Genome-Wide Identification and Expression Profiling Analysis of the Xyloglucan Endotransglucosylase/Hydrolase Gene Family in Tobacco (Nicotiana tabacum L.)

Abstract
:1. Introduction
2. Materials and Methods
2.1. Identification of Xyloglucan Endo-Transglucosylase/Hydrolase Family Members in Tobacco and Other Species
2.2. Phylogenetic Analysis
2.3. Gene Structure Analysis and Weblogo of Conserved Catalytic Site
2.4. Chromosomal Location and Gene Duplication
2.5. Ka/Ks Values Estimation
2.6. Microarray Expression Profiles of NtXTHs
2.7. Plant Materials, Growth Conditions, and Stress Treatments
2.8. RNA Extraction and qRT-PCR Analysis
2.9. Amplification and Sequencing of NtXTH cDNAs
2.10. Subcellular Localization of NtXTH Proteins
3. Results
3.1. Identification of NtXTHs Based on the SGN Database
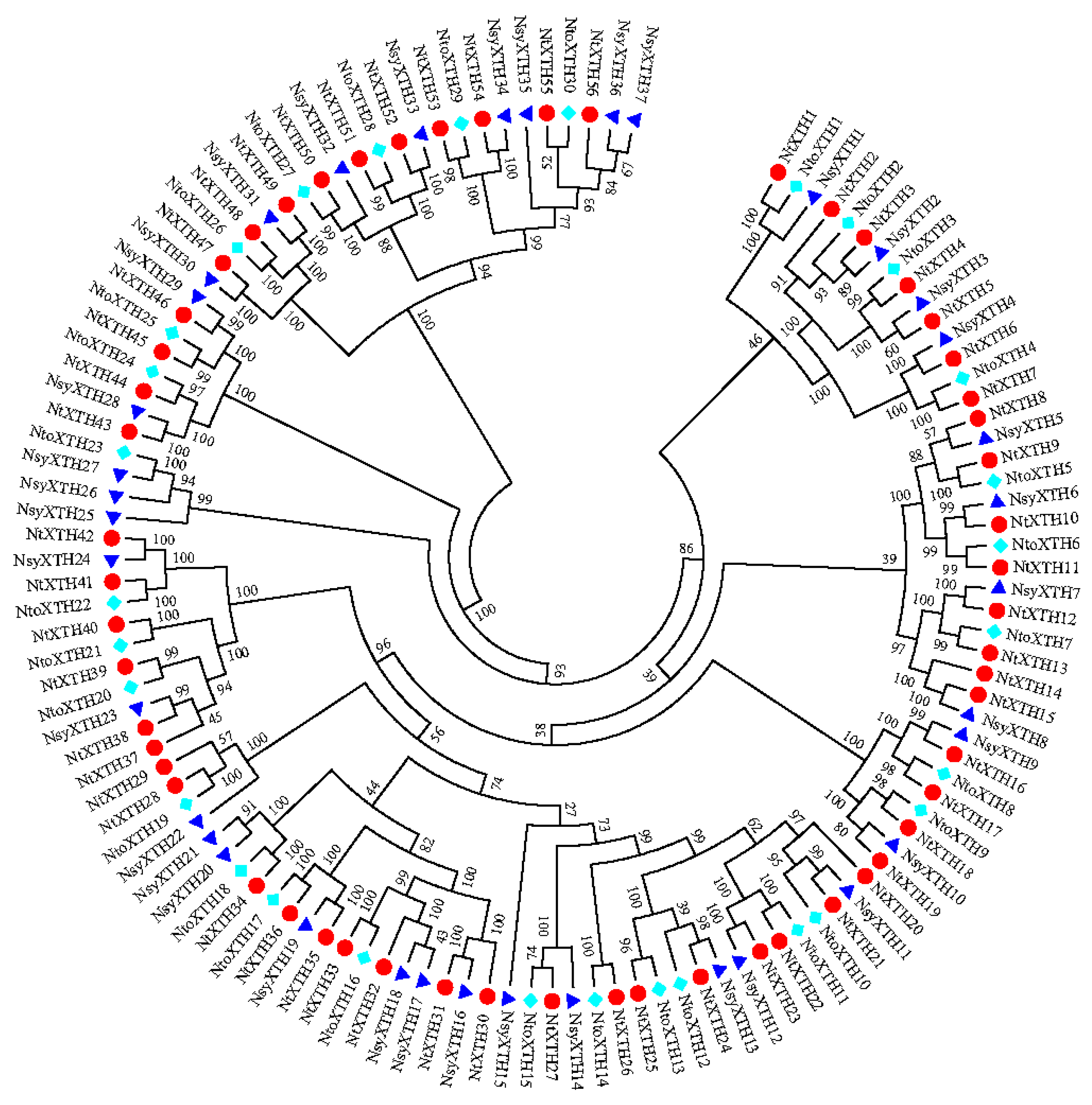
3.2. Phylogenetic and Structural Analyses of NtXTH Genes
3.3. Chromosomal Location, Gene Duplication, and Evolution
3.4. Analysis of XTH Genes in N. tabacum, N. tomentosiformis, N. sylvestris, Arabidopsis, and Rice
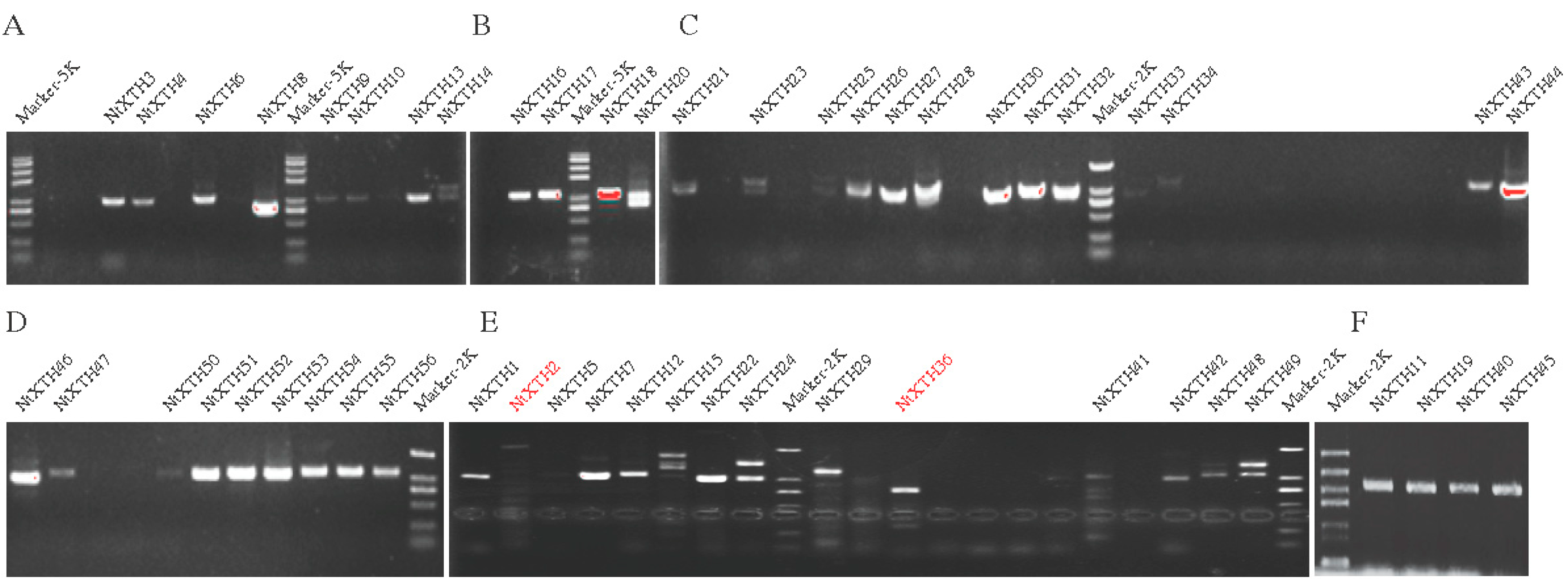
3.5. Isolation and Sequencing of NtXTH cDNAs
3.6. Subcellular Localization of NtXTH Proteins
3.7. Expression of NtXTH Genes during the Growth Cycle
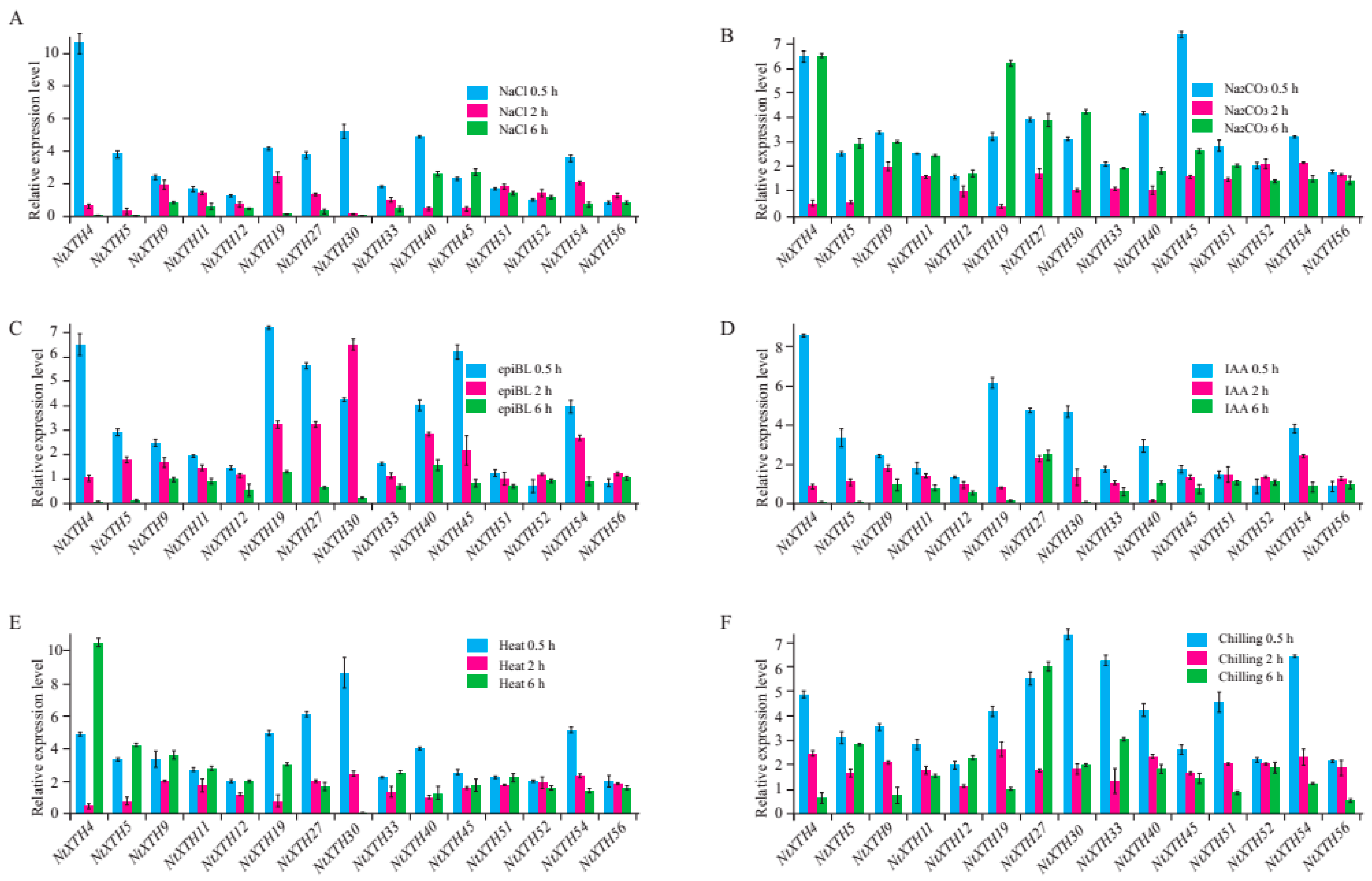
3.8. Expression Profiling of NtXTH Genes under Abiotic Stress Using qRT-PCR
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Acknowledgments
Conflicts of Interest
References
- O’Neill, M.A.; York, W.S. The composition and structure of plant primary cell walls. Plant Cell Wall 2003, 1–54. [Google Scholar] [CrossRef]
- Saladie, M.; Rose, J.K.; Cosgrove, D.J.; Catala, C. Characterization of a new xyloglucan endotransglucosylase/hydrolase (XTH) from ripening tomato fruit and implications for the diverse modes of enzymic action. Plant J. 2006, 47, 282–295. [Google Scholar] [CrossRef] [PubMed]
- Buckeridge, M.S.; Santos, H.P.; Tiné, M.A.S. Mobilisation of storage cell wall polysaccharides in seeds. Plant Physiol. Biochem. 2000, 38, 141–156. [Google Scholar] [CrossRef]
- Cantarel, B.L.; Coutinho, P.M.; Rancurel, C.; Bernard, T.; Lombard, V.; Henrissat, B. The Carbohydrate-Active EnZymes database (CAZy): An expert resource for Glycogenomics. Nucleic Acids Res. 2009, 37, D233–D238. [Google Scholar] [CrossRef] [PubMed]
- Atkinson, R.G.; Johnston, S.L.; Yauk, Y.K.; Sharma, N.N.; Schröder, R. Analysis of xyloglucan endotransglucosylase/hydrolase (XTH) gene families in kiwifruit and apple. Postharvest Biol. Technol. 2009, 51, 149–157. [Google Scholar] [CrossRef]
- Geisler, L.J.; Geisler, M.; Coutinho, P.M.; Segerman, B.; Nishikubo, N.; Takahashi, J.; Aspeborg, H.; Djerbi, S.; Master, E.; Andersson, G.S.; et al. Poplar carbohydrate-active enzymes. Gene identification and expression analyses. Plant Physiol. 2006, 140, 946–962. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Liu, D.; Zhang, H.; Gao, H.; Guo, X.; Wang, D.; Zhang, X.; Zhang, A. The alpha- and beta-expansin and xyloglucan endotransglucosylase/hydrolase gene families of wheat: Molecular cloning, gene expression, and EST data mining. Genomics 2007, 90, 516–529. [Google Scholar] [CrossRef] [PubMed]
