression patterns in tissues may reflect the variability in their functions in different fish species, as well as in different individual animals. High Rnase2 and Rnase3 expression in different tissues might indicate multiple functions. As mentioned in our study, high expression in the thymus indicates that a gene may be involved in immunity-related functions; in the pancreas, it may possess digestion activity as an RNA-degrading enzyme; in the gut, it may be associated with intestinal microbiota [29]. Indeed, the existence of multiple copies of Rnase2 and Rnase3 may make their biological activity more powerful and promote the evolution of new protein functions [53]. Only three copies of Rnase2 and one Rnase3 are expressed. The specific expression profiles of different Ma-Rnase2 and 3 paralogs indicate multiple functions, such as RnaseA activity, antibacterial and antiviral activity, etc. Therefore, we conducted a bacterial infection experiment to further investigate the antibacterial activity of Rnase2 and Rnase3.
Rnase2b and Rnase2c mRNA expression after the A. hydrophila challenge was induced in all five M. amblycephala tissues at 12 hpi. Rnase2e is significantly (p < 0.01) up-regulated in kidney at 12 hpi after A. hydrophila infection, which could reflect the fact that kidney, as a major immune organ in fish, is more susceptible to A. hydrophila infection than liver and spleen. Rnase3a was up-regulated in liver, gut and blood 12 hpi after A. hydrophila infection. Much higher up-regulated in more tissues observed for Rnase3 in comparison to Rnase2 (after A. hydrophila infection) could be an indication that Rnase3 has a stronger antibacterial activity than Rnase2. This would be consistent with their corresponding functions in other animals: Rnase3 has a predominantly antimicrobial activity in zebrafish and humans [45], whereas Rnase2 has a predominantly antiviral activity in humans [54]. Previous studies have shown that RNase2 plays an important role in the activation of dendrocytes, immune response modulation, and TLR2 activation [55]. A novel antibacterial activity emerged in Rnase3 after its duplication [28]. Rnase3 is an antimicrobial protein secreted in response to infection, and is critical for neutralizing bacterial lipopolysaccharides (LPS) and inhibiting tumor necrosis factor production in human macrophages [56].
5. Conclusions
In conclusion, we have characterized the structure, expression and response to a bacterial challenge of Ma-Rnase2 and Ma-Rnase3 from the molecular aspect. This study demonstrates preliminarily that, although Rnase2 and Rnase3 are very similar, they might remarkably differ in their biological activities. Rnase3 is more susceptible to A. hydrophila infection than Rnase2. In following studies, we will focus on the protein expression and biological function of Ma-Rnase2 and Ma-Rnase3; especially on the antibacterial mechanism of Rnase3 and experimental studies of bacterial infection both in vitro and in vivo. This will provide a more clear and comprehensive picture of functions and evolutionary history of Rnase2 and Rnase3 in M. amblycephala.
Supplementary Materials
The following are available online at https://www.mdpi.com/2073-4425/9/2/95/s1. Figure S1: Nucleotide sequences of Megalobrama amblycephala Rnase2a, Rnase2b, Rnase2c, Rnase2d, Rnase2e, Rnase2f, Rnase3a, Rnase3b and Rnase3c, Figure S2: The 3D protein structure prediction for M. amblycephala different Rnases, Table S1: Number of basic amino acids of Rnase1 to Rnase3 in fish Rnases, as well as Homo sapiens and Bos taurus Rnases known to be bactericidal, Table S2: The Ct values of Rnase2a, Rnase2d, Rnase2f, Rnase3b, Rnase3c and β-actin at different stages of development and growth.
Acknowledgments
This work was financially supported by the Central Public-interest Scientific Institution Basal Research Fund, CAFS (2017HY-XKQ0205), the National Natural Science Foundation of China (31702315), the China Postdoctoral Science Foundation (2016M600600) and the Hubei Provincial Natural Science Foundation of China (2017CFB166).
Author Contributions
H.L. conceived and designed the experiments and modified the manuscript; R.G. performed the experiments, analyzed the data and wrote the manuscript; W.W. contributed reagents and modified the manuscript; all authors reviewed and approved the final manuscript.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Barnard, E.A. Biological function of pancreatic ribonuclease. Nature 1969, 221, 340–344. [Google Scholar] [CrossRef] [PubMed]
- Cho, S.; Beintema, J.J.; Zhang, J. The ribonuclease A superfamily of mammals and birds: Identifying new members and tracing evolutionary histories. Genomics 2005, 85, 208–220. [Google Scholar] [CrossRef] [PubMed]
- Cho, S.; Zhang, J. Zebrafish ribonucleases are bactericidal: Implications for the origin of the vertebrate RnaseA superfamily. Mol. Biol. Evol. 2007, 24, 1259–1268. [Google Scholar] [CrossRef] [PubMed]
- Kazakou, K.; Holloway, D.E.; Prior, S.H.; Subramanian, V.; Acharya, K.R. Ribonuclease A homologues of the zebrafish: Polymorphism, crystal structures of two representatives and their evolutionary implications. J. Mol. Biol. 2008, 380, 206–222. [Google Scholar] [CrossRef] [PubMed]
- Pizzo, E.; Buonanno, P.; Di, M.A.; Ponticelli, S.; De, F.S.; Quarto, N.; Cubellis, M.V.; D’Alessio, G. Ribonucleases and angiogenins from fish. J. Biol. Chem. 2006, 281, 27454–27460. [Google Scholar] [CrossRef] [PubMed]
- Pizzo, E.; Merlino, A.; Turano, M.; Russo, K.I.; Coscia, F.; Zanfardino, A.; Varcamonti, M.; Furia, A.; Giancola, C.; Mazzarella, L.; et al. A new Rnase sheds light on the Rnase/angiogenin subfamily from zebrafish. Biochem. J. 2011, 433, 345–355. [Google Scholar] [CrossRef] [PubMed]
