3.1. Comparison of Hydroclimate Variables for 2003 and 2004
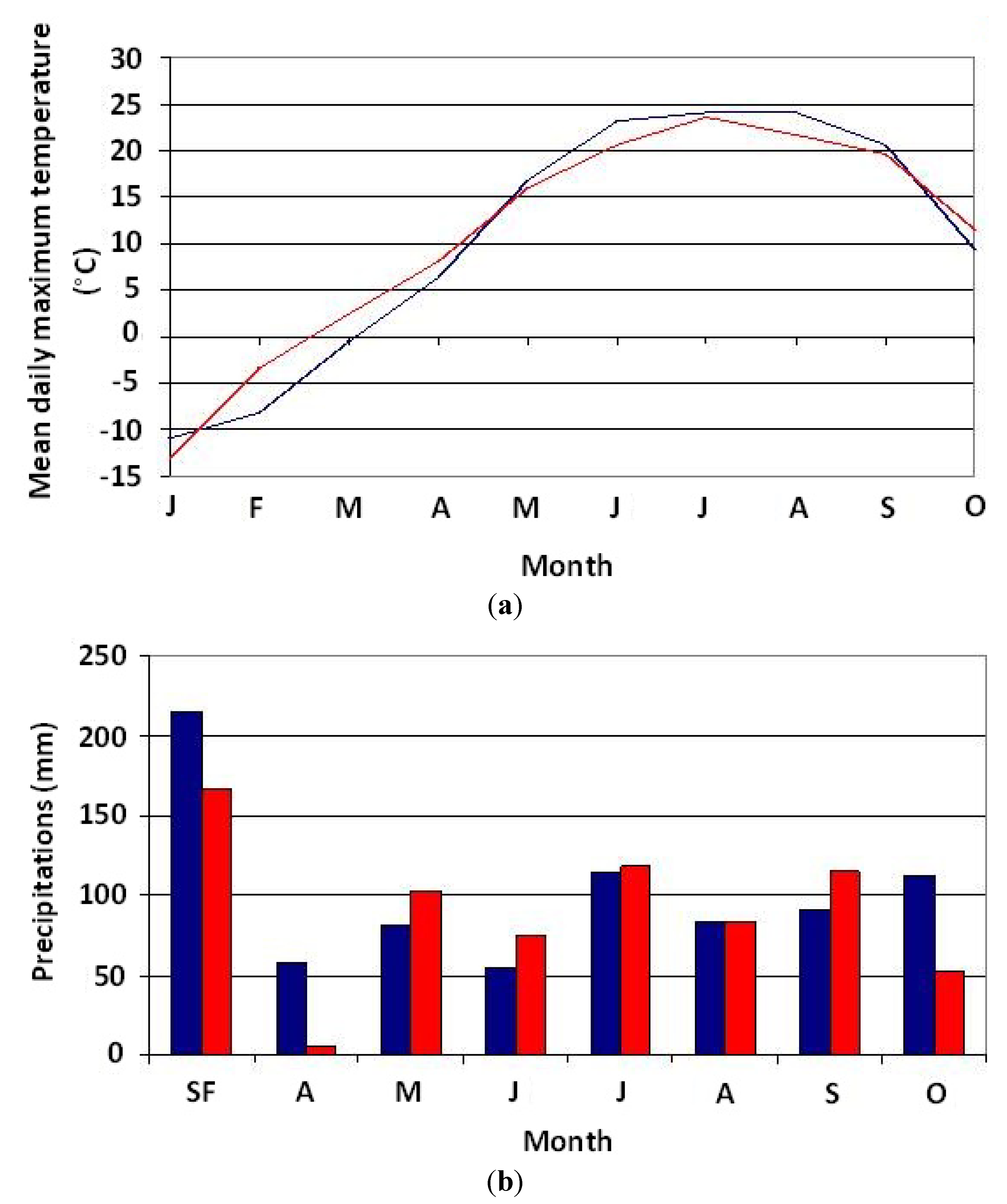
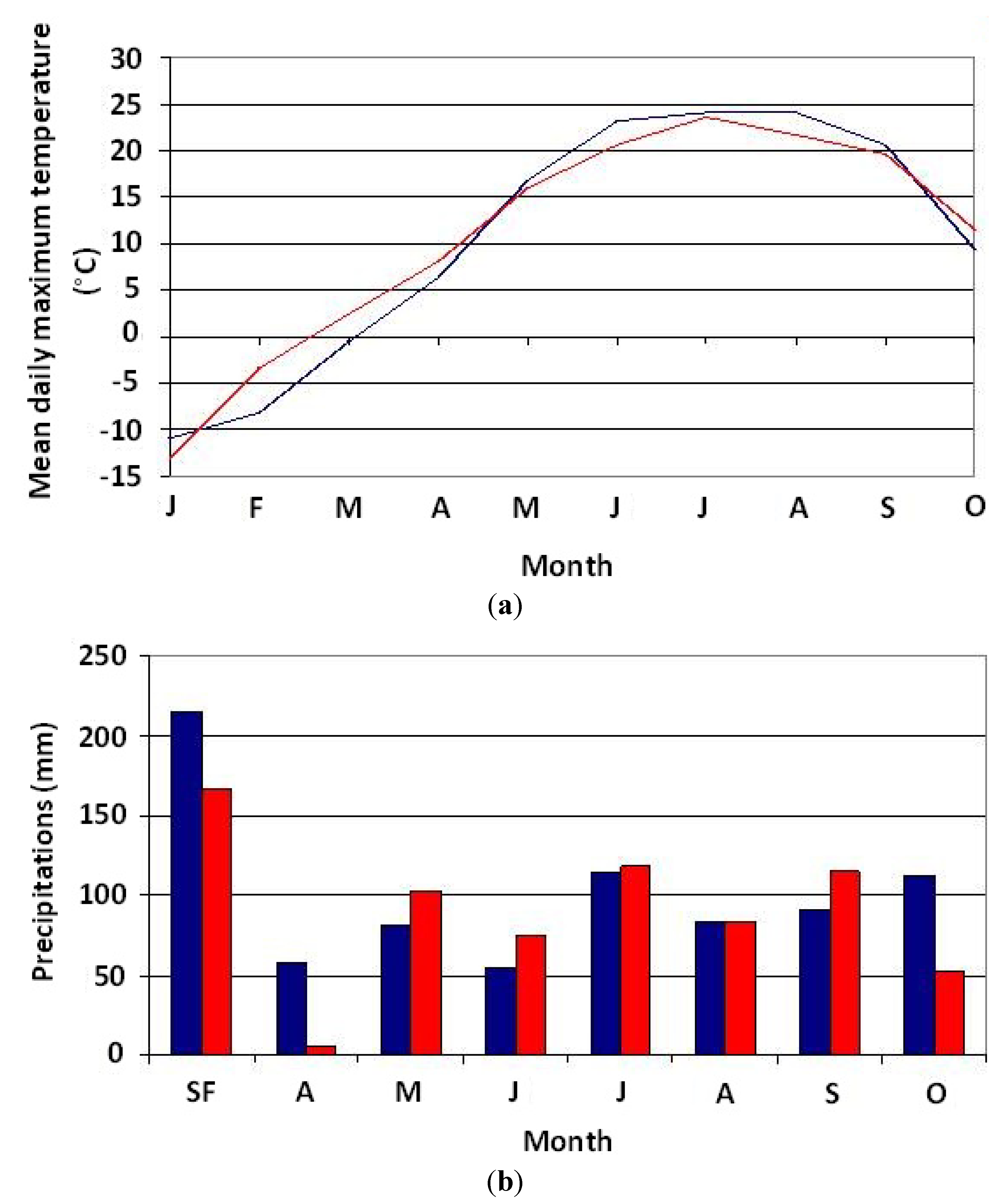
Temperature and precipitation variability in 2003 and 2004 is shown in
Figure 3a,b. Results of the comparison of measurements of these two variables at the monthly scale for the two years using the paired
t-test are shown in
Table 1. These results reveal that there is no statistically significant difference between the two years. Thus, climate conditions for 2003 and 2004 are similar. As for streamflow (
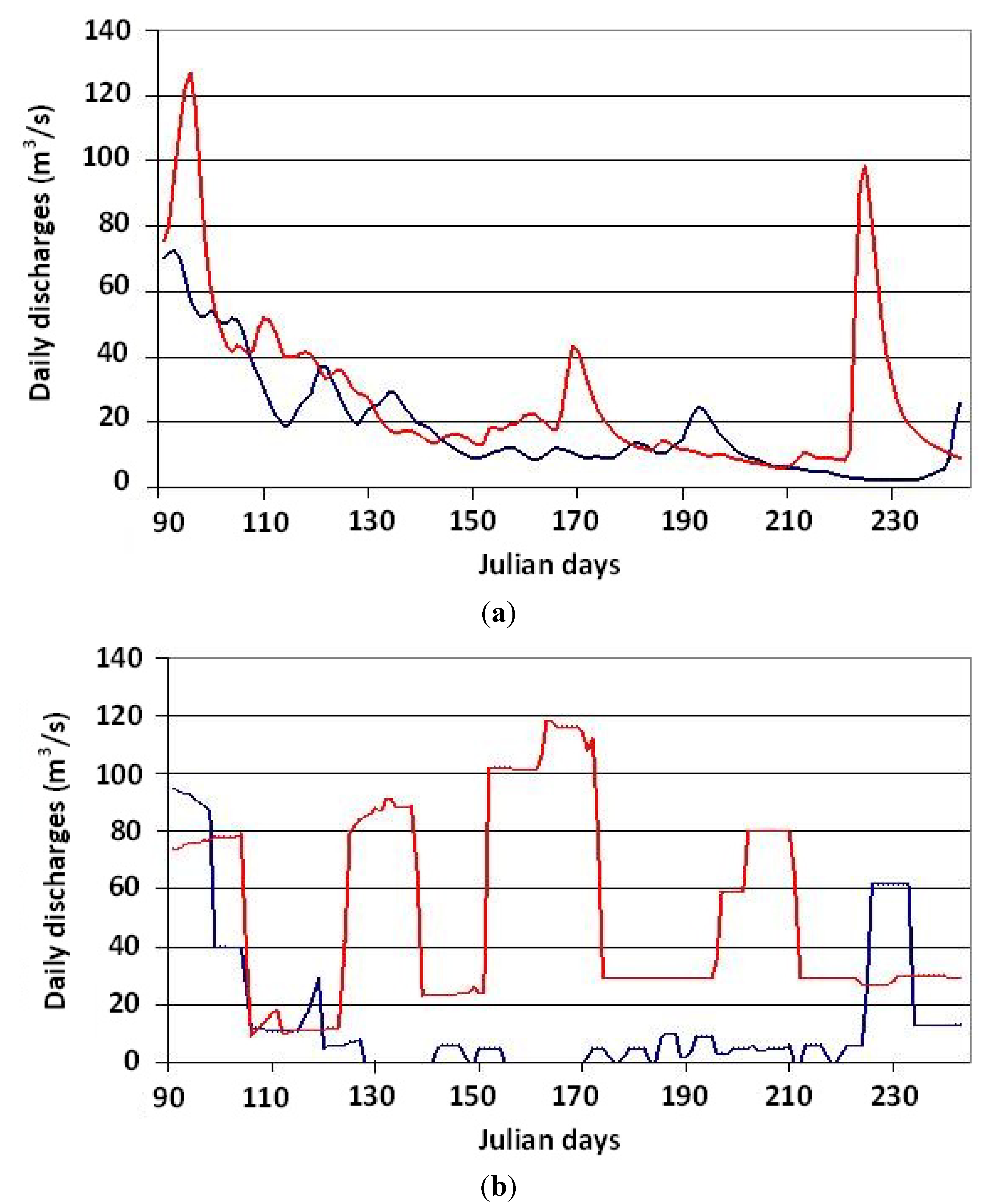
Figure 4), no significant difference was observed upstream from the dam between 2003 and 2004 at the monthly scale. At the daily scale, however, a significant difference is observed that is due to the fact that three floods occurred in 2004, while there were none in 2003 (
Figure 4). In contrast, downstream from the dam, the difference in streamflow between the two years is large (
Table 1). Over the growing season as a whole (except for a few days in August), flows are systematically lower in 2003 than in 2004. Indeed, mean daily and monthly flows during the growing season were roughly four times larger in 2004 than in 2003. The most important observation is the large number of days with very low daily flows (<1 L/s/km