- Miedes, E.; Lorences, E.P. Xyloglucan endotransglucosylase/hydrolases (XTHs) during tomato fruit growth and ripening. J. Plant Physiol. 2009, 166, 489–498. [Google Scholar] [CrossRef] [PubMed]
- Rai, K.M.; Thu, S.W.; Balasubramanian, V.K.; Cobos, C.J.; Disasa, T.; Mendu, V. Identification, characterization, and expression analysis of cell wall related genes in Sorghum bicolor (L.) Moench, a food, fodder, and biofuel crop. Front. Plant Sci. 2016, 7, 1287. [Google Scholar] [CrossRef] [PubMed]
- Yokoyama, R.; Nishitani, K. A comprehensive expression analysis of all members of a gene family encoding cell-wall enzymes allowed us to predict cis-regulatory regions involved in cell-wall construction in specific organs of Arabidopsis. Plant Cell Physiol. 2001, 42, 1025–1033. [Google Scholar] [CrossRef] [PubMed]
- Yokoyama, R.; Rose, J.K.; Nishitani, K. A surprising diversity and abundance of xyloglucan endotransglucosylase/hydrolases in rice. Classification and expression analysis. Plant Physiol. 2004, 134, 1088–1099. [Google Scholar] [CrossRef] [PubMed]
- Rose, J.K.; Braam, J.; Fry, S.C.; Nishitani, K. The XTH family of enzymes involved in xyloglucan endotransglucosylation and endohydrolysis: Current perspectives and a new unifying nomenclature. Plant Cell Physiol. 2002, 43, 1421–1435. [Google Scholar] [CrossRef] [PubMed]
- Cosgrove, D.J. Growth of the plant cell wall. Nat. Rev. Mol. Cell Biol. 2005, 6, 850–861. [Google Scholar] [CrossRef] [PubMed]
- Osato, Y.; Yokoyama, R.; Nishitani, K. A principal role for AtXTH18 in Arabidopsis thaliana root growth: A functional analysis using RNAi plants. J. Plant Res. 2006, 119, 153–162. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Jeong, B.R.; Fry, S.C.; Boyer, J.S. Change in XET activities, cell wall extensibility and hypocotyl elongation of soybean seedlings at low water potential. Planta 2005, 220, 593–601. [Google Scholar] [CrossRef] [PubMed]
- Matsui, A.; Yokoyama, R.; Seki, M.; Ito, T.; Shinozaki, K.; Takahashi, T.; Komeda, Y.; Nishitani, K. AtXTH27 plays an essential role in cell wall modification during the development of tracheary elements. Plant J. 2005, 42, 525–534. [Google Scholar] [CrossRef] [PubMed]
- Harada, T.; Torii, Y.; Morita, S.; Onodera, R.; Hara, Y.; Yokoyama, R.; Nishitani, K.; Satoh, S. Cloning, characterization, and expression of xyloglucan endotransglucosylase/hydrolase and expansin genes associated with petal growth and development during carnation flower opening. J. Exp. Bot. 2011, 62, 815–823. [Google Scholar] [CrossRef] [PubMed]
- Singh, A.P.; Tripathi, S.K.; Nath, P.; Sane, A.P. Petal abscission in rose is associated with the differential expression of two ethylene-responsive xyloglucan endotransglucosylase/hydrolase genes, RbXTH1 and RbXTH2. J. Exp. Bot. 2011, 62, 5091–5103. [Google Scholar] [CrossRef] [PubMed]
- Nishikubo, N.; Takahashi, J.; Roos, A.A.; Derba, M.M.; Piens, K.; Brumer, H.; Teeri, T.T.; Stalbrand, H.; Mellerowicz, E.J. Xyloglucan endo-transglycosylase-mediated xyloglucan rearrangements in developing wood of hybrid aspen. Plant Physiol. 2011, 155, 399–413. [Google Scholar] [CrossRef] [PubMed]