- Pizzo, E.; Varcamonti, M.; Di, M.A.; Zanfardino, A.; Giancola, C.; D’Alessio, G. Ribonucleases with angiogenic and bactericidal activities from the Atlantic salmon. FFBS J. 2008, 275, 1283–1295. [Google Scholar] [CrossRef] [PubMed]
- Lang, D.; Zhang, Y.; Yu, L. Molecular evolution of the ribonuclease A superfamily. Hereditas 2014, 36, 165–192. [Google Scholar]
- Wheeler, T.T.; Maqbool, N.J.; Gupta, S.K. Mapping, phylogenetic and expression analysis of the Rnase (RnaseA) locus in cattle. J. Mol. Evol. 2012, 74, 237–248. [Google Scholar] [CrossRef] [PubMed]
- Tao, F.Y.; Wei, Z.; Ma, R.Y. Progress in bactericidal activity of ribonuclease A superfamily members. Chin. J. Antibiot. 2009, 34, 325–379. [Google Scholar]
- Xu, H.; Li, D. Research progress of human Ribonuclease A superfamily. Chem. Life 2012, 32, 174–179. [Google Scholar]
- Gupta, S.K.; Haigh, B.J.; Griffin, F.J.; Wheeler, T.T. The mammalian secreted Rnases: Mechanisms of action in host defence. Innate Immun. 2013, 19, 86–97. [Google Scholar] [CrossRef] [PubMed]
- Sorrentino, S. The eight human “canonical” ribonucleases: Molecular diversity, catalytic properties, and special biological actions of the enzyme proteins. FEBS Lett. 2010, 584, 2194–2200. [Google Scholar] [CrossRef] [PubMed]
- Attery, A.; Batra, J.K. Mouse eosinophil associated ribonucleases: Mechanism of cytotoxic, antibacterial and antiparasitic activities. Int. J. Biol. Macromol. 2017, 94, 445–450. [Google Scholar] [CrossRef] [PubMed]
- Acharya, K.R.; Ackerman, S.J. Eosinophil granule proteins: form and function. J. Biol. Chem. 2014, 289, 17406–17415. [Google Scholar] [CrossRef] [PubMed]
- Futami, J. Tissue-specific expression of pancreatic-type RNases and RNase inhibitor in humans. DNA Cell. Biol. 1997, 16, 413–419. [Google Scholar] [CrossRef] [PubMed]
- Rosenberg, H.F. Eosinophil-derived Neurotoxin/RNase 2: Connecting the past, the present and the future. Curr. Pharm. Biotechnol. 2008, 9, 135–140. [Google Scholar] [CrossRef] [PubMed]
- Bedoya, V.I.; Boasso, A.; Hardy, A.W.; Rybak, S.; Shearer, G.M.; Rugeles, M.T. Ribonucleases in HIV type 1 inhibition: effect of recombinant RNases on infection of primary T cells and immune activation-induced RNase gene and protein expression. Aids Res. Hum. Retrov. 2006, 22, 897–907. [Google Scholar] [CrossRef] [PubMed]
- Rosenberg, H.F.; Dyer, K.D.; Domachowske, J.B. Respiratory viruses and eosinophils: Exploring the connections. Antivir. Res. 2009, 83, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Koczera, P.; Martin, L.; Marx, G.; Tobias, S. The ribonuclease A superfamily in humans: Canonical Rnases as the buttress of innate immunity. Int. J. Mol. Sci. 2016, 17, 1278. [Google Scholar] [CrossRef] [PubMed]
- Gullberg, U.; Widegren, B.; Arnason, U.; Egesten, A.; Olsson, I. The cytotoxic eosinophil cationic protein (ECP) has ribonuclease activity. Biochem. Bioph. Res. Co. 1986, 139, 1239–1242. [Google Scholar] [CrossRef]
- Rosenberg, H.F.; Tenen, D.G.; Ackerman, S.J. Molecular cloning of the human eosinophil-derived neurotoxin: a member of the ribonuclease gene family. Proc. Natl. Acad. Sci. USA 1989, 86, 4460–4464. [Google Scholar] [CrossRef] [PubMed]
- Rosenberg, H.F.; Dyer, K.D. Eosinophil cationic protein and eosinophil-derived neurotoxin, evolution of novel function in a primate ribonuclease gene family. J. Biol. Chem. 1995, 270, 21539–21542. [Google Scholar] [CrossRef] [PubMed]
- Carreras, E.; Boix, E.; Rosenberg, H.F.; Cuchillo, C.M.; Nogués, M.V. Both aromatic and cationic residues contribute to the membrane-lytic and bactericidal activity of eosinophil cationic protein. Biochemistry 2003, 42, 6636–6644. [Google Scholar] [CrossRef] [PubMed]
- Singh, A.; Batra, J.K. Role of unique basic residues in cytotoxic, antibacterial and antiparasitic activities of human eosinophil cationic protein. Biol. Chem. 2011, 392, 337–346. [Google Scholar] [CrossRef] [PubMed]
- Pulido, D.; Torrent, M.; Andreu, D.; Nogués, M.V.; Boix, E. Two human host defense ribonucleases against mycobacteria, the eosinophil cationic protein (RNase3) and RNase7. Antimicrob. Agents. Chemother. 2013, 57, 3797–3805. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Rosenberg, H.F.; Nei, M. Positive Darwinian selection after gene duplication in primate ribonuclease genes. Proc. Natl. Acad. Sci. USA 1998, 95, 3708–3713. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J. Evolution by gene duplication: An update. Trends Ecol. Evol. 2003, 18, 292–298. [Google Scholar] [CrossRef]
- Liu, H.; Wang, W. Expression patterns and functional novelty of ribonuclease 1 in herbivorous Megalobrama amblycephala. Int. J. Mol. Sci. 2016, 17, 786. [Google Scholar] [CrossRef] [PubMed]
- Gao, Z.; Luo, W.; Liu, H.; Zeng, C.; Liu, X.; Yi, S.; Wang, W. Transcriptome analysis and SSR/SNP markers information of the blunt snout bream (Megalobrama amblycephala). PLoS ONE 2012, 7, e42637. [Google Scholar] [CrossRef] [PubMed]
- Liu, B.; Zhao, Z.; Brown, P.B.; Cui, H.; Xie, J.; Tsion, H.M.; Ge, X. Dietary vitamin A requirement of juvenile Wuchang bream (Megalobrama amblycephala) determined by growth and disease resistance. Aquaculture 2016, 450, 23–30. [Google Scholar] [CrossRef]
- Liu, H.; Chen, C.; Gao, Z.; Min, J.; Gu, Y.; Jian, J.; Jiang, X.; Cai, H.; Ebersberger, I.; Xu, M.; et al. The draft genome of blunt snout bream (Megalobrama amblycephala) reveals the development of intermuscular bone and adaptation to herbivorous diet. Gigascience 2017, 6, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Artimo, P.; Jonnalagedda, M.; Arnold, K.; Baratin, D.; Csardi, G.; Castro, E.; Duvaud, S.; Flegel, V.; Fortier, A.; Gasteiger, E.; et al. ExPASy: SIB bioinformatics resource portal. Nucleic Acids Res. 2012, 40, 597–603. [Google Scholar] [CrossRef] [PubMed]
- SignalP4.1 Server. Available online: http://www.cbs.dtu.dk/services/SignalP/ (accessed on 10 August 2017).
