The Case of X and Y Localization of Nucleolus Organizer Regions (NORs) in Tragulus javanicus (Cetartiodactyla, Mammalia)

 , and
, and 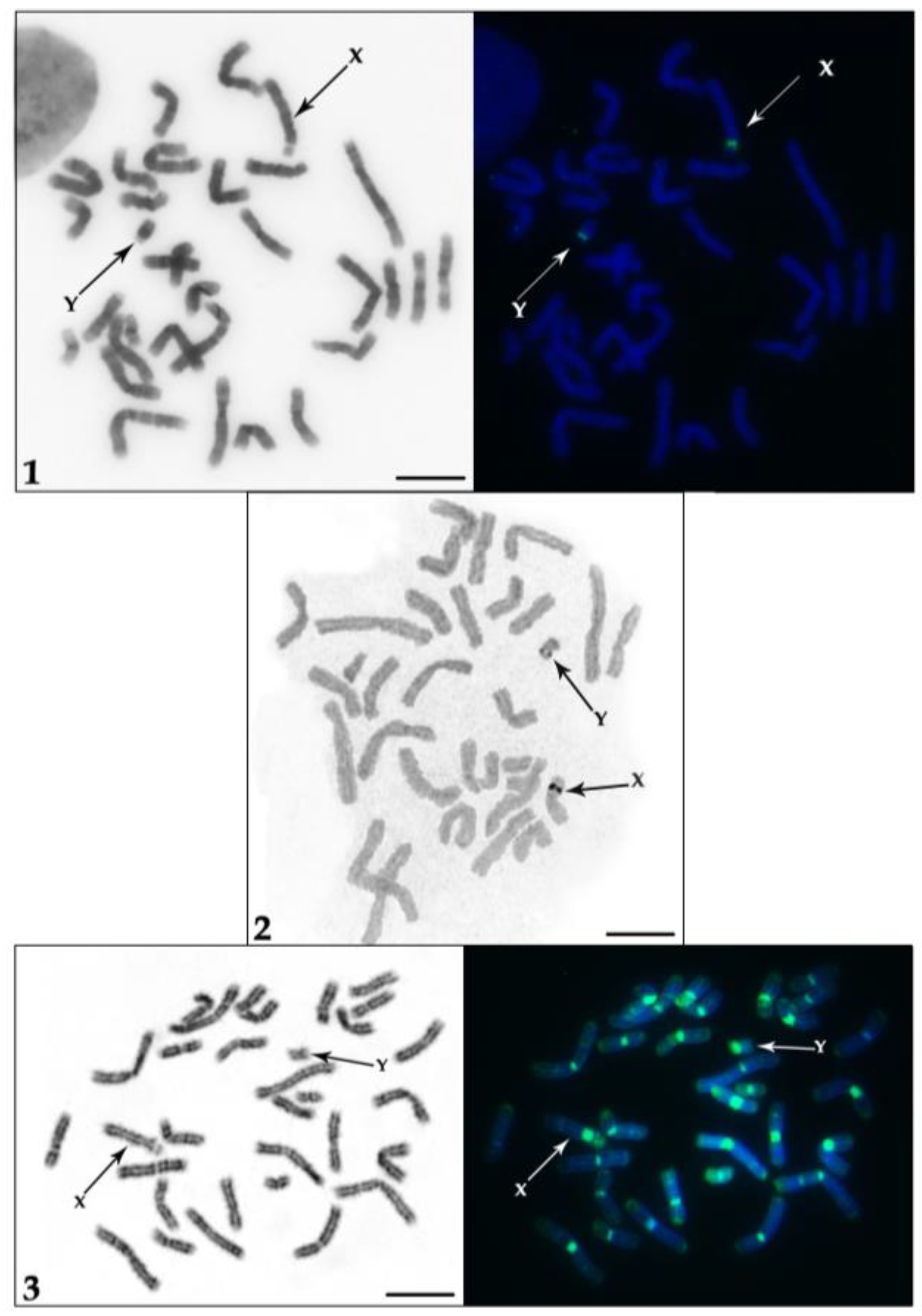{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Culture and Chromosome Preparation
2.2. Nucleolus Organizer Regions Detection
2.3. Silver Staining of Metaphase Chromosome
2.4. The Combined Method of Heterogeneous Heterochromatin Detection (CDAG)
3. Results
4. Discussion
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Olson, M.O.J. The Nucleolus; Springer: Berlin, Germany, 2011; ISBN 978-1-4614-0514-6. [Google Scholar]
- Wilson, G.N. The structure and organization of human ribosomal genes. Cell Nucl. 1982, 10, 289–318. [Google Scholar]
- Winking, H.; Nielsen, K.; Gropp, A. Variable positions of NORs in Mus musculus. Cytogenet. Genome Res. 1980, 26, 158–164. [Google Scholar] [CrossRef] [PubMed]
- Britton-Davidian, J.; Cazaux, B.; Catalan, J. Chromosomal dynamics of nucleolar organizer regions (NORs) in the house mouse: micro-evolutionary insights. Heredity 2012, 108, 68–74. [Google Scholar] [CrossRef] [PubMed]
- Stults, D.M.; Killen, M.W.; Williamson, E.P.; Hourigan, J.S.; Vargas, H.D.; Arnold, S.M.; Moscow, J.A.; Pierce, A.J. Human rRNA gene clusters are recombinational hotspots in cancer. Cancer Res. 2009, 69, 9096–9104. [Google Scholar] [CrossRef] [PubMed]
- Hsu, T.C.; Spirito, S.E.; Pardue, M.L. Distribution of 18+ 28S ribosomal genes in mammalian genomes. Chromosoma 1975, 53, 25–36. [Google Scholar] [CrossRef] [PubMed]
- Gerbault-Seureau, M.; Cacheux, L.; Dutrillaux, B. The Relationship between the (In-) Stability of NORs and their chromosomal location: The example of Cercopithecidae and a short review of other primates. Cytogenet. Genome Res. 2017, 153, 138–146. [Google Scholar] [CrossRef] [PubMed]
- Guillén, A.K.Z.; Hirai, Y.; Tanoue, T.; Hirai, H. Transcriptional repression mechanisms of nucleolus organizer regions (NORs) in humans and chimpanzees. Chromosome Res. 2004, 12, 225–237. [Google Scholar] [CrossRef] [PubMed]
- Hall, K.J.; Parker, J.S. Stable chromosome fission associated with rDNA mobility. Chromosome Res. 1995, 3, 417–422. [Google Scholar] [CrossRef] [PubMed]
- Henderson, A.S.; Warburton, D.; Atwood, K.C. Location of ribosomal DNA in the human chromosome complement. Proc. Natl. Acad. Sci. USA 1972, 69, 3394–3398. [Google Scholar] [CrossRef] [PubMed]
- Henderson, A.S.; Atwood, K.C.; Warburton, D. Chromosomal distribution of rDNA in Pan paniscus, Gorilla gorilla beringei, and Symphalangus syndactylus: Comparison to related primates. Chromosoma 1976, 59, 147–155. [Google Scholar] [CrossRef] [PubMed]
- Cabrero, J.; Camacho, J.P.M. Location and expression of ribosomal RNA genes in grasshoppers: Abundance of silent and cryptic loci. Chromosome Res. 2008, 16, 595–607. [Google Scholar] [CrossRef] [PubMed]