- Cho, S.K.; Kim, J.E.; Park, J.A.; Eom, T.J.; Kim, W.T. Constitutive expression of abiotic stress-inducible hot pepper CaXTH3, which encodes a xyloglucan endotransglucosylase/hydrolase homolog, improves drought and salt tolerance in transgenic Arabidopsis plants. FEBS Lett. 2006, 580, 3136–3144. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.; Burns, T.H.; Light, G.; Sun, Y.; Fokar, M.; Kasukabe, Y.; Fujisawa, K.; Maekawa, Y.; Allen, R.D. Xyloglucan endotransglycosylase/hydrolase genes in cotton and their role in fiber elongation. Planta 2010, 232, 1191–1205. [Google Scholar] [CrossRef] [PubMed]
- Xu, W.; Campbell, P.; Vargheese, A.K.; Braam, J. The Arabidopsis XET-related gene family: Environmental and hormonal regulation of expression. Plant J. 1996, 9, 879–889. [Google Scholar] [CrossRef] [PubMed]
- Choi, J.Y.; Seo, Y.S.; Kim, S.J.; Kim, W.T.; Shin, J.S. Constitutive expression of CaXTH3, a hot pepper xyloglucan endotransglucosylase/hydrolase, enhanced tolerance to salt and drought stresses without phenotypic defects in tomato plants (Solanum lycopersicum cv. Dotaerang. Plant Cell Rep. 2011, 30, 867–877. [Google Scholar] [CrossRef] [PubMed]
- Zhu, J.; Alvarez, S.; Marsh, E.L.; Lenoble, M.E.; Cho, I.J.; Sivaguru, M.; Chen, S.; Nguyen, H.T.; Wu, Y.; Schachtman, D.P.; et al. Cell wall proteome in the maize primary root elongation zone. II. Region-specific changes in water soluble and lightly ionically bound proteins under water deficit. Plant Physiol. 2007, 145, 1533–1548. [Google Scholar] [CrossRef] [PubMed]
- Yang, K.A.; Lim, C.J.; Hong, J.K.; Park, C.Y.; Cheong, Y.H.; Chung, W.S.; Lee, K.O.; Lee, S.Y.; Cho, M.J.; Lim, C.O. Identification of cell wall genes modified by a permissive high temperature in Chinese cabbage. Plant Sci. 2006, 171, 175–182. [Google Scholar] [CrossRef]
- Dong, J.; Jiang, Y.; Chen, R.; Xu, Z.; Gao, X. Isolation of a novel xyloglucan endotransglucosylase (OsXET9) gene from rice and analysis of the response of this gene to abiotic stresses. Afr. J. Biotechnol. 2011, 10, 17424–17434. [Google Scholar]
- Kuluev, B.; Mikhaylova, E.; Berezhneva, Z.; Nikonorov, Y.; Postrigan, B.; Kudoyarova, G.; Chemeris, A. Expression profiles and hormonal regulation of tobacco NtEXGT gene and its involvement in abiotic stress response. Plant Physiol. Biochem. 2017, 111, 203–215. [Google Scholar] [CrossRef] [PubMed]
- Alexandersson, E.; Becker, J.V.; Jacobson, D.; Nguema, E.O.; Steyn, C.; Denby, K.J.; Vivier, M.A. Constitutive expression of a grapevine polygalacturonase-inhibiting protein affects gene expression and cell wall properties in uninfected tobacco. BMC Res. Notes 2011, 4, 493. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sierro, N.; Battey, J.N.; Ouadi, S.; Bakaher, N.; Bovet, L.; Willig, A.; Goepfert, S.; Peitsch, M.C.; Ivanov, N.V. The tobacco genome sequence and its comparison with those of tomato and potato. Nat. Commun. 2014, 5, 3833. [Google Scholar] [CrossRef] [PubMed]