- Biasini, M.; Bienert, S.; Waterhouse, A.; Arnold, K.; Studer, G.; Schmidt, T.; Kiefer, F.; Gallo Cassarino, T.; Bertoni, M.; Bordoli, L.; et al. SWISS-MODEL: modelling protein tertiary and quaternary structure using evolutionary information. Nucleic. Acids Res. 2014, 42, W252. [Google Scholar] [CrossRef] [PubMed]
- Gouet, P.; Robert, X.; Courcelle, E. ESPript/ENDscript: Extracting and rendering sequence and 3D information from atomic structures of proteins. Nucleic. Acids. Res. 2003, 31, 3320–3323. [Google Scholar] [CrossRef] [PubMed]
- De Lano, W.L. The Py MOL Molecular Graphics System; De Lano Scientific: San Carlos, CA, USA, 2002. [Google Scholar]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis Version 7.0 for Bigger Datasets. Mol Biol Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [PubMed]
- GenBank. Available online: https://www.ncbi.nlm.nih.gov/ (accessed on 10 August 2017).
- Saitou, N.; Nei, M. The neighbor-joining method: A new method for reconstructing phylogenetic trees. Mol. Biol. Evol. 1987, 4, 406–425. [Google Scholar] [PubMed]
- Li, Z.; Yang, L.; Wang, J.; Shi, W.; Pawar, R.A.; Liu, Y.; Xu, C.; Cong, W.; Hu, Q.; Lu, T.; et al. β-Actin is a useful internal control for tissue-specific gene expression studies using quantitative real-time PCR in the half-smooth tongue sole Cynoglossus semilaevis challenged with LPS or Vibrio anguillarum. Fish Shellfish Immun. 2010, 29, 89–93. [Google Scholar] [CrossRef] [PubMed]
- Schmittgen, T.D.; Livak, K.J. Analyzing real-time PCR data by the comparative CT method. Nat. Protoc. 2008, 3, 1101–1108. [Google Scholar] [CrossRef] [PubMed]
- Giusti, D.; Gatouillat, G.; Jan, S.L.; Plée, J.; Bernard, P.; Antonicelli, F.; Pham, B.N. Eosinophil Cationic Protein (ECP), a predictive marker of bullous pemphigoid severity and outcome. Sci. Rep. 2017, 7, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Arnold, U. Stability and folding of amphibian ribonuclease A superfamily members in comparison with mammalian homologues. FEBS J. 2014, 281, 3559–3575. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Rosenberg, H.F. Complementary advantageous substitutions in the evolution of an antiviral RNase of higher primates. Proc. Natl. Acad. Sci. USA 2002, 99, 5486–5491. [Google Scholar] [CrossRef] [PubMed]
- Harder, J.; Schröder, J.M. RNase7, a novel innate immune defense antimicrobial protein of healthy human skin. J. Biol. Chem. 2002, 277, 46779–46784. [Google Scholar] [CrossRef] [PubMed]
- Nitto, T.; Dyer, K.D.; Czapiga, M.; Rosenberg, H.F. Evolution and function of leukocyte RNaseA ribonucleases of the avian species, Gallus gallus. J. Biol. Chem. 2006, 281, 25622–25634. [Google Scholar] [CrossRef] [PubMed]
- Hooper, L.V.; Stappenbeck, T.S.; Hong, C.V.; Gordon, J.I. Angiogenins: A new class of microbicidal proteins involved in innate immunity. Nat. Immun. 2003, 4, 269–273. [Google Scholar] [CrossRef] [PubMed]