2) in 2003, including a total of 54 days during the growing season (
Table 2). The longest sequence of no-flow days exceeds 15 days in June. As a result, the interannual variability of streamflow was higher downstream than upstream from the dam. This high variability, which is not observed in climate data and streamflow measurements upstream from the dam, is due to the mode of management of the dam, which consists in storing large amounts of water in the reservoir in the spring and summer for use in winter for hydroelectric power generation. There does not, however, seem to be any link between climate (temperature and precipitation) and the amount of water stored in the reservoir, as described below.
Table 1.
Comparison of mean values of hydroclimatic variables measured in 2003 and 2004 using the paired t-test.
Table 1.
Comparison of mean values of hydroclimatic variables measured in 2003 and 2004 using the paired t-test.
| Variables | Variables | Mean for 2003 | Mean for 2004 | p-value |
|---|
| Temperature (°C) | Maximums | 10.5 (13.42) | 10.7 (12.18) | 0.800 |
| Minimums | −2.3 (13.72) | −2.0 (12.70) | 0.690 |
| Precipitations (mm) | 101.3 (50.80) | 90 (48.26) | 0.406 |
| Monthly mean flows (m3/s) & | Upstream from dam | 24.5 (14.28) | 35.4 (24.79) | 0.132 |
| Downstream from dam | 14.5 (16.66) | 51.8 (21.08) | 0.068 * |
| Daily mean flows (m3/s) & | Upstream from dam | 21.3 (19.91) | 33.9 (30.64) | 0.000 |
| Downstream from dam | 14.4 (23.52) | 51.6 (33.30) | 0.000 |
Figure 3.
(a) Comparison of mean monthly temperatures measured at the Saint-Michel-des-Saints station in 2003 (blue curve) and 2004 (red curve); (b) Comparison of total monthly precipitation measured at the Saint-Michel-des-Saints station in 2003 (blue curve) and 2004 (red curve). SF = total snowfall in fall and winter.
Figure 3.
(a) Comparison of mean monthly temperatures measured at the Saint-Michel-des-Saints station in 2003 (blue curve) and 2004 (red curve); (b) Comparison of total monthly precipitation measured at the Saint-Michel-des-Saints station in 2003 (blue curve) and 2004 (red curve). SF = total snowfall in fall and winter.
Table 2.
Comparison of the total number of days for which daily specific flows during the growing season were nil or nearly so (<1 L/s/km2) downstream from the dam in 2003 and 2004.
Table 2.