- Edwards, K.D.; Bombarely, A.; Story, G.W.; Allen, F.; Mueller, L.A.; Coates, S.A.; Jones, L. TobEA: An atlas of tobacco gene expression from seed to senescence. BMC Genom. 2010, 11, 142. [Google Scholar] [CrossRef] [PubMed]
- Ali, J.; Chen, X.; Chen, Z.; Zhao, H.; Zhao, Y.; Cheng, B.; Xiang, Y. Genome-wide analysis of soybean HD-Zip gene family and expression profiling under salinity and drought treatments. PLoS ONE 2014, 9, e87156. [Google Scholar]
- He, H.; Serraj, R.; Yang, Q. Changes in OsXTH gene expression, ABA content, and peduncle elongation in rice subjected to drought at the reproductive stage. Acta Physiol. Plant. 2009, 31, 749–756. [Google Scholar] [CrossRef]
- Leitch, I.J.; Hanson, L.; Lim, K.Y.; Kovarik, A.; Chase, M.W.; Clarkson, J.J.; Leitch, A.R. The ups and downs of genome size evolution in polyploid species of Nicotiana (Solanaceae). Ann. Bot. 2008, 101, 805–814. [Google Scholar] [CrossRef] [PubMed]
- Tamura, K.; Stecher, G.; Peterson, D.; Filipski, A.; Kumar, S. MEGA6: Molecular evolutionary genetics analysis version 6.0. Mol. Biol. Evol. 2013, 30, 2725–2729. [Google Scholar] [CrossRef] [PubMed]
- Guo, M.; Lu, J.P.; Zhai, Y.F.; Chai, W.G.; Gong, Z.H.; Lu, M.H. Genome-wide analysis, expression profile of heat shock factor gene family (CaHsfs) and characterisation of CaHsfA2 in pepper (Capsicum annuum L.). BMC Plant Biol. 2015, 15, 151. [Google Scholar] [CrossRef] [PubMed]
- Hu, B.; Jin, J.; Guo, A.Y.; Zhang, H.; Luo, J.; Gao, G. GSDS 2.0: An upgraded gene feature visualization server. Bioinformatics 2015, 31, 1296–1297. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Guo, K.; Yu, L.; Tu, Y.; Hu, H.; Wang, B.; Cui, X.; Peng, L. Expression profiling and integrative analysis of the CESA/CSL superfamily in rice. BMC Plant Biol. 2010, 10, 282. [Google Scholar] [CrossRef] [PubMed]
- Librado, P.; Rozas, J. DnaSP v5: A software for comprehensive analysis of DNA polymorphism data. Bioinformatics 2009, 25, 1451–1452. [Google Scholar] [CrossRef] [PubMed]
- Doerks, T.; Copley, R.R.; Schultz, J.; Ponting, C.P.; Bork, P. Systematic identification of novel protein domain families associated with nuclear functions. Genome Res. 2002, 12, 47–56. [Google Scholar] [CrossRef] [PubMed]
- Lynch, M.; Conery, J.S. The evolutionary fate and consequences of duplicate genes. Science 2000, 290, 1151–1155. [Google Scholar] [CrossRef] [PubMed]
- Saeed, A.I.; Sharov, V.; White, J.; Li, J.; Liang, W.; Bhagabati, N.; Braisted, J.; Klapa, M.; Currier, T.; Thiagarajan, M.; et al. TM4: A free, open-source system for microarray data management and analysis. Biotechniques 2003, 34, 374–378. [Google Scholar] [PubMed]
- Han, Y.; Wang, W.; Sun, J.; Ding, M.; Zhao, R.; Deng, S.; Wang, F.; Hu, Y.; Wang, Y.; Lu, Y.; et al. Populus euphratica XTH overexpression enhances salinity tolerance by the development of leaf succulence in transgenic tobacco plants. J. Exp. Bot. 2013, 64, 4225–4238. [Google Scholar] [CrossRef] [PubMed]