- Boix, E.; Salazar, V.A.; Torrent, M.; Pulido, D.; Nogués, M.V.; Moussaoui, M. Structural determinants of the eosinophil cationic protein antimicrobial activity. Biol. Chem. 2012, 393, 801–815. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J. Parallel adaptive origins of digestive RNases in Asian and African leaf monkeys. Nat. Genet. 2006, 38, 819–823. [Google Scholar] [CrossRef] [PubMed]
- Rosenberg, H.F. Eosinophil-derived neurotoxin (EDN/RNase2) and the mouse eosinophil-associated RNases (mEars): Expanding roles in promoting host defense. Int. J. Mol. Sci. 2015, 16, 15442–15455. [Google Scholar] [CrossRef] [PubMed]
- Goo, S.M.; Cho, S. The expansion and functional diversification of the mammalian ribonuclease a superfamily epitomizes the efficiency of multigene families at generating biological novelty. Genome Biol. Evol. 2013, 5, 2124. [Google Scholar] [CrossRef] [PubMed]
- Lynch, M.; Conery, J.S. The evolutionary fate and consequences of duplicate genes. Science 2000, 290, 1151–1155. [Google Scholar] [CrossRef] [PubMed]
- Rugeles, M.T.; Trubey, C.M.; Bedoya, V.I.; Pinto, L.A.; Oppenheim, J.J.; Rybak, S.M.; Shearer, G.M. Ribonuclease is partly responsible for the HIV-1 inhibitory effect activated by HLA alloantigen recognition. Aids. 2003, 17, 481–486. [Google Scholar] [CrossRef] [PubMed]
- Gagne, D.; Narayanan, C.; Bafna, K.; Charest, L.A.; Agarwal, P.K.; Doucet, N. Sequence-specific backbone resonance assignments and microsecond timescale molecular dynamics simulation of human eosinophil-derived neurotoxin. Biomol. NMR Assignm. 2017, 11, 143–149. [Google Scholar] [CrossRef] [PubMed]
- Pulido, D.; Garcia-Mayoral, M.F.; Moussaoui, M.; Velázquez, D.; Torrent, M.; Bruix, M.; Boix, E. Structural basis for endotoxin neutralization by the Eosinophil Cationic Protein. FEBS J. 2016, 283, 4176–4191. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Multiple protein sequence alignments of Rnase1-3 in Megalobrama amblycephala and Danio rerio. Dashes indicate alignment gaps. The locations of the signal peptides are marked between the first two arrows. The isoelectric points and molecular weights (kDa) of the mature peptides are indicated by pI and Mw, respectively. The conserved “CKXXNTF” motif is boxed. The positions of six structural cysteines (active-site residues) are marked with blue circles. The three red triangles show the three catalytic residues.
Figure 1.
Multiple protein sequence alignments of Rnase1-3 in Megalobrama amblycephala and Danio rerio. Dashes indicate alignment gaps. The locations of the signal peptides are marked between the first two arrows. The isoelectric points and molecular weights (kDa) of the mature peptides are indicated by pI and Mw, respectively. The conserved “CKXXNTF” motif is boxed. The positions of six structural cysteines (active-site residues) are marked with blue circles. The three red triangles show the three catalytic residues.
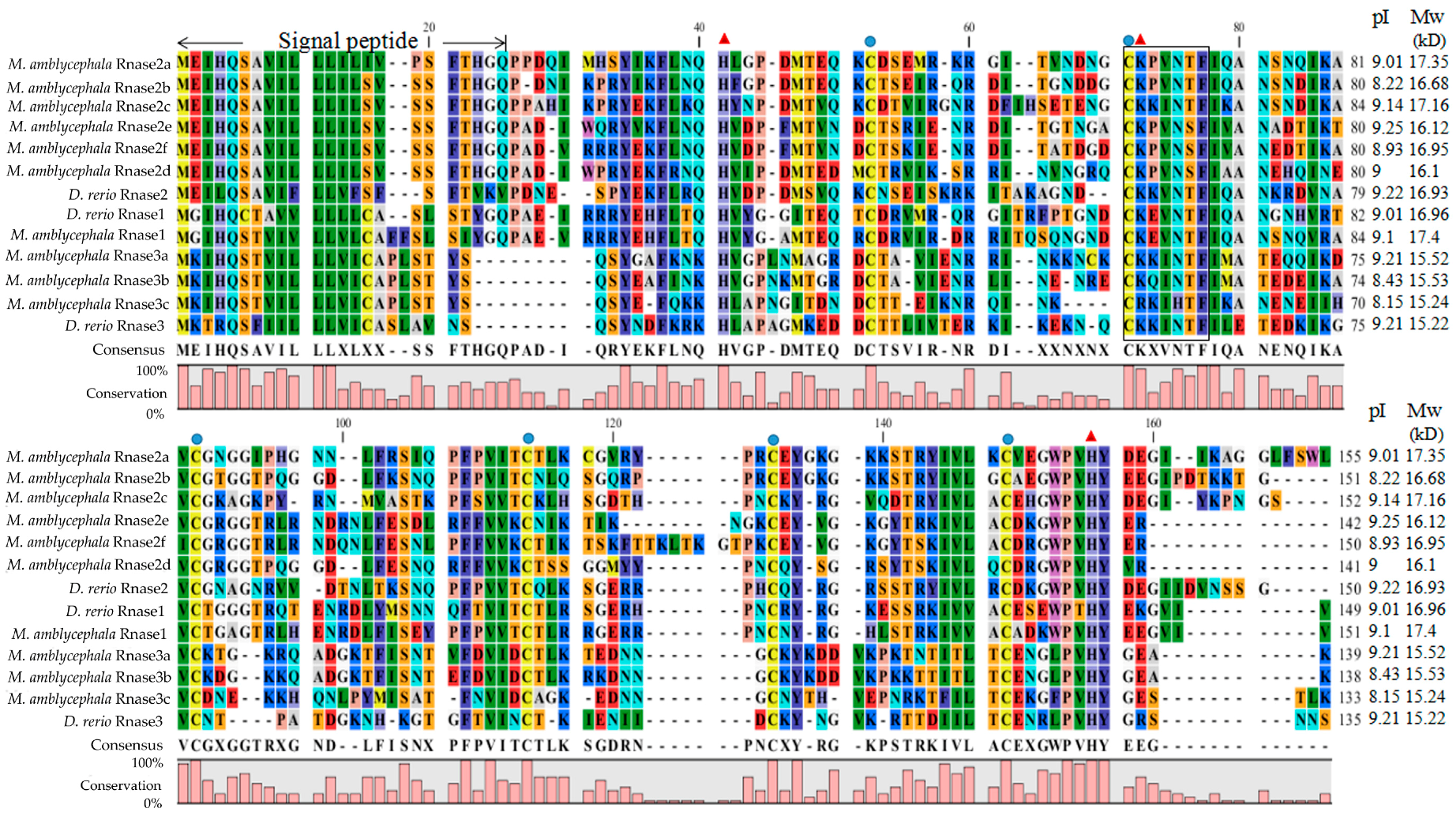
Figure 2.