- Kulemzina, A.I.; Proskuryakova, A.A.; Beklemisheva, V.R.; Lemskaya, N.A.; Perelman, P.L.; Graphodatsky, A.S. Comparative chromosome map and heterochromatin features of the gray whale karyotype (Cetacea). Cytogenet. Genome Res. 2016, 148, 25–34. [Google Scholar] [CrossRef] [PubMed]
- Dutrillaux, A.-M.; Carton, B.; Cacheux, L.; Dutrillaux, B. Interstitial NORs, Fragile sites, and chromosome evolution: a not so simple relationship-the example of Melolontha melolontha and genus Protaetia (Coleoptera: Scarabaeidae). Cytogenet. Genome Res. 2016, 149, 304–311. [Google Scholar] [CrossRef] [PubMed]
- Gallagher, D.S.; Davis, S.K.; De Donato, M.; Burzlaff, J.D.; Womack, J.E.; Taylor, J.F.; Kumamoto, A.T. A karyotypic analysis of nilgai, Boselaphus tragocamelus (Artiodactyla: Bovidae). Chromosome Res. 1998, 6, 505–514. [Google Scholar] [CrossRef] [PubMed]
- Volleth, M. Differences in the location of nucleolus organizer regions in European vespertilionid bats. Cytogenet. Genome Res. 1987, 44, 186–197. [Google Scholar] [CrossRef] [PubMed]
- Nirchio, M.; Oliveira, C.; Ferreira, I.A.; Granado, A.; Ron, E. Extensive polymorphism and chromosomal characteristics of ribosomal DNA in the characid fish Triportheus venezuelensis (Characiformes, Characidae). Genet. Mol. Biol. 2007, 30, 25–30. [Google Scholar] [CrossRef] [Green Version]
- Scacchetti, P.C.; Utsunomia, R.; Pansonato-Alves, J.C.; da Costa Silva, G.J.; Vicari, M.R.; Artoni, R.F.; Oliveira, C.; Foresti, F. Repetitive DNA sequences and evolution of ZZ/ZW sex chromosomes in Characidium (Teleostei: Characiformes). PLoS ONE 2015, 10, e0137231. [Google Scholar] [CrossRef] [PubMed]
- Machado, T.C.; Pansonato-Alves, J.C.; Pucci, M.B.; Nogaroto, V.; Almeida, M.C.; Oliveira, C.; Foresti, F.; Bertollo, L.A.; Moreira-Filho, O.; Artoni, R.F.; et al. Chromosomal painting and ZW sex chromosomes differentiation in Characidium (Characiformes, Crenuchidae). BMC Genet. 2011, 12, 65. [Google Scholar] [CrossRef] [PubMed]
- Schartl, M.; Schmid, M.; Nanda, I. Dynamics of vertebrate sex chromosome evolution: from equal size to giants and dwarfs. Chromosoma 2016, 125, 553–571. [Google Scholar] [CrossRef] [PubMed]
- Yano, C.F.; Bertollo, L.A.C.; Molina, W.F.; Liehr, T.; de Bello Cioffi, M. Genomic organization of repetitive DNAs and its implications for male karyotype and the neo-Y chromosome differentiation in Erythrinus erythrinus (Characiformes, Erythrinidae). Comp. Cytogenet. 2014, 8, 139. [Google Scholar] [CrossRef] [PubMed]
- Cooper, D.W.; Johnston, P.G.; Watson, J.M.; Graves, J.A.M. X-inactivation in marsupials and monotremes. In Seminars in Developmental Biology; Elsevier: Amsterdam, The Netherlands, 1993; Volume 4, pp. 117–128. [Google Scholar]
- Merry, D.E.; Pathak, S.; VandeBerg, J.L. Differential NOR activities in somatic and germ cells of Monodelphis domestica (Marsupialia, Mammalia). Cytogenet. Genome Res. 1983, 35, 244–251. [Google Scholar] [CrossRef] [PubMed]