- Xu, Z.; Wang, M.; Shi, D.; Zhou, G.; Niu, T.; Hahn, M.G.; O’Neill, M.A.; Kong, Y. DGE-seq analysis of MUR3-related Arabidopsis mutants provides insight into how dysfunctional xyloglucan affects cell elongation. Plant Sci. 2017, 258, 156–169. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2-ΔΔCt method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Kimura, S.; Laosinchai, W.; Itoh, T.; Cui, X.; Linder, C.R.; Brown, R.M. Immunogold labeling of rosette terminal cellulose-synthesizing complexes in the vascular plant Vigna angularis. Plant Cell 1999, 11, 2075–2085. [Google Scholar] [CrossRef] [PubMed]
- Batoko, H.; Zheng, H.Q.; Hawes, C.; Moore, I. A Rab1 GTPase is required for transport between the endoplasmic reticulum and golgi apparatus and for normal golgi movement in plants. Plant Cell 2000, 12, 2201–2217. [Google Scholar] [CrossRef] [PubMed]
- Hu, R.; Qi, G.; Kong, Y.; Kong, D.; Gao, Q.; Zhou, G. Comprehensive analysis of NAC domain transcription factor gene family in Populus trichocarpa. BMC Plant Biol. 2010, 10, 145. [Google Scholar] [CrossRef] [PubMed]
- Pena, M.J.; Darvill, A.G.; Eberhard, S.; York, W.S.; O’Neill, M.A. Moss and liverwort xyloglucans contain galacturonic acid and are structurally distinct from the xyloglucans synthesized by hornworts and vascular plants. Glycobiology 2008, 18, 891–904. [Google Scholar] [CrossRef] [PubMed]
- Leitch, I.J.; Bennett, M.D. Genome downsizing in polyploid plants. Biol. J. Linn. Soc. 2004, 82, 651–663. [Google Scholar] [CrossRef]
- Sierro, N.; Battey, J.N.; Ouadi, S.; Bovet, L.; Goepfert, S.; Bakaher, N.; Peitsch, M.C.; Ivanov, N.V. Reference genomes and transcriptomes of Nicotiana sylvestris and Nicotiana tomentosiformis. Genome Biol. 2013, 14, R60. [Google Scholar] [CrossRef] [PubMed]
- Okazawa, K.; Sato, Y.; Nakagawa, T.; Asada, K.; Kato, I.; Tomita, E.; Nishitani, K. Molecular cloning and cDNA sequencing of endoxyloglucan transferase, a novel class of glycosyltransferase that mediates molecular grafting between matrix polysaccharides in plant cell walls. J. Biol. Chem. 1993, 268, 25364–25368. [Google Scholar] [PubMed]
- Campbell, P.; Braam, J. Xyloglucan endotransglycosylases: Diversity of genes, enzymes and potential wall-modifying functions. Trends Plant Sci. 1999, 4, 361–366. [Google Scholar] [CrossRef]
- Campbell, P.; Braam, J. Co- and/or post-translational modifications are critical for TCH4 XET activity. Plant J. 1998, 15, 553–561. [Google Scholar] [CrossRef] [PubMed]
- Han, Y.; Ban, Q.; Li, H.; Hou, Y.; Jin, M.; Han, S.; Rao, J. DkXTH8, a novel xyloglucan endotransglucosylase/hydrolase in persimmon, alters cell wall structure and promotes leaf senescence and fruit postharvest softening. Sci. Rep. 2016, 6, 39155. [Google Scholar] [CrossRef] [PubMed]
- Xu, W.; Purugganan, M.M.; Polisensky, D.H.; Antosiewicz, D.M.; Fry, S.C.; Braam, J. Arabidopsis TCH4, regulated by hormones and the environment, encodes a xyloglucan endotransglycosylase. Plant Cell 1995, 7, 1555–1567. [Google Scholar] [CrossRef] [PubMed]
- Vissenberg, K.; Fry, S.C.; Verbelen, J.P. Root hair initiation is coupled to a highly localized increase of xyloglucan endotransglycosylase action in Arabidopsis roots. Plant Physiol. 2001, 127, 1125–1135. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | Accession NO. | Previous Gene Name | Signal Position | Orthologous in Arabidopsis (ID%) | Protein Length | Intron | Sub-Cellular Localization | MW (Da) | PI |