Comparison of the total number of days for which daily specific flows during the growing season were nil or nearly so (<1 L/s/km2) downstream from the dam in 2003 and 2004.
| Year | April | May | June | July | August | Total |
|---|
| 2003 | 0 | 17 | 21 | 12 | 4 | 54 |
| 2004 | 0 | 0 | 0 | 0 | 0 | 0 |
Figure 4.
Comparison of daily flows in 2003 (blue curve) and 2004 (red curve) during the growing season, upstream (a); and downstream (b) from the Matawin dam.
Figure 4.
Comparison of daily flows in 2003 (blue curve) and 2004 (red curve) during the growing season, upstream (a); and downstream (b) from the Matawin dam.
3.2. Comparison of Abiotic and Biotic Variables in 2003 and 2004
Results of grain-size distribution analysis for sediments from the six sand bars are presented in
Table 3. No significant difference in grain-size distribution is observed between 2003 and 2004. Bars upstream and downstream from the dam are mainly comprised of sand. Moreover, no significant difference is observed in the chemistry of sediments sampled in 2003 and 2004 (
Table 4). As for vegetation, a complete list of species collected on the six sites is presented in
Table A1,
Table A2. No significant change is observed from 2003 to 2004 on the three bars located upstream from the dam for ecological group species richness (
Table 5). In contrast, downstream from the dam, two significant changes are observed and confirmed using the Chi-square test at the 5% significance level. The first change is a decrease in the total number of species per site. Thus, the average number of species per site fell by 23%, from 30 in 2003 to 23 in 2004. This decrease in the total number of species is due to a decrease in the number of terrestrial species (second change), the mean frequency per site for these species going from 25.4% in 2003 to 13% in 2004. Incidentally, a comparison of species frequency for the three ecological groups upstream and downstream from the dam reveals that obligate wetland species are more abundant than species in the other two groups and that the difference in the number of obligate wetland species and the number of species in the other two groups is higher upstream than downstream from the dam. Conversely, the proportion of terrestrial species becomes very small. Sand bars located at the edge of the low-flow channel are frequently and regularly submerged by flood waters, so that they are effectively wetlands. This likely accounts for the high frequency of obligate wetland species compared to species in the other two ecological groups, in particular terrestrial species. Sand bar submergence under flood waters is less frequent and regular downstream from the dam as a result of spring flood suppression and the storage of large amounts of water in the reservoir. This leads to the “terrestrialization” of bars located at the edge of the low-flow channel, which promotes their invasion by terrestrial plants which do not germinate effectively in sites characterized by prolonged, frequent and regular submergence (e.g., [
38]). The loss of terrestrial species observed on the three bars downstream from the dam in 2004 is due to the fact that, unlike in 2003 when the lack of water releases meant that they were exposed for many days at a time, the bars remained submerged for long periods in 2004. It follows that man-made “artificial drought” affects terrestrial species more strongly than obligate wetland species in the low-flow channel downstream from the dam. In a study of species dynamics in the St. Lawrence River alluvial plain in Quebec, [
22,
23] observed that, in dry years (climate-induced drought), the number of aquatic species decreased while the number of exotic or aggressive terrestrial species increased.
Table 3.
Grain-size distribution (%) in 2003 and 2004 in sites located upstream (UD) and downstream (DD) from the dam.
Table 3.
Grain-size distribution (%) in 2003 and 2004 in sites located upstream (UD) and downstream (DD) from the dam.
| Grain-size | Upstream from dam | Downstream from dam |
|---|
| 2003 | 2004 | 2003 | 2004 |
|---|
| UD1 | UD2 | UD3 | UD1 | UD2 | UD3 | DD1 | DD2 | DD3 | DD1 | DD2 | DD3 |
|---|
| Sand | 98 | 98.6 | 97.6 | 97.9 | 98.4 | 97.6 | 98.0 | 98.6 | 98.0 | 97.9 | 98.4 | 98.1 |
| Silt | 1.8 | 1.3 | 1.9 | 1.9 | 1.6 | 1.9 | 1.8 | 1.3 | 1.8 | 1.7 | 1.6 | 1.5 |
| Clay | 0.2 | 0.1 | 0.5 | 0.1 | 0.0 | 0.5 | 0.2 | 0.1 | 0.2 | 0.6 | 0 | 0.4 |
Table 4.
Nutrients concentration in three bars upstream and downstream from the dam in 2003 and 2004. TOC = total organic carbon (%); TN = total nitrogen (%); TP = total phosphorus (ppm).
Table 4.
Nutrients concentration in three bars upstream and downstream from the dam in 2003 and 2004. TOC = total organic carbon (%); TN = total nitrogen (%); TP = total phosphorus (ppm).
| Nutrients | Upstream from dam | Downstream from dam |
|---|
| 2003 | 2004 | 2003 | 2004 |
|---|
| UD1 | UD2 | UD3 | UD1 | UD2 | UD3 | DD1 | DD2 | DD3 | DD1 | DD2 | DD3 |
|---|
| TOC | 1.23 | 0.12 | 0.06 | 1.46 | 0.12 | 0.04 | 0.49 | 0.74 | 0.57 | 0.44 | 0.62 | 0.55 |
| TN | 0.09 | 0.02 | 0.10 | 0.073 | 0.012 | 0.10 | 0.02 | 0.02 | 0.02 | 0.02 | 0.02 | 0.02 |
| TP | 64.9 | 9.9 | 11.9 | 63.4 | 9.6 | 11.9 | 29.3 | 36.3 | 36.7 | 30.3 | 36 | 38.7 |
Table 5.