- Goodpasture, C.; Bloom, S.E. Visualization of nucleolar organizer regions in mammalian chromosomes using silver staining. Chromosoma 1975, 53, 37–50. [Google Scholar] [CrossRef] [PubMed]
- Stitou, S.; Jimenez, R.; De La Guardia, R.D.; Burgos, M. Sex-chromosome pairing through heterochromatin in the African rodent Lemniscomys barbarus (Rodentia, Muridae). A synaptonemal complex study. Chromosome Res. 2000, 8, 277–283. [Google Scholar] [CrossRef] [PubMed]
- Pardue, M.L.; Hsu, T.C. Locations of 18S and 28S ribosomal genes on the chromosomes of the Indian muntjac. J. Cell Biol. 1975, 64, 251–254. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stanyon, R.; Galleni, L. A rapid fibroblast culture technique for high resolution karyotypes. Ital. J. Zool. 1991, 58, 81–83. [Google Scholar] [CrossRef] [Green Version]
- Yang, F.; Graphodatsky, A.S. Animal probes and ZOO-FISH. In Fluorescence In Situ Hybridization (FISH); T. Liehr: Berlin, Germany, 2017; pp. 323–346. [Google Scholar]
- Maden, B.E.H.; Dent, C.L.; Farrell, T.E.; Garde, J.; McCALLUM, F.S.; Wakeman, J.A. Clones of human ribosomal DNA containing the complete 18 S-rRNA and 28 S-rRNA genes. Characterization, a detailed map of the human ribosomal transcription unit and diversity among clones. Biochem. J. 1987, 246, 519–527. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seabright, M. A rapid banding technique for human chromosomes. Lancet 1971, 298, 971–972. [Google Scholar] [CrossRef]
- Graphodatsky, A.S.; Radjabli, S.I. Chromosomes of Agricultural and Laboratory Animals. Atlas; Nauka: Novosibirsk, Russia, 1988. [Google Scholar]
- Lemskaya, N.A.; Kulemzina, A.I.; Beklemisheva, V.R.; Biltueva, L.S.; Proskuryakova, A.A.; Perelman, P.L.; Graphodatsky, A.S. The combined method of heterogeneous heterochromatin detection (CDAG) in different mammalian species. Chromosoma 2018. [Google Scholar]
- Gallagher, D.S.; Houck, M.L.; Ryan, A.M.; Womack, J.E.; Kumamoto, A.T. A karyotypic analysis of the lesser Malay chevrotain, Tragulus javanicus (Artiodactyla: Tragulidae). Chromosome Res. 1996, 4, 545–551. [Google Scholar] [CrossRef] [PubMed]
- Mäkinen, A.; Zijlstra, C.; de Haan, N.A.; Mellink, C.H.M.; Bosma, A.A. Localization of 18S+28S and 5S ribosomal RNA genes in the dog by fluorescence in situ hybridization. Cytogenet. Genome Res. 1997, 78, 231–235. [Google Scholar] [CrossRef]
- Kulemzina, A.I.; Yang, F.; Trifonov, V.A.; Ryder, O.A.; Ferguson-Smith, M.A.; Graphodatsky, A.S. Chromosome painting in Tragulidae facilitates the reconstruction of Ruminantia ancestral karyotype. Chromosome Res. 2011, 19, 531. [Google Scholar] [CrossRef] [PubMed]
- Gallagher, D.S.; Davis, S.K.; De Donato, M.; Burzlaff, J.D.; Womack, J.E.; Taylor, J.F.; Kumamoto, A.T. A Molecular Cytogenetic Analysis of the Tribe Bovini (Artiodactyla: Bovidae: Bovinae) with an Emphasis on Sex Сhromosome Morphology and NOR Distribution. Chromosome Res. 1999, 7, 481–492. [Google Scholar] [CrossRef] [PubMed]