|---|---|---|---|---|---|---|---|---|---|
| NtXTH1 | mRNA_14204 | 1-23 | 67.59 | 293 | 4 | Plasma membrane | 34,168.57 | 7.68 | |
| NtXTH2 | mRNA_63239 | 1-16 | 79.09 | 219 | 4 | Plasma membrane | 25,787.6 | 4.76 | |
| NtXTH3 | mRNA_137463 | 1-20 | 72.26 | 291 | 4 | Plasma membrane | 33,977.82 | 4.72 | |
| NtXTH4 | mRNA_134286 | 1-27 | 73.56 | 298 | 4 | Plasma membrane | 35,038.32 | 5.46 | |
| NtXTH5 | mRNA_87242 | 1-27 | 73.47 | 298 | 4 | Plasma membrane | 34,932.28 | 5.46 | |
| NtXTH6 | mRNA_14392 | 1-27 | 68.58 | 299 | 4 | Plasma membrane | 34,613.85 | 6.05 | |
| NtXTH7 | mRNA_82272 | 1-24 | 68.29 | 296 | 4 | Plasma membrane | 34,346.59 | 6.23 | |
| NtXTH8 | mRNA_35364 | 1-18 | 82.53 | 242 | 3 | Plasma membrane | 27,862.47 | 6.99 | |
| NtXTH9 | mRNA_93233 | 1-27 | 79.25 | 291 | 3 | Plasma membrane | 33,061.33 | 7.04 | |
| NtXTH10 | mRNA_35366 | 1-28 | 76.16 | 292 | 4 | Plasma membrane | 33,239.62 | 8.15 | |
| NtXTH11 | mRNA_50079 | KJ730270 [27] | 1-28 | 76.51 | 292 | 4 | Plasma membrane | 33,221.52 | 7.64 |
| NtXTH12 | mRNA_23269 | 1-25 | 70.93 | 298 | 4 | Plasma membrane | 34,042.1 | 5.69 | |
| NtXTH13 | mRNA_58849 | 1-25 | 71.28 | 298 | 4 | Plasma membrane | 33,987.21 | 5.89 | |
| NtXTH14 | mRNA_78608 | 1-20 | 54.68 | 283 | 4 | Plasma membrane | 31,667.75 | 5.74 | |
| NtXTH15 | mRNA_92550 | 1-21 | 56.49 | 288 | 5 | Plasma membrane | 32,624.85 | 6.75 | |
| NtXTH16 | mRNA_86110 | 1-18 | 73.08 | 299 | 4 | Plasma membrane | 34,646.33 | 8.87 | |
| NtXTH17 | mRNA_68366 | 1-21 | 74.13 | 302 | 4 | Plasma membrane | 35,183.88 | 8.95 | |
| NtXTH18 | mRNA_104426 | AB017025.1 [28] | 1-21 | 70.85 | 295 | 4 | Plasma membrane | 33,923.42 | 8.65 |
| NtXTH19 | mRNA_123885 | D86730.1 [27] | 1-21 | 71.77 | 295 | 4 | Plasma membrane | 33,882.41 | 8.64 |
| NtXTH20 | mRNA_95691 | 1-10 | 66.51 | 217 | 3 | Plasma membrane | 24,690.52 | 8.95 | |
| NtXTH21 | mRNA_39798 | 1-25 | 61.97 | 291 | 3 | Plasma membrane | 32,880.96 | 9.13 | |
| NtXTH22 | mRNA_18536 | 1-20 | 63.54 | 284 | 3 | Plasma membrane | 31,877.64 | 8.7 | |
| NtXTH23 | mRNA_34533 | 1-20 | 62.95 | 284 | 3 | Plasma membrane | 31,884.77 | 8.98 | |
| NtXTH24 | mRNA_9973 | 1-25 | 61.05 | 290 | 3 | Plasma membrane | 32,549.69 | 8.97 | |
| NtXTH25 | mRNA_122713 | 1-25 | 60.87 | 290 | 3 | Plasma membrane | 32,705.83 | 8.97 | |
| NtXTH26 | mRNA_60328 | 1-30 | 63.25 | 300 | 3 | Plasma membrane | 33,809.7 | 8.17 | |
| NtXTH27 | mRNA_52501 | 1-21 | 73.19 | 273 | 3 | Plasma membrane | 31,689.83 | 9.3 | |
| NtXTH28 | mRNA_99357 | 1-36 | 62.31 | 337 | 5 | Plasma membrane | 38,692.06 | 9.37 | |
| NtXTH29 | mRNA_110655 | 1-36 | 61.98 | 337 | 5 | Plasma membrane | 38,660.91 | 9.17 | |
| NtXTH30 | mRNA_75358 | 1-23 | 66.78 | 285 | 3 | Plasma membrane | 32,501.78 | 7.58 | |
| NtXTH31 | mRNA_51471 | 1-23 | 67.25 | 288 | 3 | Plasma membrane | 32,728.07 | 6.82 | |
| NtXTH32 | mRNA_84816 | 1-17 | 71.54 | 279 | 3 | Plasma membrane | 31,762.65 | 7.61 | |