Relative species abundance (%) by ecological group, upstream and downstream from the dam in 2003 and 2004. OBL = obligate wetland species; FAC = facultative wetland species; TS = terrestrial species.
Table 5.
Relative species abundance (%) by ecological group, upstream and downstream from the dam in 2003 and 2004. OBL = obligate wetland species; FAC = facultative wetland species; TS = terrestrial species.
| Ecological group | Upstream from dam | Downstream from dam |
|---|
| 2003 | 2004 | 2003 | 2004 |
|---|
| UD1 | UD2 | UD3 | UD1 | UD2 | UD3 | DD1 | DD2 | DD3 | DD1 | DD2 | DD3 |
|---|
| OBL (%) | 63 | 53.9 | 60.6 | 63 | 53.9 | 60.6 | 40.6 | 38.7 | 37 | 48 | 60 | 47.7 |
| FAC (%) | 25.9 | 33.3 | 27.3 | 25.9 | 33.3 | 27.3 | 34.4 | 32.3 | 40.8 | 40 | 24 | 33.3 |
| TS (%) | 11.1 | 12.8 | 9.1 | 11.1 | 12.8 | 9.1 | 25.0 | 29.0 | 22.2 | 12 | 16 | 11.1 |
| Total number | 27 | 39 | 33 | 27 | 39 | 33 | 32 | 31 | 27 | 25 | 25 | 18 |
In contrast, these latter species disappeared when the alluvial plain was flooded while the number of aquatic plants increased. Similarly, in Australia, increases in the magnitude of flooding downstream from a dam led to the senescence of flood-intolerant species in the Murray River alluvial plain [
38], an observation also confirmed by other authors (e.g., [
39]). Very low water table variability in the low-flow channel relative to the alluvial plain could account for the maintenance of obligate wetland species abundance under drought conditions.
To constrain better the effect of the interannual variability of daily streamflow on the number of species downstream from the dam, we calculated the coefficients of correlation between abiotic and biotic variables and applied principal component analysis to the data shown in
Table 5. Coefficients of correlation are shown in
Table 6, which indicates that there is no significant difference in coefficient of correlation values between 2003 and 2004 due to the significant lack of variability in abiotic variables between the two years. However, for the two years, total nitrogen (TN) is positively correlated with the proportion of wetland obligate species (OBL) and, to a lesser extent, negatively correlated with the proportion of facultative species (FAC). It should also be pointed out that TN did not vary downstream from the dam from 2003 to 2004 (see
Table 4), so that nitrogen cannot account for changes observed downstream from the dam in 2004.
Table 6.