(a) Sequences alignment of M. amblycephala Rnase1, Rnase2a, Rnase2b, Rnase2c, Rnase2d, Rnase2e, Rnase2f, Rnase3a, Rnase3b and Rnase3c. Secondary structure elements of Rnases are depicted at the top. Cysteine pairings (disulfide bridges) are numbered in green below the columns. Locations of three α-helices (α1-α3) and six β-strands (β1-β6) are shown above the sequences. The figure was created using ESPript software. (b) Representation of the superimposed three-dimensional structures of all ten Rnases of M. amblycephala. The image was drawn using PYMOL. Catalytic residue regions are shown in red (A1-A3), conserved binding sites in green (B1, B2), and disulfide bond binding sites in blue (C1-C3).
Figure 2.
(a) Sequences alignment of M. amblycephala Rnase1, Rnase2a, Rnase2b, Rnase2c, Rnase2d, Rnase2e, Rnase2f, Rnase3a, Rnase3b and Rnase3c. Secondary structure elements of Rnases are depicted at the top. Cysteine pairings (disulfide bridges) are numbered in green below the columns. Locations of three α-helices (α1-α3) and six β-strands (β1-β6) are shown above the sequences. The figure was created using ESPript software. (b) Representation of the superimposed three-dimensional structures of all ten Rnases of M. amblycephala. The image was drawn using PYMOL. Catalytic residue regions are shown in red (A1-A3), conserved binding sites in green (B1, B2), and disulfide bond binding sites in blue (C1-C3).
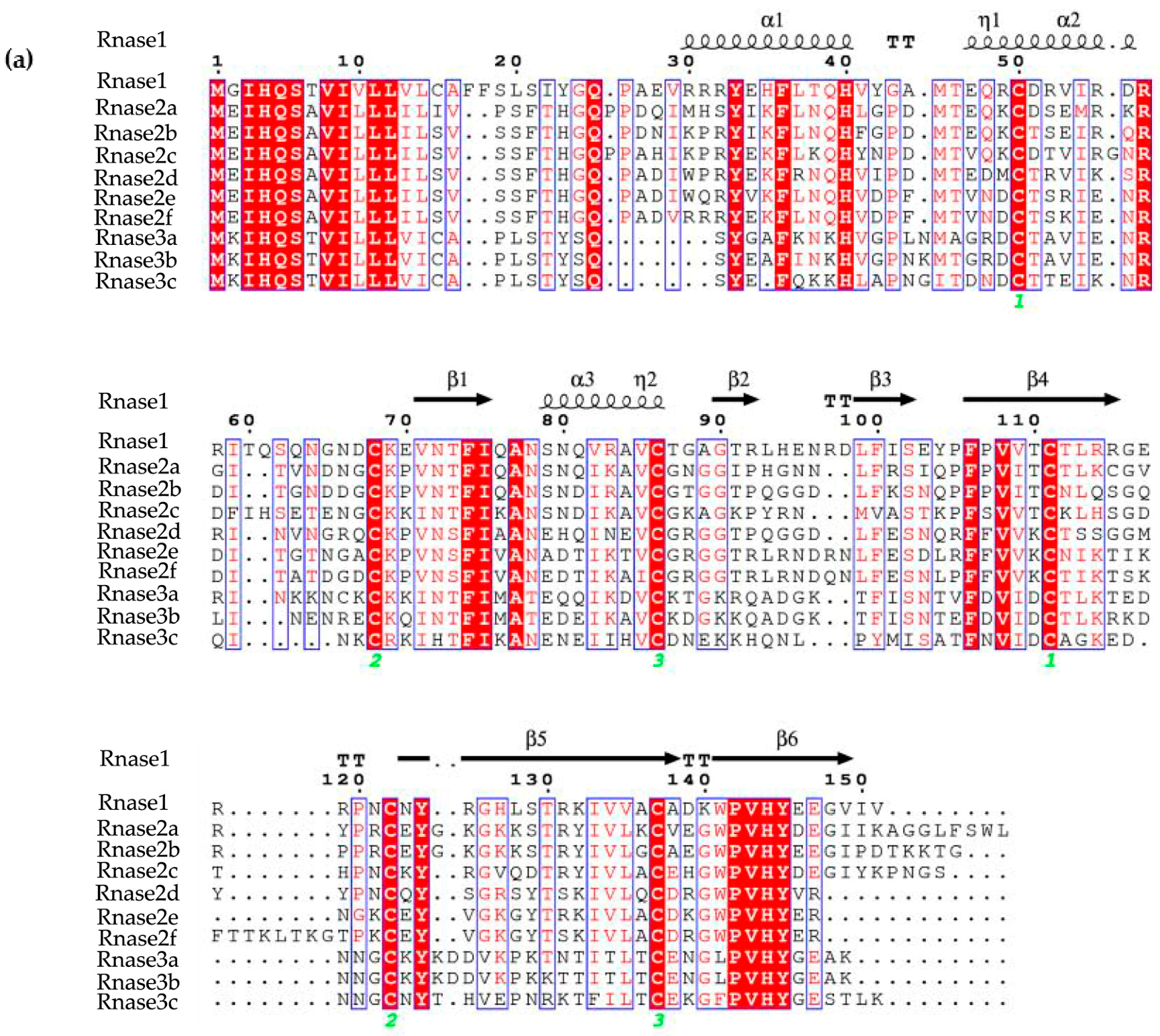
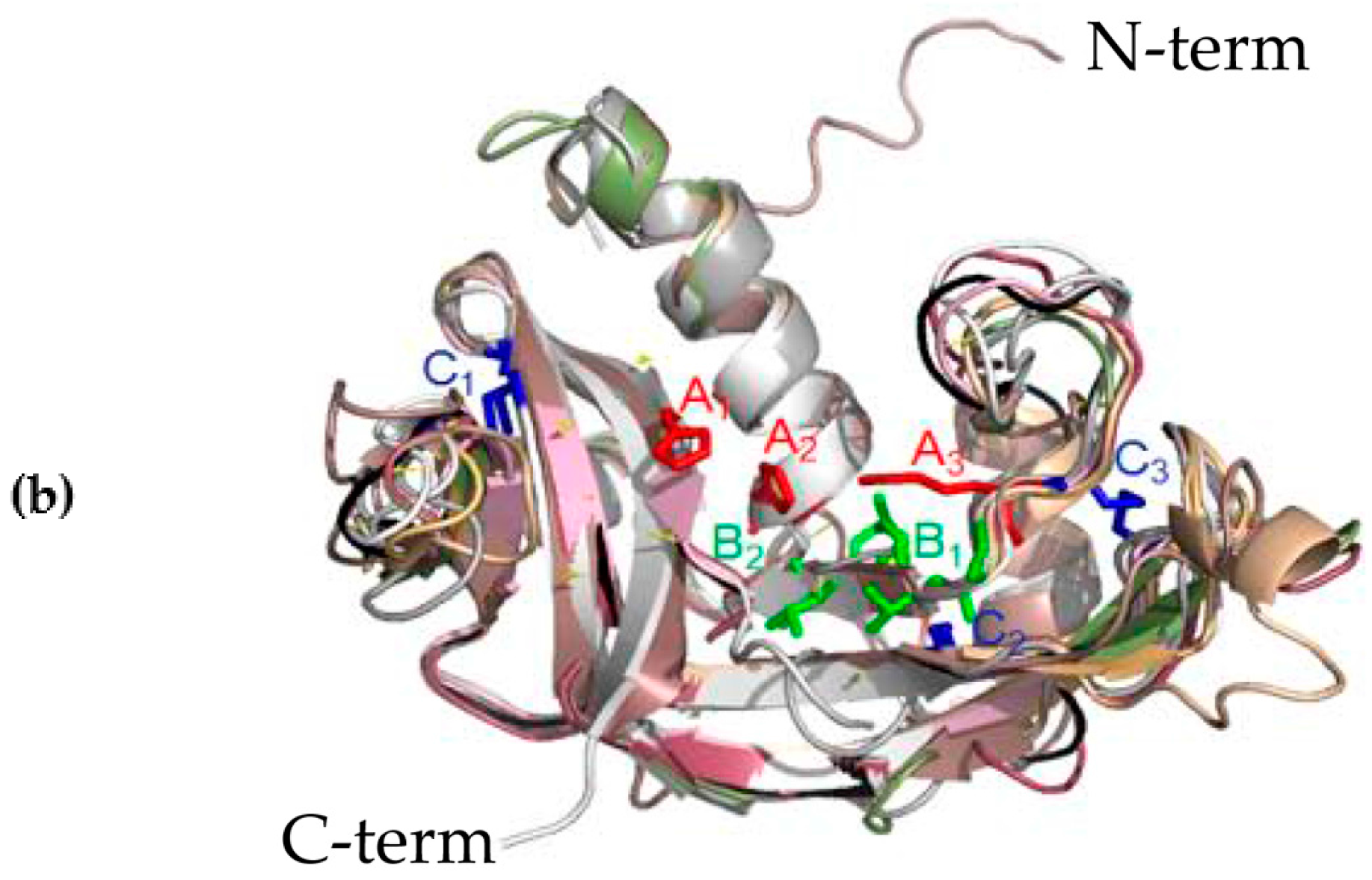
Figure 3.
Neighbor-Joining (NJ) phylogenetic tree showing the relationships among protein sequences of Rnase1, 2 and 3 of various bony fishes and mammals. Numbers at the branches indicate bootstrap probabilities inferred with 1000 replicates.
Figure 3.
Neighbor-Joining (NJ) phylogenetic tree showing the relationships among protein sequences of Rnase1, 2 and 3 of various bony fishes and mammals. Numbers at the branches indicate bootstrap probabilities inferred with 1000 replicates.