| NtXTH33 | mRNA_55584 | 1-17 | 71.59 | 279 | 3 | Plasma membrane | 31,527.43 | 7.61 | |
| NtXTH34 | mRNA_83457 | 1-22 | 69 | 272 | 3 | Plasma membrane | 30,676.35 | 5.84 | |
| NtXTH35 | mRNA_8391 | HQ108341.1 [27] | 1-23 | 71.26 | 286 | 3 | Plasma membrane | 32,690.84 | 8.47 |
| NtXTH36 | mRNA_83477 | 1-23 | 71.1 | 289 | 3 | Plasma membrane | 32,898.04 | 8.12 | |
| NtXTH37 | mRNA_21254 | 1-26 | 60.5 | 290 | 4 | Plasma membrane | 33,221.98 | 9.11 | |
| NtXTH38 | mRNA_77290 | 1-26 | 62.06 | 290 | 4 | Plasma membrane | 33,268.95 | 9 | |
| NtXTH39 | mRNA_11279 | 1-26 | 61.21 | 290 | 4 | Plasma membrane | 33,184.82 | 9.11 | |
| NtXTH40 | mRNA_126220 | 1-25 | 60.76 | 289 | 4 | Plasma membrane | 33,142.73 | 9.1 | |
| NtXTH41 | mRNA_126221 | 1-30 | 59.01 | 294 | 4 | Plasma membrane | 33,286.39 | 6.48 | |
| NtXTH42 | mRNA_21253 | 1-30 | 60.5 | 294 | 4 | Plasma membrane | 33,467.71 | 7.66 | |
| NtXTH43 | mRNA_18656 | 1-20 | 82.5 | 296 | 4 | Extracellular | 34,255.72 | 9.4 | |
| NtXTH44 | mRNA_2368 | 1-20 | 80.99 | 296 | 4 | Extracellular | 34,223.71 | 9.38 | |
| NtXTH45 | mRNA_110606 | 1-20 | 70.71 | 295 | 4 | Extracellular | 33,679.71 | 8.73 | |
| NtXTH46 | mRNA_99979 | 1-19 | 71.01 | 294 | 4 | Extracellular | 33,602.71 | 8.95 | |
| NtXTH47 | mRNA_36960 | 1-26 | 57.1 | 316 | 4 | Extracellular | 36,336.03 | 6.26 | |
| NtXTH48 | mRNA_124647 | 1-26 | 58.09 | 316 | 4 | Plasma membrane | 35,865.25 | 6.24 | |
| NtXTH49 | mRNA_98504 | 1-26 | 58.39 | 316 | 4 | Plasma membrane | 36,054.57 | 6.14 | |
| NtXTH50 | mRNA_32955 | 1-30 | 63.88 | 365 | 4 | Extracellular | 41,821.13 | 8.95 | |
| NtXTH51 | mRNA_60999 | 1-23 | 63.73 | 339 | 4 | Extracellular | 39,602.53 | 7.19 | |
| NtXTH52 | mRNA_33005 | 1-25 | 61.95 | 343 | 4 | Extracellular | 39,847.83 | 6.92 | |
| NtXTH53 | mRNA_20939 | 1-20 | 67.7 | 331 | 4 | Plasma membrane | 37,918.84 | 6.64 | |
| NtXTH54 | mRNA_8840 | 1-20 | 67.61 | 331 | 4 | Plasma membrane | 37,970.88 | 6.64 | |
| NtXTH55 | mRNA_119817 | 1-20 | 68.1 | 330 | 4 | Plasma membrane | 37,969.77 | 6.56 | |
| NtXTH56 | mRNA_89227 | 1-20 | 69.9 | 330 | 4 | Plasma membrane | 37,820.69 | 7.64 |
| Paralogous pairs | Score | Identities | Similarity | Gaps | SynDif | SynPos | Ks | NSynDif | NSynPos | Ka | Ka/Ks | T (Mya) |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| NtXTH2-NtXTH3 | 1234 | 214/291 (73.5%) | 218/291 (74.9%) | 72/291 (24.7%) | 16 | 138.4 | 0.13 | 5 | 518.58 | 0.01 | 0.08 | 10.29 |
| NtXTH4-NtXTH5 | 1655 | 292/298 (98.0%) | 297/298 (99.7%) | 0/298 (0.0%) | 31 | 184.5 | 0.19 | 6 | 709.50 | 0.01 | 0.04 | 15.59 |
| NtXTH6-NtXTH7 | 1574 | 282/299 (94.3%) | 287/299 (96.0%) | 3/299 (1.0%) | 35 | 186.5 | 0.22 | 15 | 701.50 | 0.02 | 0.10 | 17.70 |
| NtXTH10-NtXTH11 | 1569 | 287/292 (98.3%) | 289/292 (99.0%) | 0/292 (0.0%) | 23 | 199.3 | 0.13 | 5 | 676.67 | 0.01 | 0.06 | 10.27 |
| NtXTH12-NtXTH13 | 1565 | 287/298 (96.3%) | 291/298 (97.7%) | 0/298 (0.0%) | 39 | 197.1 | 0.23 | 14 | 696.92 | 0.02 | 0.09 | 18.83 |
| NtXTH14-NtXTH15 | 1390 | 256/288 (88.9%) | 266/288 (92.4%) | 5/288 (1.7%) | 19.3 | 191.1 | 0.11 | 44.7 | 654.92 | 0.07 | 0.66 | 8.91 |