Coefficients of correlation calculated between abiotic and biotic variables for 2003–2004.
Table 6.
Coefficients of correlation calculated between abiotic and biotic variables for 2003–2004.
| Abiotic variables | 2003 (n = 6) | 2004 (n = 6) | 2003 and 2004 (n = 12) |
|---|
| OBL | FAC | TS | OBL | FAC | TS | OBL | FAC | TS |
|---|
| Sand | −0.343 | 0.244 | 0.424 | −0.159 | −0.048 | 0.846 | −0.307 | 0.123 | 0.461 |
| Silt | 0.241 | −0.132 | −0.319 | 0.680 | −0.349 | −0.549 | 0.372 | −0.226 | −0.352 |
| Clay | 0.470 | −0.406 | −0.538 | −0.494 | 0.571 | −0.627 | 0.029 | 0.215 | −0.407 |
| TOC | 0.001 | −0.198 | 0.207 | 0.358 | −0.345 | 0.102 | 0.129 | −0.279 | 0.129 |
| TN | 0.847 | −0.819 | −0.763 | 0.668 | −0.493 | −0.683 | 0.691 ** | −0.630 * | −0.547 |
| TP | 0.035 | −0.181 | 0.139 | 0.223 | −0.280 | 0.097 | 0.095 | −0.230 | 0.097 |
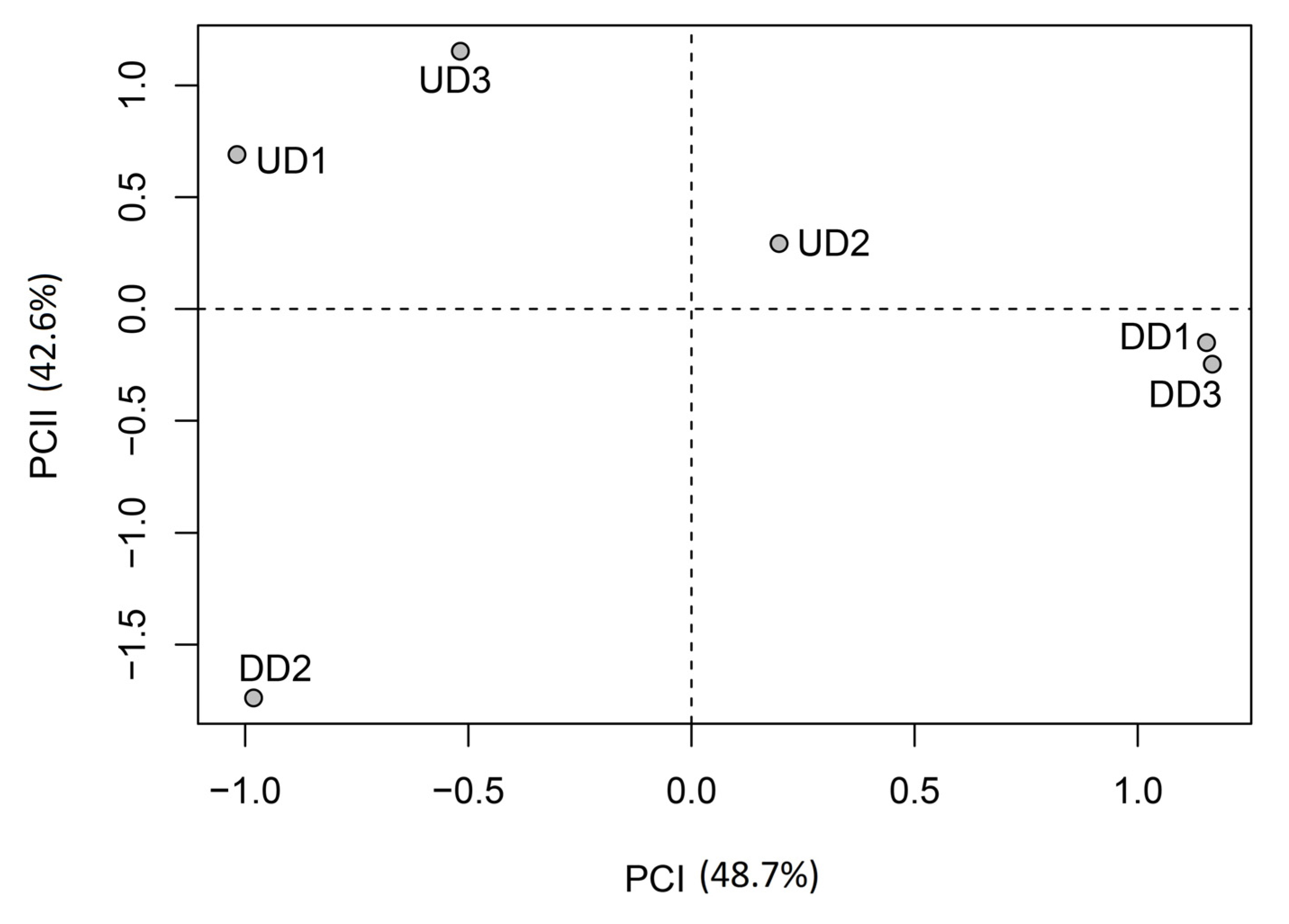
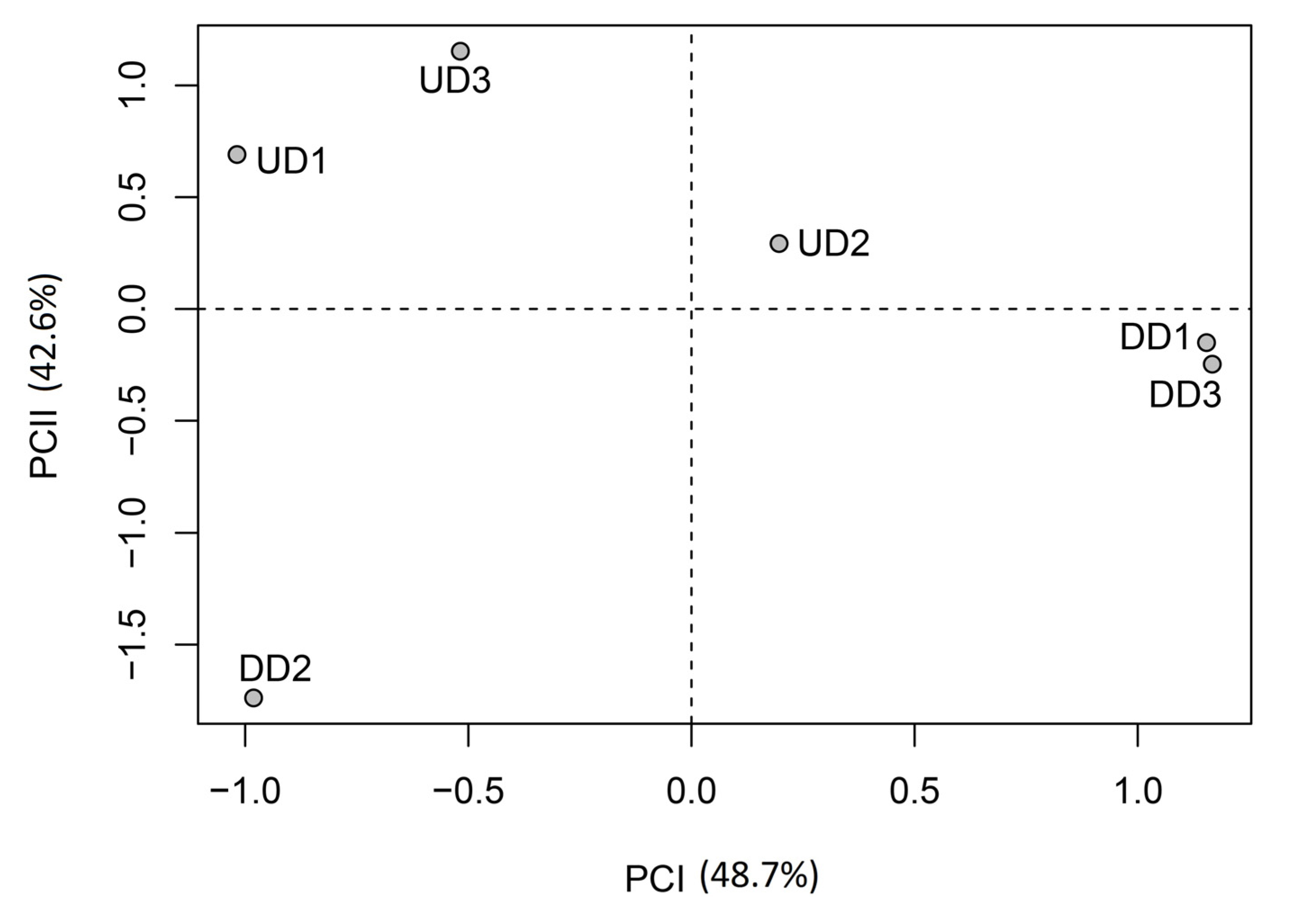
Results of principal component analysis are shown in
Table 7 and
Figure 5. Applying the Kayser criterion, only the first two components are found to be statistically significant, their eigenvalues being >1. Their cumulative total explained variance reaches 90%.
Table 7 shows that analysis of loading values does not distinguish between the two years based on the variations in the proportions of species in each of the three ecological groups. Indeed, the two principal components are strongly correlated (loading values >0.800) to the proportions of species in two of the three ecological groups in 2003 and 2004. Thus, for instance, the first principal component is strongly correlated with proportions of OBL and FACW group species in 2004, and with the proportion of FACW group species in 2003 (
Table 6). In contrast, principal component analysis highlighted variations between sites (
Figure 5). Thus, sites with high proportions (>60%) of obligate wetland species during at least one year have negative scores on the first principal component while the other sites have positive scores. None of the abiotic factors analyzed seems to account for this grouping, that is, the difference in the number of species in each ecological group observed between sites. It should be noted that this difference may be explained by many other abiotic factors which were not analyzed and cannot all be listed, for one thing, and/or by various ecological processes (
i.e., grazers, seed bank,