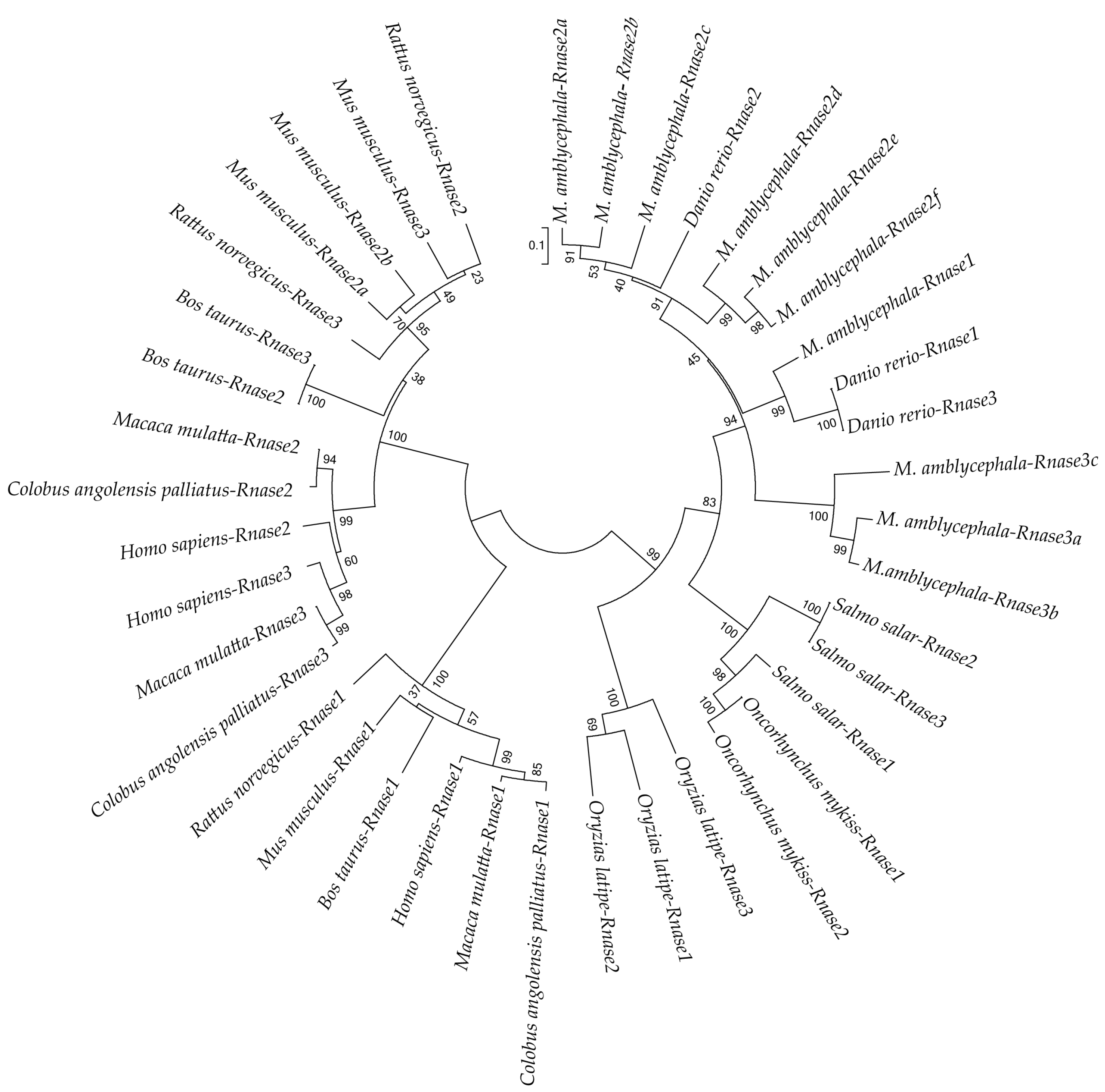
Figure 4.
Expression patterns of (a) Rnase2b, (b) Rnase2c, (c) Rnase2e and (d) Rnase3a genes in different developmental stages and tissues of M. amblycephala. β-actin was used as the reference gene. Tissues: brain (B), heart (H), liver (L), spleen (S), kidney (K), gut (G), muscle (M), and testis (T). Developmental stages: 0 h post-fertilization (hpf) (fertilized egg), 16 hpf (late gastrulastage), 38 hpf (heart appearance), 48 hpf (hatching), 88 hpf (eye appearance), 120 hpf (air bladder formation) and 140 hpf (intestine appearance). The expressions in different developmental stages were normalized to the fertilized egg stage, while expressions in tissues were normalized to the expressions in brain tissue (both set as 1, or control group). Differences were statistically analyzed using one-way analysis of variance (one-way analysis of variance (ANOVA)). Highly significant differences from the control group (p < 0.01) are marked with **, and significant differences (p < 0.05) with *.
Figure 4.
Expression patterns of (a) Rnase2b, (b) Rnase2c, (c) Rnase2e and (d) Rnase3a genes in different developmental stages and tissues of M. amblycephala. β-actin was used as the reference gene. Tissues: brain (B), heart (H), liver (L), spleen (S), kidney (K), gut (G), muscle (M), and testis (T). Developmental stages: 0 h post-fertilization (hpf) (fertilized egg), 16 hpf (late gastrulastage), 38 hpf (heart appearance), 48 hpf (hatching), 88 hpf (eye appearance), 120 hpf (air bladder formation) and 140 hpf (intestine appearance). The expressions in different developmental stages were normalized to the fertilized egg stage, while expressions in tissues were normalized to the expressions in brain tissue (both set as 1, or control group). Differences were statistically analyzed using one-way analysis of variance (one-way analysis of variance (ANOVA)). Highly significant differences from the control group (p < 0.01) are marked with **, and significant differences (p < 0.05) with *.