- Vozdova, M.; Fröhlich, J.; Kubickova, S.; Sebestova, H.; Rubes, J. Meiotic Recombination in the Giraffe (G. reticulata). Cytogenet. Genome Res. 2017, 152, 73–80. [Google Scholar] [CrossRef] [PubMed]
- Eickbush, T.H.; Eickbush, D.G. Finely orchestrated movements: evolution of the ribosomal RNA genes. Genetics 2007, 175, 477–485. [Google Scholar] [CrossRef] [PubMed]
- Raskina, O.; Barber, J.C.; Nevo, E.; Belyayev, A. Repetitive DNA and chromosomal rearrangements: speciation-related events in plant genomes. Cytogenet. Genome Res. 2008, 120, 351–357. [Google Scholar] [CrossRef] [PubMed]
- Grozdanov, P.; Georgiev, O.; Karagyozov, L. Complete sequence of the 45-kb mouse ribosomal DNA repeat: analysis of the intergenic spacer. Genomics 2003, 82, 637–643. [Google Scholar] [CrossRef]
- Solano, E.; Taylor, P.J.; Rautenbach, A.; Ropiquet, A.; Castiglia, R. Cryptic speciation and chromosomal repatterning in the South African climbing mice Dendromus (Rodentia, Nesomyidae). PLoS ONE 2014, 9, e88799. [Google Scholar] [CrossRef] [PubMed]
- Proskuryakova, A.A.; Kulemzina, A.I.; Perelman, P.L.; Makunin, A.I.; Larkin, D.M.; Farré, M.; Kukekova, A.V.; Johnson, J.L.; Lemskaya, N.A.; Beklemisheva, V.R.; et al. X Chromosome Evolution in Cetartiodactyla. Genes 2017, 8, 216. [Google Scholar] [CrossRef] [PubMed]
- Srivastava, R.; Srivastava, R.; Ahn, S.H. The epigenetic pathways to ribosomal DNA silencing. Microbiol. Mol. Biol. Rev. 2016, 80, 545–563. [Google Scholar] [CrossRef] [PubMed]
- Gottlieb, S.; Esposito, R.E. A new role for a yeast transcriptional silencer gene, SIR2, in regulation of recombination in ribosomal DNA. Cell 1989, 56, 771–776. [Google Scholar] [CrossRef]
- Li, G.; Davis, B.W.; Raudsepp, T.; Wilkerson, A.J.P.; Mason, V.C.; Ferguson-Smith, M.; O’Brien, P.C.; Waters, P.D.; Murphy, W.J. Comparative analysis of mammalian Y chromosomes illuminates ancestral structure and lineage-specific evolution. Genome Res. 2013, 23, 1486–1495. [Google Scholar] [CrossRef] [PubMed] [Green Version]


© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Proskuryakova, A.A.; Kulemzina, A.I.; Perelman, P.L.; Serdukova, N.A.; Ryder, O.A.; Graphodatsky, A.S. The Case of X and Y Localization of Nucleolus Organizer Regions (NORs) in Tragulus javanicus (Cetartiodactyla, Mammalia). Genes 2018, 9, 312. https://doi.org/10.3390/genes9060312
Proskuryakova AA, Kulemzina AI, Perelman PL, Serdukova NA, Ryder OA, Graphodatsky AS. The Case of X and Y Localization of Nucleolus Organizer Regions (NORs) in Tragulus javanicus (Cetartiodactyla, Mammalia). Genes. 2018; 9(6):312. https://doi.org/10.3390/genes9060312
Chicago/Turabian StyleProskuryakova, Anastasia A., Anastasia I. Kulemzina, Polina L. Perelman, Natalia A. Serdukova, Oliver A. Ryder, and Alexander S. Graphodatsky. 2018. "The Case of X and Y Localization of Nucleolus Organizer Regions (NORs) in Tragulus javanicus (Cetartiodactyla, Mammalia)" Genes 9, no. 6: 312. https://doi.org/10.3390/genes9060312