etc.), for another. The second principal component, for its part, distinguishes between sites according to their location with respect to the dam. The three sites located upstream from the dam have positive scores on this component while the sites located downstream from the dam have negative scores. Thus, this component reflects the variation in species proportions observed between 2003 and 2004. As previously mentioned, sites located upstream from the dam show no change in species composition between 2003 and 2004, while in sites located downstream, the number of terrestrial species decreased in 2004. This diminution is thought to result from the high variability of streamflow observed between 2003 and 2004 downstream from the dam which, as previously noted, is due to the occurrence of episodes of nil or quasi nil flow (<1 L/s/km²) during the growing season. Thus, despite a significant increase in daily flows in 2004, no change in the number of species is observed in the three sites located upstream from the dam. Furthermore, from a hydroclimatic standpoint, 2002 was similar to 2004. Consequently, the absence of some species observed in 2003 downstream from dam can only be the result of the drought observed that year. Based on these observations, we conclude that the first principal component reflects the effect of variations in species proportions, particularly those of obligate wetland species, according to location (spatial effect), while the second component reflects the effect of the interannual variability of streamflow downstream from the dam on species proportions (temporal effect).
Table 7.
Loading of species proportions in the three ecological groups on the first two significant principal components for 2003 and 2004. Results obtained after Varimax axis rotation.
Table 7.
Loading of species proportions in the three ecological groups on the first two significant principal components for 2003 and 2004. Results obtained after Varimax axis rotation.
Principal
component | 2003 | 2004 | EV (%) |
|---|
| OBL | FAC | TS | OBL | FAC | TS |
|---|
| PC I | −0.566 | 0.808 | 0.310 | −0.983 | 0.918 | −0.208 | 48.7 |
| PC II | 0.802 | −0.460 | −0.914 | 0.175 | 0.104 | −0.906 | 42.6 |
Figure 5.
Location of sites upstream (UD) and downstream (DD) from the dam in the space defined by the first two significant components.
Figure 5.
Location of sites upstream (UD) and downstream (DD) from the dam in the space defined by the first two significant components.