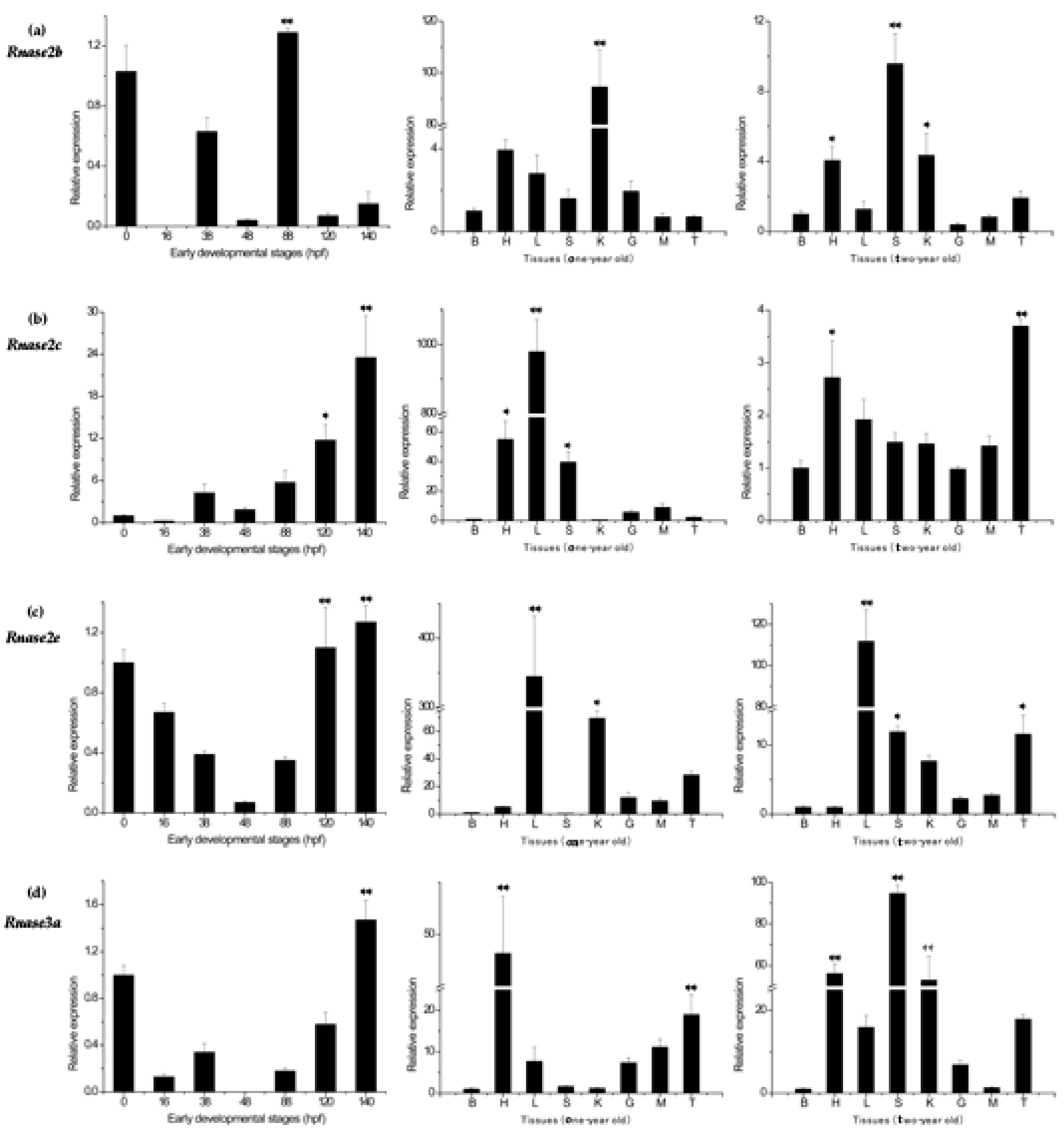
Figure 5.
Messenger RNA (mRNA) after Aeromonas hydrophila infection in five different tissues (a) Expression of Rnase2b, (b) Rnase2c, (c) Rnase2e and (d) Rnase3a. β-actin was used as the internal reference. Results are presented as the mean ± standard error among three replicates, where significant differences (p < 0.05) between control and test groups are marked with *, and highly significant differences (p < 0.01) with **. # p < 0.05 and ## p < 0.01 show significant differences between the designated groups. 6h: 6 h after A. hydrophila infection; 12h: 12 h after A. hydrophila infection.
Figure 5.
Messenger RNA (mRNA) after Aeromonas hydrophila infection in five different tissues (a) Expression of Rnase2b, (b) Rnase2c, (c) Rnase2e and (d) Rnase3a. β-actin was used as the internal reference. Results are presented as the mean ± standard error among three replicates, where significant differences (p < 0.05) between control and test groups are marked with *, and highly significant differences (p < 0.01) with **. # p < 0.05 and ## p < 0.01 show significant differences between the designated groups. 6h: 6 h after A. hydrophila infection; 12h: 12 h after A. hydrophila infection.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Primers for Megalobrama amblycephala Rnase2 and Rnase3 expression analysis.
| Primer Name | Primer Sequence (5′-3′) | Fragment Length |
|---|---|---|
| β-actin | F:ACCCACACCGTGCCCATCTA R:CGGACAATTTCTCTTTCGGCTG | 204 |
| Rnase2a | F:GTCAACCACCAGACCAAAT R:AAGGCTGAATGCTCCTAAAC | 237 |
| Rnase2b | F:GATGGCTGCAAACCTGTCA R:CGAGTGGACTTCTTCCCTTT | 194 |
| Rnase2c | F:GATATGACCGTGCAGAAGTG R:TCTGTATTTACAGTTTGGGTGT | 240 |
| Rnase2d | F:CCTTCACTCACGGACAACC R:GCACTGACGACCATTTACAT | 140 |
| Rnase2e | F:AGAGGAGGAACTCGACTGAG R:AGCCTTTATCACAAGCCAAC | 157 |
| Rnase2f | F:AAGCAATTTGTGGCAGAG R:GGAGTTCCCTTAGTTAGTTTAG | 133 |
| Rnase3a | F:AGGCAAGCGGATGGAAAG R:CACCATAATGTACTGGGAGACC | 166 |
| Rnase3b | F:AATTAAAGCCGTTTGTAAGG R:ACCATAATGTACTGGGAGACC | 193 |
| Rnase3c | F:TTGTCCACTTACAGCCAGAG R:AACCGTTGTTATCTTCTTTGC | 256 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Geng, R.; Liu, H.; Wang, W. Differential Expression of Six Rnase2 and Three Rnase3 Paralogs Identified in Blunt Snout Bream in Response to Aeromonas hydrophila Infection. Genes 2018, 9, 95. https://doi.org/10.3390/genes9020095
AMA Style
Geng R, Liu H, Wang W. Differential Expression of Six Rnase2 and Three Rnase3 Paralogs Identified in Blunt Snout Bream in Response to Aeromonas hydrophila Infection. Genes. 2018; 9(2):95. https://doi.org/10.3390/genes9020095
Chicago/Turabian StyleGeng, Ruijing, Han Liu, and Weimin Wang. 2018. "Differential Expression of Six Rnase2 and Three Rnase3 Paralogs Identified in Blunt Snout Bream in Response to Aeromonas hydrophila Infection" Genes 9, no. 2: 95. https://doi.org/10.3390/genes9020095
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.