| NtXTH16-NtXTH17 | 1610 | 292/302 (96.7%) | 296/302 (98.0%) | 3/302 (1.0%) | 24 | 189.3 | 0.14 | 11 | 707.67 | 0.02 | 0.11 | 11.39 |
| NtXTH18-NtXTH19 | 1603 | 291/295 (98.6%) | 294/295 (99.7%) | 0/295 (0.0%) | 27 | 200.6 | 0.15 | 4 | 684.42 | 0.01 | 0.04 | 12.16 |
| NtXTH20-NtXTH21 | 1179 | 213/291 (73.2%) | 213/291 (73.2%) | 74/291 (25.4%) | 19 | 142.7 | 0.15 | 5 | 508.33 | 0.01 | 0.07 | 12.02 |
| NtXTH22-NtXTH23 | 1510 | 276/284 (97.2%) | 280/284 (98.6%) | 0/284 (0.0%) | 35 | 191.8 | 0.21 | 9 | 660.25 | 0.01 | 0.07 | 17.15 |
| NtXTH24-NtXTH25 | 1543 | 284/290 (97.9%) | 286/290 (98.6%) | 0/290 (0.0%) | 41.5 | 195.8 | 0.25 | 7.5 | 674.17 | 0.01 | 0.04 | 20.41 |
| NtXTH28-NtXTH29 | 1757 | 326/337 (96.7%) | 333/337 (98.8%) | 0/337 (0.0%) | 23 | 227.7 | 0.11 | 11 | 783.33 | 0.01 | 0.13 | 8.89 |
| NtXTH30-NtXTH31 | 1455.5 | 271/288 (94.1%) | 277/288 (96.2%) | 3/288 (1.0%) | 24 | 191.3 | 0.14 | 15 | 663.67 | 0.02 | 0.17 | 11.25 |
| NtXTH32-NtXTH33 | 1441 | 264/279 (94.6%) | 274/279 (98.2%) | 0/279 (0.0%) | 18 | 184.9 | 0.10 | 15 | 652.08 | 0.02 | 0.22 | 8.55 |
| NtXTH35-NtXTH36 | 1458 | 271/289 (93.8%) | 274/289 (94.8%) | 3/289 (1.0%) | 30 | 187.9 | 0.18 | 16 | 670.08 | 0.02 | 0.14 | 14.71 |
| NtXTH37-NtXTH38 | 1540 | 277/290 (95.5%) | 283/290 (97.6%) | 0/290 (0.0%) | 28 | 191.2 | 0.16 | 13 | 678.83 | 0.02 | 0.12 | 13.36 |
| NtXTH41-NtXTH42 | 1562 | 282/294 (95.9%) | 285/294 (96.9%) | 0/294 (0.0%) | 22 | 195.2 | 0.12 | 15 | 686.83 | 0.02 | 0.18 | 10.02 |
| NtXTH43-NtXTH44 | 1625 | 288/296 (97.3%) | 294/296 (99.3%) | 0/296 (0.0%) | 23 | 204.9 | 0.12 | 8 | 683.08 | 0.01 | 0.10 | 9.97 |
| NtXTH45-NtXTH46 | 1558 | 283/295 (95.9%) | 291/295 (98.6%) | 1/295 (0.3%) | 13 | 204.9 | 0.07 | 11 | 677.08 | 0.02 | 0.25 | 5.43 |
| NtXTH48-NtXTH49 | 1591 | 295/316 (93.4%) | 303/316 (95.9%) | 0/316 (0.0%) | 15 | 222.5 | 0.07 | 22 | 725.50 | 0.03 | 0.44 | 5.79 |
| NtXTH51-NtXTH52 | 1772 | 330/343 (96.2%) | 333/343 (97.1%) | 4/343 (1.2%) | 25.5 | 219.4 | 0.13 | 10.5 | 797.58 | 0.01 | 0.11 | 10.35 |
| NtXTH53-NtXTH54 | 1739 | 320/331 (96.7%) | 325/331 (98.2%) | 0/331 (0.0%) | 16 | 227.0 | 0.07 | 11 | 766.60 | 0.01 | 0.20 | 6.07 |
| NtXTH55-NtXTH56 | 1692 | 313/330 (94.8%) | 320/330 (97.0%) | 0/330 (0.0%) | 34 | 225.8 | 0.17 | 18 | 764.25 | 0.02 | 0.14 | 13.78 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, M.; Xu, Z.; Ding, A.; Kong, Y. Genome-Wide Identification and Expression Profiling Analysis of the Xyloglucan Endotransglucosylase/Hydrolase Gene Family in Tobacco (Nicotiana tabacum L.). Genes 2018, 9, 273. https://doi.org/10.3390/genes9060273
Wang M, Xu Z, Ding A, Kong Y. Genome-Wide Identification and Expression Profiling Analysis of the Xyloglucan Endotransglucosylase/Hydrolase Gene Family in Tobacco (Nicotiana tabacum L.). Genes. 2018; 9(6):273. https://doi.org/10.3390/genes9060273
Chicago/Turabian StyleWang, Meng, Zongchang Xu, Anming Ding, and Yingzhen Kong. 2018. "Genome-Wide Identification and Expression Profiling Analysis of the Xyloglucan Endotransglucosylase/Hydrolase Gene Family in Tobacco (Nicotiana tabacum L.)" Genes 9, no. 6: 273. https://doi.org/10.3390/genes9060273