Many models have been proposed to account for the relationship between water levels and riparian and alluvial plain vegetation. Most of these models, however, were developed for lentic settings (e.g., [
3,
23,
40,
41,
42]). In Quebec, for instance, [
23] developed a model for the St. Lawrence River. For reasons already mentioned, the fact that this model was developed for a lentic fluvial setting with low interannual variability of water levels means that it is not applicable to the present case. Although a few models have been proposed for natural and regulated rivers (e.g., [
9,
23,
40,
41,
42]), their generalization remains limited due to the complexity of factors which affect species abundance at any given site. In addition, our study aimed at measuring the extent of the presence of terrestrial plants in the low-flow channel downstream from the dam as it relates to flow inversion and man-made drought. In this perspective, the study did not consider some aspects such as the role of seed bank and the analysis of the frequency of drawdown. The effect of this factor has already been demonstrated by many authors (e.g., [
3,
4,
8,
38]). It is reasonable to expect that this factor could in part account for the variation in the number of species observed downstream from the dam between 2003 and 2004 on the one hand, and for the variation in the number of species observed between sites, on the other hand. Finally, models and theories developed for rivers are more applicable to riparian vegetation than to low-flow channel vegetation in a given river.


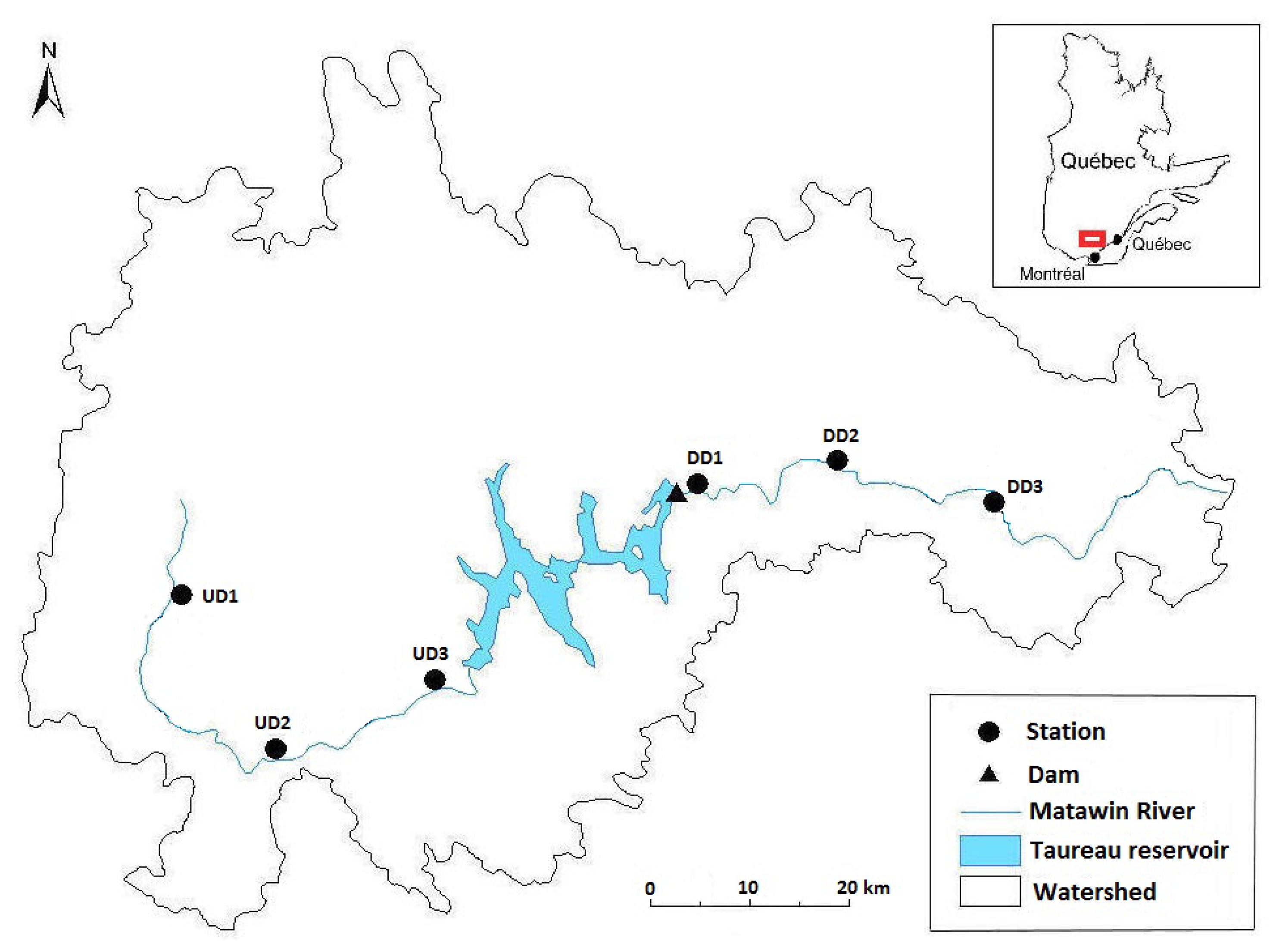

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}